
Periplasmic Protein Mobility for Extracellular Electron Transport in Shewanella oneidensis


Abstract
1. Introduction
2. Materials and Methods
2.1. Organisms and Microbial Growth
2.2. Crosslinking
2.3. Fractionation
2.4. Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis (SDS-PAGE) and Heme-Staining
2.5. Fumarate Reductase Assay
2.6. Bioelectrochemistry
3. Results
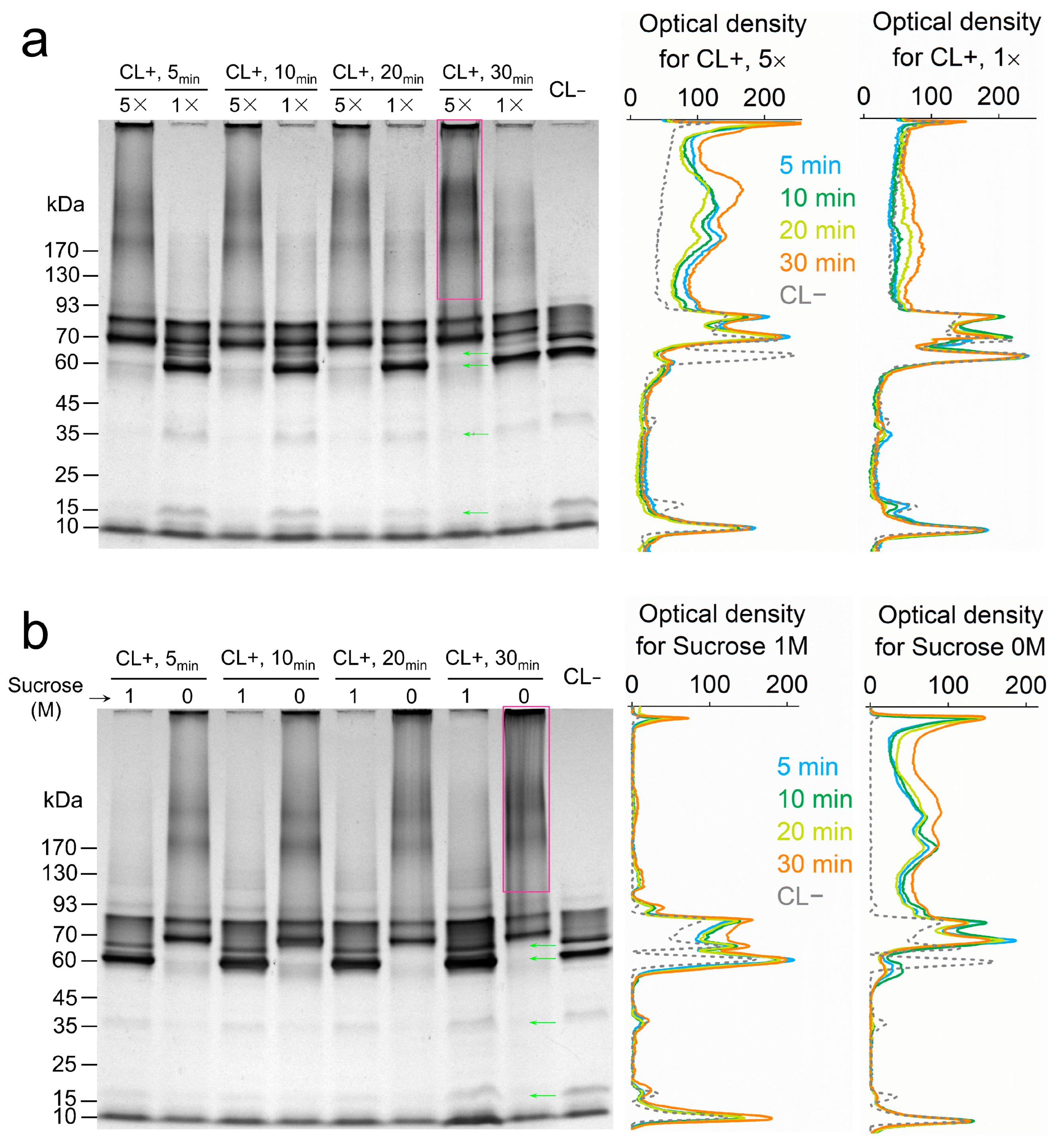
3.1. Dependence of Cytochrome Crosslinking on Protein Mobility
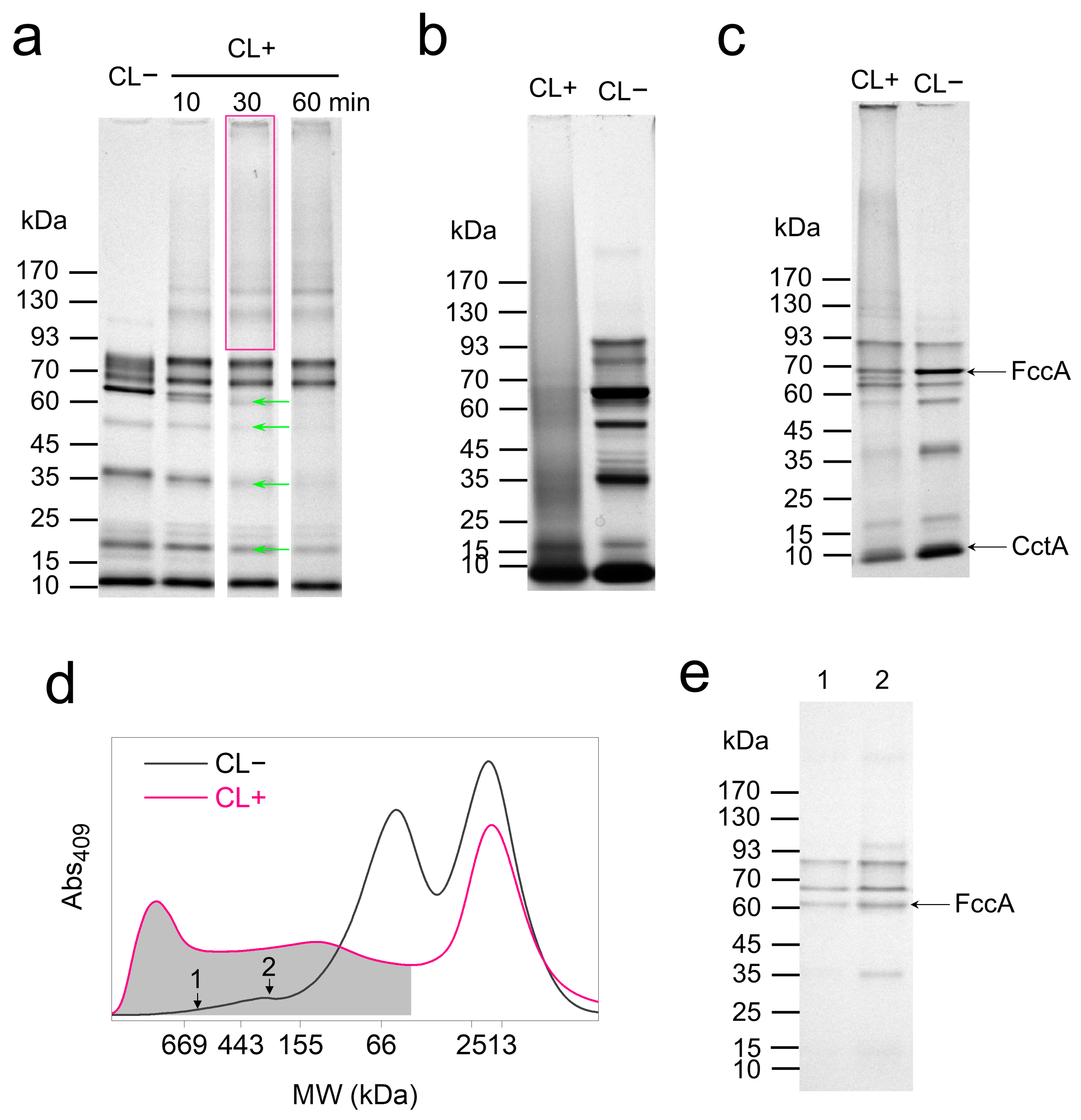
3.2. In Vivo Crosslinking of Periplasmic Cytochromes
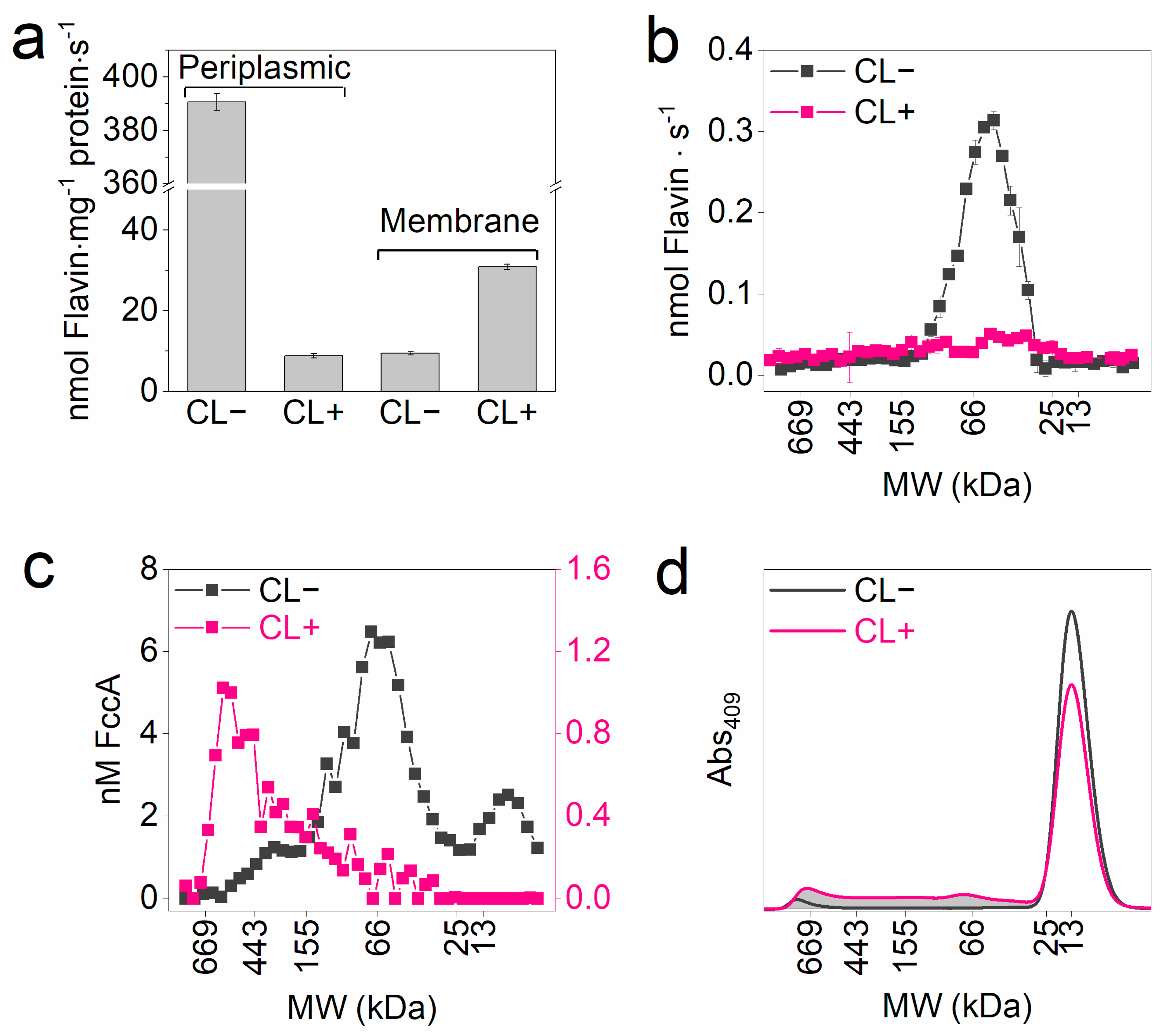
3.3. Differentiated Crosslinking of FccA and CctA
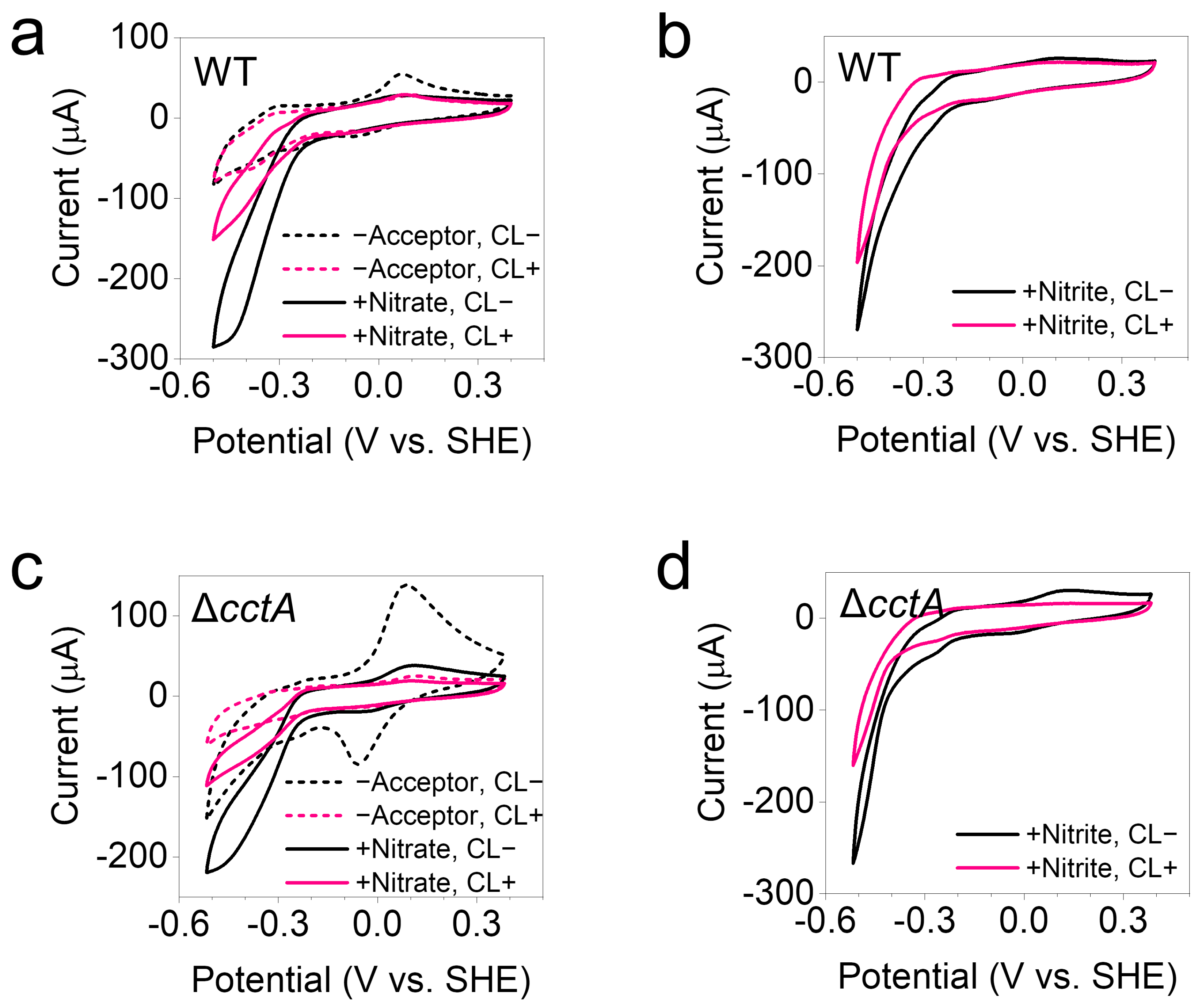
3.4. Preservation of EET Activity After Crosslinking
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CL+ | Crosslinked |
| CL− | Non-crosslinked |
| CM | Cytoplasmic membrane |
| EET | Extracellular electron transport |
| FAD | Flavin adenine dinucleotide |
| FMN | Flavin mononucleotide |
| MHCs | Multi-heme c-type cytochromes |
| MW | Molecular weight |
| OM | Outer membrane |
| SDS-PAGE | Sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| SEC | Size-exclusion chromatography |
| WT | Wild-type |
References
- Richter, K.; Schicklberger, M.; Gescher, J. Dissimilatory reduction of extracellular electron acceptors in anaerobic respiration. Appl. Environ. Microbiol. 2012, 78, 913–921. [Google Scholar] [CrossRef] [PubMed]
- Madigan, M.T.; Bender, K.S.; Buckley, D.H.; Sattley, W.M.; Stahl, D.A. Metabolic Diversity of Microorganisms. In Brock Biology of Microorganisms, 15th Global ed.; Pearson Education Limited: New York, NY, USA, 2019; pp. 428–486. [Google Scholar]
- Hau, H.H.; Gralnick, J.A. Ecology and biotechnology of the genus Shewanella. Annu. Rev. Microbiol. 2007, 61, 237–258. [Google Scholar] [CrossRef] [PubMed]
- Paquete, C.M.; Morgado, L.; Salgueiro, C.A.; Louro, R.O. Molecular mechanisms of microbial extracellular electron transfer: The importance of multiheme cytochromes. Front. Biosci. Landmark 2022, 27, 174. [Google Scholar] [CrossRef] [PubMed]
- Breuer, M.; Rosso, K.M.; Blumberger, J.; Butt, J.N. Multi-haem cytochromes in Shewanella oneidensis MR-1: Structures, functions and opportunities. J. Royal Soc. Interface 2015, 12, 20141117. [Google Scholar] [CrossRef]
- Edwards, M.J.; White, G.F.; Butt, J.N.; Richardson, D.J.; Clarke, T.A. The crystal structure of a biological insulated transmembrane molecular wire. Cell 2020, 181, 665–673. [Google Scholar] [CrossRef]
- Ross, D.E.; Ruebush, S.S.; Brantley, S.L.; Hartshorne, R.S.; Clarke, T.A.; Richardson, D.J.; Tien, M. Characterization of protein-protein interactions involved in iron reduction by Shewanella oneidensis MR-1. Appl. Environ. Microbiol. 2007, 73, 5797–5808. [Google Scholar] [CrossRef]
- Pandeya, A.; Ojo, I.; Alegun, O.; Wei, Y. Periplasmic targets for the development of effective antimicrobials against gram-negative bacteria. ACS Infect. Dis. 2020, 6, 2337–2354. [Google Scholar] [CrossRef]
- Pedebos, C.; Smith, I.P.S.; Boags, A.; Khalid, S. The hitchhiker’s guide to the periplasm: Unexpected molecular interactions of polymyxin B1 in E. coli. Structure 2021, 29, 444–456. [Google Scholar] [CrossRef]
- Alves, M.N.; Neto, S.E.; Alves, A.S.; Fonseca, B.M.; Carrelo, A.; Pacheco, I.; Paquete, C.M.; Soares, C.M.; Louro, R.O. Characterization of the periplasmic redox network that sustains the versatile anaerobic metabolism of Shewanella oneidensis MR-1. Front. Microbiol. 2015, 6, 665. [Google Scholar] [CrossRef]
- Tran, B.M.; Punter, C.M.; Linnik, D.; Iyer, A.; Poolman, B. Single-protein diffusion in the periplasm of Escherichia coli. J. Mol. Biol. 2024, 436, 168420. [Google Scholar] [CrossRef]
- Sochacki, K.A.; Shkel, I.A.; Record, M.T.; Weisshaar, J.C. Protein diffusion in the periplasm of E. coli under osmotic stress. Biophys. J. 2011, 100, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, B.M.; Paquete, C.M.; Neto, S.E.; Pacheco, I.; Soares, C.M.; Louro, R.O. Mind the gap: Cytochrome interactions reveal electron pathways across the periplasm of Shewanella oneidensis MR-1. Biochem. J. 2013, 449, 101–108. [Google Scholar] [CrossRef]
- Bretschger, O.; Obraztsova, A.; Sturm, C.A.; Chang, I.S.; Gorby, Y.A.; Reed, S.B.; Culley, D.E.; Reardon, C.L.; Barua, S.; Romine, M.F.; et al. Current production and metal oxide reduction by Shewanella oneidensis MR-1 wild type and mutants. Appl. Environ. Microbiol. 2007, 73, 7003–7012. [Google Scholar] [CrossRef]
- Liu, H.; Newton, G.J.; Nakamura, R.; Hashimoto, K.; Nakanishi, S. Electrochemical characterization of a single electricity-producing bacterial cell of Shewanella by using optical tweezers. Angew. Chem. Int. Ed. 2010, 49, 6596–6599. [Google Scholar] [CrossRef]
- Edwards, M.J.; Richardson, D.J.; Paquete, C.M.; Clarke, T.A. Role of multiheme cytochromes involved in extracellular anaerobic respiration in bacteria. Protein Sci. 2020, 29, 830–842. [Google Scholar] [CrossRef]
- Tayri-Wilk, T.; Slavin, M.; Zamel, J.; Blass, A.; Cohen, S.; Motzik, A.; Sun, X.; Shalev, D.E.; Ram, O.; Kalisman, N. Mass spectrometry reveals the chemistry of formaldehyde cross-linking in structured proteins. Nat. Commun. 2020, 11, 3128. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, B.W.; Toews, J.; Kast, J. Utility of formaldehyde cross-linking and mass spectrometry in the study of protein-protein interactions. J. Mass Spectrom. 2008, 43, 699–715. [Google Scholar] [CrossRef]
- Page, C.C.; Moser, C.C.; Dutton, P.L. Mechanism for electron transfer within and between proteins. Curr. Opin. Chem. Biol. 2003, 7, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Page, C.C.; Moser, C.C.; Chen, X.X.; Dutton, P.L. Natural engineering principles of electron tunnelling in biological oxidation-reduction. Nature 1999, 402, 47–52. [Google Scholar] [CrossRef]
- Asadi, M. Tables. In Beet-Sugar Handbook; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006; pp. 779–801. [Google Scholar]
- Schavemaker, P.E.; Boersma, A.J.; Poolman, B. How important is protein diffusion in prokaryotes? Front. Mol. Biosci. 2018, 5, 93. [Google Scholar] [CrossRef]
- Ross, D.E.; Flynn, J.M.; Baron, D.B.; Gralnick, J.A.; Bond, D.R. Towards electrosynthesis in Shewanella: Energetics of reversing the Mtr pathway for reductive metabolism. PLoS ONE 2011, 6, e16649. [Google Scholar] [CrossRef] [PubMed]
- Dohnalkova, A.C.; Marshall, M.J.; Arey, B.W.; Williams, K.H.; Buck, E.C.; Fredrickson, J.K. Imaging hydrated microbial extracellular polymers: Comparative analysis by electron microscopy. Appl. Environ. Microbiol. 2011, 77, 1254–1262. [Google Scholar] [CrossRef] [PubMed]
- Oswald, F.; Bank, E.L.M.; Bollen, Y.J.M.; Peterman, E.J.G. Imaging and quantification of trans-membrane protein diffusion in living bacteria. Phys. Chem. Chem. Phys. 2014, 16, 12625–12634. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Chong, K.; Massefski, W.; Evans, R. Quantitative interpretation of protein diffusion coefficients in mixed protiated-deuteriated aqueous solvents. J. Phys. Chem. B 2022, 126, 5887–5895. [Google Scholar] [CrossRef]
- Sturm, G.; Richter, K.; Doetsch, A.; Heide, H.; Louro, R.O.; Gescher, J. A dynamic periplasmic electron transfer network enables respiratory flexibility beyond a thermodynamic regulatory regime. ISME J. 2015, 9, 1802–1811. [Google Scholar] [CrossRef]
- Beblawy, S.; Bursac, T.; Paquete, C.; Louro, R.; Clarke, T.A.; Gescher, J. Extracellular reduction of solid electron acceptors by Shewanella oneidensis. Mol. Microbiol. 2018, 109, 571–583. [Google Scholar] [CrossRef]
- Lemaire, O.N.; Mejean, V.; Iobbi-Nivol, C. The Shewanella genus: Ubiquitous organisms sustaining and preserving aquatic ecosystems. FEMS Microbiol. Rev. 2020, 44, 155–170. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Source | Band Position (kDa) | Predicted Protein | Band Change |
|---|---|---|---|---|
| 1 | Cross-linked | 170 | - | Emerged |
| 2 | Cross-linked | 159 | OmcA-MtrC | Emerged |
| 3 | Cross-linked | 143 | MtrB-MtrA | Emerged |
| 4 | Cross-linked | 115 | - | Emerged |
| 5 | Native | 83 | OmcA | None |
| 6 | Native | 78 | Unknown | Vanished |
| 7 | Native | 73 | MtrC | None |
| 8 | Native | 64 | FccA | Vanished |
| 9 | Cross-linked | 62 | - | Emerged and vanished |
| 10 | Native | 52 | NrfA | Vanished |
| 11 | Native | 36 | MtrA | Vanished |
| 12 and 13 | Native | 23 | SO_4048/CytcB | Weaker |
| 21 | ||||
| 14 | Native | 19 | CymA | Weaker |
| 15 | Native | 16 | NapB | Vanished |
| 16 | Native | 11 | CctA | Weaker |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Zheng, X.; Yang, Y.; Xu, M. Periplasmic Protein Mobility for Extracellular Electron Transport in Shewanella oneidensis. Microorganisms 2025, 13, 1144. https://doi.org/10.3390/microorganisms13051144
Li D, Zheng X, Yang Y, Xu M. Periplasmic Protein Mobility for Extracellular Electron Transport in Shewanella oneidensis. Microorganisms. 2025; 13(5):1144. https://doi.org/10.3390/microorganisms13051144
Chicago/Turabian StyleLi, Daobo, Xiaodan Zheng, Yonggang Yang, and Meiying Xu. 2025. "Periplasmic Protein Mobility for Extracellular Electron Transport in Shewanella oneidensis" Microorganisms 13, no. 5: 1144. https://doi.org/10.3390/microorganisms13051144
APA StyleLi, D., Zheng, X., Yang, Y., & Xu, M. (2025). Periplasmic Protein Mobility for Extracellular Electron Transport in Shewanella oneidensis. Microorganisms, 13(5), 1144. https://doi.org/10.3390/microorganisms13051144