
Identification of Nigrospora oryzae Causing Leaf Spot Disease in Tomato and Screening of Its Potential Antagonistic Bacteria

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Pathogen Isolation and Culture Conditions
2.2. Strains Used in This Study
2.3. Assays for Pathogenicity and Morphological Observations
2.4. Genomic DNA Extraction and PCR Amplification
2.5. Phylogenetic Analysis
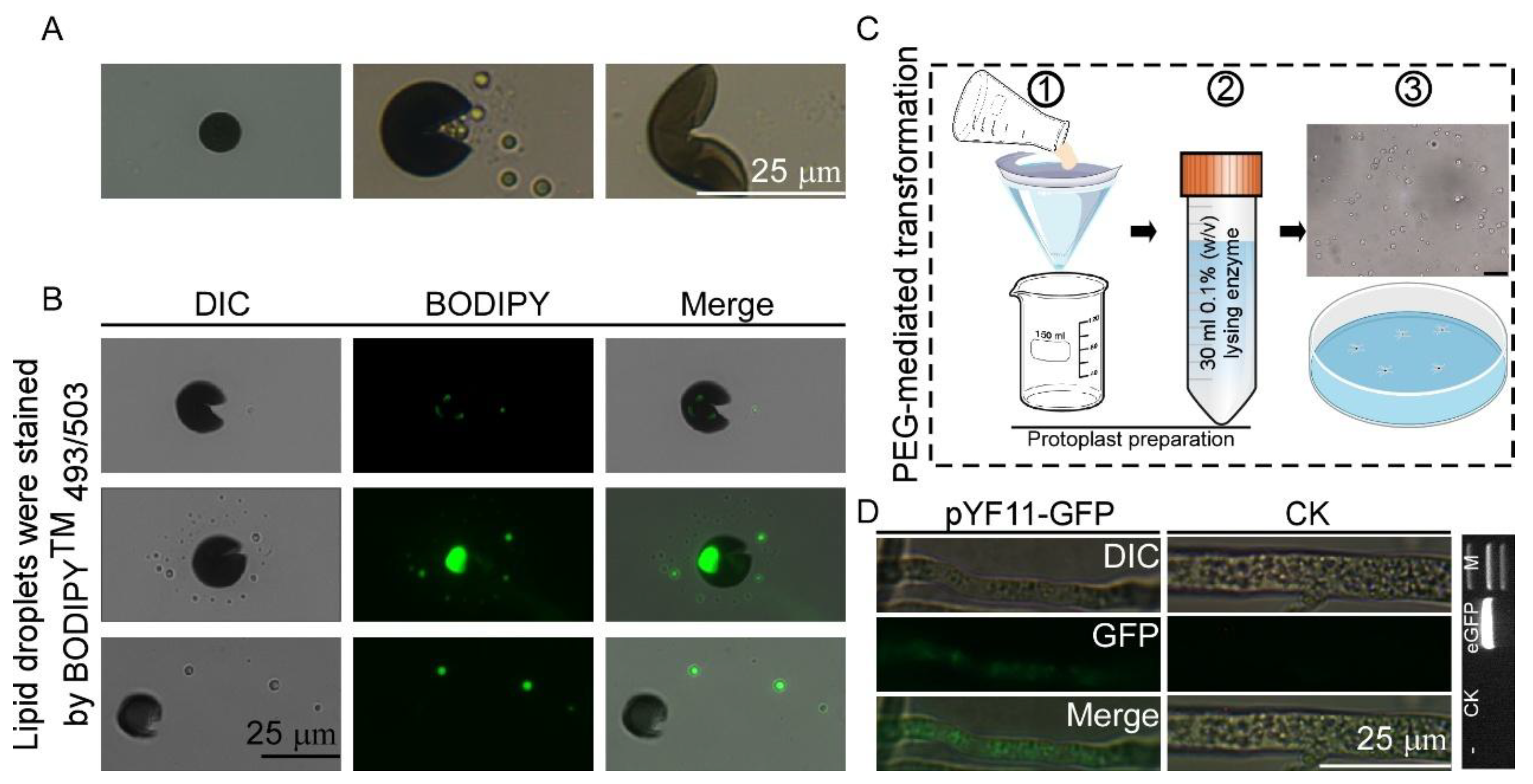
2.6. Polyethylene Glycol (PEG)-Mediated Transformation of N. oryzae
2.7. The Extracellular Laccase and Peroxidase Activity of N. oryzae
2.8. Epifluorescence Microscopy Observation
2.9. In Vitro Antagonism Against N. oryzae
2.10. Statistical Analysis
2.11. Accession Numbers
3. Results
3.1. Plant Disease Symptoms and Pathogenicity Test
3.2. Morphological Characteristics of Conidia
3.3. Establishment of Genetic Transformation Methods for N. oryzae
3.4. N. oryzae-YN Lost the Extracellular Laccase and Peroxidase Activity
3.5. Screening of Antagonistic Bacteria
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yin, R.; Wang, Z.; Xu, F. Multi-scenario simulation of China’s dynamic relationship between water-land resources allocation and cultivated land use based on shared socioeconomic pathways. J. Environ. Manag. 2023, 341, 118062. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Dong, X.; Wang, Z.; An, M.; Yang, X.; Xia, Z.; Wu, Y. First report of tomato brown rugose fruit virus infecting Solanum lycopersicum in Northeast China. Plant Dis. 2024, 108, 542. [Google Scholar] [CrossRef] [PubMed]
- Foolad, M.R.; Merk, H.L.; Ashrafi, H. Genetics, genomics and breeding of late blight and early blight resistance in tomato. Crit. Rev. Plant Sci. 2008, 27, 75–107. [Google Scholar] [CrossRef]
- Panthee, D.R.; Chen, F. Genomics of fungal disease resistance in tomato. Curr. Genom. 2010, 11, 30–39. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Wang, Y.; Yao, J.; Li, Z.; Huo, J.; Zhou, S.; Liu, W.; Wu, H. Genome sequence resource for Nigrospora oryzae, an important pathogenic fungus threatening crop production. Mol. Plant Microbe Interact. 2021, 34, 835–838. [Google Scholar] [CrossRef]
- Zeng, Y.; Jiang, Y.; Luo, M.; Yang, M.F. First report of leaf spot on Zanthoxylum bungeanum caused by Nigrospora sphaerica in China. Plant Dis. 2024, 108, 812. [Google Scholar] [CrossRef]
- Petrović, E.; Vrandečić, K.; Ćosić, J.; Đermić, E.; Godena, S. First report of Nigrospora species causing leaf spot on Olive (Olea europaea L.). Horticulturae 2023, 9, 1067. [Google Scholar] [CrossRef]
- Han, Y.Z.; Fan, Z.W.; Wu, C.F.; Li, M.Y.; Zhou, D.D. First report of Nigrospora leaf blight on elephant grass caused by Nigrospora sphaerica in China. Plant Dis. 2019, 103, 2681. [Google Scholar] [CrossRef]
- Kee, Y.J.; Hafifi, A.B.M.; Huda-Shakirah, A.R.; Wong, K.L.; Jin, X.L.; Siti Nordahliawate, M.S.; Zakaria, L.; Mohd, M.H. First report of reddish brown spot disease of red-fleshed dragon fruit (Hylocereus polyrhizus) caused by Nigrospora lacticolonia and Nigrospora sphaerica in Malaysia. Crop. Prot. 2019, 122, 165–170. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, K.; Zhao, Y.; Zhang, Y.; Fu, Q.; Shiwen, H. Nigrospora oryzae causing panicle branch rot disease on Oryza sativa (rice). Plant Dis. 2021, 105, 2724. [Google Scholar] [CrossRef]
- Tao, Y.; Yang, C.; Yu, S.; Fu, F.; Dai, T. New occurrence of Nigrospora oryzae causing leaf blight in Ginkgo biloba in China and biocontrol screening of endophytic bacteria. Microorganisms 2024, 12, 2125. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Dong, W.; Li, S.; Lu, X.; Wang, P.; Zhang, X.; Wang, Y.; Ma, P. Fengycin produced by Bacillus subtilis NCD-2 plays a major role in biocontrol of cotton seedling damping-off disease. Microbiol. Res. 2014, 169, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Chen, X.; Liu, X.; Guo, Q.; Li, S.; Lu, X.; Zhang, X.; Wang, P.; Dong, L.; Zhao, W.; et al. Genome mining and UHPLC-QTOF-MS/MS to identify the potential antimicrobial compounds and determine the specificity of biosynthetic gene clusters in Bacillus subtilis NCD-2. BMC Genom. 2020, 21, 767. [Google Scholar] [CrossRef]
- De la Cruz-Lopez, N.; Cruz-Lopez, L.; Holguin-Melendez, F.; Guillen-Navarro, G.K.; Huerta-Palacios, G. Volatile organic compounds produced by Cacao endophytic bacteria and their inhibitory activity on Moniliophthora roreri. Curr. Microbiol. 2022, 79, 35. [Google Scholar] [CrossRef]
- Liu, Z.; Friesen, T.L. Polyethylene glycol (PEG)-mediated transformation in filamentous fungal pathogens. Methods Mol. Biol. 2012, 835, 365–375. [Google Scholar]
- Liu, X.; Qian, B.; Gao, C.; Huang, S.; Cai, Y.; Zhang, H.; Zheng, X.; Wang, P.; Zhang, Z. The putative protein phosphatase MoYvh1 functions upstream of MoPdeH to regulate the development and pathogenicity in Magnaporthe oryzae. Mol. Plant Microbe Interact. 2016, 29, 496–507. [Google Scholar] [CrossRef]
- Terron, M.C.; Lopez-Fernandez, M.; Carbajo, J.M.; Junca, H.; Tellez, A.; Yague, S.; Arana-Cuenca, A.; Gonzalez, T.; Gonzalez, A.E. Tannic acid interferes with the commonly used laccase-detection assay based on ABTS as the substrate. Biochimie 2004, 86, 519–522. [Google Scholar] [CrossRef]
- Wang, J.; Qin, S.; Fan, R.; Peng, Q.; Hu, X.; Yang, L.; Liu, Z.; Baccelli, I.; Migheli, Q.; Berg, G.; et al. Plant growth promotion and biocontrol of leaf blight caused by Nigrospora sphaerica on passion fruit by Endophytic Bacillus subtilis strain GUCC4. J. Fungi 2023, 9, 132. [Google Scholar] [CrossRef]
- Zhang, J.; Li, H.; Gu, W.; Zhang, K.; Liu, X.; Liu, M.; Yang, L.; Li, G.; Zhang, Z.; Zhang, H. Peroxisome dynamics determines host-derived ROS accumulation and infectious growth of the rice blast fungus. mBio 2023, 14, e0238123. [Google Scholar] [CrossRef]
- Demoor, A.; Silar, P.; Brun, S. Appressorium: The breakthrough in Dikarya. J. Fungi 2019, 5, 72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tang, W.; Liu, K.; Huang, Q.; Zhang, X.; Yan, X.; Chen, Y.; Wang, J.; Qi, Z.; Wang, Z.; et al. Eight RGS and RGS-like proteins orchestrate growth, differentiation, and pathogenicity of Magnaporthe oryzae. PLoS Pathog. 2011, 7, e1002450. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Singh, A.K.; Kumar, A. Disease management of tomato through PGPB: Current trends and future perspective. 3 Biotech. 2017, 7, 255. [Google Scholar] [CrossRef] [PubMed]
- Karacic, V.; Miljakovic, D.; Marinkovic, J.; Ignjatov, M.; Milosevic, D.; Tamindzic, G.; Ivanovic, M. Bacillus Species: Excellent biocontrol agents against tomato diseases. Microorganisms 2024, 12, 457. [Google Scholar] [CrossRef]
- Wang, M.; Liu, F.; Crous, P.W.; Cai, L. Phylogenetic reassessment of Nigrospora: Ubiquitous endophytes, plant and human pathogens. Persoonia 2017, 39, 118–142. [Google Scholar] [CrossRef]
- Hao, Y.; Aluthmuhandiram, J.; Chethana, K.; Manawasinghe, I.S.; Li, X.; Liu, M.; Hyde, K.D.; Phillips, A.; Zhang, W. Nigrospora species associated with various hosts from Shandong Peninsula, China. Mycobiology 2020, 48, 169–183. [Google Scholar] [CrossRef]
- Qiu, C.; Zhu, W.; Niu, T.; Liu, Z. Nigrospora oryzae causing leaf spot on Asiatic Dayflower in Chongqing, China. Plant Dis. 2022, 106, 763. [Google Scholar] [CrossRef]
- Han, S.; Yu, S.; Zhu, T.; Li, S.; Qiao, T.; Liu, Y.; Lin, T.; Yang, C. Nigrospora oryzae causing black leaf spot disease of Hibiscus mutabilis in China. Plant Dis. 2021, 105, 2255. [Google Scholar] [CrossRef]
- Kim, S.M.; Kim, S.; Lee, J.; Choi, S.Y.; Chung, H.; Chun, J.; Seo, B.Y.; Lim, J.R.; Choi, N.J. First Report of Nigrospora oryzae causing leaf spot on Peanut (Arachis hypogaea L.) in the Republic of Korea. Plant Dis. 2024, 108, 3202. [Google Scholar] [CrossRef]
- Zhao, H.; Liu, H.Y.; Yang, X.S.; Liu, Y.X.; Ni, Y.X.; Wang, F.; Tang, L. First report of Nigrospora leaf blight on sesame caused by Nigrospora sphaerica in China. Plant Dis. 2014, 98, 842. [Google Scholar] [CrossRef]
- Liu, X.; Yang, J.; Qian, B.; Cai, Y.; Zou, X.; Zhang, H.; Zheng, X.; Wang, P.; Zhang, Z. MoYvh1 subverts rice defense through functions of ribosomal protein MoMrt4 in Magnaporthe oryzae. PLoS Pathog. 2018, 14, e1007016. [Google Scholar] [CrossRef] [PubMed]
- Thalavaipandian, A.; Ramesh, V.; Bagyalakshmi; Muthuramkumar, S.; Rajendran, A. Diversity of fungal endophytes in medicinal plants of Courtallam Hills, Western Ghats, India. Mycosphere 2011, 2, 575–582. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′-3′) | Remark |
|---|---|---|
| TUB2-F | GGTAACCAAATCGGTGCTGCTTTC | Amplification of TUB2 sequence |
| TUB2-R | ACCCTCAGTGTAGTGACCCTTGGC | Amplification of TUB2 sequence |
| 5EF1-728F | CATCGAGAAGTTCGAGAAGG | Amplification of TEF1-α sequence |
| EF1-986R | TACTTGAAGGAACCCTTACC | Amplification of TEF1-α sequence |
| ITS1 | CTTGGTCATTTAGAGGAAGTAA | Amplification of ITS1 sequence |
| ITS4 | TCCTCCGCTTATTGATATGC | Amplification of ITS1 sequence |
| Organism | Accession | Host |
|---|---|---|
| Apiosporaceae sp. | KU747955 | Lomariopsis mexicana |
| Buergenerula spartinae | MN644634 | Sporobolus alterniflorus |
| Chromocleista sp. | MN644566 | Sporobolus pumilus |
| Fungal endophyte | KY765249 | Taxodium distichum |
| Fungal sp. | PQ320869 | Tachigali versicolor |
| Kohlmeyeriopsis medullaris | MN644565 | Sporobolus pumilus |
| Nigrospora oryzae | - | Solanum lycopersicum |
| Nigrospora oryzae | GQ221861 | Zea mays |
| Nigrospora oryzae | GQ328855 | Zea mays |
| Nigrospora oryzae | KX958066 | sediment below ocean floor |
| Nigrospora sp. | MN153965 | Zea mays |
| Nigrospora sp. | MF942937 | Dicranum scoparium |
| Nigrospora sphaerica | KT462720 | surface sterilized cow pea seed |
| Sordariomycetes sp. | KP991424 | Quercus hypoleucoides |
| Sordariomycetes sp. | JQ759919 | Pinus elliottii |
| Trichoderma afroharzianum | MN644564 | Sporobolus pumilus |
| Gene | Organism | Accession | Identity | Host |
|---|---|---|---|---|
| ITS1 | N. oryzae 2684 | EU272488.1 | 99.43% | Espeletia sp. |
| TUB2 | N. oryzae GUCC:19-5152 | OP450803.1 | 89.60% | Broadbean |
| TEF1-a | N. oryzae LC6969 | KY019386.1 | 72.46% | Citrus reticulata |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Yang, F.; Zhang, A.; Guo, Q.; Sun, X.; Zhang, S.; Di, D. Identification of Nigrospora oryzae Causing Leaf Spot Disease in Tomato and Screening of Its Potential Antagonistic Bacteria. Microorganisms 2025, 13, 1128. https://doi.org/10.3390/microorganisms13051128
Zhang J, Yang F, Zhang A, Guo Q, Sun X, Zhang S, Di D. Identification of Nigrospora oryzae Causing Leaf Spot Disease in Tomato and Screening of Its Potential Antagonistic Bacteria. Microorganisms. 2025; 13(5):1128. https://doi.org/10.3390/microorganisms13051128
Chicago/Turabian StyleZhang, Jun, Fei Yang, Aihong Zhang, Qinggang Guo, Xiangrui Sun, Shangqing Zhang, and Dianping Di. 2025. "Identification of Nigrospora oryzae Causing Leaf Spot Disease in Tomato and Screening of Its Potential Antagonistic Bacteria" Microorganisms 13, no. 5: 1128. https://doi.org/10.3390/microorganisms13051128
APA StyleZhang, J., Yang, F., Zhang, A., Guo, Q., Sun, X., Zhang, S., & Di, D. (2025). Identification of Nigrospora oryzae Causing Leaf Spot Disease in Tomato and Screening of Its Potential Antagonistic Bacteria. Microorganisms, 13(5), 1128. https://doi.org/10.3390/microorganisms13051128