1. Introduction
Porcine epidemic diarrhea virus (PEDV), a porcine enteropathogenic coronavirus belonging to the
Alphacoronavirus genus in the
Coronaviridae family, is the causative agent of porcine epidemic diarrhea (PED), causing acute diarrhea, vomiting, and dehydration in pigs, and leading to a high mortality in newborn piglets [
1]. PEDV was initially isolated in Belgium in 1978 [
2]. Since 2010, new highly virulent variants of PEDV have led to the pandemic of PED in China and gradually spread to regions across Asia, Europe, America, and beyond, inflicting substantial economic losses on pig farms worldwide [
3,
4,
5]. In recent years, PED has continued to be highly prevalent and has been ranked among the most critical diseases in Chinese pig farms [
6]. Currently, the prevention of PED mainly relies on vaccines, with antiviral drugs as effective supplements.
Among numerous anti-coronavirus drugs from various sources, nucleoside analogues, such as GS-441524, remdesivir, and molnupiravir, have shown broad-spectrum antiviral activity [
7,
8,
9,
10]. For example, GS-441524, the nucleoside precursor of remdesivir, has been proven highly efficacious against feline infectious peritonitis coronavirus (FIPV) and is well tolerated in cats [
11]. GS-441524 also showed a similar efficacy to remdesivir in inhibiting the replication of SARS-CoV-2 [
12]. Compared with remdesivir, GS-441524 has a number of pharmacological advantages, including needing fewer steps to be synthesized in vitro and be activated in vivo, less toxicity, which allows for higher concentrations of aerosol and oral administration, and better oral bioavailability [
13,
14]. Therefore, GS-441524 exerts more potent clinical value in the treatment of infectious diseases caused by coronaviruses. Recently, the anti-PEDV activities of GS-441524 and remdesivir were also tested. Xie et al. showed that GS-441524 exhibited stronger anti-PEDV activity and lower cytotoxicity than remdesivir in Vero cells and showed a good safety profile in cells and mice [
15]. Although it is evident that GS-441524 and remdesivir exert their anti-PEDV capability by directly inhibiting the viral RNA-dependent RNA polymerase (RdRp) activity, there are still several issues worth clarifying. For example, the antiviral activity of GS-441524 against trypsin-independent PEDV is still unknown. Is there a significant difference in the antiviral activity of GS-441524 against trypsin-dependent and trypsin-independent PEDVs? The answer to this question requires further investigation into the impact of GS-441524 on PEDV proliferation. In this study, we utilized trypsin-dependent and trypsin-independent strains to further study the inhibitory activity of GS-441524 against different PEDV strains and its impact on the viral cycle. We found that GS-441524 significantly inhibited the replication of trypsin-dependent and trypsin-independent PEDV. Further experiments showed that GS-441524 affected replication stages of the viral life cycle, including viral genome amplification, viral protein expression, and virion production. These results indicate that GS-441524 can serve as a potential therapeutic alternative for PED control.
2. Materials and Methods
2.1. Cells, Virus, and Compound
Vero CCL-81 cells (African green monkey kidney cells, purchased from ATCC) were cultured in Dulbecco’s Modified Eagle Medium (DMEM) (Hyclone, Logan, UT, USA), supplemented with 10% fetal bovine serum (FBS) (Gibco BRL, Gaithersburg, MD, USA), 100 U/mL of penicillin, and 100 μg/mL of streptomycin, at 37 °C in a 5% CO
2-enriched atmosphere. The trypsin-independent attenuated PEDV DR13
att (JQ023162; isolated from a commercial vaccine of Green Cross, Yongin, Republic of Korea), rPEDV-∆ORF3-GFP (recombinant PEDV DR13
att with ORF3 gene replaced by GFP gene) [
16], and trypsin-dependent rPEDV-S (recombinant PEDV DR13
att with S gene replaced by S gene from a trypsin-dependent strain (KF650375) were propagated in Vero cells. Virus titers were determined using the Reed-Muench method [
17] and expressed as tissue culture infective dose 50% (TCID
50). GS-441524 (Yuanye, Shanghai, China) was dissolved in 100% dimethylsulfoxide (DMSO) to form a 10 mM stock solution and diluted with DMEM to the specified concentrations before use.
2.2. Cytotoxicity Determination
The cytotoxicity of GS441524 was determined in Vero cells using the MTT assay. The cells were seeded at a density of 2 × 104 cells/well into 96-well plates. When the cells reached 90% confluence within about 24 h, the supernatant was discarded, and the cells were washed three times with PBS. Subsequently, two-fold serial diluted GS-441524, starting from an initial concentration of 400 μM, was added to the cells and incubated for 3 d, with six replicates for each concentration. Cell control and vehicle (DMSO) control were set up. The cell viability was detected using the MTT assay. In brief, 20 μL MTT solution (5 mg/mL) was added to each well and incubated at 37 °C for 4 h. Then, the reaction solution was removed, and 150 μL DMSO was added to each well, followed by slow shaking of the plate at room temperature for 10 min. The optical density (OD) of each well was measured at a wavelength of 490 nm. The relative viability of the cells was calculated according to the formula: cell survival rate (%) = OD (sample)/OD (control) × 100%. The half cytotoxic concentration (CC50) of GS-441524 was calculated.
2.3. Determination of the Half Maximal Effective Concentration (EC50) of GS-441524
The half maximal effective concentration (EC50) of GS-441524 against trypsin-independent PEDV DR13att was determined with Vero cells cultured in 96-well plates. Briefly, Vero cells with 90% confluency were infected with DR13att at a multiplicity of infection (MOI) of 0.01 and cultured for 3 d in the presence of two-fold serial dilutions of GS-441524. Cell control (without virus and compound), virus control (without compound and vehicle), and vehicle (DMSO) control were set up. The cell viability was determined using the MTT assay. The relative inhibitory rate of GS-441524 on PEDV was calculated according to the following formula: inhibitory rate (%) = [OD (sample) − OD (virus control)]/[OD (cell control) − OD (virus control)] × 100%, and the EC50 of GS-441524 was calculated.
2.4. Antiviral Effect of GS-441524 Against Trypsin-Independent PEDV
To analyze the inhibitory effect of GS-441524 on PEDV, Vero cells inoculated with PEDV DR13att were treated with different concentrations of GS-441524. The expression of PEDV (M and N) proteins was detected using IFA or Western Blot, and the viral titers in the supernatant were determined. The detailed protocols were as follows.
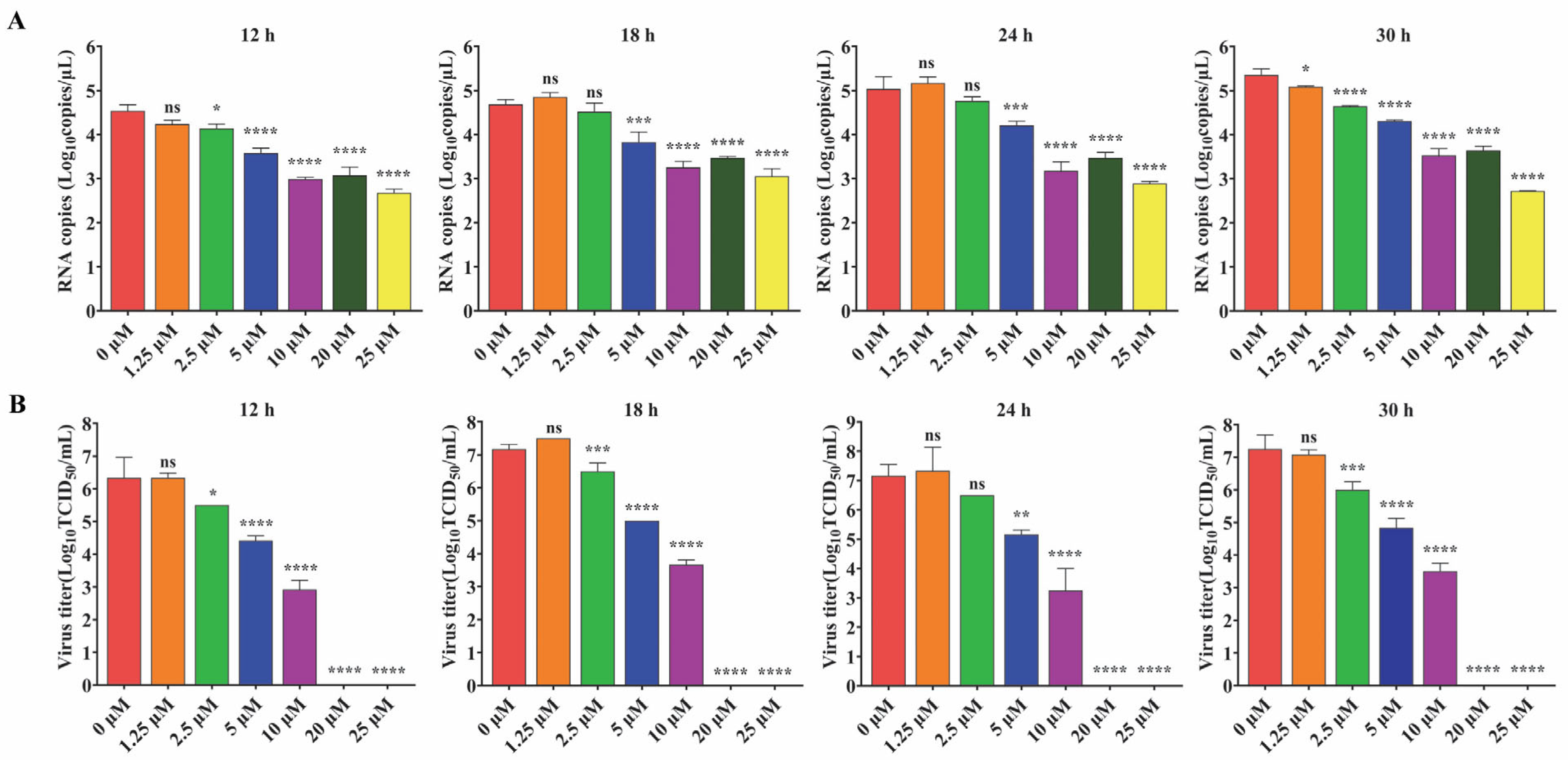
For IFA analysis, Vero cells were seeded into 48-well plates at 4 × 104 cells/well and cultured for about 24 h. Then, the cells were inoculated with PEDV DR13att at MOI 0.1. The inoculum was removed after 2 h incubation, and fresh maintenance medium containing 0, 1.25, 2.5, 5, 10, and 20 μM GS-441524 was added. The cells were fixed at 24 h post inoculation (hpi) and underwent immunofluorescent staining to analyze the expression of the M protein.
For Western Blot and virus titration analysis, Vero cells were seeded into 6-well plates at 4 × 105 cells/well. After about 24 h, the cells were inoculated with PEDV DR13att at an MOI of 0.1. The inoculum was removed after 2 h incubation, and fresh maintenance medium containing 0, 1.25, 2.5, 5, 10, and 20 μM GS-441524 was added. The viral titers (TCID50) and the mRNA levels of M protein in the supernatant were determined at 12, 18, 24, and 30 hpi. The cells collected at 30 hpi were lysed and analyzed for N protein expression by Western Blot.
2.5. Comparison of the Antiviral Activity of GS-441524 Against Trypsin-Dependent and -Independent PEDVs
Unlike cell-adapted strains that do not require trypsin, most clinical strains of PEDV require additional trypsin for proliferation. To further explore the anti-PEDV activity of GS-441524, Vero cells infected with DR13att (trypsin-independent) and rPEDV-S (trypsin-dependent, by substituting the S gene of DR13att with that of a trypsin-dependent strain) were treated with different concentrations of GS-441524. Cytopathic effects were observed, and the supernatant was collected at 24 and 48 hpi for viral titration (TCID50).
2.6. Effects of GS-441524 on the Life Cycle Phases of Trypsin-Independent PEDV
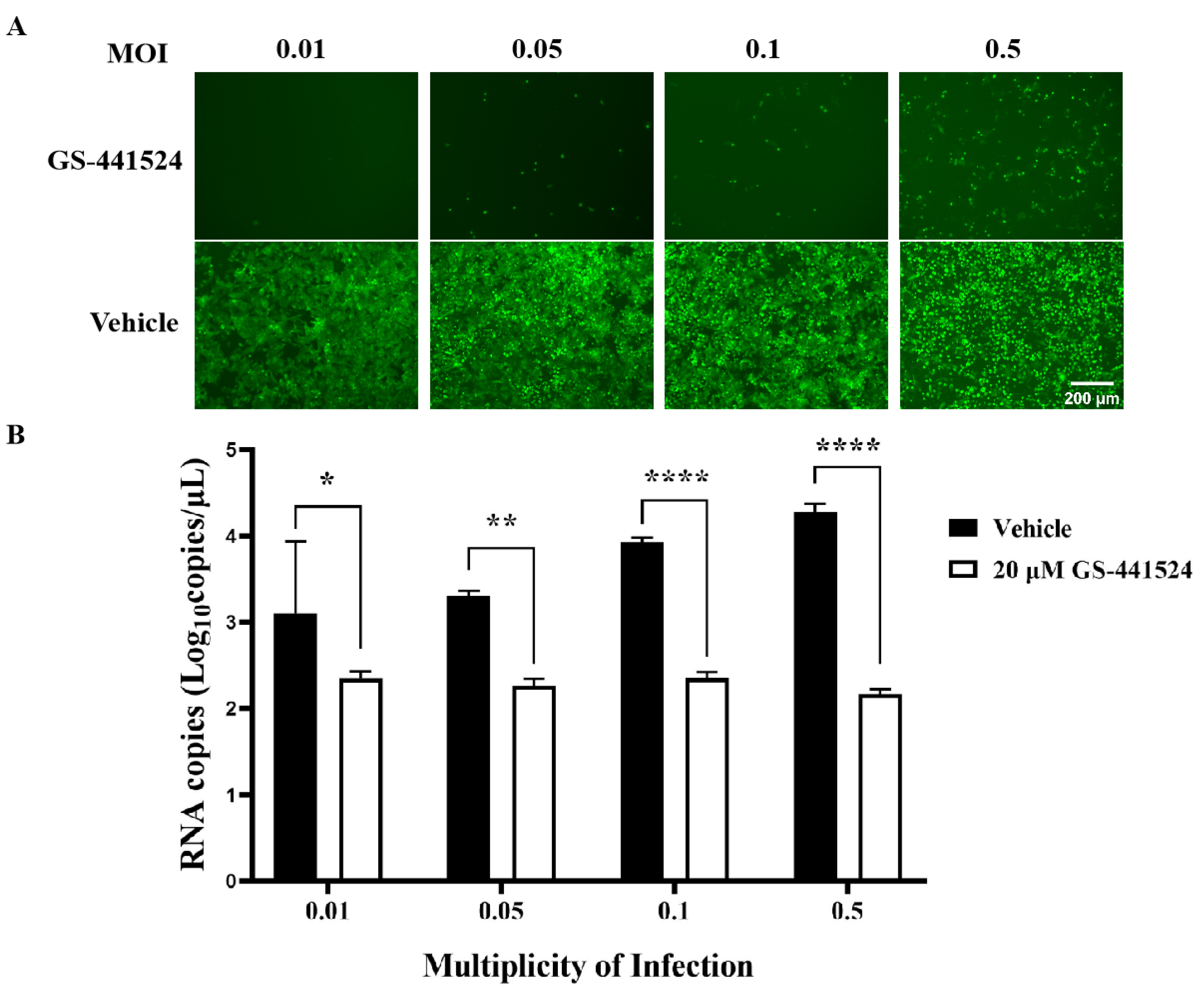
To assess the impact of GS-441524 on the life cycle of PEDV, Vero cells (1 × 105 cells/well) were seeded into 24-well plates and cultured until 90% confluence. Then, the Vero cells were treated with GS-441524 according to the following protocols. For attachment, the cells were incubated with rPEDV-∆ORF3-GFP (MOI 0.01) and 20 μM GS-441524 at 4 °C for 2 h. Then, the inoculum was removed, and the cells were cultured with fresh DMEM for another 48 h at 37 °C. For entry, the cells were infected with rPEDV-∆ORF3-GFP at MOI 0.01 at 4 °C for 2 h. Then, the inoculum was discarded, and the cells were treated with 20 μM GS-441524 at 37 °C for 2 h. After removing GS-441524, the cells were further cultured with fresh DMEM at 37 °C for 46 h. For replication, the cells were incubated with rPEDV-∆ORF3-GFP at MOI 0.01 at 4 °C for 2 h. After discarding the inoculum, the cells were cultured in fresh DMEM for another 6 h at 37 °C. Then, GS-441524 was added to the culture up to 20 μM and incubated for another 42 h. For release, the cells were infected with rPEDV-∆ORF3-GFP at MOI 0.01 at 4 °C for 2 h, and then cultured at 37 °C for 44 h. Then, the supernatant was discarded, and the cells were cultured in fresh DMEM with 20 μM GS-441524 for another 4 h. To determine the capability of the drug to inactivate PEDV, rPEDV-∆ORF3-GFP was preincubated with GS-441524 at 37 °C for 1 h. Following a 2 h incubation at 4 °C with the cells, the inocula were removed, and the cells were further cultured in fresh DMEM at 37 °C for 48 h. Lastly, the viral titers and the mRNA levels of M protein in the supernatant were determined for samples collected after each treatment. Images of cells were captured using an EVOS fluorescence microscope (M7000, Thermo Fisher Scientific, Waltham, MA, USA).
2.7. Immunofluorescence Assay (IFA)
For immunofluorescence staining, PEDV-infected cells were washed twice with PBS, fixed with 4% paraformaldehyde for 15 min, and then permeabilized with 0.1% Triton X-100 in PBS at room temperature for 15 min. After being blocked with 5% bovine serum albumin (BSA), the cells were incubated with anti-PEDV M polyclonal antibody (1:100) for 1 h, then rinsed three times with PBS and incubated with Alexa Fluor 488-conjugated goat anti-rabbit antibody (1:200, Beyotime, Shanghai, China) for 1 h. The nuclei were visualized by DAPI nuclear staining. Images of the immunofluorescent cells were captured using an EVOS fluorescence microscope at a magnification of 200×.
2.8. Western Blot
The cells were lysed in RIPA lysis buffer supplemented with a protease inhibitor cocktail (TransGen Biotech, Beijing, China) and subjected to sodium dodecyl sulphate–polyacrylamide gel electrophoresis (SDS–PAGE). The proteins separated by SDS–PAGE in the gel were then transferred onto a polyvinylidene fluoride (PVDF) membrane. Next, the membrane was blocked with 5% non-fat milk in Tris-buffered saline-Tween (TBST) and incubated with either anti-PEDV N monoclonal antibody (1:3000, Shanghai Ango Biotechnology Ltd., Shanghai, China) or anti-GAPDH monoclonal antibody (1:2000, Sangon Biotech, Shanghai, China) at room temperature for 1 h. Finally, the membrane was incubated with horseradish peroxidase (HRP) conjugated goat anti-rabbit IgG (1:20,000, Sangon Biotech, Shanghai, China). The proteins were detected using the Amersham ECL Western Blotting Analysis System (GE Healthcare, Chicago, IL, USA).
2.9. RNA Extraction and Real-Time RT-PCR
Total RNA was extracted from cells using the TIANamp viral RNA kit (TIANGEN, Beijing, China). Subsequently, the extracted RNA was subjected to reverse transcription with reverse transcription reagent (Promega, Madison, WI, USA). The standards for SYBR Green real-time RT-PCR were produced by amplifying a 486 bp membrane (M) gene fragment and then cloning it into the pJET1.2/blunt vector (Thermo Fisher Scientific, Waltham, MA, USA) to construct a recombinant plasmid. The primers for standards (sense: 5′-TATTCCCGTTGATGAGGT-3′; antisense: 5′-GCAACCTTATAGCCCTCT-3′) and for qPCR (sense: 5′-TCTTGTGTTGGCACTGTCAC-3′; antisense: 5′-TGCAAGCCATAAGGATGCTG-3′) were synthesized by Sangon Company (Shanghai, China). Real-time RT-PCR was carried out using ABI 7500-fast Real-Time PCR systems (ABI, Foster City, CA, USA). Each 20 μL qPCR reaction system contained 2 μL reverse transcription sample, 10 μL TliRNaseH Plus (2×), 0.4 μL forward and reverse primers (10 μM), 0.4 μL ROX Reference Dye II (50×), and 6.8 μL sterile purified water. Amplification conditions were 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s and 60 °C for 34 s. All samples and standards were carried out in triplicate. The PEDV standards (recombinant plasmid) were diluted serially tenfold (101 to 108 copies/μL) to perform qPCR and establish a standard curve. The concentration of each sample was calculated by plotting the Ct value against the standard curve.
2.10. Statistical Analysis
Statistical analyses (one-way ANOVA) were performed using GraphPad Prism 8.0 software. All experiments were performed in three independent experiments, and data were expressed as mean values ± standard error of the mean (SEM). The statistical significances were defined as p < 0.05 (*), and the higher significance was denoted by p < 0.01 (**), p < 0.001 (***), and p < 0.0001 (****). Data relating to viral RNA copies and virus titers were converted to log10 to maintain a normal distribution.
4. Discussion
GS-441524, like the active form of remdesivir in vivo, is phosphorylated into GS-441524 triphosphate by cell kinases, competing against the endogenous ATP for recognition and incorporation by the viral RNA-dependent RNA-polymerases (RdRp) [
18]. To date, GS-441524 has been reported to exhibit broad-spectrum antiviral activity against a variety of RNA viruses, including a broad spectrum of coronaviruses, such as SARS-CoV [
12], Middle Eastern respiratory syndrome coronavirus (MERS-CoV) [
19], mouse hepatitis virus (MHV) [
20], and other zoonotic coronaviruses [
21]. GS-441524 treatment can rapidly reverse the disease symptoms of FIP, allowing sick cats to return to normal without significant toxicity [
21]. GS-441524 and remdesivir potently inhibited SARS-CoV-2 replication in a dose-dependent manner, and GS-441524 had stronger anti-SARS-CoV-2 activity than remdesivir [
22]. In addition, GS-441524 displayed a favorable oral bioavailability of 57% and produced adequate drug exposure in mouse plasma and lungs, suggesting that it could be a promising oral antiviral drug for the treatment of COVID-19 [
13].
In this study, we investigated the antiviral activity of GS-441524 against trypsin-independent PEDV and confirmed that it could also significantly inhibit the proliferation of trypsin-independent PEDV, with an EC
50 value of 2.6 μM and the highest SIs (selective indices) of 40.15. We further showed that GS-441524 was capable of markedly suppressing viral amplification and virion production of PEDV in a dose-dependent manner within the concentration of 1.25~20 μM, and 20 μM GS-441524 could almost completely inhibit the proliferation of PEDV. Given that in vitro proliferation of clinical strains of PEDV usually requires the addition of exogenous trypsin, we further investigated the inhibitory effect of GS-441524 on trypsin-dependent PEDV. In order to minimize the influence of genetic background, two PEDV strains, DR13
att strain (trypsin-independent PEDV) and rPEDV-S strain (trypsin-dependent PEDV), with different S genes (the determinant of trypsin-dependency) were selected. The results showed that GS-441524 could potently inhibit the proliferation of trypsin-dependent and trypsin-independent PEDVs tested, but had a stronger inhibitory effect on trypsin-independent PEDV strain. As the genetic backgrounds of the two PEDV strains, including the coding gene for RdRp, which is the target of GS-441524, are identical except for the S gene, we speculate that the differential inhibitory activities of GS-441524 on the trypsin-dependent and trypsin-independent PEDVs might be attributed to the influence of trypsin on drug transmembrane transport. The transmembrane transport of nucleoside analog drugs mainly relies on equilibrative nucleoside transporters (ENTs) [
23]. Trypsin added during rPEDV-S cultivation may reduce the transport efficiency of GS-441524 by degrading ENTs or altering cell membrane structure, thereby reducing the drug’s inhibitory efficacy against trypsin-dependent PEDV. However, this hypothesis and its underlying mechanisms require further experimental verification.
It is well-established that GS-441524 inhibits the activity of RdRp in its triphosphate form. Consistent with this, we found that administering 20 μM GS-441524 six hours after PEDV entered Vero cells could completely inhibit virus proliferation, while administering it during the adsorption and entry stages of PEDV infection had no significant effect on virus titers and structural protein expression. In addition, 20 μM GS-441524 effectively inhibited the proliferation of rPEDV-∆ORF3-GFP with an MOI less than 0.5. Although the fluorescence signal of the GFP protein intensifies with an increase in viral infection dose, the virus titer could not be detected. Taken together, these results suggested that GS-441524 was a potential candidate drug for the treatment of PEDV infection.
Several studies have shown that nucleoside analogs hold the potential as antiviral drugs against PEDV. Specifically, molnupiravir, N6-methyladenosine, beta-D-N(4)-hydroxycytidine, and ribavirin have been shown to have a significant inhibitory effect on PEDV [
10,
24,
25]. Xie’s research revealed that remdesivir nucleoside (RDV-N) was more efficacious (EC
50, 0.31 μM) in Vero E6 cells than remdesivir (EC
50, 0.74 μM) and beta-D-N(4)-hydroxycytidine (EC
50, 1.17 μM) [
15]. Huang et al. compared the anti-PEDV activity of molnupiravir and remdesivir and found that remdesivir has stronger anti-PEDV activity than molnupiravir [
26]. In our study, GS-441524 showed strong anti-PEDV activity with an EC
50 of 2.6 μM, which is higher than the EC
50 value of 0.31 reported in a previous study [
15]. The antiviral activity of the compound is closely related to the viral strain, cell line, and viral infection dose. In the two studies, the viral infection dose was the same, both being 0.01 MOI. However, the viral strains and cell lines used were different. This may lead to the difference in the EC
50 values. In our study, GS-441524 showed a low cytotoxicity (CC
50 = 104.4 μM), which was far lower than that of remdesivir (CC
50, 1.7–15.0 μM) [
27], indicating that GS-441524 has distinct advantages over remdesivir in treating PEDV infection.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}