
Genetic Basis of Gap Formation Between Migrating Helicobacter pylori Colonies in Soft Agar Assays



Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Soft Agar Motility Assays
2.3. Isolation of Tn-7 Mutants Without Clear Gap Formation Phenotype
2.4. Identification of Transposon Insertion Sites
2.5. Genomic Transformations
2.6. Microscopy
2.7. Growth Curves
2.8. Clear Gap Material Assessment
3. Results
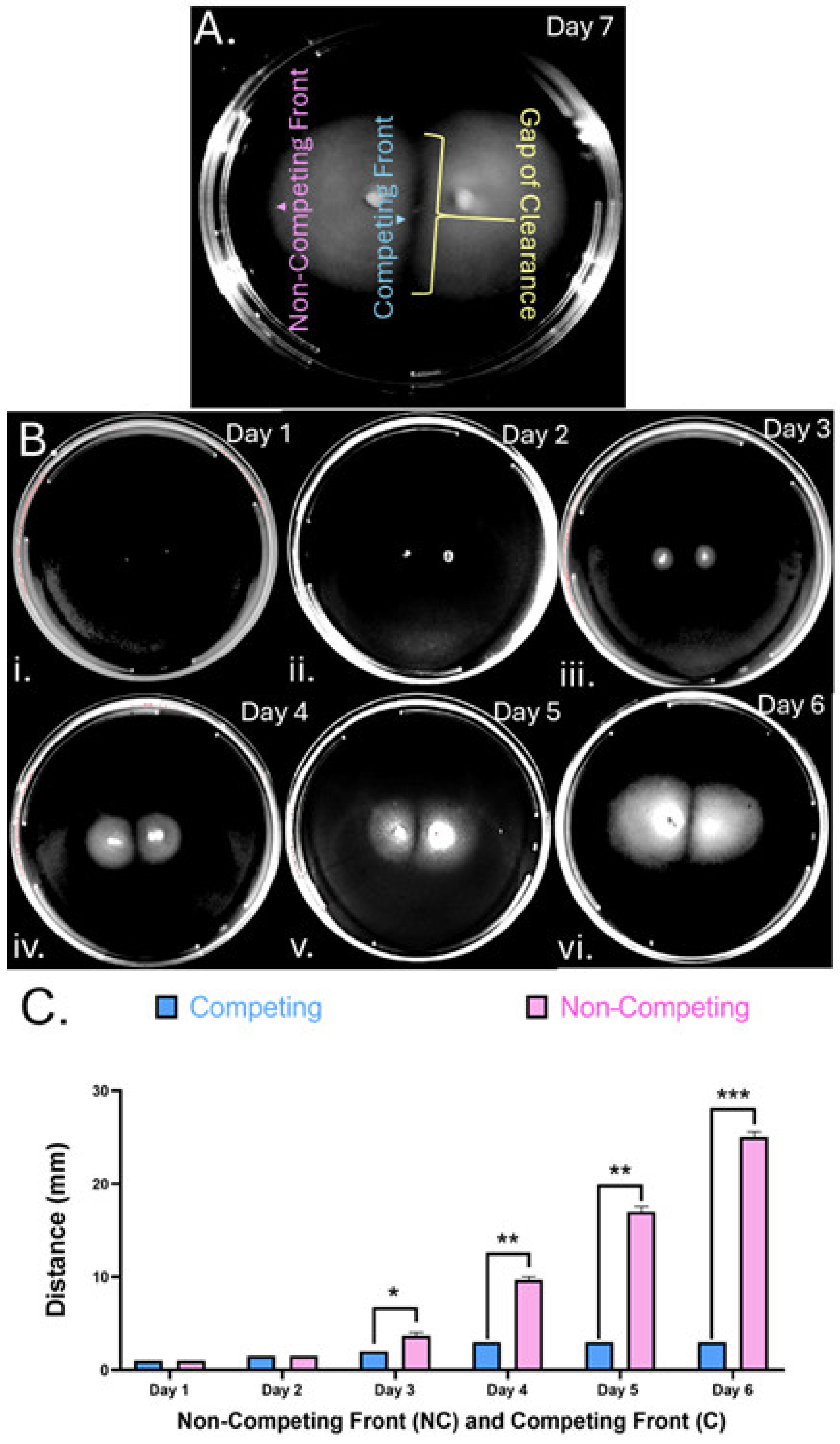
3.1. H. pylori Ceases Soft Agar Migration and Forms a Gap When Two Colonies Meet
3.2. Formation of Colony Gaps Is Chemotaxis-Independent
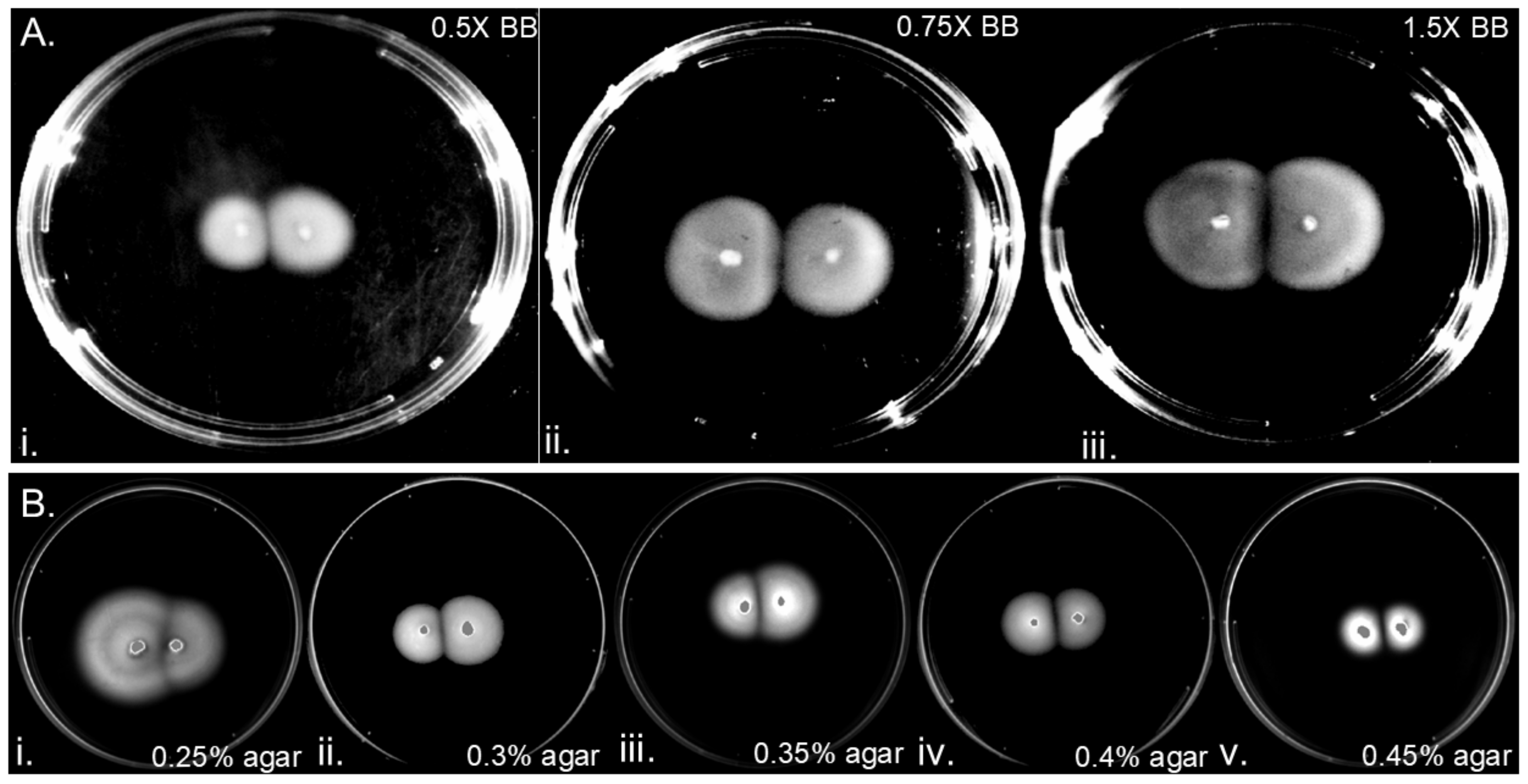
3.3. H. pylori Colony Gap Formation Is Independent of Nutrient Level and Agar Percentages
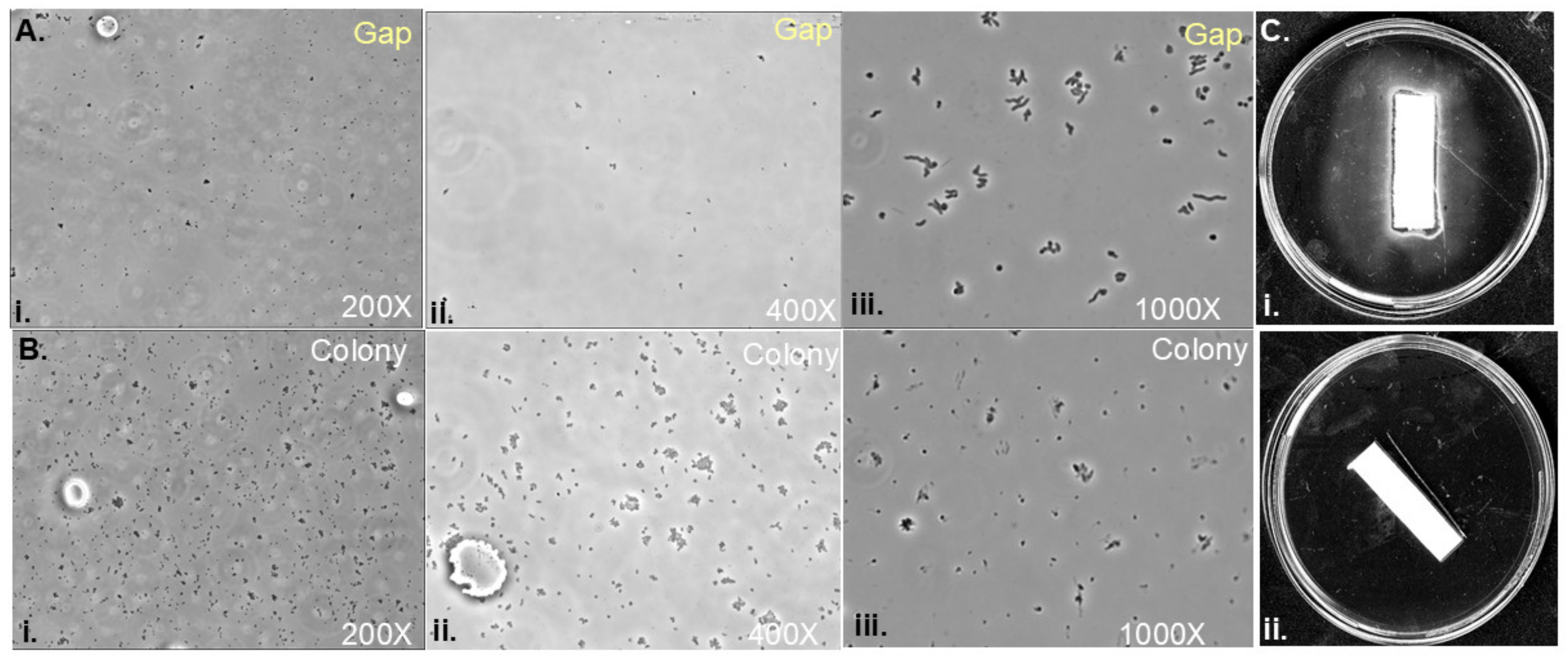
3.4. Soft Agar Colony Gaps Contain Low Densities of H. pylori Cells with Spiral Morphology
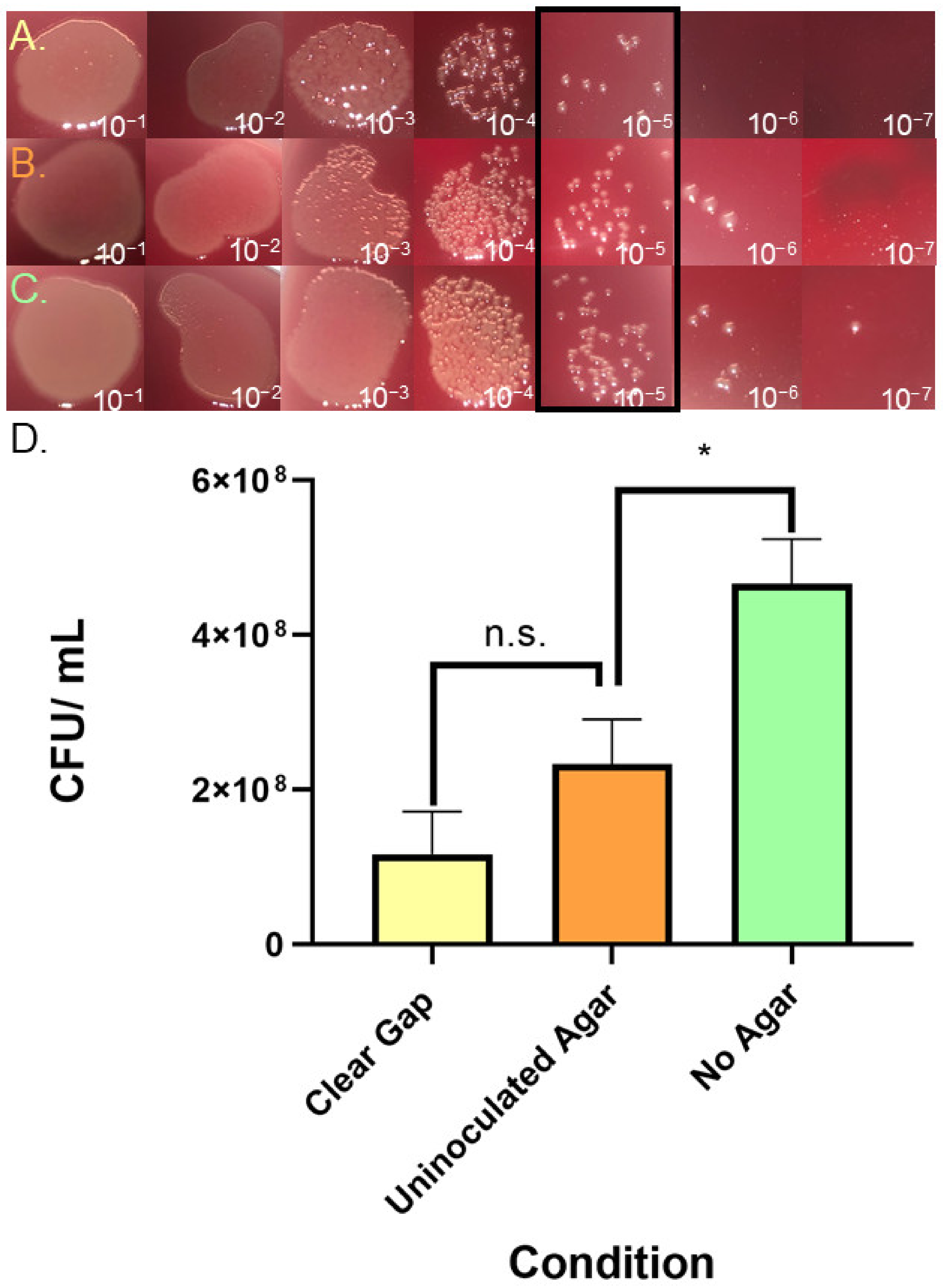
3.5. Assessing Inhibitory Compounds in Gap Regions
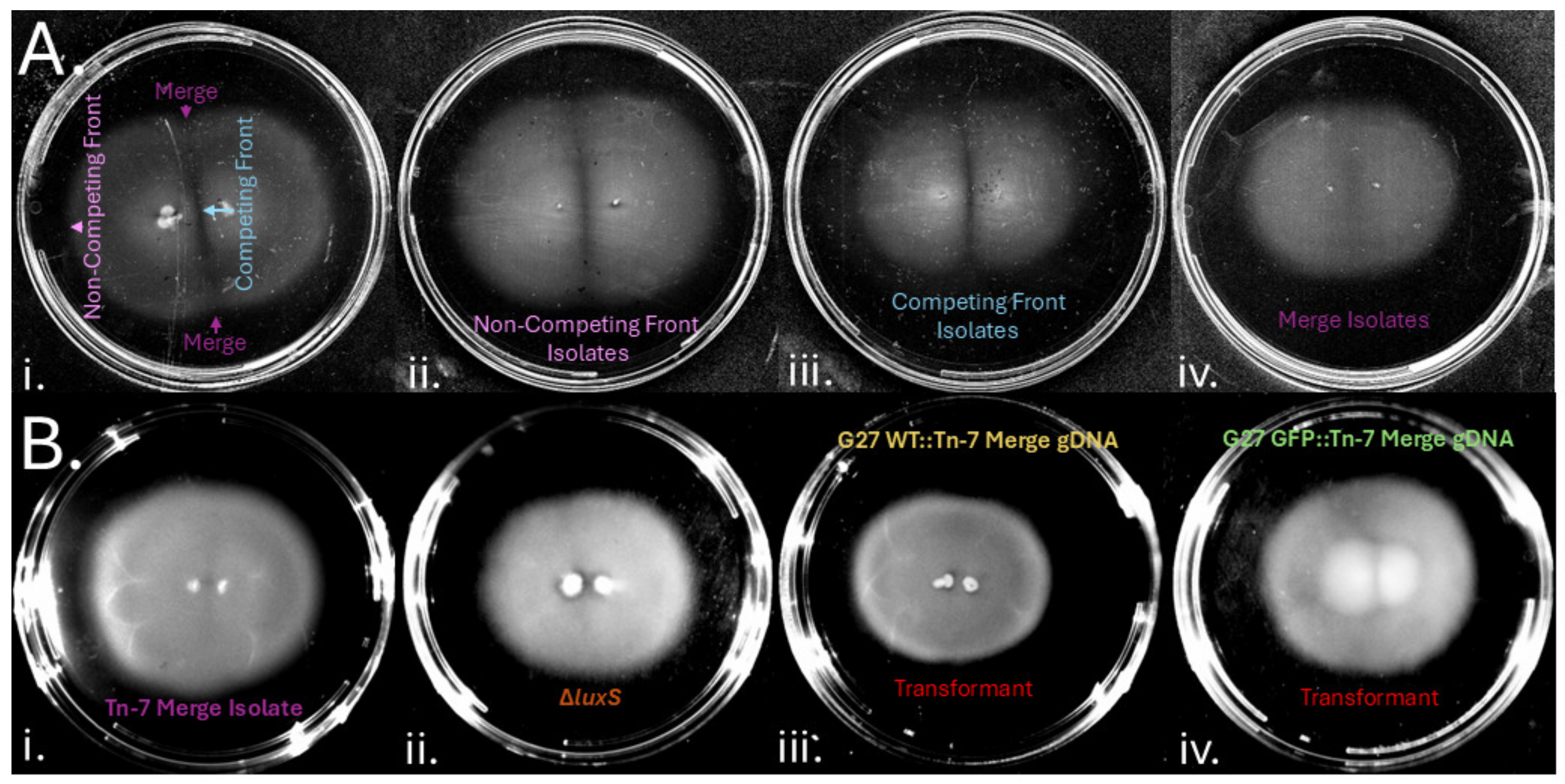
3.6. H. pylori Gap Formation Has a Genetic Basis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wuichet, K.; Zhulin, I.B. Origins and Diversification of a Complex Signal Transduction System in Prokaryotes. Sci. Signal. 2010, 3, ra50. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, A.J.; Berg, H.C. Migration of Bacteria in Semisolid Agar. Proc. Natl. Acad. Sci. USA 1989, 86, 6973–6977. [Google Scholar] [CrossRef] [PubMed]
- Croze, O.A.; Ferguson, G.P.; Cates, M.E.; Poon, W.C.K. Migration of Chemotactic Bacteria in Soft Agar: Role of Gel Concentration. Biophys. J. 2011, 101, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Sanders, L.; Andermann, T.M.; Ottemann, K.M. A Supplemented Soft Agar Chemotaxis Assay Demonstrates the Helicobacter pylori Chemotactic Response to Zinc and Nickel. Microbiology 2013, 159, 46–57. [Google Scholar] [CrossRef]
- Sekowska, A.; Masson, J.-B.; Celani, A.; Danchin, A.; Vergassola, M. Repulsion and Metabolic Switches in the Collective Behavior of Bacterial Colonies. Biophys. J. 2009, 97, 688–698. [Google Scholar] [CrossRef]
- Espeso, D.R.; Martínez-García, E.; De Lorenzo, V.; Goñi-Moreno, Á. Physical Forces Shape Group Identity of Swimming Pseudomonas putida Cells. Front. Microbiol. 2016, 7, 01437. [Google Scholar] [CrossRef]
- Strassmann, J.E.; Gilbert, O.M.; Queller, D.C. Kin Discrimination and Cooperation in Microbes. Annu. Rev. Microbiol. 2011, 65, 349–367. [Google Scholar] [CrossRef]
- Tipping, M.J.; Gibbs, K.A. Peer Pressure from a Proteus mirabilis Self-Recognition System Controls Participation in Cooperative Swarm Motility. PLoS Pathog. 2019, 15, e1007885. [Google Scholar] [CrossRef]
- Budding, A.E.; Ingham, C.J.; Bitter, W.; Vandenbroucke-Grauls, C.M.; Schneeberger, P.M. The Dienes Phenomenon: Competition and Territoriality in Swarming Proteus mirabilis. J. Bacteriol. 2009, 191, 3892–3900. [Google Scholar] [CrossRef]
- Nadell, C.D.; Drescher, K.; Wingreen, N.S.; Bassler, B.L. Extracellular Matrix Structure Governs Invasion Resistance in Bacterial Biofilms. ISME J. 2015, 9, 1700–1709. [Google Scholar] [CrossRef]
- Fung, C.; Tan, S.; Nakajima, M.; Skoog, E.C.; Camarillo-Guerrero, L.F.; Klein, J.A.; Lawley, T.D.; Solnick, J.V.; Fukami, T.; Amieva, M.R. High-Resolution Mapping Reveals That Microniches in the Gastric Glands Control Helicobacter pylori Colonization of the Stomach. PLoS Biol. 2019, 17, e3000231. [Google Scholar] [CrossRef] [PubMed]
- Foynes, S.; Dorrell, N.; Ward, S.J.; Stabler, R.A.; McColm, A.A.; Rycroft, A.N.; Wren, B.W. Helicobacter pylori Possesses Two CheY Response Regulators and a Histidine Kinase Sensor, CheA, Which Are Essential for Chemotaxis and Colonization of the Gastric Mucosa. Infect. Immun. 2000, 68, 2016–2023. [Google Scholar] [CrossRef] [PubMed]
- Sagoo, J.; Abedrabbo, S.; Liu, X.; Ottemann, K.M. Helicobacter pylori cheV1 Mutants Recover Semisolid Agar Migration Due to Loss of a Previously Uncharacterized Type IV Filament Membrane Alignment Complex Homolog. J. Bacteriol. 2024, 206, e00406-23. [Google Scholar] [CrossRef] [PubMed]
- Malfertheiner, P.; Camargo, M.C.; El-Omar, E.; Liou, J.-M.; Peek, R.; Schulz, C.; Smith, S.I.; Suerbaum, S. Helicobacter pylori Infection. Nat. Rev. Dis. Primers 2023, 9, 19. [Google Scholar] [CrossRef]
- Uemura, N.; Yamaguchi, S.; Schlemper, R.J. Helicobacter pylori Infection and the Development of Gastric Cancer. N. Engl. J. Med. 2001, 345, 784–789. [Google Scholar] [CrossRef]
- Polk, D.B.; Peek, R.M. Helicobacter pylori: Gastric Cancer and Beyond. Nat. Rev. Cancer 2010, 10, 403–414. [Google Scholar] [CrossRef]
- Hu, Q.; Zhang, Y.; Zhang, X.; Fu, K. Gastric Mucosa-Associated Lymphoid Tissue Lymphoma and Helicobacter pylori Infection: A Review of Current Diagnosis and Management. Biomark. Res. 2016, 4, 15. [Google Scholar] [CrossRef]
- Spagnuolo, R.; Scarlata, G.G.M.; Paravati, M.R.; Abenavoli, L.; Luzza, F. Change in Diagnosis of Helicobacter pylori Infection in the Treatment-Failure Era. Antibiotics 2024, 13, 357. [Google Scholar] [CrossRef]
- Dang, B.N.; Graham, D.Y. Helicobacter pylori Infection and Antibiotic Resistance: A WHO High Priority? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 383–384. [Google Scholar] [CrossRef]
- Kumar, S.; Araque, M.; Stark, V.S.; Kleyman, L.S.; Cohen, D.A.; Goldberg, D.S. Barriers to Community-Based Eradication of Helicobacter pylori Infection. Clin. Gastroenterol. Hepatol. 2024, 22, 2140–2142.e1. [Google Scholar] [CrossRef]
- Ottemann, K.M.; Lowenthal, A.C. Helicobacter pylori Uses Motility for Initial Colonization and To Attain Robust Infection. Infect. Immun. 2002, 70, 1984–1990. [Google Scholar] [CrossRef] [PubMed]
- Terry, K.; Williams, S.M.; Connolly, L.; Ottemann, K.M. Chemotaxis Plays Multiple Roles during Helicobacter pylori Animal Infection. Infect. Immun. 2005, 73, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.S.; Ottemann, K.M. Colonization, Localization, and Inflammation: The Roles of H. pylori Chemotaxis in Vivo. Curr. Opin. Microbiol. 2018, 41, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Keilberg, D.; Zavros, Y.; Shepherd, B.; Salama, N.R.; Ottemann, K.M. Spatial and Temporal Shifts in Bacterial Biogeography and Gland Occupation during the Development of a Chronic Infection. mBio 2016, 7, e01705-16. [Google Scholar] [CrossRef]
- Liu, X.; Roujeinikova, A.; Ottemann, K.M. FliL Functions in Diverse Microbes to Negatively Modulate Motor Output via Its N-Terminal Region. mBio 2023, 14, e00283-23. [Google Scholar] [CrossRef]
- Censini, S.; Lange, C.; Xiang, Z.; Crabtree, J.E.; Ghiara, P.; Borodovsky, M.; Rappuoli, R.; Covacci, A. Cag, a Pathogenicity Island of Helicobacter pylori, Encodes Type I-Specific and Disease-Associated Virulence Factors. Proc. Natl. Acad. Sci. USA 1996, 93, 14648–14653. [Google Scholar] [CrossRef]
- Tomb, J.-F.; White, O.; Kerlavage, A.R.; Clayton, R.A.; Sutton, G.G.; Fleischmann, R.D.; Ketchum, K.A.; Klenk, H.P.; Gill, S.; Dougherty, B.A.; et al. The Complete Genome Sequence of the Gastric Pathogen Helicobacter pylori. Nature 1997, 388, 539–547. [Google Scholar] [CrossRef]
- Arnold, I.C.; Lee, J.Y.; Amieva, M.R.; Roers, A.; Flavell, R.A.; Sparwasser, T.; Müller, A. Tolerance Rather Than Immunity Protects from Helicobacter pylori–Induced Gastric Preneoplasia. Gastroenterology 2011, 140, 199–209.e8. [Google Scholar] [CrossRef]
- Lee, A.; O’Rourke, J.; De Ungria, M.; Robertson, B.; Daskalopoulos, G.; Dixon, M. A Standardized Mouse Model of Helicobacter pylori Infection: Introducing the Sydney Strain. Gastroenterology 1997, 112, 1386–1397. [Google Scholar] [CrossRef]
- Howitt, M.R.; Lee, J.Y.; Lertsethtakarn, P.; Vogelmann, R.; Joubert, L.-M.; Ottemann, K.M.; Amieva, M.R. ChePep Controls Helicobacter pylori Infection of the Gastric Glands and Chemotaxis in the Epsilonproteobacteria. mBio 2011, 2, e00098-11. [Google Scholar] [CrossRef]
- Pittman, M.S.; Goodwin, M.; Kelly, D.J. Chemotaxis in the Human Gastric Pathogen Helicobacter pylori: Different Roles for CheW and the Three CheV Paralogues, and Evidence for CheV2 Phosphorylation. Microbiology 2001, 147, 2493–2504. [Google Scholar] [CrossRef] [PubMed]
- Lowenthal, A.C.; Simon, C.; Fair, A.S.; Mehmood, K.; Terry, K.; Anastasia, S.; Ottemann, K.M. A Fixed-Time Diffusion Analysis Method Determines That the Three Chev Genes of Helicobacter pylori Differentially Affect Motility. Microbiology 2009, 155, 1181–1191. [Google Scholar] [CrossRef] [PubMed]
- Andermann, T.M.; Chen, Y.-T.; Ottemann, K.M. Two Predicted Chemoreceptors of Helicobacter pylori Promote Stomach Infection. Infect. Immun. 2002, 70, 5877–5881. [Google Scholar] [CrossRef] [PubMed]
- McGee, D.J.; Langford, M.L.; Watson, E.L.; Carter, J.E.; Chen, Y.-T.; Ottemann, K.M. Colonization and Inflammation Deficiencies in Mongolian Gerbils Infected by Helicobacter pylori Chemotaxis Mutants. Infect. Immun. 2005, 73, 1820–1827. [Google Scholar] [CrossRef]
- Collins, K.D.; Andermann, T.M.; Draper, J.; Sanders, L.; Williams, S.M.; Araghi, C.; Ottemann, K.M. The Helicobacter pylori CZB Cytoplasmic Chemoreceptor TlpD Forms an Autonomous Polar Chemotaxis Signaling Complex That Mediates a Tactic Response to Oxidative Stress. J. Bacteriol. 2016, 198, 1563–1575. [Google Scholar] [CrossRef]
- Salama, N.R.; Shepherd, B.; Falkow, S. Global Transposon Mutagenesis and Essential Gene Analysis of Helicobacter pylori. J. Bacteriol. 2004, 186, 7926–7935. [Google Scholar] [CrossRef]
- DeLoney-Marino, C.R.; Wolfe, A.J.; Visick, K.L. Chemoattraction of Vibrio fischeri to Serine, Nucleosides, and N-Acetylneuraminic Acid, a Component of Squid Light-Organ Mucus. Appl. Environ. Microbiol. 2003, 69, 7527–7530. [Google Scholar] [CrossRef]
- Shen, P.; Lees, J.A.; Bee, G.C.W.; Brown, S.P.; Weiser, J.N. Pneumococcal Quorum Sensing Drives an Asymmetric Owner–Intruder Competitive Strategy during Carriage via the Competence Regulon. Nat. Microbiol. 2018, 4, 198–208. [Google Scholar] [CrossRef]
- Odenbreit, S.; Faller, G.; Haas, R. Role of the AlpAB Proteins and Lipopolysaccharide in Adhesion of Helicobacter pylori to Human Gastric Tissue. Int. J. Med. Microbiol. 2002, 292, 247–256. [Google Scholar] [CrossRef]
- Odenbreit, S.; Swoboda, K.; Barwig, I.; Ruhl, S.; Borén, T.; Koletzko, S.; Haas, R. Outer Membrane Protein Expression Profile in Helicobacter pylori Clinical Isolates. Infect. Immun. 2009, 77, 3782–3790. [Google Scholar] [CrossRef]
- Senkovich, O.A.; Yin, J.; Ekshyyan, V.; Conant, C.; Traylor, J.; Adegboyega, P.; McGee, D.J.; Rhoads, R.E.; Slepenkov, S.; Testerman, T.L. Helicobacter pylori AlpA and AlpB Bind Host Laminin and Influence Gastric Inflammation in Gerbils. Infect. Immun. 2011, 79, 3106–3116. [Google Scholar] [CrossRef] [PubMed]
- Yonezawa, H.; Osaki, T.; Woo, T.; Kurata, S.; Zaman, C.; Hojo, F.; Hanawa, T.; Kato, S.; Kamiya, S. Analysis of Outer Membrane Vesicle Protein Involved in Biofilm Formation of Helicobacter pylori. Anaerobe 2011, 17, 388–390. [Google Scholar] [CrossRef] [PubMed]
- Yonezawa, H.; Osaki, T.; Fukutomi, T.; Hanawa, T.; Kurata, S.; Zaman, C.; Hojo, F.; Kamiya, S. Diversification of the AlpB Outer Membrane Protein of Helicobacter pylori Affects Biofilm Formation and Cellular Adhesion. J. Bacteriol. 2017, 199, e00729-16. [Google Scholar] [CrossRef] [PubMed]
- De Jonge, R.; Durrani, Z.; Rijpkema, S.G.; Kuipers, E.J.; Van Vliet, A.H.M.; Kusters, J.G. Role of the Helicobacter pylori Outer-Membrane Proteins AlpA and AlpB in Colonization of the Guinea Pig Stomach. J. Med. Microbiol. 2004, 53, 375–379. [Google Scholar] [CrossRef]
- Odenbreit, S.; Till, M.; Hofreuter, D.; Faller, G.; Haas, R. Genetic and Functional Characterization of the alpAB Gene Locus Essential for the Adhesion of Helicobacter pylori to Human Gastric Tissue. Mol. Microbiol. 1999, 31, 1537–1548. [Google Scholar] [CrossRef]
- Lu, H.; Wu, J.Y.; Beswick, E.J.; Ohno, T.; Odenbreit, S.; Haas, R.; Reyes, V.E.; Kita, M.; Graham, D.Y.; Yamaoka, Y. Functional and Intracellular Signaling Differences Associated with the Helicobacter pylori AlpAB Adhesin from Western and East Asian Strains. J. Biol. Chem. 2007, 282, 6242–6254. [Google Scholar] [CrossRef]
- Hathroubi, S.; Hu, S.; Ottemann, K.M. Genetic Requirements and Transcriptomics of Helicobacter pylori Biofilm Formation on Abiotic and Biotic Surfaces. NPJ Biofilms Microbiomes 2020, 6, 56. [Google Scholar] [CrossRef]
- Voss, B.J.; Loh, J.T.; Hill, S.; Rose, K.L.; McDonald, W.H.; Cover, T.L. Alteration of the Helicobacter pylori Membrane Proteome in Response to Changes in Environmental Salt Concentration. Proteom. Clin. Appl. 2015, 9, 1021–1034. [Google Scholar] [CrossRef]
- Caston, R.R.; Loh, J.T.; Voss, B.J.; McDonald, W.H.; Scholz, M.B.; McClain, M.S.; Cover, T.L. Effect of Environmental Salt Concentration on the Helicobacter pylori Exoproteome. J. Proteom. 2019, 202, 103374. [Google Scholar] [CrossRef]
- Joyce, E.A.; Bassler, B.L.; Wright, A. Evidence for a Signaling System in Helicobacter pylori: Detection of a luxS-Encoded Autoinducer. J. Bacteriol. 2000, 182, 3638–3643. [Google Scholar] [CrossRef]
- Rader, B.A.; Campagna, S.R.; Semmelhack, M.F.; Bassler, B.L.; Guillemin, K. The Quorum-Sensing Molecule Autoinducer 2 Regulates Motility and Flagellar Morphogenesis in Helicobacter pylori. J. Bacteriol. 2007, 189, 6109–6117. [Google Scholar] [CrossRef] [PubMed]
- Rader, B.A.; Wreden, C.; Hicks, K.G.; Sweeney, E.G.; Ottemann, K.M.; Guillemin, K. Helicobacter pylori Perceives the Quorum-Sensing Molecule AI-2 as a Chemorepellent via the Chemoreceptor TlpB. Microbiology 2011, 157, 2445–2455. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Huang, X.; Zhang, X.; Zhang, X.; Xu, X.; She, F.; Wen, Y. AI-2 Induces Urease Expression Through Downregulation of Orphan Response Regulator HP1021 in Helicobacter pylori. Front. Med. 2022, 9, 790994. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.K.; Huang, J.Y.; Wreden, C.; Sweeney, E.G.; Goers, J.; Remington, S.J.; Guillemin, K. Chemorepulsion from the Quorum Signal Autoinducer-2 Promotes Helicobacter pylori Biofilm Dispersal. mBio 2015, 6, e00379-15. [Google Scholar] [CrossRef]
- Wen, Y.; Huang, H.; Tang, T.; Yang, H.; Wang, X.; Huang, X.; Gong, Y.; Zhang, X.; She, F. AI-2 represses CagA expression and bacterial adhesion, attenuating the Helicobacter pylori-induced inflammatory response of gastric epithelial cells. Helicobacter 2021, 26, e12778. [Google Scholar] [CrossRef]
- Czechowska, K.; McKeithen-Mead, S.; Al Moussawi, K.; Kazmierczak, B.I. Cheating by Type 3 Secretion System-Negative Pseudomonas aeruginosa during Pulmonary Infection. Proc. Natl. Acad. Sci. USA 2014, 111, 7801–7806. [Google Scholar] [CrossRef]
- Rizvanov, A.A.; Haertlé, T.; Bogomolnaya, L.; Talebi Bezmin Abadi, A. Helicobacter pylori and Its Antibiotic Heteroresistance: A Neglected Issue in Published Guidelines. Front. Microbiol. 2019, 10, 1796. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Disrupted by Tn-7 | Gene Function | Number of Isolates |
|---|---|---|
| HPG27_843 | Outer membrane protein HopC/AlpA | 3 |
| Intergenic HPG27_843&844 | Outer membrane protein HopCB/AlpAB locus | 2 |
| HPG27_147 | Cysteine-rich protein D | 1 |
| HPG27_412 | Phosphatidyl glycerophosphate synthase | 1 |
| HPG27_413 | Hypothetical protein | 1 |
| HPG27_462 | Hypothetical protein | 1 |
| HPG27_1061 | Cysteine-rich protein X | 1 |
| HPG27_1062 | Hypothetical protein | 1 |
| HPG27_1299 | Type II endorestriction nuclease HpyAIV | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elshenawi, Y.; Hathroubi, S.; Hu, S.; Liu, X.; Ottemann, K.M. Genetic Basis of Gap Formation Between Migrating Helicobacter pylori Colonies in Soft Agar Assays. Microorganisms 2025, 13, 1087. https://doi.org/10.3390/microorganisms13051087
Elshenawi Y, Hathroubi S, Hu S, Liu X, Ottemann KM. Genetic Basis of Gap Formation Between Migrating Helicobacter pylori Colonies in Soft Agar Assays. Microorganisms. 2025; 13(5):1087. https://doi.org/10.3390/microorganisms13051087
Chicago/Turabian StyleElshenawi, Yasmine, Skander Hathroubi, Shuai Hu, Xiaolin Liu, and Karen M. Ottemann. 2025. "Genetic Basis of Gap Formation Between Migrating Helicobacter pylori Colonies in Soft Agar Assays" Microorganisms 13, no. 5: 1087. https://doi.org/10.3390/microorganisms13051087
APA StyleElshenawi, Y., Hathroubi, S., Hu, S., Liu, X., & Ottemann, K. M. (2025). Genetic Basis of Gap Formation Between Migrating Helicobacter pylori Colonies in Soft Agar Assays. Microorganisms, 13(5), 1087. https://doi.org/10.3390/microorganisms13051087