
Diversity, Composition, and Ecological Function of Endophytic Fungal Communities Associated with Erigeron breviscapus in China


Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Sample Pretreatment
2.3. DNA Extraction and Sequencing
2.4. Processing of Sequencing Data
2.5. Statistical Analysis
3. Results
3.1. Sequencing Yields
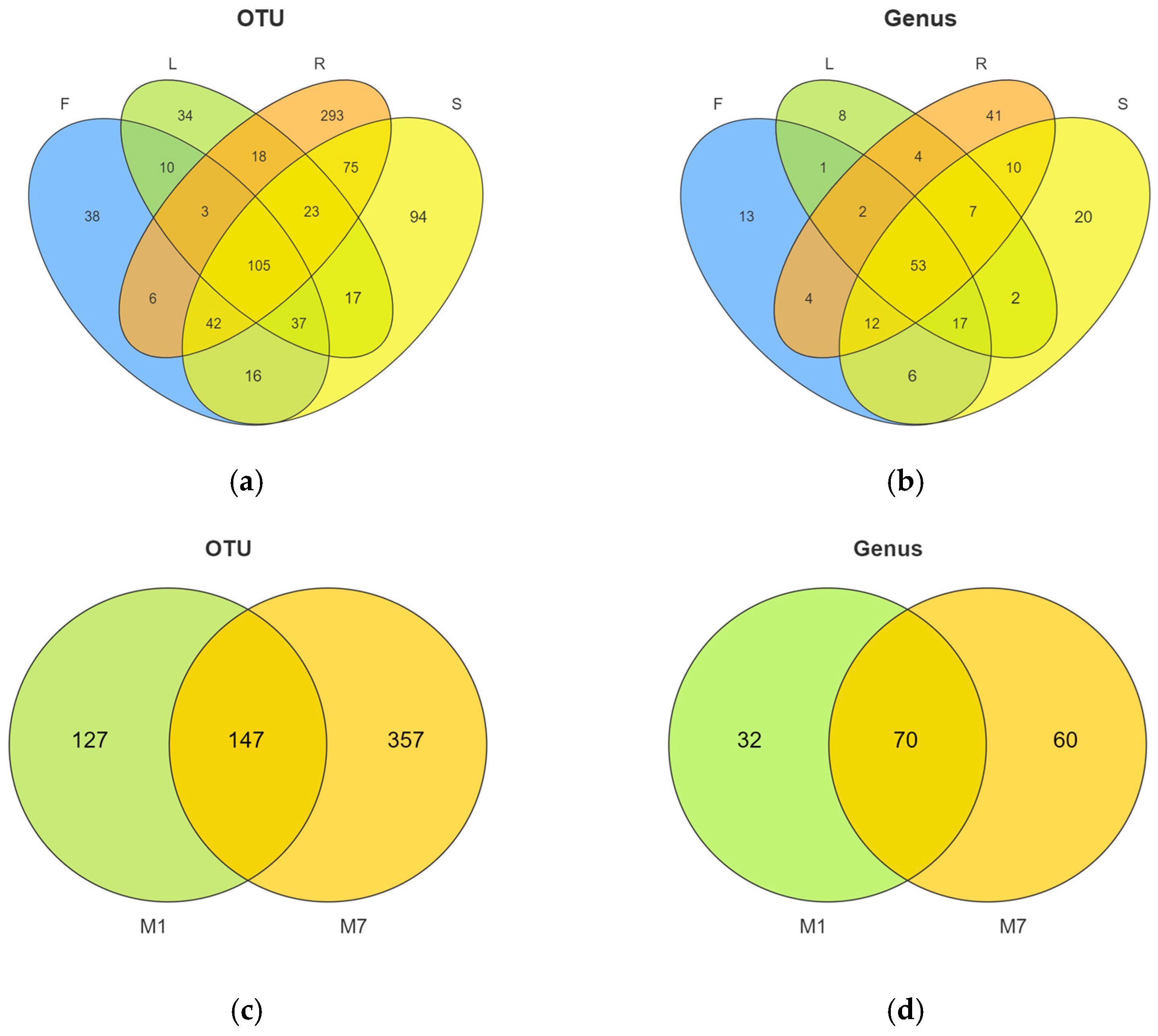
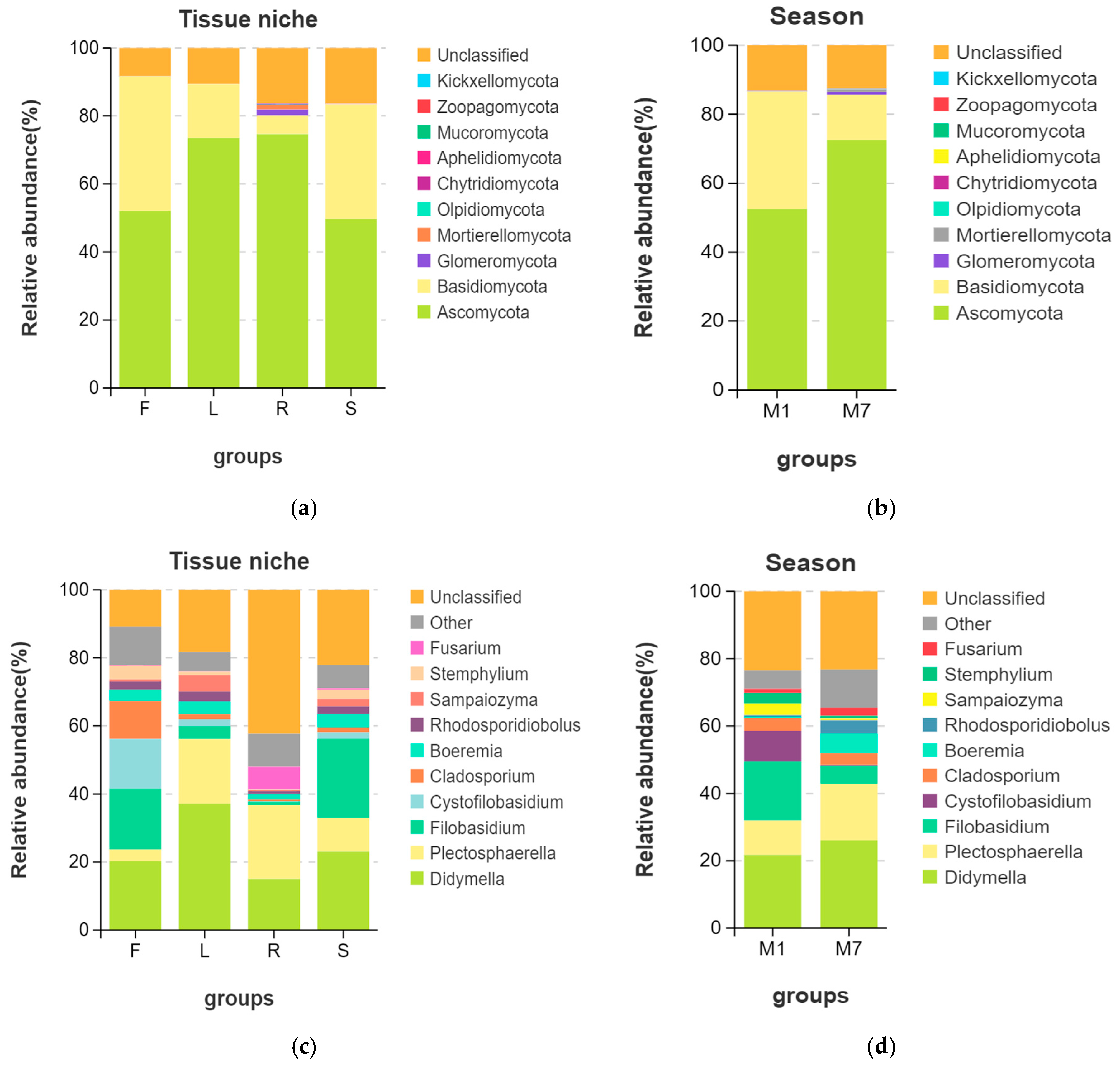
3.2. Fungal Endophytic Community Composition
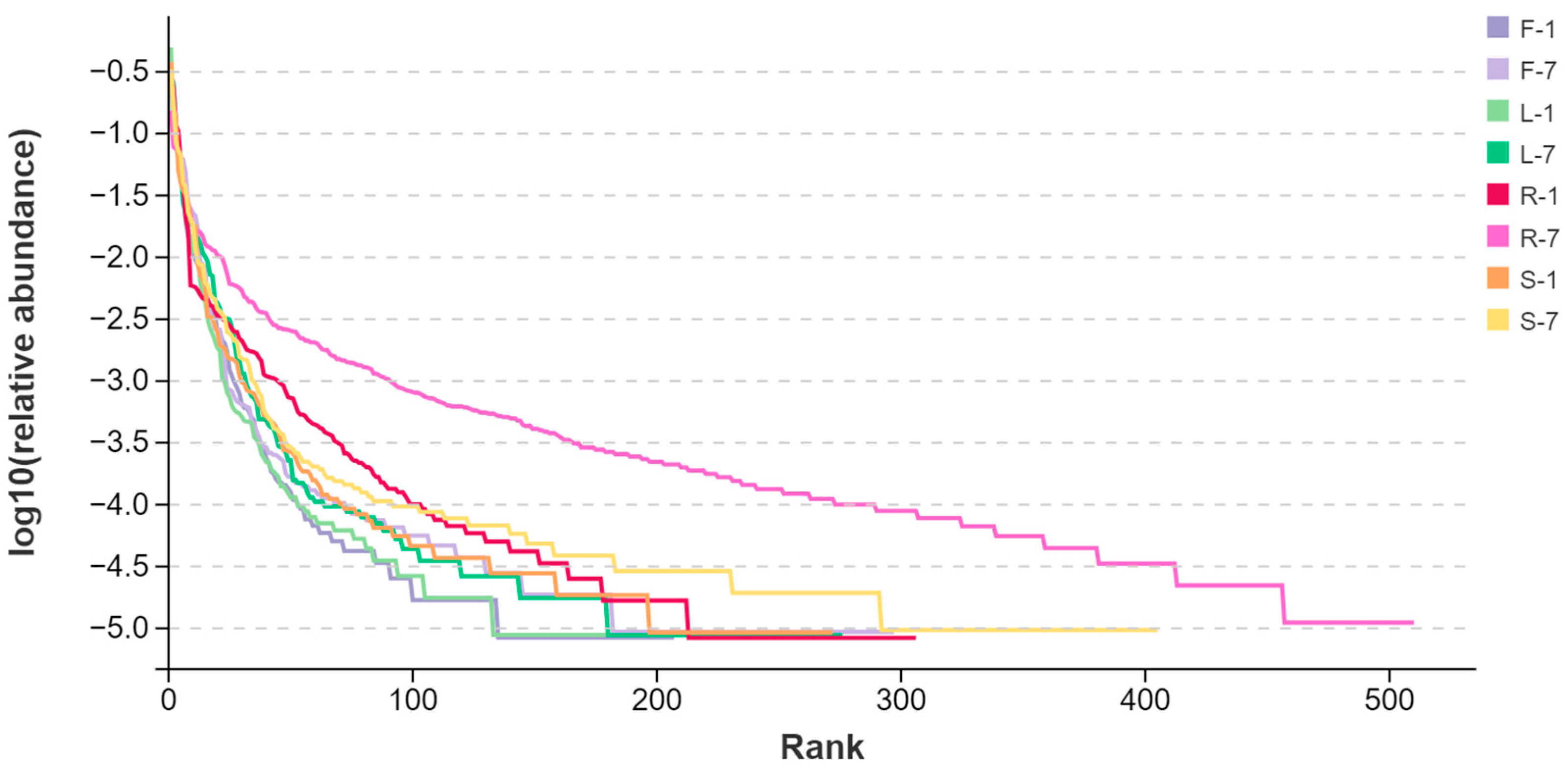
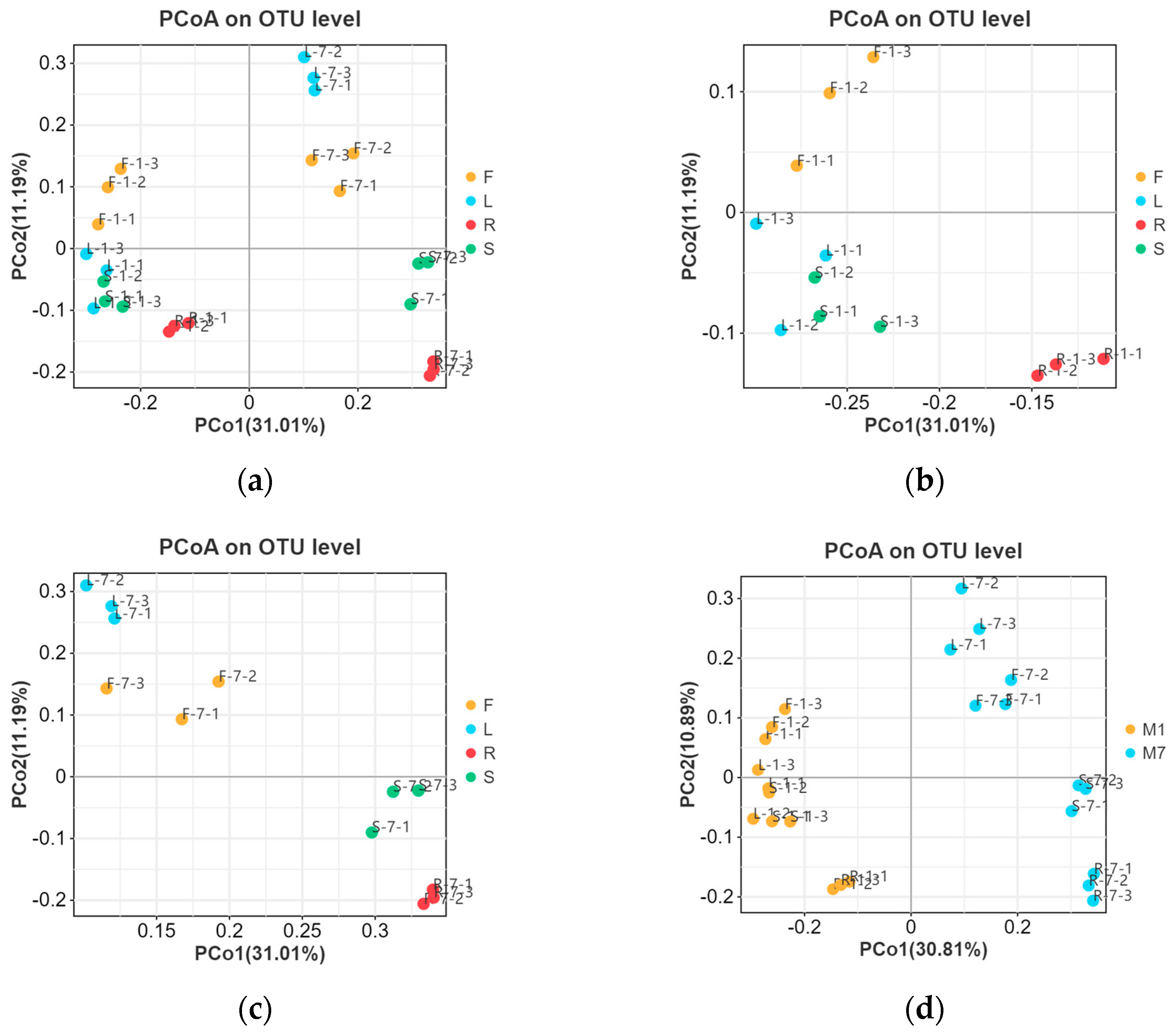
3.3. Diversity of Fungal Endophytes
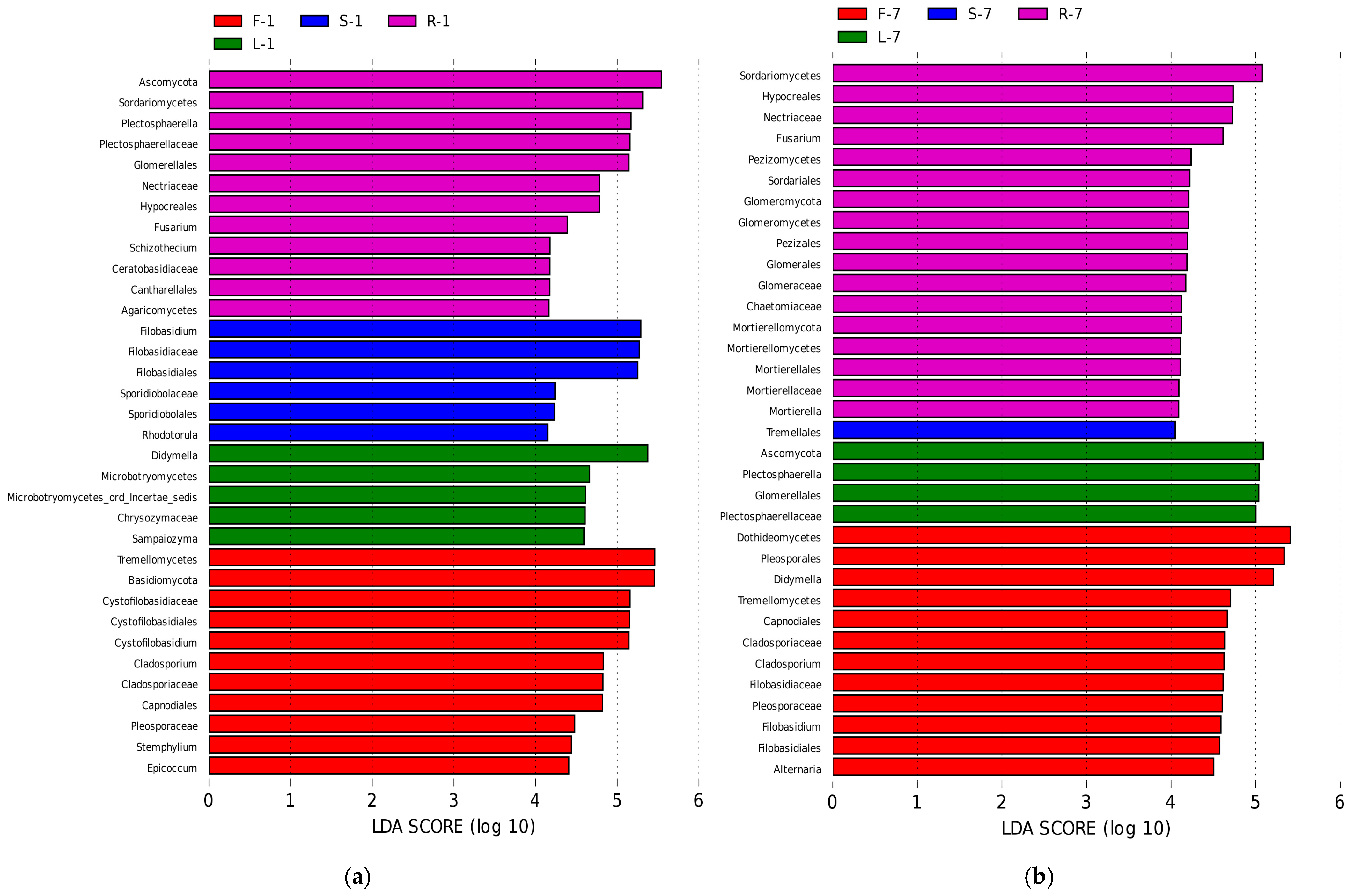
3.4. Biomarkers in Different Organs
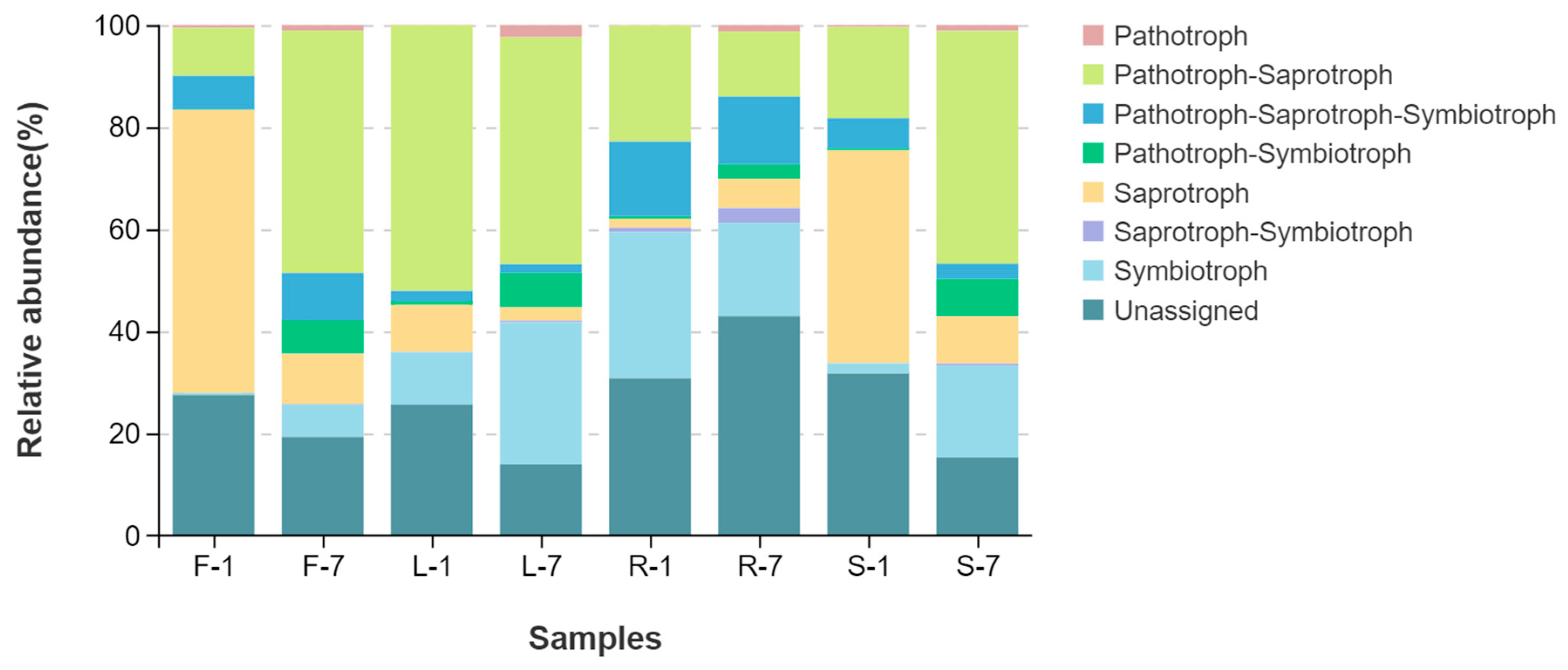
3.5. Fungal Ecological Functions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, X.; Cheng, J.; Zhang, G.; Ding, W.; Duan, L.; Yang, J.; Kui, L.; Cheng, X.; Ruan, J.; Fan, W.; et al. Engineering yeast for the production of breviscapine by genomic analysis and synthetic biology approaches. Nat. Commun. 2018, 9, 448. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Liang, Y.; Xu, M.; Sun, M.; Sun, W.; Zhou, Y.; Huang, X.; Song, W.; Liang, Y.; Wang, Z. Protective mechanism of Erigeron breviscapus injection on blood–brain barrier injury induced by cerebral ischemia in rats. Sci. Rep. 2021, 11, 18451. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yu, Y.; Guo, J.; Zhang, Y.; Huang, L. Rapid and Efficient Optimization Method for a Genetic Transformation System of Medicinal Plants Erigeron breviscapus. Int. J. Mol. Sci. 2023, 24, 5611. [Google Scholar] [CrossRef]
- Dong, X.; Qu, S. Erigeron breviscapus (Vant.) Hand-Mazz.: A Promising Natural Neuroprotective Agent for Alzheimer’s Disease. Front. Pharmacol. 2022, 13, 877872. [Google Scholar] [CrossRef]
- Wu, R.; Liang, Y.; Xu, M.; Fu, K.; Zhang, Y.; Wu, L.; Wang, Z. Advances in Chemical Constituents, Clinical Applications, Pharmacology, Pharmacokinetics and Toxicology of Erigeron Breviscapus. Front. Pharmacol. 2021, 12, 656335. [Google Scholar] [CrossRef]
- Yu, Y.; Ouyang, Z.; Guo, J.; Zeng, W.; Zhao, Y.; Huang, L. Complete Chloroplast Genome Sequence of Erigeron breviscapus and Characterization of Chloroplast Regulatory Elements. Front. Plant Sci. 2021, 12, 758290. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Chen, Y.; Zhao, P.; Yang, S.; He, S.; Long, G. Sulfur fertiliser enhancement of Erigeron breviscapus (Asteraceae) quality by improving plant physiological responses and reducing soil cadmium bioavailability. Environ. Sci. Pollut. Res. Int. 2022, 29, 70508–70519. [Google Scholar] [CrossRef]
- Li, X.; Zhang, S.; Yang, Z.; Song, K.; Yi, T. Conservation genetics and population diversity of Erigeron breviscapus (Asteraceae), an important Chinese herb. Biochem. Syst. Ecol. 2013, 49, 156–166. [Google Scholar] [CrossRef]
- He, S.; Dong, X.; Zhang, G.; Fan, W.; Duan, S.; Shi, H.; Li, D.; Li, R.; Chen, G.; Long, G.; et al. High quality genome of Erigeron Breviscapus provides a reference for herbal plants in Asteraceae. Mol. Ecol. Resour. 2021, 21, 153–169. [Google Scholar] [CrossRef]
- Zhang, W.; Yang, S.C.; Zhang, G.H.; Su, B. Research on situation and countermeasure of Erigeron breviscapus plant production. Chin. Herbal. Med. 2013, 38, 2227–2230. [Google Scholar]
- Chouhan, S.; Agrawal, L.; Prakash, A. Amelioration in traditional farming system by exploring the different plant growth-promoting attributes of endophytes for sustainable agriculture. Arch. Microbiol. 2022, 204, 151. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; Van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Salehi, M.; Safaie, N. Editorial: Endophytic Fungi: Secondary Metabolites and Plant Biotic and Abiotic Stress Management. Front. Microbiol. 2024, 15, 1345210. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Li, S.; Zhang, X.; Zhao, C. Biological Activities of Some New Secondary Metabolites Isolated from Endophytic Fungi: A Review Study. Int. J. Mol. Sci. 2021, 22, 959. [Google Scholar] [CrossRef]
- Nagarajan, K.; Ibrahim, B.; Ahmad Bawadikji, A.; Lim, J.-W.; Tong, W.-Y.; Leong, C.-R.; Khaw, K.Y.; Tan, W.-N. Recent Developments in Metabolomics Studies of Endophytic Fungi. J. Fungi 2021, 8, 28. [Google Scholar] [CrossRef]
- Tsipinana, S.; Husseiny, S.; Alayande, K.A.; Raslan, M.; Amoo, S.; Adeleke, R. Contribution of endophytes towards improving plant bioactive metabolites: A rescue option against red-taping of medicinal plants. Front. Plant Sci. 2023, 14, 1248319. [Google Scholar] [CrossRef]
- Nwobodo, D.C.; Okoye, N.N.; Sifir Mudkhur, M.; Ikem, J.C.; Eze, P.M.; Okoye, F.B.C.; Saki, M.; Esimone, C.O. In Vitro antiplasmodial and anticancer analyses of endophytic fungal extracts isolated from selected Nigerian medicinal plants. Sci. Rep. 2024, 14, 19765. [Google Scholar] [CrossRef]
- Zhao, Y.; Xiong, Z.; Wu, G.; Bai, W.; Zhu, Z.; Gao, Y.; Parmar, S.; Sharma, V.K.; Li, H. Fungal Endophytic Communities of Two Wild Rosa Varieties with Different Powdery Mildew Susceptibilities. Front. Microbiol. 2018, 9, 2462. [Google Scholar] [CrossRef]
- Fan, L.; Li, Y.; Wang, X.; Leng, F.; Li, S.; Zhu, N.; Chen, K.; Wang, Y. Culturable endophytic fungi community structure isolated from Codonopsis pilosula roots and effect of season and geographic location on their structures. BMC Microbiol. 2023, 23, 132. [Google Scholar] [CrossRef]
- Vannier, N.; Bittebiere, A.-K.; Mony, C.; Vandenkoornhuyse, P. Root endophytic Fungi Impact Host Plant Biomass and Respond to Plant Composition at Varying Spatio-Temporal Scales. Fungal Ecol. 2020, 44, 100907. [Google Scholar] [CrossRef]
- Li, H.-Y.; Shen, M.; Zhou, Z.-P.; Li, T.; Wei, Y.; Lin, L. Diversity and cold adaptation of endophytic fungi from five dominant plant species collected from the Baima Snow Mountain, Southwest China. Fungal Divers. 2012, 54, 79–86. [Google Scholar] [CrossRef]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-Coverage ITS Primers for the DNA-Based Identification of Ascomycetes and Basidiomycetes in Environmental Samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinforma. Oxf. Engl. 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinforma. Oxf. Engl. 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinforma. Oxf. Engl. 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Abarenkov, K.; Henrik Nilsson, R.; Larsson, K.-H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE Database for Molecular Identification of Fungi--Recent Updates and Future Perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef]
- Leray, M.; Knowlton, N. Visualizing Patterns of Marine Eukaryotic Diversity from Metabarcoding Data Using QIIME. Methods Mol. Biol. Clifton NJ 2016, 1452, 219–235. [Google Scholar]
- Chen, H.; Boutros, P.C. Venn Diagram: A Package for the Generation of Highly-Customizable Venn and Euler Diagrams in R. BMC Bioinformatics 2011, 12, 35. [Google Scholar] [CrossRef]
- Forsberg, K.J.; Patel, S.; Gibson, M.K.; Lauber, C.L.; Knight, R.; Fierer, N.; Dantas, G. Bacterial Phylogeny Structures Soil Resistomes across Habitats. Nature 2014, 509, 612–616. [Google Scholar] [CrossRef]
- Desgarennes, D.; Garrido, E.; Torres-Gomez, M.J.; Peña-Cabriales, J.J.; Partida-Martinez, L.P. Diazotrophic potential among bacterial communities associated with wild and cultivated agave species. FEMS Microbiol. Ecol. 2014, 90, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Tanunchai, B.; Ji, L.; Schroeter, S.A.; Wahdan, S.F.M.; Hossen, S.; Delelegn, Y.; Buscot, F.; Lehnert, A.-S.; Alves, E.G.; Hilke, I.; et al. FungalTraits vs. FUNGuild: Comparison of Ecological Functional As-signments of Leaf- and Needle-Associated Fungi Across 12 Temperate Tree Species. Microb. Ecol. 2023, 85, 411–428. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Yang, J.; Zhou, L.; Yang, Z.; Liang, J.; Liu, Y.; Ma, X.; Qian, Z.; Hong, P.; Kalueff, A.V.; et al. Marine fungal metabolite butyrolactone I prevents cognitive deficits by relieving inflammation and intestinal microbiota imbalance on aluminum trichloride-injured zebrafish. J. Neuroinflammation 2022, 19, 39. [Google Scholar] [CrossRef]
- Li, L.-Y.; Han, J.; Wu, L.; Fang, C.; Li, W.-G.; Gu, J.-M.; Deng, T.; Qin, C.-J.; Nie, J.-Y.; Zeng, X.-T. Alterations of gut microbiota diversity, composition and metabonomics in testosterone-induced benign prostatic hyperplasia rats. Mil. Med. Res. 2022, 9, 12. [Google Scholar] [CrossRef]
- Li, Z.-Y.; Yang, L.-Y.; Zhou, B.; Li, S.-L.; Chen, Y.-W.; Zhang, Q. Study on endophytic fungi in Erigeron breviscapus Ι: Isolation and identification of endophytic fungi. J. Yunnan Univ., Nat. Sci. Ed. 2003, 25, 65–68. (In Chinese) [Google Scholar]
- Tang, W.; Gong, W.; Xiao, R.; Mao, W.; Zhao, L.; Song, J.; Awais, M.; Ji, X.; Li, H. Endophytic Fungal Community of Stellera chamaejasme L. and Its Possible Role in Improving Host Plants’ Ecological Flexibility in Degraded Grasslands. J. Fungi 2023, 9, 465. [Google Scholar] [CrossRef]
- Jo, Y.; Back, C.-G.; Kim, K.-H.; Chu, H.; Lee, J.H.; Moh, S.H.; Cho, W.K. Using RNA-Sequencing Data to Examine Tissue-Specific Garlic Microbiomes. Int. J. Mol. Sci. 2021, 22, 6791. [Google Scholar] [CrossRef]
- Bonthond, G.; Barilo, A.; Allen, R.J.; Cunliffe, M.; Krueger-Hadfield, S.A. Fungal endophytes vary by species, tissue type, and life cycle stage in intertidal macroalgae. J. Phycol. 2022, 58, 330–342. [Google Scholar] [CrossRef]
- Sharon, O.; Kagan-Trushina, N.; Sharon, A. Wheat fungal endophyte communities are inseparable from the host and influence plant development. mBio 2024, 15, e0253323. [Google Scholar] [CrossRef]
- Luo, J.; Liu, T.; Diao, F.; Hao, B.; Zhang, Z.; Hou, Y.; Guo, W. Shift in rhizospheric and endophytic microbial communities of dominant plants around Sunit Alkaline Lake. Sci. Total Environ. 2023, 867, 161503. [Google Scholar] [CrossRef] [PubMed]
- Perazzolli, M.; Vicelli, B.; Antonielli, L.; Longa, C.M.O.; Bozza, E.; Bertini, L.; Caruso, C.; Pertot, I. Simulated global warming affects endophytic bacterial and fungal communities of Antarctic pearlwort leaves and some bacterial isolates support plant growth at low temperatures. Sci. Rep. 2022, 12, 18839. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Li, X.; Yang, J.; Liu, J.; Zhao, L.; He, X. Fungal Endophytic Community and Diversity Associated with Desert Shrubs Driven by Plant Identity and Organ Differentiation in Extremely Arid Desert Ecosystem. J. Fungi 2021, 7, 578. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, Q.; Ju, M.; Yan, S.; Zhang, Q.; Gu, P. The Endophytic Fungi Diversity, Community Structure, and Ecological Function Prediction of Sophora alopecuroides in Ningxia, China. Microorganisms 2022, 10, 2099. [Google Scholar] [CrossRef]
- Liao, C.; Doilom, M.; Jeewon, R.; Hyde, K.D.; Manawasinghe, I.S.; Chethana, K.W.T.; Balasuriya, A.; Thakshila, S.A.D.; Luo, M.; Mapook, A.; et al. Challenges and Update on Fungal Endophytes: Classification, Definition, Diversity, Ecology, Evolution and Functions. Fungal Divers. 2025, 131, 301–367. [Google Scholar]
- Chen, R.; Chen, X.; Zhu, T.; Liu, J.; Xiang, X.; Yu, J.; Tan, H.; Gao, S.; Li, Q.; Fang, Y.; et al. Integrated Transcript and Metabolite Profiles Reveal That Eb CHI Plays an Important Role in Scutellarin Accumulation in Erigeron breviscapus Hairy Roots. Front. Plant Sci. 2018, 9, 789. [Google Scholar]
- Lu, Y.; Zhang, R.Y.; Duan, Y.; Chen, L.Y.; Zhang, W. Establishment of HPLC Fingerprint for Erigeron breviscapus and Content Determination of Four Components. J. Chin. Med. Mater. 2020, 43, 927–931. (In Chinese) [Google Scholar]
- Pang, B.; Yin, D.; Zhai, Y.; He, A.; Qiu, L.; Liu, Q.; Ma, N.; Shen, H.; Jia, Q.; Liang, Z.; et al. Diversity of Endophytic Fungal Community in Huperzia Serrata from Different Ecological Areas and Their Correlation with Hup A Content. BMC Microbiol. 2022, 22, 191. [Google Scholar] [CrossRef]
- Ma, N.; Yin, D.; Liu, Y.; Gao, Z.; Cao, Y.; Chen, T.; Huang, Z.; Jia, Q.; Wang, D. Succession of Endophytic Fungi and Rhizosphere Soil Fungi and Their Correlation with Secondary Metabolites in Fagopyrum dibotrys. Front. Microbiol. 2023, 14, 1220431. [Google Scholar] [CrossRef]
- Li, N.; Mao, Y.; Huang, Y.; Zhang, L.; Hou, L.; Liu, X.; Du, Y.; Chen, D.; Sun, K. Seasonal Succession of Endophyte and the Association with Active Ingredients in Rheum palmatum. Microbiol. Spectr. 2024, 12, e0118424. [Google Scholar] [CrossRef]
- Mendes, L.W.; Raaijmakers, J.M.; De Hollander, M.; Mendes, R.; Tsai, S.M. Influence of resistance breeding in common bean on rhizosphere microbiome composition and function. ISME J. 2018, 12, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.-Y.; Guo, L.-D.; Hyde, K.D. Response of endophytic fungi of Stipa grandis to experimental plant function group removal in Inner Mongolia steppe, China. Fungal Divers. 2010, 43, 93–101. [Google Scholar] [CrossRef]
- De Oliveira, A.A.; de Oliveira Ramalho, M.; Moreau, C.S.; de Carvalho Campos, A.E.; Harakava, R.; Bueno, O.C. Exploring the diversity and potential interactions of bacterial and fungal endophytes associated with different cultivars of olive (Olea europaea) in Brazil. Microbiol. Res. 2022, 263, 127128. [Google Scholar] [CrossRef]
- Muhammad Atif, M.; Huang, X.; Hou, W.; Zhang, Y.; Cai, Y.; Munir, M.Z.; Wu, L.; Zheng, C. Response of Fungal Diversity, Community Composition, and Functions to Nutrients Management in Red Soil. J. Fungi 2021, 7, 554. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Yang, W.; Diao, L.; Wang, Y.; Yang, X.; Zhang, H.; Wang, J.; Luo, Y.; An, S.; Cheng, X. Responses of soil fungal communities and functional guilds to ~160 years of natural revegetation in the Loess Plateau of China. Front. Microbiol. 2022, 13, 967565. [Google Scholar] [CrossRef]
- Pham, P.; Shi, Y.; Khan, I.; Sumarah, M.; Renaud, J.; Sunohara, M.; Craiovan, E.; Lapen, D.; Aris-Brosou, S.; Chen, W. The functions and factors governing fungal communities and diversity in agricultural waters: Insights into the ecosystem services aquatic mycobiota provide. Front. Microbiol. 2024, 15, 1460330. [Google Scholar] [CrossRef]
- Liang, M.; Liu, X.; Parker, I.M.; Johnson, D.; Zheng, Y.; Luo, S.; Gilbert, G.S.; Yu, S. Soil microbes drive phylogenetic diversity-productivity relationships in a subtropical forest. Sci. Adv. 2019, 5, eaax5088. [Google Scholar] [CrossRef]
- Millberg, H.; Boberg, J.; Stenlid, J. Changes in fungal community of Scots Pine (Pinus sylvestris) needles along a latitudinal gradient in Sweden. Fungal Ecol. 2015, 17, 126–139. [Google Scholar] [CrossRef]
- Li, J.; Hou, L.; Zhang, G.; Cheng, L.; Liu, Y. Comparative Analysis of Rhizosphere and Endosphere Fungal Communities in Healthy and Diseased Faba Bean Plants. J. Fungi 2024, 10, 84. [Google Scholar] [CrossRef]
- Chen, Q.; Jiang, J.R.; Zhang, G.Z.; Cai, L.; Crous, P.W. Resolving the Phoma enigma. Stud. Mycol. 2015, 82, 137–217. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-H.; Choi, K.M.; Jung, C.R.; Lee, S.-H. First report of Didymella bellidis causing Leaf Spots on Angelica gigas in South Korea. J. Plant Pathol. 2020, 102, 1297. [Google Scholar] [CrossRef]
- Wang, X.; Wu, X.; Jiang, S.; Yin, Q.; Li, D.; Wang, Y.; Wang, D.; Chen, Z. Whole Genome Sequence and Gene Annotation Resource for Didymella bellidis Associated with Tea Leaf Spot. Plant Dis. 2021, 105, 1168–1170. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Hu, X.; Liu, C. Isolation and identification of the pathogens causing leaf rot disease of Erigeron breviscapus. Plant Prot. 2006, 32, 92–93. (In Chinese) [Google Scholar]
- Varghese, S.; Jisha, M.S.; Rajeshkumar, K.C.; Gajbhiye, V.; Alrefaei, A.F.; Jeewon, R. Endophytic Fungi: A future prospect for breast cancer therapeutics and drug development. Heliyon 2024, 10, e33995. [Google Scholar] [CrossRef]
- Pal, P.P.; Shaik, A.B.; Begum, A.S. Prospective Leads from Endophytic fungi for Anti-Inflammatory Drug Discovery. Planta Med. 2020, 86, 941–959. [Google Scholar] [CrossRef]
- Yu, C.; Wang, P.; Ding, H.; Hu, Y.; Wang, F.; Chen, H.; Chen, L.; Liu, Y. Effects of the epiphytic patterns on endophytes and metabolites of Dendrobium nobile Lindl. Front. Plant Sci. 2024, 15, 1326998. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, M.-F.; Zhang, Q.; Yan, B.; Jiang, Y.-L. Screening for broad-spectrum antimicrobial endophytes from Rosa roxburghii and multi-omic analyses of biosynthetic capacity. Front. Plant Sci. 2022, 13, 1060478. [Google Scholar] [CrossRef]
- Yao, L.; Wang, J.; He, J.; Huang, L.; Gao, W. Endophytes, biotransforming microorganisms, and engineering microbial factories for triterpenoid saponins production. Crit. Rev. Biotechnol. 2021, 41, 249–272. [Google Scholar] [CrossRef]
- Fan, H.; Lin, P.; Kang, Q.; Zhao, Z.-L.; Wang, J.; Cheng, J.-Y. Metabolism and Pharmacological Mechanisms of Active Ingredients in Erigeron Breviscapus. Curr. Drug Metab. 2021, 22, 24–39. [Google Scholar] [CrossRef]
- Su, S.; Zhao, D.; Yuan, B.; Ma, Y.; Zhu, S.; Xu, K.; Lee, G.; Ho, C.-T.; Huang, Q. Biosynthesis of 6- and 7-mono-demethylated nobiletins by a newly isolated strain of yeast. J. Agric. Food Chem. 2022, 70, 15439–15448. [Google Scholar] [CrossRef]
- He, Y.; Zhan, F.; Xuan, L.; Zu, Y.; Gao, Z.; Li, Y. Selection and flavonoid producing capacity of endophytic fungi isolated from Erigeron breviscapus. J. Agro-Environ. Sci. 2010, 29, 226–229. (In Chinese) [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Df | Sums of Sqs. | Mean Sqs. | F | R2 | p |
|---|---|---|---|---|---|---|
| Tissue | 3 | 1.6996 | 0.5665 | 4.2071 | 0.3869 | 0.002 * |
| season | 1 | 0.8545 | 0.8545 | 5.3128 | 0.1945 | 0.001 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Wu, H.; Wang, F.; Zhao, L.; Gong, W.; Li, H. Diversity, Composition, and Ecological Function of Endophytic Fungal Communities Associated with Erigeron breviscapus in China. Microorganisms 2025, 13, 1080. https://doi.org/10.3390/microorganisms13051080
Zhao Y, Wu H, Wang F, Zhao L, Gong W, Li H. Diversity, Composition, and Ecological Function of Endophytic Fungal Communities Associated with Erigeron breviscapus in China. Microorganisms. 2025; 13(5):1080. https://doi.org/10.3390/microorganisms13051080
Chicago/Turabian StyleZhao, Yi, Hui Wu, Fang Wang, Liangzhou Zhao, Weijun Gong, and Haiyan Li. 2025. "Diversity, Composition, and Ecological Function of Endophytic Fungal Communities Associated with Erigeron breviscapus in China" Microorganisms 13, no. 5: 1080. https://doi.org/10.3390/microorganisms13051080
APA StyleZhao, Y., Wu, H., Wang, F., Zhao, L., Gong, W., & Li, H. (2025). Diversity, Composition, and Ecological Function of Endophytic Fungal Communities Associated with Erigeron breviscapus in China. Microorganisms, 13(5), 1080. https://doi.org/10.3390/microorganisms13051080