Abstract
Ticks are key vectors of zoonotic pathogens, and their expanding distribution in Europe heightens public health concerns. In Bosnia and Herzegovina, while tick distribution is well documented, molecular data on tick-borne pathogens remain limited. This study aimed to illustrate the presence and diversity of these pathogens, focusing on areas with high human activity. Ticks (n = 556) were collected in April 2022 from eight diverse locations, including urban parks, private properties, and rural sites. PCR-based screening was employed to detect Anaplasmataceae, Borrelia, Francisella, Piroplasmida, Rickettsia, and tick-borne encephalitis virus (TBEV), with subsequent sequencing to confirm results. Further characterization of Borrelia burgdorferi sensu lato was achieved via reverse line blotting (RLB) hybridization and sequencing. Ixodes ricinus was the most prevalent species, followed by Dermacentor marginatus and D. reticulatus. Our analysis revealed an overall infection rate of 22.1% in questing ticks, with Rickettsia spp. and Borrelia spp. predominating. Notably, seven Borrelia species were identified in I. ricinus, alongside Anaplasma phagocytophilum, Rickettsia helvetica, and R. monacensis, with co-infections mainly observed in peri-urban areas. This study provides the first molecular evidence of multiple tick-borne pathogens in the region, underscoring the need for further surveillance and risk assessment of tick-borne diseases in the region.
Keywords:
Anaplasma; Balkan; barcoding; Borrelia; Neoehrlichia mikurensis; PCR; reverse line blotting; Rickettsia 1. Introduction
Among all arthropod vectors, ixodid or hard ticks (Acari: Ixodida: Ixodidae) transmit the widest range of pathogens, including protozoa, bacteria, and viruses, posing significant medical and veterinary risks [1]. Hard ticks are widely present in Europe, and climate change is expanding their habitats and thus increasing the risk of regular contact to tick-borne pathogens (TBPs) for both animals and humans [2,3]. The prevalence of TBPs impacting human health has risen significantly over recent decades, reflecting a growing public health challenge [3].
Ixodes ricinus is a three-host tick and the most prevalent tick species throughout the western Palearctic, including Central and (south)eastern Europe. New locations are continuously recorded in Europe [4,5], and endemic areas have reported a notable increase in abundance [6]. Feeding patterns of this species are highly diverse, allowing ticks to feed on various animal species, from small mammals and birds to larger animals, which contributes to the transmission of various pathogens [7,8]. The recent description of Ixodes inopinatus as a distinct species from I. ricinus emphasizes the need for further investigations into its distribution and ecological role. Both species co-exist in overlapping geographical regions, including the Mediterranean basin; however, morphological and molecular identification is challenging [9,10].
The genus Dermacentor comprises 33 species; Dermacentor marginatus and D. reticulatus are the most abundant tick species in the Balkans. Both species quest by latching onto hosts from low vegetation [11]. Dermacentor marginatus commonly targets medium and large mammals including deer, whereas D. reticulatus targets livestock and wildlife, as it is found in open habitats [12,13].
Ixodes ricinus is a highly competent vector for a variety of pathogens, of which the spirochete bacteria Borrelia are the most prevalent. Borrelia burgdorferi s.l. display the most important and the most prevalent tick-borne pathogens in the Northern Hemisphere [14]. Species within the B. burgdorferi s.l. complex, including B. burgdorferi s.s., Borrelia afzelii, and Borrelia garinii, are associated with the progression of Lyme borreliosis (LB) due to their unique pathogenic profiles. Borrelia afzelii is commonly associated with persistent cutaneous symptoms, while B. garinii frequently leads to neurological complications [15,16]. The newly described species B. spielmanii has been identified in a few skin isolates from patients with erythema migrans [17], although its pathogenic potential remains to be fully investigated and confirmed.
Tick-borne encephalitis virus (TBEV) (Flaviviridae) presents a significant public health threat, and infections can cause neurological disorders, including meningitis, encephalitis, and meningoencephalitis [18]. Ixodes ricinus is the key vector for the transmission of the TBE virus across Europe and parts of Central Asia. Among ticks in the genus Dermacentor, the TBEV has also been detected in D. marginatus and D. reticulatus, the latter being a competent vector under experimental conditions [19], expanding its potential transmission range [20].
Other tick-borne bacteria of medical and particularly veterinary relevance are Rickettsia, Anaplasma, and Ehrlichia. The obligate intracellular bacteria of the genus Rickettsia are transmitted by Ixodes and Dermacentor ticks. The latter contributes to the spread of rickettsial pathogens that cause diseases such as spotted fever. Spotted fever group (SFG) Rickettsia spp., namely R. slovaca and R. raoultii, are the causative agents of tick-borne lymphadenopathy (TIBOLA) or Dermacentor-borne necrosis erythema and lymphadenopathy (DEBONEL) [21]. On the other hand, the role of R. monacensis in human disease remains unclear due to its limited pathogenicity despite its occasional detection in human cases [22,23], and the pathogenicity of R. helvetica has been suspected but never proven [24].
The genera Anaplasma, Ehrlichia, and two Neoehrlichia species comprise the intracellular bacteria of the Anaplasmataceae family that are transmitted by ticks to various mammalian hosts, including humans. Species such as Anaplasma phagocytophilum, A. capra, and A. marginale cause anaplasmosis with symptoms including fatigue, fever, and muscle aches; in severe cases, it may lead to complications like immune system dysregulation or reduced platelet levels (thrombocytopenia) [25].
Babesiosis, also known as piroplasmosis, is caused by intraerythrocytic protozoa of the genus Babesia. Symptoms often range from flu-like symptoms such as fever, chills, sweat, and fatigue, with severe cases potentially leading to hemolytic anemia due to red blood cell destruction. The risk of complications is higher in immune-compromised, splenectomised, or elderly individuals [26].
In Balkan countries, data on tick distribution are available, but tick-borne pathogens have been neglected for a long time. Tick surveillance in Bosnia and Herzegovina has demonstrated 19 endemic species [27], of which I. ricinus is the most prevalent species followed by D. marginatus [28]. A few studies, mostly including ticks collected from animals, revealed B. burgdorferi s.l. was predominantly detected in I. ricinus ticks, with detection rates ranging from 2 to 5% depending on the collection site and environmental factors [29]. Similarly, Rickettsia spp., including R. slovaca and R. raoultii, have been detected in Dermacentor and Ixodes ticks, with reported prevalence rates surpassing 10% [30]. Omeragić et al. [29] reported a minimal infection rate of 3.1% for Babesia and 8.8% for A. phagocytophilum in I. ricinus pools. These findings highlight the presence of pathogens in Bosnia and Herzegovina and a dynamic of infection that affects both animals and humans. Although surveys on tick distribution have been conducted, molecular data confirming pathogen presence by PCR and sequencing, particularly from individually analyzed specimens, are scarce but important to assess true prevalence rates. In the current study, we aimed to provide new insights into the prevalence of tick-borne pathogens and analyze sequence data in central and northeastern Bosnia and Herzegovina, particularly at locations with frequent human interaction such as recreational areas.
2. Materials and Methods
2.1. Tick Collection Sites
The survey was conducted at eight different peri-urban and rural collection sites in central and northeastern Bosnia and Herzegovina. Collection sites were chosen based on potential suitability for questing ticks (e.g., uncut grass, meadow/forest intersection) and accessibility (public or private with access granted by owners) in April 2022 (Figure 1). All collection sites were sampled once. The attributes of collection sites are given in Table 1.
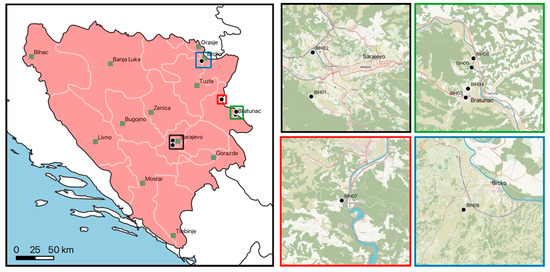
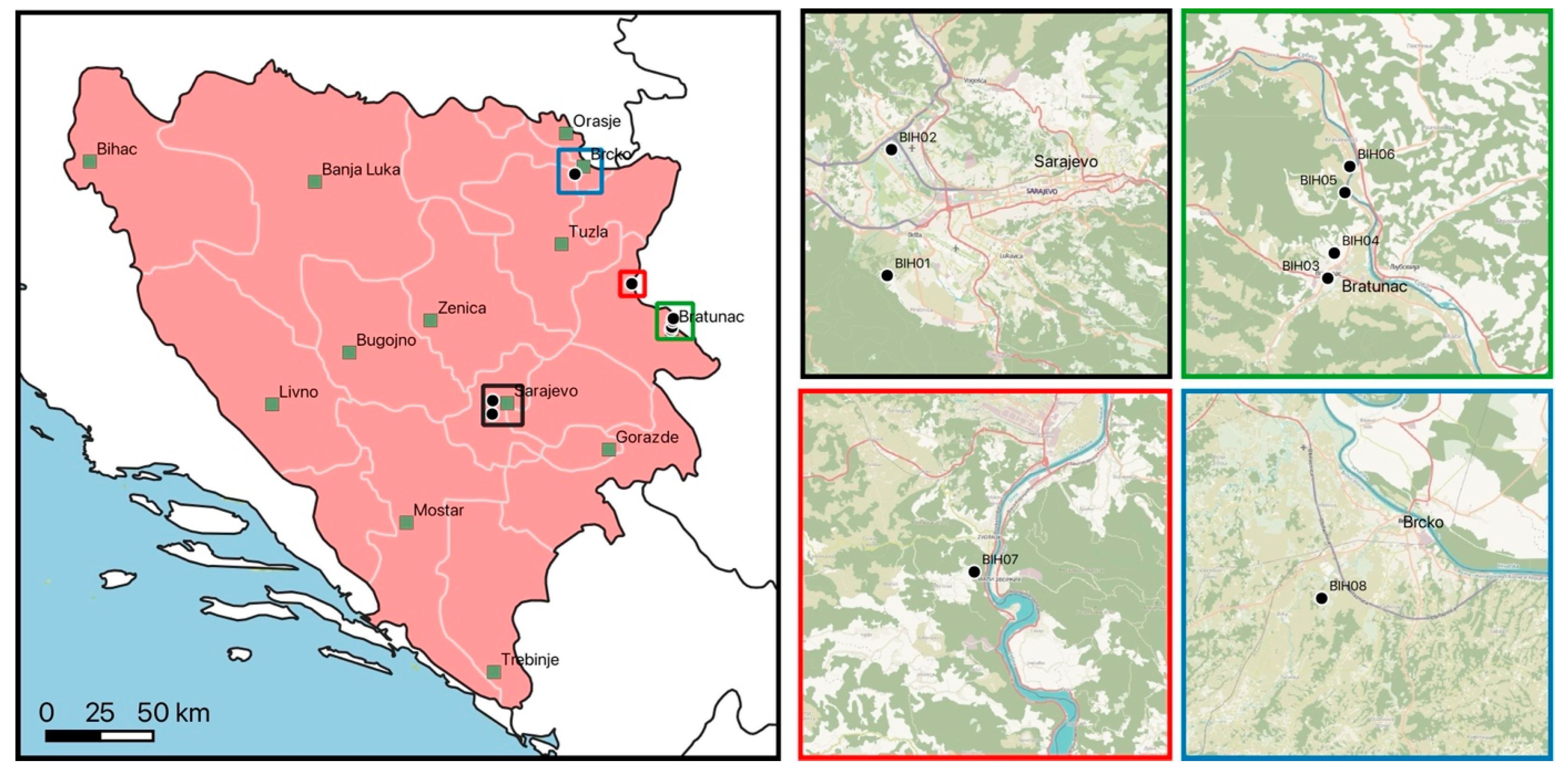
Figure 1.
Collection sites in Bosnia and Herzegovina. Major cities are indicated as black-framed green squares. Colored squares in the main map are displayed as magnified areas on the right. Country borders, first-level administrative divisions, and sea data were included using Natural Earth (www.naturalearthdata.com, accessed on 28 November 2024).

Table 1.
Attributes of tick collection sites.
Questing ticks were collected using a 1 × 1 m white flannel flag, flagging along predefined transects (on average 10 × 50 m2) for an hour. At location BIH1, sheep close by were additionally sampled (with consent of the owners) by carefully removing engorged ticks from animal hosts using tweezers to ensure specimen integrity. Collected ticks were put in 2 mL tubes with screw caps, stored in dry ice before storage in −80 °C.
2.2. Nucleic Acid Extraction
For nucleic acid extraction, ticks were cut longitudinally into two halves. From one half, DNA was isolated using a DNeasy® Blood and Tissue Kit (Qiagen, Hilden, Germany) by incubating in 180 µL ATL buffer and 20 µL proteinase K overnight following the manufacturer’s instructions.
For RNA isolation, halves of ticks were pooled (maximum of six individuals), sorted by location and sex. To each pool, 180 µL phosphate-buffered saline (PBS) was added and crushed with 3 mm stainless steel beads for 5 min at 6000× g with a Qiagen TissueLyser LT (Qiagen, Hilden, Germany). The homogenate was centrifuged at 18,000× g for 5 min, and the supernatant was transferred to a new tube. Consecutively, a Qiagen RNeasy Mini Kit (Qiagen, Hilden, Germany) was used, adding 560 µL of AVL-AVE Lysis buffer (containing RNA carrier) to the homogenate, followed by 350 µL of 70% ethanol, only mixing by pipetting. Subsequent steps were performed based on the manufacturer’s instructions, with a 50 µL final elution volume. The RNA eluate was stored at −20 °C until further use.
2.3. Morphological and Molecular Tick Identification
The morphological identification of ticks was performed under a stereomicroscope using the identification keys from Estrada-Peña et al. [31]. Due to morphological and genetic discrepancies between I. ricinus and I. inopinatus, we refer to I. ricinus sensu lato in this study.
The molecular identification of chosen ticks was based on the amplification of a 16S rRNA gene segment using the primer combination 16S+1 (5′-CTGCTCAATGATTTTTTAAATTGCTGTGG-3′) and 16S-1 (5′-CCGGTCTGAACTCAGATCAAGT-3′) by Black and Piesman [32]. The PCR conditions were 94 °C for 5 min for initial denaturation, followed by 38 cycles of 94 °C for 1 min, 52 °C for 1 min, and 72 °C for 1 min, with a final elongation at 72 °C for 10 min. For all PCR amplifications, the 2× EmeraldAmp® GT PCR Master Mix (Takara Bio Europe, Paris, France) was used with 2 μL template DNA and sterile H2O, adding up to a final volume of 25 μL. Reactions were run on an Eppendorf Mastercycler (Eppendorf AG, Hamburg, Germany). Bands were analyzed with a Gel DocTM XR+ Imager (Bio-Rad Laboratories, Inc., Hercules, CA, USA), cut out and subsequently purified with the IllustraTM GFXTM PCR DNA and Gel Purification Kit (GE Healthcare, Buckinghamshire, UK); then, they were sent to Microsynth Austria GmbH for Sanger sequencing.
Sequences were obtained from both strands, aligned with Clustal X 2.1 [33], and a consensus sequence was generated in GenDoc 2.7.0 [34]. The obtained sequences were uploaded to GenBank and compared to available sequences in the GenBank database using the Basic Local Alignment Search Tool (BLAST) (https://blast.ncbi.nlm.nih.gov/Blast.cgi; accessed on 25 March 2025).
2.4. DNA-Based Pathogen Detection
Samples were screened for the presence of Anaplasmataceae, Borrelia, Francisella, Piroplasmida, and Rickettsia DNA. For the detection of pathogen DNA, published protocols were used (Table 2). Negative controls (H2O) and respective positive controls (DNA at our disposal of Anaplasma marginale originating from bovine blood, Borrelia burgdorferi s.s. B31 strain, Babesia venatorum originating from I. ricinus, Rickettsia raoultii Jongejan strain, and Francisella tularensis subsp. holarctica) were used in each PCR run.
Table 2.
PCR protocols for DNA-based pathogen detection.
2.5. RNA-Based Pathogen Detection
A reverse transcriptase (RT) qPCR with the “universal” flavivirus primer set PF1S (5′-TGYRTBTAYAACATGATGGG-3′) and PF2Rbis (5′-GTGTCCCADCCDGCDGTRTC-3′) and the Luna® Universal One-Step RT-qPCR Kit (New England Biolabs, Ipswich, MA, USA) was used to detect TBEV RNA [46]. For all reactions, 2.5 µL of RNA template was used with the following PCR conditions: 55 °C for 10 min, followed by 45 cycles of 95 °C for 1 min, 95 °C for 10 s, and 50 °C for 1 min. Sterile H2O was used as a negative control, and RNA extracted from a live Yellow fever virus vaccine (Stamaril®, Sanofi, Paris, France) was used as a positive control.
2.6. Borrelia Burgdorferi Sensu Lato Species Discrimination by Reverse Line Blotting (RLB)
All samples positive for B. burgdorferi s.l. by PCR were subjected to PCR-RLB hybridization. A genus-specific RLB-PCR was carried out as reported previously [47,48], using the biotinylated primer pairs shown in Table 1. Each PCR reaction mix with a 25 μL total volume contained the following: 5 μL (5×) of Phire reaction buffer, 200 nmol/L of each dNTP (Solis BioDyne, Tartu, Estonia), 400 nmol/L of each primer per specific primer pair, 0.125 μL Phire Hot Start II DNA Polymerase (Thermo Scientific, Vienna, Austria), PCR-grade water (Sigma-Aldrich, Vienna, Austria), and 2.5 μL of template DNA [49]. The resulting amplicons were analyzed using RLB hybridization as described previously [50].
The further discrimination of B. garinii and B. bavariensis was achieved by amplification and sequencing of the housekeeping gene clpA [41]. A nested PCR was carried out using the primer pairs shown in Table 1. Each PCR reaction was prepared in a total of 25 μL volume containing the following: 5 μL (5×) of Phire reaction buffer, 200 nmol/L of each dNTP (Solis BioDyne, Tartu, Estonia), 400 nmol/L of each primer per specific primer pair (first reaction: clpAF1237 and clpAR2218 and second reaction: clpAF1255 and clpAR2104), 0.5 μL Phire Hot Start II DNA Polymerase (Thermo Scientific, Vienna, Austria), PCR-grade water (Sigma-Aldrich, Vienna, Austria), and lastly 2.5 μL of template DNA. Bands were analyzed with iBright CL750 Imaging System (Thermo Fisher Scientific, Vienna, Austria), cut out, and subsequently purified using QIAquick Gel Extraction kit (Qiagen, Hilden, Germany); then, they were sent to sequencing at Microsynth (Microsynth AG, Vienna, Austria).
2.7. Statistical Analysis
All data were analyzed using Microsoft Excel 16.82 for Mac and R 3.6.2 [51]. Due to the heterogeneity of prevalence rates between locations and the low number of Dermacentor spp., we refrained from further statistical analyses.
3. Results
3.1. Tick Collection
Overall, 556 ticks were collected, comprising three species from eight locations. Of these, 511 (91.9%) were Ixodes ricinus, 42 (7.6%) were Dermacentor marginatus, and 3 (0.5%) were D. reticulatus. Of all, 507 (91.2%) specimens were questing, 16 (2.9%) were unfed from a host, and 33 (5.9%) were engorged females (Table 3). Only I. ricinus and D. marginatus were collected from host animals, which were sheep exclusively at location BIH1 (Stojčevac public park).
Table 3.
Number of collected ticks by species, sex, and developmental stage.
Ixodes ricinus was collected at seven locations, except an open agricultural field (BIH2), and numbers varied considerably between locations. Dermacentor marginatus was sampled at three locations (engorged at BIH1 from sheep close to public park, questing at BIH2 open agricultural field, and BIH8 private property) and D. reticulatus collected at only one location (questing at BIH2 private property) (Table 4).
Table 4.
Sampled tick species by location.
At location BIH1, I. ricinus and D. marginatus were both collected from sheep, and sympatric occurrence of questing D. marginatus and D. reticulatus was observed at location BIH2 (open agricultural field) (Table 4).
3.2. Tick Barcoding
Altogether, 16S rDNA barcoding of 70 specimens was performed. All barcodes allowed for identification to the species level. Haplotyping revealed no genetic pattern associated with geographical origin. Twenty-three I. ricinus sequences with a length of 406 to 408 base pairs (bp) resulted in 12 haplotypes, all showing 99.75% to 100% identity with various I. ricinus sequences originating, e.g., from Poland (MK671578), Slovakia (MN947216), and Portugal (KY039161). Thirty-four barcodes with a length of 414 to 416 bp were obtained from D. marginatus, divided into eight haplotypes, which showed 99.76% to 100% identity with sequences from China (OM368304), Turkey (MT229170), and Spain (MH645513). All three D. reticulatus sequences with a length of 403 bp displaying a single haplotype were 100% identical to reference sequences from Russia (OR936112) and Poland (MK671579) (Table 5).
Table 5.
Generated barcodes, haplotypes, and accession numbers of ticks from BIH.
3.3. DNA-Based Pathogen Screening
Altogether, 112 (112/507; 22.1%) questing ticks, nine (9/16; 56.3%) unfed ticks collected from hosts, and four (4/33; 12.1%) engorged ticks were positive for at least one pathogen. Double infections were detected in questing ticks (11/507; 2.2%) and unfed ticks from hosts (2/16; 12.5%), and triple infections (5/507; 1.0%) were only from questing I. ricinus ticks (Table 6).
Table 6.
Detected pathogens by feeding status.
Of 495 questing Ixodes ricinus specimens, 108 (21.1%) were infected with at least one pathogen, 13 (2.5%) were infected with two pathogens, and 5 (1.0%) with three pathogens (Table 6). Of all 16 engorged female I. ricinus collected from sheep, only one (6.3%) was infected with R. monacensis. The highest single infection rates were observed for Rickettsia (58/511; 11.4%) and Borrelia (39/511; 7.9%), followed by Anaplasma (15/511; 2.9%) and Neoehrlichia mikurensis (1/511; 0.2%) (Table 7).
Table 7.
Single pathogens and co-infections detected in questing I. ricinus ticks.
For Anaplasma, only A. phagocytophilum was detected, with highest rates in female ticks (4.8%). Borrelia comprised B. burgdorferi s.l. (37/511; 7.2%), as well as B. miyamotoi (2/511; 0.4%). One sample (nymph) was positive for Neoehrlichia mikurensis (0.2%), and Rickettsia positive samples were split into R. helvetica (42/511; 8.2%) and R. monacensis (16/511; 3.3%) (Table 7).
Further discrimination of B. burgdorferi s.l. by RLB revealed six species, namely B. afzelii (10/511; 2.0%), B. burgdorferi s.s. (14/511; 2.7%), B. garinii (2/511; 0.4%), B. lusitaniae (18/511; 3.5%), B. spielmanii (2/511; 0.4%), and B. valaisiana (5/511; 1.0%) (Table 7).
The majority of double infections (7/11; 63.6%) detected in I. ricinus comprised two B. burgdorferi s.l. species, as well as three (3/11; 27.2%) B. lusitaniae + R. helvetica infections and one (1/11; 9.0%) B. burgdorferi s.s. + N. mikurensis infection (Table 7). Triple infections involved several B. burgdorferi s.l. species, A. phagocytophilum, and R. monacensis.
Neither Babesia/Theileria or Francisella DNA nor flavivirus RNA were detected in the samples.
In D. marginatus, five (5/16; 31.3%) male specimens collected from hosts were positive for A. ovis and three (3/7; 42.9%) questing females, three (3/17; 17.6%) unengorged females, and four (4/16; 25.0%) males from a host were positive for R. raoultii. One (1/7; 14.3%) questing female and two (2/16; 12.5%) males collected from a host were positive for R. slovaca. Double infections were only detected in males collected from a host, which comprised one (1/16; 6.3%) A. ovis + R. raoultii co-infection and one (1/16; 6.3%) A. ovis + R. slovaca infection.
Of the three D. reticulatus specimens collected questing, one (1/3; 33.3%) was infected with R. raoultii.
As a byproduct of the Anaplasmataceae PCR, Wolbachia spp. and Candidatus Midichloria mitochondrii DNA were detected and sequenced. In 25 of 507 (4.9%) questing I. ricinus, Wolbachia spp. DNA was detected, but only in ticks from location BIH4 (9/187; 4.8%) and BIH5 (16/132; 12.1%). The majority of Wolbachia spp. DNA was detected in nymphs (23/211; 10.9%) and only few females (1/152; 0.7%) and males (1/132; 0.8%). All Wolbachia sp. sequences showed highest similarity with Wolbachia subgroup A. Additionally, in six (6/152; 4.0%) questing female I. ricinus, Candidatus Midichloria mitochondrii DNA was detected.
3.4. Detected Pathogens by Location
While maintaining the same collection effort at all eight locations, the number of ticks and the number of detected pathogens varied considerably between locations. At two locations (BIH3 a private house with small garden and BIH6 along Drina riverbank), no pathogens were detected. At location BIH2 (around agricultural field), only Dermacentor spp. were detected, being infected with R. raoultii and R. slovaca (Supplementary Table S1). The highest diversity of pathogens in questing I. ricinus was detected at location BIH4 (10 pathogens), followed by location BIH5 (eight pathogens) and location BIH1 (seven pathogens), with all three peri-urban collection sites exhibiting the interface of meadows and mixed forests (Figure 2). Also, double and triple infections (BIH1 and BIH4) were only detected in I. ricinus from these three sites (Table S1).
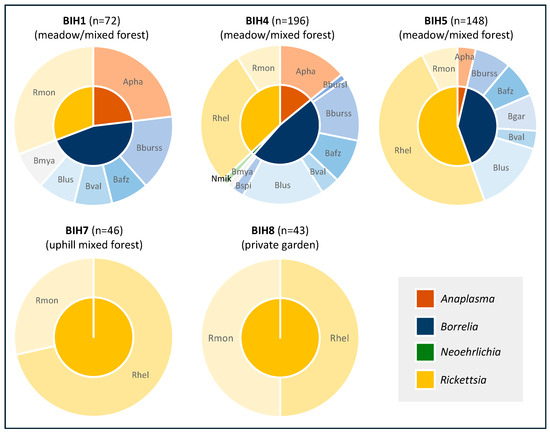
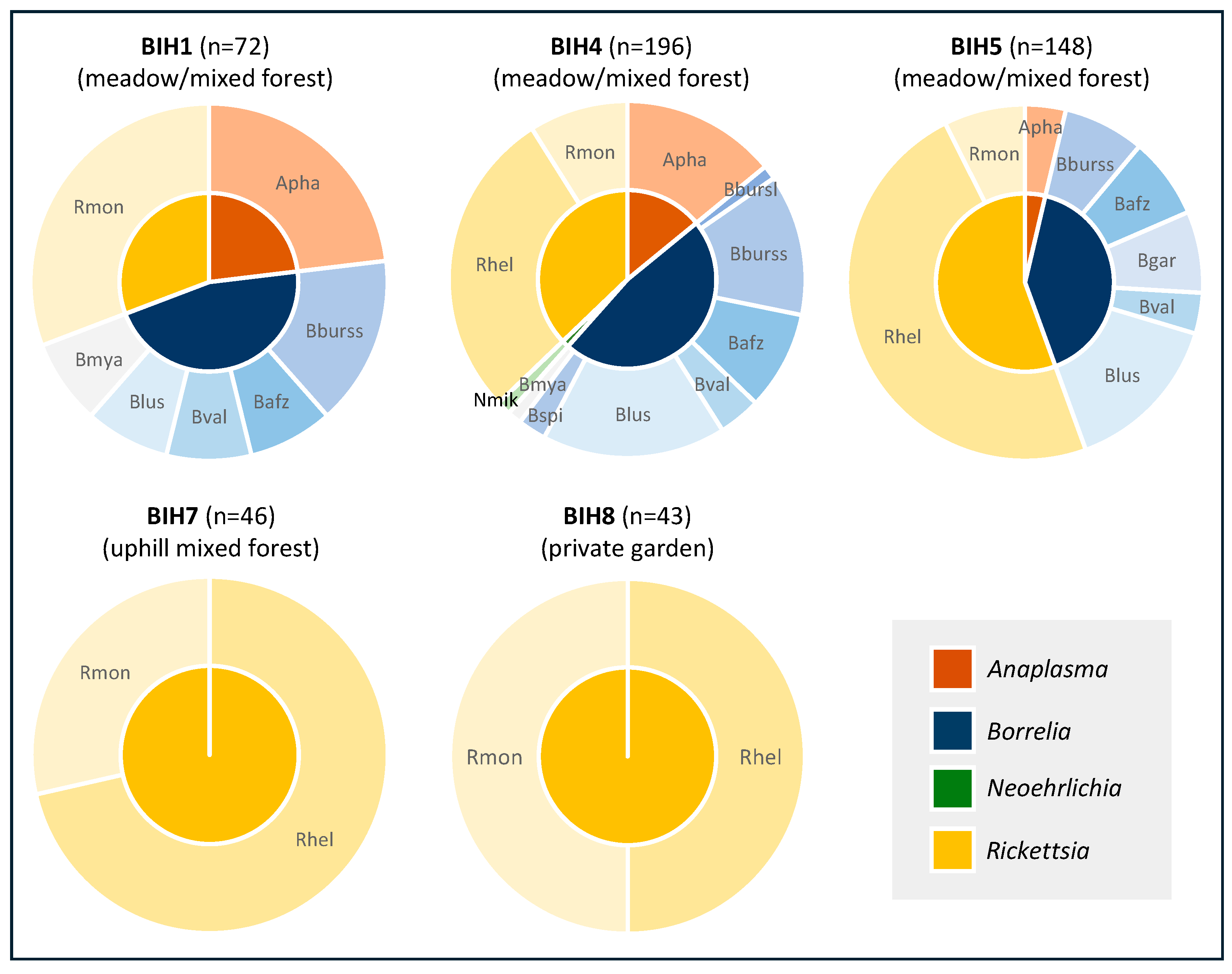
Figure 2.
Pathogen diversity in questing I. ricinus by sampling location. Apha, Anaplasma phagocytophilum; Nmik, Neoehrlichia mikurensis; Bbursl, Borrelia burgdorferi s.l.; Bburss, B. burgdorferi s.s.; Bafz, B. afzeli; Bgar, B. garinii; Blus, B. lusitaniae; Bval, B. valaisiana; Bspi, B. spielmanii; Bmya, B. myamotoi; Rslo, Rickettsia slovaca; Rhel, R. helvetica; Rmon, R. monacensis.
3.5. Pathogen Typing
Sequences of pathogens were uploaded to GenBank and compared to available reference sequences. All 14 A. phagocytophilum sequences comprised a single haplotype, showing 100% identity with reference sequences originating from ticks (I. ricinus, MW922753; I. scapularis, HG916766; or R. sanguineus sensu lato, OR976127) and animals such as dogs (MK814412), horses (MK811374), or rats (OL690564). For A. ovis, the confirmatory PCR based on the major surface protein 4 (msp4) gene showed 100% identity with isolates from sheep in Pakistan (PQ616034) or China (MH908943) (Table 8).
Table 8.
Generated sequences, haplotypes, and accession numbers of detected pathogens.
For Borrelia, only B. myamotoi was subjected to sequence analysis, revealing 100% identity with reference sequences originating from I. ricinus in Czech Republic (KJ847049) and Poland (KF422749). Typing of B. burgdorferi s.l. species was accomplished by RLB only due to low specificity of the applied PCR.
A single sequence of N. mikurensis showed 99.26% identity to Candidatus Neoehrlichia sp. (OP269946, OP269947) and 98.9% identity to the sequence of the Ca. Neoehrlichia mikurensis reference genome (CP066557).
For Rickettsia, six R. monacensis sequences comprising two haplotypes were 99.71% to 100% identical to a strain from Poland (JQ796867) and the type strain (LN794217). Four R. helvetica sequences comprising two haplotypes showed 99.8% to 100% identity to reference sequences from Poland (JQ796866) and Austria (EU057990). Five R. raoultii sequences consisting of a single haplotype showed 100% identity to the “Khabarovsk” strain (CP010969), and three R. slovaca sequences (single haplotype) were 100% identical to strains from Pakistan (MN581971) and Portugal (AY125009) (Table 8).
4. Discussion
Our study provides a snapshot and a valuable cross-section through tick-borne pathogens in local tick populations from central and northeastern Bosnia and Herzegovina, thereby demonstrating a high diversity of pathogens. Ixodes ricinus, the predominant tick species in the area, showed an infection rate of more than 20% with at least one pathogen, including co-infections (2.5%) and triple infections (1.0%). We provide the first molecular data on Borrelia genotyping in the region, identifying six distinct B. burgdorferi sensu lato genospecies in addition to B. myamotoi.
4.1. Pathogens in Questing Ticks
Previous studies in Bosnia and Herzegovina have confirmed the presence of B. burgdorferi s.l.; however, the identification of specific genospecies has been limited. Omeragić et al. [29] conducted molecular screening of tick-borne pathogens in I. ricinus ticks, detecting B. burgdorferi s.l. in 3.4% of specimens exclusively in those collected from domestic animals. On the contrary, our study is the first to report the presence of six distinct species within the B. burgdorferi complex in this region, namely B. afzelii, B. garinii, B. burgdorferi s.s., B. lusitaniae, B. spielmanii, B. valaisiana, and additionally B. miyamotoi, a relapsing fever spirochete considered an emerging human pathogen [52]. Among the genospecies that infect humans, B. burgdorferi s.s., B. afzelii, and B. garinii are the primary agents of Lyme borreliosis in Europe [53,54]. Recently, Lasić et al. [55] detected B. burgdorferi s.l. in I. ricinus ticks collected from patients in Sarajevo, the capital of Bosnia and Herzegovina, and recent data from the Institute of Public Health [56] indicate a rising trend in reported Lyme borreliosis cases.
In Bosnia and Herzegovina, previous studies have identified the presence of Rickettsia species in ticks. For instance, Omeragić et al. [29] detected Rickettsia spp. in ticks collected from cats, sheep, goats, and dogs, while all ticks collected from vegetation tested negative. In contrast, our study found the highest Rickettsia spp. positivity rates in ticks sampled by flagging, with R. helvetica (8.2%) as the most prevalent, followed by R. monacensis (3.3%). This is in line with Hodžić et al. [57], who reported R. helvetica (5.7%) and R. monacensis (1.1%) in questing I. ricinus in the region. The molecular detection of Rickettsia spp. was reported in patients with a history of tick bites who sought medical care, confirming the presence of R. helvetica, R. monacensis, and R. felis, highlighting their potential pathogenicity in humans [58]. Boretti et al. [59] assessed the public health significance of R. helvetica, detecting the pathogen in dogs, foxes, humans, and Ixodes ticks. Rickettsia helvetica infections in humans manifest as a non-specific febrile illness and in more severe cases, R. helvetica-associated meningitis has been reported [60,61]. On the other hand, R. monacensis infections can present with fever, rash, or headache, though their full clinical spectrum remains incompletely understood [62].
DNA of A. phagocytophilum was detected in 3% of questing I. ricinus, contrary to a recent study by Omeragić et al. [29], who reported absence in pools of questing adult I. ricinus. However, the presence of A. phagocytophilum DNA was recently confirmed in various animal hosts, as well as in ticks collected from dogs across all regions of BIH, with infections ranging from 0.9 to 23.8% [29]. As an emerging tick-borne pathogen, A. phagocytophilum represents a significant veterinary and public health concern [63]. The low A. phagocytophilum infection rates observed in this study are in line with previous findings in Bosnia and Herzegovina [57] and more broadly across Eastern Europe. Although there are no officially reported human cases of A. phagocytophilum infections, the presence of Anaplasma spp. has been confirmed in blood samples collected from stray dogs in Bosnia and Herzegovina [64], which highlights the need for further surveillance and awareness of this emerging pathogen, particularly given its zoonotic potential [63] and the role of domestic animals as potential reservoirs. Neoehrlichia mikurensis was detected in only a single I. ricinus tick, highlighting its rare occurrence in the sampled tick population. This emerging tick-borne pathogen is of clinical relevance, particularly in immunocompromised individuals [65].
We report the detection of Wolbachia spp. and Candidatus Midichloria mitochondrii as a rather incidental finding while screening for TBPs of the genera Anaplasma and Ehrlichia, suggesting that the true prevalence of these symbionts might be higher if specific PCR assays were employed. Wolbachia subgroup A in ticks is typically associated with insect endoparasitoids like Ixodiphagus hookeri, which parasitize ticks, leading to incidental detection in tick samples [66]. Midichloria species are known endosymbionts of ticks, residing in ovarian tissues and occasionally in salivary glands, with potential implications for tick biology and pathogen transmission [67].
4.2. Co-Infections
Double and triple co-infections with Borrelia spp. or other tick-borne pathogens are more common in I. ricinus due to its diverse host range, and they may exacerbate disease severity in infected individuals [68,69,70]. In the present study, Borrelia-infected ticks were exclusively collected from peri-urban locations with frequent human presence, aligning with the findings on I. ricinus ticks collected from peri-urban locations in Kosovo [71]. The proximity to forested areas, the presence of wildlife species, and the presence of stray dogs likely contribute to the maintenance and transmission of Borrelia spp. and other tick-borne pathogens [72].
4.3. Pathogen Absence
Interestingly, no Babesia spp. DNA was detected in the analyzed tick samples, aligning with previous studies that reported low prevalence or sporadic detection in the region [73]. While I. ricinus is a known vector of Babesia spp., with human infections reported in Europe, its circulation in Bosnia and Herzegovina remains poorly characterized. Similarly, no evidence of tick-borne encephalitis virus (TBEV) was found, which may be attributed not only to inactive TBEV foci and a lower risk of transmission in the sampled areas but also to the relatively low number of ticks examined, given that TBEV infection rates in ticks are typically < 1:1000. However, considering the particular focal distribution of the TBEV and the endemic presence in neighboring countries, continued surveillance is crucial for assessing potential spillover risks [74,75].
4.4. Sympatric Occurrence of Dermacentor spp.
We observed sympatric occurrences of both D. marginatus and D. reticulatus species questing at the same locations, a phenomenon that has been documented in other regions. For instance, Drehmann et al. [76] reported overlapping distributions of these species across Germany, suggesting potential ecological interactions and shared habitats. While previous research confirms the presence of both Dermacentor species, Omeragić et al. [28] also noted that the previously registered abundance of D. marginatus had nearly doubled. In our study, A. ovis was detected in 31.3% of male D. marginatus ticks from hosts, while R. raoultii and R. slovaca were identified in both questing and host-collected ticks.
4.5. Differences in Pathogen Diversity
The absence or low diversity of tick-borne pathogens in rural locations may be attributed to limited sample sizes, reducing the likelihood of encountering infected ticks. However, pathogen prevalence is not solely dependent on sample size; host-related factors, including the availability, density, and reservoir competence of the main tick hosts also play a crucial role in pathogen circulation [77,78]. For instance, despite similar trapping effort, tick density and pathogen prevalence varied between the peri-urban and rural settings, as the difference in habitat supports distinct wildlife reservoirs and host communities, which affect tick feeding and infection cycles [72,79]. Low pathogen detection in certain locations may be attributed to the absence of key reservoir hosts, reducing pathogen circulation in local tick populations [6]. Areas with fragmented landscapes and lower host densities have been linked to decreased infection rates in ticks [80]. Additionally, climatic factors such as temperature, precipitation, and humidity play a crucial role in tick survival and pathogen transmission. For example, one site with few collected specimens and no pathogen detection in this study was located along the Drina River, where periodic flooding may disrupt tick activity and reduce pathogen persistence in the environment [81,82].
5. Conclusions
Despite sampling only once per location within a single year and the collection sites being heterogeneous, potentially influencing pathogen detection rates, our findings underscore the veterinary and medical significance of tick-borne pathogens in Bosnia and Herzegovina, particularly Borrelia species (including cases of co-infection), A. phagocytophilum, N. mikurensis, and Rickettsia spp., all of which were detected in our study. Continuous surveillance is essential, especially in recreational areas, where human–tick encounters are frequent, to monitor the dynamics of TBP circulation. Future research should aim for longitudinal sampling over multiple seasons across diverse habitats to better understand the dynamics of tick-borne diseases and their implications for public health and veterinary medicine.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms13051054/s1, Table S1: Detected pathogens by sampling location.
Author Contributions
Conceptualization, I.H., J.W., E.K. and A.A.; data curation, I.H., M.R., M.W., (Michiel Wijnveld) and E.K.; formal analysis, I.H., E.K. and A.A.; funding acquisition, A.G.O., M.W., (Martin Weiler) and E.K.; methodology, I.H., J.D., M.R., M.W. (Michiel Wijnveld), B.J., E.K. and A.A.; project administration, A.G.O., M.W. (Martin Weiler) and E.K.; writing—original draft, I.H. and E.K.; writing—review and editing, I.H., J.D., M.R., M.W. (Michiel Wijnveld), A.G.O., B.J., M.W. (Martin Weiler), J.W., E.K. and A.A. All authors have read and agreed to the published version of the manuscript.
Funding
The study has been funded by the Austrian defense research program FORTE of the Federal Ministry of Finance (BMF) (project no. 886318) and in part by the Austrian Science Fund (FWF) (grant number: P33867).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated and analyzed during this study are included in the article and Supplementary Materials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, 3–14. [Google Scholar] [CrossRef]
- Gray, J.S. The ecology of ticks transmitting Lyme borreliosis. Exp. Appl. Acarol. 1998, 22, 249–258. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; De La Fuente, J. The ecology of ticks and epidemiology of tick-borne viral diseases. Antivir. Res. 2014, 108, 104–128. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A. Forecasting habitat suitability for ticks and prevention of tick-borne diseases. Vet. Parasitol. 2001, 98, 111–132. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Farkas, R.; Jaenson, T.G.T.; Koenen, F.; Madder, M.; Pascucci, I.; Salman, M.; Tarrés-Call, J.; Jongejan, F. Association of environmental traits with the geographic ranges of ticks (Acari: Ixodidae) of medical and veterinary importance in the western Palearctic. A digital data set. Exp. Appl. Acarol. 2012, 59, 351. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.-C.; Golovljova, I.; Jaenson, T.G.T.; Jensen, J.-K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasit. Vectors 2013, 6, 1. [Google Scholar] [CrossRef]
- Estrada-Peña, A. Ticks as vectors: Taxonomy, biology and ecology. Rev. Sci. Tech. 2015, 34, 53–65. [Google Scholar] [CrossRef]
- Sonenshine, D.E.; Roe, R.M. (Eds.) Biology of Ticks; Oxford University Press: Oxford, UK, 2014; Volume 2, ISBN 9780199744053. [Google Scholar]
- Estrada-Peña, A.; Nava, S.; Petney, T. Description of all the stages of Ixodes inopinatus n. sp. (Acari: Ixodidae). Ticks Tick-Borne Dis. 2014, 5, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Chitimia-Dobler, L.; Rieß, R.; Kahl, O.; Wölfel, S.; Dobler, G.; Nava, S.; Estrada-Peña, A. Ixodes inopinatus—Occurring also outside the Mediterranean region. Ticks Tick-Borne Dis. 2018, 9, 196–200. [Google Scholar] [CrossRef]
- Zając, Z.; Bartosik, K.; Woźniak, A. Monitoring Dermacentor reticulatus Host-Seeking Activity in Natural Conditions. Insects 2020, 11, 264. [Google Scholar] [CrossRef]
- Bilbija, B.; Spitzweg, C.; Papoušek, I.; Fritz, U.; Földvári, G.; Mullett, M.; Ihlow, F.; Sprong, H.; Civáňová Křížová, K.; Anisimov, N.; et al. Dermacentor reticulatus—A tick on its way from glacial refugia to a panmictic Eurasian population. Int. J. Parasitol. 2023, 53, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Rubel, F.; Brugger, K.; Pfeffer, M.; Chitimia-Dobler, L.; Didyk, Y.M.; Leverenz, S.; Dautel, H.; Kahl, O. Geographical distribution of Dermacentor marginatus and Dermacentor reticulatus in Europe. Ticks Tick-Borne Dis. 2016, 7, 224–233. [Google Scholar] [CrossRef]
- Stanek, G.; Reiter, M. The expanding Lyme Borrelia complex-clinical significance of genomic species? Clin. Microbiol. Infect. 2011, 17, 487–493. [Google Scholar] [CrossRef]
- Rauer, S.; Kastenbauer, S.; Hofmann, H.; Fingerle, V.; Huppertz, H.I.; Hunfeld, K.P.; Krause, A.; Ruf, B.; Dersch, R. Guidelines for diagnosis and treatment in neurology—Lyme neuroborreliosis. Ger. Med. Sci. 2020, 18, Doc03. [Google Scholar] [CrossRef]
- Steinbrink, A.; Brugger, K.; Margos, G.; Kraiczy, P.; Klimpel, S. The evolving story of Borrelia burgdorferi sensu lato transmission in Europe. Parasitol. Res. 2022, 121, 781–803. [Google Scholar] [CrossRef]
- Fingerle, V.; Schulte-Spechtel, U.C.; Ruzic-Sabljic, E.; Leonhard, S.; Hofmann, H.; Weber, K.; Pfister, K.; Strle, F.; Wilske, B. Epidemiological aspects and molecular characterization of Borrelia burgdorferi s.l. from southern Germany with special respect to the new species Borrelia spielmanii sp. nov. Int. J. Med. Microbiol. 2008, 298, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, L.; Vapalahti, O. Tick-borne encephalitis. Lancet 2008, 371, 1861–1871. [Google Scholar] [CrossRef]
- Ličková, M.; Fumačová Havlíková, S.; Sláviková, M.; Slovák, M.; Drexler, J.F.; Klempa, B. Dermacentor reticulatus is a vector of tick-borne encephalitis virus. Ticks Tick-Borne Dis. 2020, 11, 101414. [Google Scholar] [CrossRef] [PubMed]
- Chitimia-Dobler, L. TBEV-transmission and natural cycles. In The TBE Book; Dobler, G., Erber, W., Bröker, M., Chitimia-Dobler, L., Schmitt, H.J., Eds.; Global Health Press: Singapore, 2024; Chapter 5. [Google Scholar]
- Parola, P.; Rovery, C.; Rolain, J.M.; Brouqui, P.; Davoust, B.; Raoult, D. Rickettsia slovaca and R. raoultii in Tick-borne Rickettsioses. Emerg. Infect. Dis. 2009, 15, 1105–1108. [Google Scholar] [CrossRef]
- Jado, I.; Oteo, J.A.; Aldámiz, M.; Gil, H.; Escudero, R.; Ibarra, V.; Portu, J.; Portillo, A.; Lezaun, M.J.; García-Amil, C.; et al. Rickettsia monacensis and Human Disease, Spain. Emerg. Infect. Dis. 2007, 13, 1405–1407. [Google Scholar] [CrossRef]
- de Sousa, R.; Pereira, B.I.; Nazareth, C.; Cabral, S.; Ventura, C.; Crespo, P.; Marques, N.; da Cunha, S. Rickettsia slovaca Infection in Humans, Portugal. Emerg. Infect. Dis. 2013, 19, 1627. [Google Scholar] [CrossRef] [PubMed]
- Brouqui, P.; Parola, P.; Fournier, P.E.; Raoult, D. Spotted fever rickettsioses in southern and eastern Europe. FEMS Immunol. Med. Microbiol. 2007, 49, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Rar, V.; Golovljova, I. Anaplasma, Ehrlichia, and “Candidatus Neoehrlichia” bacteria: Pathogenicity, biodiversity, and molecular genetic characteristics, a review. Infect. Genet. Evol. 2011, 11, 1842–1861. [Google Scholar] [CrossRef] [PubMed]
- Vannier, E.; Krause, P.J. Human babesiosis. N. Engl. J. Med. 2012, 366, 2397–2407. [Google Scholar] [CrossRef]
- Kapo, N.; Zuber Bogdanović, I.; Gagović, E.; Žekić, M.; Veinović, G.; Sukara, R.; Mihaljica, D.; Adžić, B.; Kadriaj, P.; Cvetkovikj, A.; et al. Ixodid ticks and zoonotic tick-borne pathogens of the Western Balkans. Parasit. Vectors 2024, 17, 45. [Google Scholar] [CrossRef]
- Omeragić, J.; Šerić-Haračić, S.; Klarić Soldo, D.; Kapo, N.; Fejzić, N.; Škapur, V.; Medlock, J. Distribution of ticks in Bosnia and Herzegovina. Ticks Tick-Borne Dis. 2022, 13, 101870. [Google Scholar] [CrossRef]
- Omeragić, J.; Kapo, N.; Goletić, Š.; Softić, A.; Terzić, I.; Šabić, E.; Škapur, V.; Klarić Soldo, D.; Goletić, T. Investigation of Tick-Borne Pathogens in Ixodes Ticks from Bosnia and Herzegovina. Animals 2024, 14, 2190. [Google Scholar] [CrossRef]
- Goletić, T.; Klarić Soldo, D.; Kapo, N.; Goletić, Š.; Koro-Spahić, A.; Alispahić, A.; Softić, A.; Škapur, V.; Omeragić, J. Tick-Borne Pathogens in Dermacentor reticulatus Ticks from Bosnia and Herzegovina. Pathogens 2024, 13, 421. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Mihalca, A.D.; Petney, T.N. Ticks of Europe and North Africa: A Guide to Species Identification; Springer: Berlin, Germany, 2018. [Google Scholar]
- Black, W.C.; Piesman, J. Phylogeny of hard- and soft-tick taxa (Acari: Ixodida) based on mitochondrial 16S rDNA sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 10034–10038. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Nicholas, K.B. Genedoc: A Tool for Editing and Annoting Multiple Sequence Alignments. 1997. Available online: https://nrbsc.org/gfx/genedoc/ (accessed on 5 January 2025).
- Schouls, L.M.; Van De Pol, I.; Rijpkema, S.G.T.; Schot, C.S. Detection and Identification of Ehrlichia, Borrelia burgdorferi Sensu Lato, and Bartonella Species in Dutch Ixodes ricinus Ticks. J. Clin. Microbiol. 1999, 37, 2215–2222. [Google Scholar] [CrossRef]
- Bekker, C.P.; de Vos, S.; Taoufik, A.; Sparagano, O.A.; Jongejan, F. Simultaneous detection of Anaplasma and Ehrlichia species in ruminants and detection of Ehrlichia ruminantium in Amblyomma variegatum ticks by reverse line blot hybridization. Vet. Microbiol. 2002, 89, 223–238. [Google Scholar] [CrossRef]
- Yang, J.; Liu, Z.; Niu, Q.; Liu, J.; Han, R.; Liu, G.; Shi, Y.; Luo, J.; Yin, H. Molecular survey and characterization of a novel Anaplasma species closely related to Anaplasma capra in ticks, northwestern China. Parasit. Vectors 2016, 9, 603. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Zhao, S.; Wang, K.; Song, J.; Yan, Y.; Zhou, Y.; Shi, K.; Jian, F.; Wang, R.; Zhang, L.; et al. A Multiplex PCR Detection Assay for the Identification of Clinically Relevant Anaplasma Species in Field Blood Samples. Front. Microbiol. 2020, 11, 511546. [Google Scholar] [CrossRef] [PubMed]
- Torina, A.; Agnone, A.; Blanda, V.; Alongi, A.; D’Agostino, R.; Caracappa, S.; Marino, A.M.F.; Di Marco, V.; de la Fuente, J. Development and validation of two PCR tests for the detection of and differentiation between Anaplasma ovis and Anaplasma marginale. Ticks Tick-Borne Dis. 2012, 3, 283–287. [Google Scholar] [CrossRef]
- Liebisch, G.; Sohns, B.; Bautsch, W. Detection and typing of Borrelia burgdorferi sensu lato in Ixodes ricinus ticks attached to human skin by PCR. J. Clin. Microbiol. 1998, 36, 3355–3358. [Google Scholar] [CrossRef]
- Margos, G.; Gatewood, A.G.; Aanensen, D.M.; Hanincová, K.; Terekhova, D.; Vollmer, S.A.; Cornet, M.; Piesman, J.; Donaghy, M.; Bormane, A.; et al. MLST of housekeeping genes captures geographic population structure and suggests a European origin of Borrelia burgdorferi. Proc. Natl. Acad. Sci. USA 2008, 105, 8730–8735. [Google Scholar] [CrossRef] [PubMed]
- Sjöstedt, A.; Eriksson, U.; Berglund, L.; Tärnvik, A. Detection of Francisella tularensis in ulcers of patients with tularemia by PCR. J. Clin. Microbiol. 1997, 35, 1045–1048. [Google Scholar] [CrossRef]
- Bonnet, S.; Jouglin, M.; L’Hostis, M.; Chauvin, A. Babesia sp. EU1 from Roe Deer and Transmission within Ixodes ricinus. Emerg. Infect. Dis. 2007, 13, 1208. [Google Scholar] [CrossRef]
- Zintl, A.; Finnerty, E.J.; Murphy, T.M.; de Waal, T.; Gray, J.S. Babesias of red deer (Cervus elaphus) in Ireland. Vet. Res. 2011, 42, 7. [Google Scholar] [CrossRef]
- Vitorino, L.; Zé-Zé, L.; Sousa, A.; Bacellar, F.; Tenreiro, R. rRNA Intergenic Spacer Regions for Phylogenetic Analysis of Rickettsia Species. Ann. N. Y. Acad. Sci. 2003, 990, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Moureau, G.; Temmam, S.; Gonzalez, J.P.; Charrel, R.N.; Grard, G.; De Lamballerie, X. A real-time RT-PCR method for the universal detection and identification of flaviviruses. Vector-Borne Zoonotic Dis. 2007, 7, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Schötta, A.-M.; Wijnveld, M.; Stockinger, H.; Stanek, G. Approaches for Reverse Line Blot-Based Detection of Microbial Pathogens in Ixodes ricinus Ticks Collected in Austria and Impact of the Chosen Method. Appl. Environ. Microbiol. 2017, 83, e00489-17. [Google Scholar] [CrossRef] [PubMed]
- Wijnveld, M.; Schötta, A.-M.; Pintér, A.; Stockinger, H.; Stanek, G. Novel Rickettsia raoultii strain isolated and propagated from Austrian Dermacentor reticulatus ticks. Parasit. Vectors 2016, 9, 567. [Google Scholar] [CrossRef]
- Wijnveld, M.; Schötta, A.M.; Stelzer, T.; Duscher, G.; Leschnik, M.; Stockinger, H.; Lindgren, P.E.; Stanek, G. Novel Protozoans in Austria Revealed through the Use of Dogs as Sentinels for Ticks and Tick-Borne Pathogens. Microorganisms 2021, 9, 1392. [Google Scholar] [CrossRef]
- Gubbels, J.M.; De Vos, A.P.; Van Der Weide, M.; Viseras, J.; Schouls, L.M.; De Vries, E.; Jongejan, F. Simultaneous detection of bovine Theileria and Babesia species by reverse line blot hybridization. J. Clin. Microbiol. 1999, 37, 1782–1789. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org (accessed on 5 January 2025).
- Krause, P.J.; Fish, D.; Narasimhan, S.; Barbour, A.G. Borrelia miyamotoi infection in nature and in humans. Clin. Microbiol. Infect. 2015, 21, 631–639. [Google Scholar] [CrossRef]
- Hubálek, Z. Epidemiology of Lyme Borreliosis. In Lyme Borreliosis; Lipsker, D., Jaulhac, B., Eds.; Karger: Basel, Switzerland, 2009; Volume 37, pp. 31–50. [Google Scholar]
- Rudenko, N.; Golovchenko, M.; Grubhoffer, L.; Oliver, J.H. Updates on Borrelia burgdorferi sensu lato complex with respect to public health. Ticks Tick-Borne Dis. 2011, 2, 123–128. [Google Scholar] [CrossRef]
- Lasić, L.; Ušanović, L.; Ćakić, S.; Hanjalić, J.; Stroil, B.K. First molecular detection of Borrelia burgdorferi sensu lato in Ixodes ricinus ticks collected from humans in the Sarajevo Canton (Bosnia and Herzegovina). Syst. Appl. Acarol. 2020, 25, 169–172. [Google Scholar] [CrossRef]
- World Health Organization. The Institute of Public Health of the Federation of Bosnia and Herzegovina. 2025. Available online: https://www.phi.rs.ba/ (accessed on 19 February 2025).
- Hodžić, A.; Fuehrer, H.P.; Duscher, G.G. First Molecular Evidence of Zoonotic Bacteria in Ticks in Bosnia and Herzegovina. Transbound. Emerg. Dis. 2017, 64, 1313–1316. [Google Scholar] [CrossRef]
- Banović, P.; Díaz-Sánchez, A.A.; Foucault-Simonin, A.; Mateos-Hernandez, L.; Wu-Chuang, A.; Galon, C.; Simin, V.; Mijatović, D.; Bogdan, I.; Corona-González, B.; et al. Emerging tick-borne spotted fever group rickettsioses in the Balkans. Infect. Genet. Evol. 2023, 107, 105400. [Google Scholar] [CrossRef] [PubMed]
- Boretti, F.S.; Perreten, A.; Meli, M.L.; Cattori, V.; Willi, B.; Wengi, N.; Hornok, S.; Honegger, H.; Hegglin, D.; Woelfel, R.; et al. Molecular Investigations of Rickettsia helvetica Infection in Dogs, Foxes, Humans, and Ixodes Ticks. Appl. Environ. Microbiol. 2009, 75, 3230–3237. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Davoust, B.; Raoult, D. Tick- and flea-borne rickettsial emerging zoonoses. Vet. Res. 2005, 36, 469–492. [Google Scholar] [CrossRef]
- Walker, D.H. Rickettsiae and Rickettsial Infections: The Current State of Knowledge. Clin. Infect. Dis. 2007, 45, S39–S44. [Google Scholar] [CrossRef] [PubMed]
- Biernat, B.; Stańczak, J.; Michalik, J.; Sikora, B.; Cieniuch, S. Rickettsia helvetica and R. monacensis infections in immature Ixodes ricinus ticks derived from sylvatic passerine birds in west-central Poland. Parasitol. Res. 2016, 115, 3469–3477. [Google Scholar] [CrossRef]
- Atif, F.A. Anaplasma marginale and Anaplasma phagocytophilum: Rickettsiales pathogens of veterinary and public health significance. Parasitol. Res. 2015, 114, 3941–3957. [Google Scholar] [CrossRef]
- Maksimović, Z.; Dervišević, M.; Zahirović, A.; Rifatbegović, M. Seroprevalence of Anaplasma spp. and Ehrlichia spp. and molecular detection of Anaplasma phagocytophilum and Anaplasma platys in stray dogs in Bosnia and Herzegovina. Ticks Tick. Borne. Dis. 2022, 13, 101875. [Google Scholar] [CrossRef]
- Jahfari, S.; Fonville, M.; Hengeveld, P.; Reusken, C.; Scholte, E.-J.; Takken, W.; Heyman, P.; Medlock, J.M.; Heylen, D.; Kleve, J.; et al. Prevalence of Neoehrlichia mikurensis in ticks and rodents from North-west Europe. Parasit. Vectors 2012, 5, 74. [Google Scholar] [CrossRef]
- Plantard, O.; Bouju-Albert, A.; Malard, M.A.; Hermouet, A.; Capron, G.; Verheyden, H. Detection of Wolbachia in the Tick Ixodes ricinus is Due to the Presence of the Hymenoptera Endoparasitoid Ixodiphagus hookeri. PLoS One 2012, 7, e30692. [Google Scholar] [CrossRef]
- Bonnet, S.I.; Binetruy, F.; Hernández-Jarguín, A.M.; Duron, O. The Tick Microbiome: Why Non-pathogenic Microorganisms Matter in Tick Biology and Pathogen Transmission. Front. Cell. Infect. Microbiol. 2017, 7, 271096. [Google Scholar] [CrossRef]
- Grunwaldt, E.; Barbour, A.; Benach, J. High Potassium in Low-Sodium Soups. N. Engl. J. Med. 1983, 308, 1166–1167. [Google Scholar] [CrossRef] [PubMed]
- Raileanu, C.; Moutailler, S.; Pavel, I.; Porea, D.; Mihalca, A.D.; Savuta, G.; Vayssier-Taussat, M. Borrelia Diversity and Co-infection with Other Tick Borne Pathogens in Ticks. Front. Cell. Infect. Microbiol. 2017, 7, 244580. [Google Scholar] [CrossRef] [PubMed]
- Moutailler, S.; Valiente Moro, C.; Vaumourin, E.; Michelet, L.; Tran, F.H.; Devillers, E.; Cosson, J.F.; Gasqui, P.; Van, V.T.; Mavingui, P.; et al. Co-infection of Ticks: The Rule Rather Than the Exception. PLoS Negl. Trop. Dis. 2016, 10, e0004539. [Google Scholar] [CrossRef] [PubMed]
- Hoxha, I.; Xhekaj, B.; Halimi, G.; Wijnveld, M.; Ruivo, M.; Çaushi, D.; Matoshi, A.; Obwaller, A.G.; Jäger, B.; Weiler, M.; et al. Zoonotic Tick-Borne Pathogens in Ixodes ricinus Complex (Acari: Ixodidae) From Urban and Peri-Urban Areas of Kosovo. Zoonoses Public Health 2025, 72, 174–183. [Google Scholar] [CrossRef]
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubálek, Z.; Földvári, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Špitalská, E.; et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: New hazards and relevance for public health. Front. Public Health 2014, 2, 251. [Google Scholar] [CrossRef]
- Hodžić, A.; Georges, I.; Postl, M.; Duscher, G.G.; Jeschke, D.; Szentiks, C.A.; Ansorge, H.; Heddergott, M. Molecular survey of tick-borne pathogens reveals a high prevalence and low genetic variability of Hepatozoon canis in free-ranging grey wolves (Canis lupus) in Germany. Ticks Tick. Borne. Dis. 2020, 11, 101389. [Google Scholar] [CrossRef]
- Gritsun, T.S.; Lashkevich, V.A.; Gould, E.A. Tick-borne encephalitis. Antiviral Res. 2003, 57, 129–146. [Google Scholar] [CrossRef]
- Süss, J. Tick-borne encephalitis 2010: Epidemiology, risk areas, and virus strains in Europe and Asia—An overview. Ticks Tick. Borne. Dis. 2011, 2, 2–15. [Google Scholar] [CrossRef]
- Drehmann, M.; Springer, A.; Lindau, A.; Fachet, K.; Mai, S.; Thoma, D.; Schneider, C.R.; Chitimia-Dobler, L.; Bröker, M.; Dobler, G.; et al. The Spatial Distribution of Dermacentor Ticks (Ixodidae) in Germany—Evidence of a Continuing Spread of Dermacentor reticulatus. Front. Vet. Sci. 2020, 7, 578220. [Google Scholar] [CrossRef]
- Halos, L.; Bord, S.; Cotté, V.; Gasqui, P.; Abrial, D.; Barnouin, J.; Boulouis, H.-J.; Vayssier-Taussat, M.; Vourc’h, G. Ecological Factors Characterizing the Prevalence of Bacterial Tick-Borne Pathogens in Ixodes ricinus Ticks in Pastures and Woodlands. Appl. Environ. Microbiol. 2010, 76, 4413–4420. [Google Scholar] [CrossRef]
- Shaw, G.; Lilly, M.; Mai, V.; Clark, J.; Summers, S.; Slater, K.; Karpathy, S.; Nakano, A.; Crews, A.; Lawrence, A.; et al. The roles of habitat isolation, landscape connectivity and host community in tick-borne pathogen ecology. R. Soc. Open Sci. 2024, 11, 240837. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Gray, J.S.; Kahl, O.; Lane, R.S.; Nijhof, A.M. Research on the ecology of ticks and tick-borne pathogens—Methodological principles and caveats. Front. Cell. Infect. Microbiol. 2013, 3, 29. [Google Scholar] [CrossRef] [PubMed]
- Heylen, D.; Lasters, R.; Adriaensen, F.; Fonville, M.; Sprong, H.; Matthysen, E. Ticks and tick-borne diseases in the city: Role of landscape connectivity and green space characteristics in a metropolitan area. Sci. Total Environ. 2019, 670, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S.E. Evidence that climate change has caused ‘emergence’ of tick-borne diseases in Europe? Int. J. Med. Microbiol. Suppl. 2004, 293, 5–15. [Google Scholar] [CrossRef]
- Pfäffle, M.; Littwin, N.; Muders, S.V.; Petney, T.N. The ecology of tick-borne diseases. Int. J. Parasitol. 2013, 43, 1059–1077. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).