
The Strategy and Application of Gene Attenuation in Metabolic Engineering



Abstract
1. Introduction
1.1. Overview of Metabolic Engineering and Its Significance
1.2. Development of Gene Editing Technology
1.3. The Importance of Gene Attenuation in Metabolic Engineering
1.4. Development and Mechanisms of Gene Attenuation Technology
2. Applications of Gene Attenuation Across Various Biological Systems

{kind=link}
{kind=link}
| System | Organisms | Strategy | Application | Ref. |
|---|---|---|---|---|
| Plant System | Rice | RNAi | It can considerably enhance the salt and drought tolerance of rice, as well as increase its yield under stress conditions. | [82] |
| Arabidopsis thaliana | asRNA | The reduction in SPS in Arabidopsis maintains carbon allocation balance. | [84] | |
| Nicotiana tabacum | asRNA | In tobacco plants, inhibiting the expression of tumor formation genes enhances disease resistance potential. | [85] | |
| Animal System | Zebrafish | RNAi | Induces double-muscle phenotype in zebrafish, enhances muscle development. | [87] |
| Insect pests | RNAi | Reduces expression of porcine endogenous retrovirus, decreasing virus transmission risk in xenotransplantation. | [91] | |
| Microbial System | E. coli | CRISPRi | The content of 4HB further increased from 1.43 mol% to 18.4 mol%, an increase of about 12.8 times. | [92] |
| Clostridium acetobutylicum | asRNA | The production of butanol and acetone was increased by about 35% and 50%, respectively, and the yield of lactic acid was effectively increased by the asRNA strategy. | [93] | |
| E. coli | asRNA | By using asRNA technology to weaken the pgi gene, the production of 2-hydroxyadipic acid was increased by about 24%, and the intracellular NADPH supply was enhanced. | [95] | |
| E. coli | sRNA | By weakening the translation of the murE gene by sRNA, the production of cadaverine could be increased to 12.6 g/L, which was 31% higher than that of the basic strain. | [98] |
3. Challenges and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CRISPRi | CRISPR interference |
| RNAi | RNA interference |
| AsRNA | Antisense RNA |
| SRNAs | Small RNAs |
| SiRNAs | Small interfering RNAs |
| TALENs | Transcription Activator-Like Effector Nucleases |
| ZFNs | Antisense RNA |
| RBS | Ribosome Binding Sites |
| CHS | Chalcone Synthase |
| PTGS | Post-transcriptional gene silencing |
| MiRNAs | MicroRNAs |
| PiRNAs | Piwi-interacting RNAs |
| DsRNA | Double-stranded RNA |
| RISC | RNA-induced silencing complex |
| RdRP | RNA-dependent RNA polymerase |
| ShRNA | Short hairpin RNA |
| LVA | Leucine-Valine-Alanine |
| PegRNA | Prime editing guide RNA |
| PAM | Protospacer adjacent motif |
References
- Bailey, J.E. Toward a science of metabolic engineering. Science 1991, 252, 1668–1675. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.; Keasling, J.D. Engineering cellular metabolism. Cell 2016, 164, 1185–1197. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.; Villadsen, J.; Gunnar, L. Bioreaction Engineering Principles, 2nd ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2003. [Google Scholar]
- Keasling, J.D. Manufacturing molecules through metabolic engineering. Science 2010, 330, 1355–1358. [Google Scholar] [CrossRef] [PubMed]
- Stephanopoulos, G.; Vallino, J.J. Network rigidity and metabolic engineering in metabolite overproduction. Science 1991, 252, 1675–1681. [Google Scholar] [CrossRef]
- Huang, Z.; Xiao, L.; Mo, W.; Zhang, Y.; Cai, Y.; Huang, S.; Chen, Z.; Long, C. Molecular Mechanism of Mok I Gene Overexpression in Enhancing Monacolin K Production in Monascus pilosus. J. Fungi 2024, 10, 721. [Google Scholar] [CrossRef]
- Shannon, K.M. Recent advances in the treatment of huntington’s disease: Targeting DNA and RNA. CNS Drugs 2020, 34, 219–228. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhao, M.; Wang, H.; Zhu, Y.; Mu, W. Metabolic engineering of Escherichia coli BL21(DE3) cocultured with glucose and xylose for efficient production of 2′-fucosyllactose. Bioresour. Technol. 2025, 419, 132062. [Google Scholar] [CrossRef]
- Xie, H.; Su, F.; Niu, Q.; Geng, L.; Cao, X.; Song, M.; Dong, J.; Zheng, Z.; Guo, R.; Zhang, Y.; et al. Knockout of miR396 genes increases seed size and yield in soybean. J. Integr. Plant. Biol. 2024, 66, 1148–1157. [Google Scholar] [CrossRef]
- Huang, C.H.; Shen, C.R.; Li, H.; Sung, L.Y.; Wu, M.Y.; Hu, Y.C. CRISPR interference (CRISPRi) for gene regulation and succinate production in cyanobacterium S. elongatus PCC 7942. Microb. Cell. Fact. 2016, 15, 196. [Google Scholar] [CrossRef]
- Rudin, N.; Haber, J.E. Efficient repair of HO-induced chromosomal breaks in Saccharomyces cerevisiae by recombination between flanking homologous sequences. Mol. Cell. Biol. 1988, 8, 3918–3928. [Google Scholar]
- Urnov, F.D.; Miller, J.C.; Lee, Y.L.; Beausejour, C.M.; Rock, J.M.; Augustus, S.; Jamieson, A.C.; Porteus, M.H.; Gregory, P.D.; Holmes, M.C. Highly efficient endogenous human gene correction using designed zinc-finger nucleases. Nature 2005, 435, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.G.; Cha, J.; Chandrasegaran, S. Hybrid restriction enzymes: Zinc finger fusions to Fok I cleavage domain. Proc. Natl. Acad. Sci. USA 1996, 93, 1156–1160. [Google Scholar] [CrossRef] [PubMed]
- Carroll, D. Genome engineering with zinc-finger nucleases. Genetics 2011, 188, 773–782. [Google Scholar] [CrossRef]
- Sun, N.; Zhao, H. Transcription activator-like effector nucleases (TALENs): A highly efficient and versatile tool for genome editing. Biotechnol. Bioeng. 2013, 110, 1811–1821. [Google Scholar] [CrossRef]
- Li, T.; Huang, S.; Zhao, X.; Wright, D.A.; Carpenter, S.; Spalding, M.H.; Weeks, D.P.; Yang, B. Modularly assembled designer TAL effector nucleases for targeted gene knockout and gene replacement in eukaryotes. Nucleic Acids Res. 2011, 39, 6315–6325. [Google Scholar] [CrossRef]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef]
- Mojica, F.J.; Díez-Villaseñor, C.; Soria, E.; Juez, G. Biological significance of a family of regularly spaced repeats in the genomes of Archaea, Bacteria and mitochondria. Mol. Microbiol. 2000, 36, 244–246. [Google Scholar] [CrossRef]
- Gilbert, L.A.; Horlbeck, M.A.; Adamson, B.; Villalta, J.E.; Chen, Y.; Whitehead, E.H.; Guimaraes, C.; Panning, B.; Ploegh, H.L.; Bassik, M.C.; et al. Genome-Scale CRISPR-Mediated control of gene repression and activation. Cell 2014, 159, 647–661. [Google Scholar] [CrossRef]
- Wang, H.; Yang, H.; Shivalila, C.S.; Dawlaty, M.M.; Cheng, A.W.; Zhang, F.; Jaenisch, R. One-step generation of mice carrying mutations in multiple genes by CRISPR/Cas-mediated genome engineering. Cell 2013, 153, 910–918. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhu, S.; Cai, C.; Yuan, P.; Li, C.; Huang, Y.; Wei, W. High-throughput screening of a CRISPR/Cas9 library for functional genomics in human cells. Nature 2014, 509, 487–491. [Google Scholar] [CrossRef]
- Shalem, O.; Sanjana, N.E.; Hartenian, E.; Shi, X.; Scott, D.A.; Mikkelson, T.; Heckl, D.; Ebert, B.L.; Root, D.E.; Doench, J.G.; et al. Genome-scale CRISPR-Cas9 knockout screening in human cells. Science 2014, 343, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Koike-Yusa, H.; Li, Y.; Tan, E.P.; Velasco-Herrera Mdel, C.; Yusa, K. Genome-wide recessive genetic screening in mammalian cells with a lentiviral CRISPR-guide RNA library. Nat. Biotechnol. 2014, 32, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wei, J.J.; Sabatini, D.M.; Lander, E.S. Genetic screens in human cells using the CRISPR-Cas9 system. Science 2014, 343, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.B.T.; Gootenberg, J.S.; Abudayyeh, O.O.; Franklin, B.; Kellner, M.J.; Joung, J.; Zhang, F. RNA editing with CRISPR-Cas13. Science 2017, 358, 1019–1027. [Google Scholar] [CrossRef]
- Chen, J.S.; Ma, E.; Harrington, L.B.; Da Costa, M.; Tian, X.; Palefsky, J.M.; Doudna, J.A. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science 2018, 360, 436–439. [Google Scholar] [CrossRef]
- Giaever, G.; Chu, A.M.; Ni, L.; Connelly, C.; Riles, L.; Véronneau, S.; Dow, S.; Lucau-Danila, A. Functional profiling of the Saccharomyces cerevisiae genome. Nature 2002, 418, 387–391. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, H.U. Systems strategies for developing industrial microbial strains. Nat. Biotechnol. 2015, 33, 1061–1072. [Google Scholar] [CrossRef]
- Chen, R.; Shi, F.; Xiang, Y.; Lai, W.; Ji, G. Establishment of CRISPR-Cpf1-assisted gene editing tool and engineering of 4-hydroxyisoleucine biosynthesis in Corynebacterium glutamicum. World J. Microbiol. Biotechnol. 2023, 39, 266. [Google Scholar] [CrossRef]
- Schultenkamper, K.; Brito, L.F.; Wendisch, V.F. Impact of CRISPR interference on strain development in biotechnology. Biotechnol. Appl. Biochem. 2020, 67, 7–21. [Google Scholar] [CrossRef]
- Zou, G.M. RNAi technique in stem cell research: Current status and future perspectives. Methods Mol. Biol. 2017, 1622, 3–14. [Google Scholar]
- Lau, C.H.; Suh, Y. In vivo epigenome editing and transcriptional modulation using CRISPR technology. Transgen. Res. 2018, 27, 489–509. [Google Scholar] [CrossRef]
- Lin, J.; Lai, G.; Guo, A.; He, L.; Yang, F.; Huang, Y.; Che, J.; Lai, C. Overexpression of LAR1 suppresses anthocyanin biosynthesis by enhancing catechin competition leading to promotion of proanthocyanidin pathway in spine grape (Vitis davidii) cells. Int. J. Mol. Sci. 2024, 25, 12087. [Google Scholar] [CrossRef]
- Li, T.; Huang, S.; Jiang, W.Z.; Wright, D.; Spalding, M.H.; Weeks, D.P.; Yang, B. TAL nucleases (TALNs): Hybrid proteins composed of TAL effectors and FokI DNA-cleavage domain. Nucleic Acids Res. 2011, 39, 359–372. [Google Scholar] [CrossRef]
- Urnov, F.D.; Rebar, E.J.; Holmes, M.C.; Zhang, H.S.; Gregory, P.D. Genome editing with engineered zinc finger nucleases. Nat. Rev. Genet. 2010, 11, 636–646. [Google Scholar] [CrossRef]
- Napoli, C.; Lemieux, C.; Jorgensen, R. Introduction of a chimeric chalcone synthase gene into petunia results in reversible co-suppression of homologous genes in trans. Plant Cell 1990, 2, 279–289. [Google Scholar] [CrossRef]
- Kooter, J.M.; Matzke, M.A.; Meyer, P. Listening to the silent genes: Transgene silencing, gene regulation and pathogen control. Trends Plant Sci. 1999, 4, 340–347. [Google Scholar] [CrossRef]
- Agrawal, N.; Dasaradhi, P.V.; Mohmmed, A.; Malhotra, P.; Bhatnagar, R.K.; Mukherjee, S.K. RNA interference: Biology, mechanism, and applications. Microbiol. Mol. Biol. Rev. 2003, 67, 657–685. [Google Scholar] [CrossRef]
- Cogoni, C.; Irelan, J.T.; Schumacher, M.; Schmidhauser, T.J.; Selker, E.U.; Macino, G. Transgene silencing of the al-1 gene in vegetative cells of Neurospora is mediated by a cytoplasmic effector and does not depend on DNA-DNA interactions or DNA methylation. Embo. J. 1996, 15, 3153–3163. [Google Scholar] [CrossRef]
- Szweykowska-Kulińska, Z.; Jarmołowski, A.; Figlerowicz, M. RNA interference and its role in the regulation of eucaryotic gene expression. Acta Biochim. Pol. 2003, 50, 217–229. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Brantl, S. Antisense-RNA regulation and RNA interference. Biochim. Biophys. Acta 2002, 1575, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Tomizawa, J.; Itoh, T.; Selzer, G.; Som, T. Inhibition of ColE1 RNA primer formation by a plasmid-specified small RNA. Proc. Natl. Acad. Sci. USA 1981, 78, 1421–1425. [Google Scholar] [CrossRef]
- Guo, L.; Liu, M.; Bi, Y.; Qi, Q.; Xian, M.; Zhao, G. Using a synthetic machinery to improve carbon yield with acetylphosphate as the core. Nat. Commun. 2023, 14, 5286. [Google Scholar] [CrossRef]
- Lamichhane, G.; Arnvig, K.B.; McDonough, K.A. Definition and annotation of (myco)bacterial non-coding RNA. Tuberculosis 2013, 93, 26–29. [Google Scholar] [CrossRef]
- Dutta, T.; Srivastava, S. Small RNA-mediated regulation in bacteria: A growing palette of diverse mechanisms. Gene 2018, 656, 60–72. [Google Scholar] [CrossRef]
- Ambros, V.; Chen, X. The regulation of genes and genomes by small RNAs. Development 2007, 134, 1635–1641. [Google Scholar] [CrossRef]
- Sun, D.; Chen, J.; Wang, Y.; Li, M.; Rao, D.; Guo, Y.; Chen, N.; Zheng, P.; Sun, J.; Ma, Y. Metabolic engineering of Corynebacterium glutamicum by synthetic small regulatory RNAs. J. Ind. Microbiol. Biotechnol. 2019, 46, 203–208. [Google Scholar] [CrossRef]
- He, Y.; Ju, C.; Zhang, X. Roles of small RNAs in the immune defense mechanisms of crustaceans. Mol. Immunol. 2015, 68, 399–403. [Google Scholar] [CrossRef]
- Filipowicz, W. RNAi: The Nuts and Bolts of the RISC Machine. Cell 2005, 122, 17–20. [Google Scholar] [CrossRef]
- Si, T.; HamediRad, M.; Zhao, H. Regulatory RNA-assisted genome engineering in microorganisms. Curr. Opin. Biotechnol. 2015, 36, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Zamore, P.D.; Tuschl, T.; Sharp, P.A.; Bartel, D.P. RNAi: Double-Stranded RNA directs the ATP-Dependent Cleavage of mRNA at 21 to 23 Nucleotide Intervals. Cell 2000, 101, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Feng, X.; Mao, H.; Li, M.; Xu, F.; Hu, K.; Guang, S. RdRP-synthesized antisense ribosomal siRNAs silence pre-rRNA via the nuclear RNAi pathway. Nat. Struct. Mol. Biol. 2017, 24, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.Y.; Chen, C.C.; Conte, D., Jr.; Moresco, J.J.; Chaves, D.A.; Mitani, S.; Yates, J.R., 3rd; Tsai, M.D.; Mello, C.C. A ribonuclease Coordinates siRNA Amplification and mRNA Cleavage during RNAi. Cell 2015, 160, 407–419. [Google Scholar] [CrossRef]
- Paddison, P.J.; Caudy, A.A.; Bernstein, E.; Hannon, G.J.; Conklin, D.S. Short hairpin RNAs (shRNAs) induce sequence-specific silencing in mammalian cells. Genes. Dev. 2002, 16, 948–958. [Google Scholar] [CrossRef]
- Stewart, C.K.; Li, J.; Golovan, S.P. Adverse effects induced by short hairpin RNA expression in porcine fetal fibroblasts. Biochem. Biophys. Res. Commun. 2008, 370, 113–117. [Google Scholar] [CrossRef]
- Hasuwa, H.; Kaseda, K.; Einarsdottir, T.; Okabe, M. Small interfering RNA and gene silencing in transgenic mice and rats. FEBS Lett. 2002, 532, 227–230. [Google Scholar] [CrossRef]
- Shmakov, S.; Smargon, A.; Scott, D.; Cox, D.; Pyzocha, N.; Yan, W.; Abudayyeh, O.O.; Gootenberg, J.S.; Makarova, K.S.; Wolf, Y.I.; et al. Diversity and evolution of class 2 CRISPR-Cas systems. Nat. Rev. Microbiol. 2017, 15, 169–182. [Google Scholar] [CrossRef]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; van der Oost, J.; Regev, A.; et al. Cpf1 is a single RNA-guided endonuclease of a class 2 CRISPR-Cas system. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Alkhnbashi, O.S.; Costa, F.; Shah, S.A.; Saunders, S.J.; Barrangou, R.; Brouns, S.J.; Charpentier, E.; Haft, D.H.; et al. An updated evolutionary classification of CRISPR-Cas systems. Nat. Rev. Microbiol. 2015, 13, 722–736. [Google Scholar] [CrossRef]
- Vigouroux, A.; Bikard, D. CRISPR tools to control gene expression in Bacteria. Microbiol. Mol. Biol. Rev. 2020, 84, 1128. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.S.; Larson, M.H.; Gilbert, L.A.; Doudna, J.A.; Weissman, J.S.; Arkin, A.P.; Lim, W.A. Repurposing CRISPR as an RNA-Guided platform for sequence-specific control of gene expression. Cell 2013, 152, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Mathur, F.; Arora, A. CRISPR-Based genetic control strategies for insect pests to mitigate classical insecticidal approaches. In Gene Editing in Plants; Springer: Singapore, 2024; pp. 667–707. [Google Scholar]
- Gilbert, L.A.; Larson, M.H.; Morsut, L.; Liu, Z.; Brar, G.A.; Torres, S.E.; Stern-Ginossar, N.; Brandman, O.; Whitehead, E.H.; Doudna, J.A.; et al. CRISPR-mediated modular RNA-guided regulation of transcription in Eukaryotes. Cell 2013, 154, 442–451. [Google Scholar] [CrossRef]
- Kampmann, M. CRISPRi and CRISPRa screens in mammalian cells for precision biology and medicine. ACS Chem. Biol. 2018, 13, 406–416. [Google Scholar] [CrossRef]
- Konermann, S.; Brigham, M.D.; Trevino, A.E.; Joung, J.; Abudayyeh, O.O.; Barcena, C.; Hsu, P.D.; Habib, N.; Gootenberg, J.S.; Nishimasu, H.; et al. Genome-scale transcriptional activation by an engineered CRISPR-Cas9 complex. Nature 2015, 517, 583–588. [Google Scholar] [CrossRef]
- Hillary, V.E.; Ceasar, S.A. A review on the mechanism and applications of CRISPR/Cas9/Cas12/Cas13/Cas14 proteins utilized for genome engineering. Mol. Biotechnol. 2023, 65, 311–325. [Google Scholar] [CrossRef]
- Li, S.; Luo, Y.; Wei, G.; Zong, W.; Zeng, W.; Xiao, D.; Zhang, H.; Song, Y.; Hao, Y.; Sun, K.; et al. Improving yield-related traits by editing the promoter of the heading date gene Ehd1 in rice. Theor. Appl. Genet. 2023, 136, 239. [Google Scholar] [CrossRef]
- Zhou, C.; Ye, B.; Cheng, S.; Zhao, L.; Liu, Y.; Jiang, J.; Yan, X. Promoter engineering enables overproduction of foreign proteins from a single copy expression cassette in Bacillus subtilis. Microb. Cell. Fact. 2019, 18, 111. [Google Scholar] [CrossRef]
- Shi, F.; Fan, Z.; Zhang, S.; Wang, Y.; Tan, S.; Li, Y. Optimization of ribosomal binding site sequences for gene expression and 4-hydroxyisoleucine biosynthesis in recombinant Corynebacterium glutamicum. Enzyme Microb. Technol. 2020, 140, 109622. [Google Scholar] [CrossRef]
- Miller, J.C.; Tan, S.; Qiao, G.; Barlow, K.A.; Wang, J.; Xia, D.F.; Meng, X.; Paschon, D.E.; Leung, E.; Hinkley, S.J.; et al. A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. 2011, 29, 143–148. [Google Scholar] [CrossRef]
- Christian, M.; Cermak, T.; Doyle, E.L.; Schmidt, C.; Zhang, F.; Hummel, A.; Bogdanove, A.J.; Voytas, D.F. Targeting DNA double-strand breaks with TAL effector nucleases. Genetics 2010, 186, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.; McLachlan, A.; Klug, A. Repetitive zinc-binding domains in the protein transcription factor IIIA from Xenopus oocytes. EMBO J. 1985, 4, 1609–1614. [Google Scholar] [CrossRef] [PubMed]
- Bitinaite, J.; Wah, D.A.; Aggarwal, A.K.; Schildkraut, I. FokI dimerization is required for DNA cleavage. Proc. Natl. Acad. Sci. USA 1998, 95, 10570–10575. [Google Scholar] [CrossRef] [PubMed]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F., 3rd. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef]
- Chen, Y.; Li, Y.; Narayan, R.; Subramanian, A.; Xie, X. Gene expression inference with deep learning. Bioinformatics 2016, 32, 1832–1839. [Google Scholar] [CrossRef]
- Badia, I.M.P.; Wessels, L.; Muller-Dott, S.; Trimbour, R.; Ramirez Flores, R.O.; Argelaguet, R.; Saez-Rodriguez, J. Gene regulatory network inference in the era of single-cell multi-omics. Nat. Rev. Genet. 2023, 24, 739–754. [Google Scholar] [CrossRef]
- Li, T.; Xu, H.; Teng, S.; Suo, M.; Bahitwa, R.; Xu, M.; Qian, Y.; Ramstein, G.P.; Song, B.; Buckler, E.S.; et al. Modeling 0.6 million genes for the rational design of functional cis-regulatory variants and de novo design of cis-regulatory sequences. Proc. Natl. Acad. Sci. USA 2024, 121, e2319811121. [Google Scholar] [CrossRef]
- Lewis, N.E.; Nagarajan, H.; Palsson, B.O. Constraining the metabolic genotype-phenotype relationship using a phylogeny of in silico methods. Nat. Rev. Microbiol. 2012, 10, 291–305. [Google Scholar] [CrossRef]
- Marchisio, M.A.; Stelling, J. Computational design of synthetic gene circuits with composable parts. Bioinformatics 2008, 24, 1903–1910. [Google Scholar] [CrossRef]
- Yadav, C.; Rawat, N.; Singla-Pareek, S.L.; Pareek, A. Knockdown of OsPHP1 leads to improved yield under salinity and drought in rice via regulating the complex Set of TCS members and cytokinin signalling. Plant Cell Environ. 2024, 48, 2769–2782. [Google Scholar] [CrossRef]
- Strand, A.; Zrenner, R.; Trevanion, S.; Stitt, M.; Gustafsson, P.; Gardeström, P. Decreased expression of two key enzymes in the sucrose biosynthesis pathway, cytosolic fructose-1,6-bisphosphatase and sucrose phosphate synthase, has remarkably different consequences for photosynthetic carbon metabolism in transgenic Arabidopsis thaliana. Plant J. 2000, 23, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Draborg, H.; Villadsen, D.; Nielsen, T.H. Transgenic arabidopsis plants with decreased activity of fructose-6-phosphate,2-kinase/fructose-2,6-bisphosphatase have altered carbon partitioning. Plant Physiol. 2001, 126, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Alekseeva, V.V.; Rukavtsova, E.B.; Golubchikova, Y.S.; Buryanov, Y.I. Inhibition of agrobacterial oncogene expression by means of antisense RNAs. Mol. Biol. 2011, 42, 153–157. [Google Scholar] [CrossRef]
- Animasaun, D.A.; Lawrence, J.A. Antisense RNA (asRNA) technology: The concept and applications in crop improvement and sustainable agriculture. Mol. Biol. Rep. 2023, 50, 9545–9557. [Google Scholar] [CrossRef]
- Acosta, J.; Carpio, Y.; Borroto, I.; Gonzalez, O.; Estrada, M.P. Myostatin gene silenced by RNAi show a zebrafish giant phenotype. J. Biotechnol. 2005, 119, 324–331. [Google Scholar] [CrossRef]
- Magee, T.R.; Artaza, J.N.; Ferrini, M.G.; Vernet, D.; Zuniga, F.I.; Cantini, L.; Reisz-Porszasz, S.; Rajfer, J.; Gonzalez-Cadavid, N.F. Myostatin short interfering hairpin RNA gene transfer increases skeletal muscle mass. J. Gene. Med. 2006, 8, 1171–1181. [Google Scholar] [CrossRef]
- Naeem, M.; Majeed, S.; Hoque, M.Z.; Ahmad, I. Latest developed strategies to minimize the Off-Target effects in CRISPR-Cas-Mediated genome editing. Cells 2020, 9, 1608. [Google Scholar] [CrossRef]
- Dieckhoff, B.; Karlas, A.; Hofmann, A.; Kues, W.A.; Petersen, B.; Pfeifer, A.; Niemann, H.; Kurth, R.; Denner, J. Inhibition of porcine endogenous retroviruses (PERVs) in primary porcine cells by RNA interference using lentiviral vectors. Arch. Virol. 2007, 152, 629–634. [Google Scholar] [CrossRef]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef]
- Lv, L.; Ren, Y.L.; Chen, J.C.; Wu, Q.; Chen, G.Q. Application of CRISPRi for prokaryotic metabolic engineering involving multiple genes, a case study: Controllable P(3HB-co-4HB) biosynthesis. Metab. Eng. 2015, 29, 160–168. [Google Scholar] [CrossRef]
- Desai, R.P.; Papoutsakis, E.T. Antisense RNA strategies for metabolic engineering of Clostridium acetobutylicum. Appl. Environ. Microbiol. 1999, 65, 936–945. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yu, H.; Shen, Z. Antisense RNA-Based strategy for enhancing surfactin production in Bacillus subtilis TS1726 via overexpression of the unconventional biotin carboxylase II To enhance ACCase activity. ACS Synth. Biol. 2019, 8, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; He, K.; Bi, H.; Wang, M.; Chen, B.; Tan, T.; Zhang, Y. Metabolic Engineering for Effective Synthesis of 2-Hydroxyadipate. ACS Synth. Biol. 2023, 12, 2475–2486. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Peng, A.; Zhang, L.; Wang, Y.; Du, G.; Chen, J.; Kang, Z. Design of artificial small regulatory trans-RNA for gene knockdown in Bacillus subtilis. Synth. Syst. Biotechnol. 2023, 8, 61–68. [Google Scholar] [CrossRef]
- Fu, J.; Zaghen, S.; Lu, H.; Konzock, O.; Poorinmohammad, N.; Kornberg, A.; Ledesma-Amaro, R.; Koseto, D.; Wentzel, A.; Di Bartolomeo, F.; et al. Reprogramming Yarrowia lipolytica metabolism for efficient synthesis of itaconic acid from flask to semipilot scale. Sci. Adv. 2024, 10, eadn0414. [Google Scholar] [CrossRef]
- Na, D.; Yoo, S.M.; Chung, H.; Park, H.; Park, J.H.; Lee, S.Y. Metabolic engineering of Escherichia coli using synthetic small regulatory RNAs. Nat. Biotechnol. 2013, 31, 170–174. [Google Scholar] [CrossRef]
- Wu, J.; Du, G.; Chen, J.; Zhou, J. Enhancing flavonoid production by systematically tuning the central metabolic pathways based on a CRISPR interference system in Escherichia coli. Sci. Rep. 2015, 5, 13477. [Google Scholar] [CrossRef]
- Kim, S.K.; Seong, W.; Han, G.H.; Lee, D.H.; Lee, S.G. CRISPR interference-guided multiplex repression of endogenous competing pathway genes for redirecting metabolic flux in Escherichia coli. Microb. Cell. Fact. 2017, 16, 188. [Google Scholar] [CrossRef]
- Li, H.; Wang, B.S.; Li, Y.R.; Zhang, L.; Ding, Z.Y.; Gu, Z.H.; Shi, G.Y. Metabolic engineering of Escherichia coli W3110 for the production of L-methionine. J. Ind. Microbiol. Biotechnol. 2017, 44, 75–88. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, Y.; Cui, J.; Guo, G.; Li, Y.; Hou, J.; Tu, Q.; Yin, Y.; Stewart, A.F.; Zhang, Y.; et al. ReaL-MGE is a tool for enhanced multiplex genome engineering and application to malonyl-CoA anabolism. Nat. Commun. 2024, 15, 9790. [Google Scholar] [CrossRef]
- Wang, X.; Fang, C.; Wang, Y.; Shi, X.; Yu, F.; Xiong, J.; Chou, S.H.; He, J. Systematic comparison and rational design of theophylline riboswitches for effective gene repression. Microbiol. Spectr. 2023, 11, e02752-22. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Xu, M.; Ma, Z.; Pan, X.; You, J.; Hu, M.; Shao, Y.; Yang, T.; Zhang, X.; Rao, Z. Significantly enhancing production of trans-4-hydroxy-l-proline by integrated system engineering in Escherichia coli. Sci. Adv. 2020, 6, eaba2383. [Google Scholar] [CrossRef] [PubMed]
- Bartoszewski, R.; Sikorski, A.F. Editorial focus: Understanding off-target effects as the key to successful RNAi therapy. Cell. Mol. Biol. Lett. 2019, 24, 69. [Google Scholar] [CrossRef] [PubMed]
- Barrangou, R.; Birmingham, A.; Wiemann, S.; Beijersbergen, R.L.; Hornung, V.; Smith, A.V.B. B. Advances in CRISPR-Cas9 genome engineering: Lessons learned from RNA interference. Nucleic Acids Res. 2015, 43, 3407–3419. [Google Scholar] [CrossRef]
- Chuang, Y.F.; Phipps, A.J.; Lin, F.L.; Hecht, V.; Hewitt, A.W.; Wang, P.Y.; Liu, G.S. Approach for in vivo delivery of CRISPR/Cas system: A recent update and future prospect. Cell. Mol. Life. Sci. 2021, 78, 2683–2708. [Google Scholar] [CrossRef]
- Birmingham, A.; Anderson, E.M.; Reynolds, A.; Ilsley-Tyree, D.; Leake, D.; Fedorov, Y.; Baskerville, S.; Maksimova, E.; Robinson, K.; Karpilow, J.; et al. 3′ UTR seed matches, but not overall identity, are associated with RNAi off-targets. Nat. Methods 2006, 3, 199–204. [Google Scholar] [CrossRef]
- Buehler, E.; Khan, A.A.; Marine, S.; Rajaram, M.; Bahl, A.; Burchard, J.; Ferrer, M. siRNA off-target effects in genome-wide screens identify signaling pathway members. Sci. Rep. 2012, 2, 428. [Google Scholar] [CrossRef]
- Jackson, A.L.; Linsley, P.S. Recognizing and avoiding siRNA off-target effects for target identification and therapeutic application. Nat. Rev. Drug. Discov. 2010, 9, 57–67. [Google Scholar] [CrossRef]
- Seok, H.; Lee, H.; Jang, E.S.; Chi, S.W. Evaluation and control of miRNA-like off-target repression for RNA interference. Cell. Mol. Life. Sci. 2018, 75, 797–814. [Google Scholar] [CrossRef]
- Elmén, J.; Thonberg, H.; Ljungberg, K.; Frieden, M.; Westergaard, M.; Xu, Y.; Wahren, B.; Liang, Z.; Ørum, H.; Koch, T.; et al. Locked nucleic acid (LNA) mediated improvements in siRNA stability and functionality. Nucleic Acids Res. 2005, 33, 439–447. [Google Scholar] [CrossRef]
- Hannus, M.; Beitzinger, M.; Engelmann, J.C.; Weickert, M.T.; Spang, R.; Hannus, S.; Meister, G. siPools: Highly complex but accurately defined siRNA pools eliminate off-target effects. Nucleic Acids Res. 2014, 42, 8049–8061. [Google Scholar] [CrossRef] [PubMed]
- Lopes, R.; Prasad, M.K. Beyond the promise: Evaluating and mitigating off-target effects in CRISPR gene editing for safer therapeutics. Front. Bioeng. Biotechnol. 2023, 11, 1339189. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.H.; Miller, S.M.; Geurts, M.H.; Tang, W.; Chen, L.; Sun, N.; Zeina, C.M.; Gao, X.; Rees, H.A.; Lin, Z.; et al. Evolved Cas9 variants with broad PAM compatibility and high DNA specificity. Nature 2018, 556, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Matson, A.W.; Hosny, N.; Swanson, Z.A.; Hering, B.J.; Burlak, C. Optimizing sgRNA length to improve target specificity and efficiency for the GGTA1 gene using the CRISPR/Cas9 gene editing system. PLoS ONE 2019, 14, e0226107. [Google Scholar] [CrossRef]
- Aquino-Jarquin, G. Current advances in overcoming obstacles of CRISPR/Cas9 off-target genome editing. Mol. Genet. Metab. 2021, 134, 77–86. [Google Scholar] [CrossRef]
- Jain, S.; Xun, G.; Abesteh, S.; Ho, S.; Lingamaneni, M.; Martin, T.A.; Tasan, I.; Yang, C.; Zhao, H. Precise regulation of Cas9-mediated genome engineering by Anti-CRISPR-Based inducible CRISPR controllers. ACS Synth. Biol. 2021, 10, 1320–1327. [Google Scholar] [CrossRef]
- Casini, A.; Olivieri, M.; Petris, G.; Montagna, C.; Reginato, G.; Maule, G.; Lorenzin, F.; Prandi, D.; Romanel, A.; Demichelis, F.; et al. A highly specific SpCas9 variant is identified by in vivo screening in yeast. Nat. Biotechnol. 2018, 36, 265–271. [Google Scholar] [CrossRef]
- Ye, C.; Yang, Y.; Chen, X.; Yang, L.; Hua, X.; Yang, M.; Zeng, X.; Qiao, S. Metabolic engineering of Escherichia coli BW25113 for the production of 5-Aminolevulinic Acid based on CRISPR/Cas9 mediated gene knockout and metabolic pathway modification. J. Biol. Eng. 2022, 16, 26. [Google Scholar] [CrossRef]
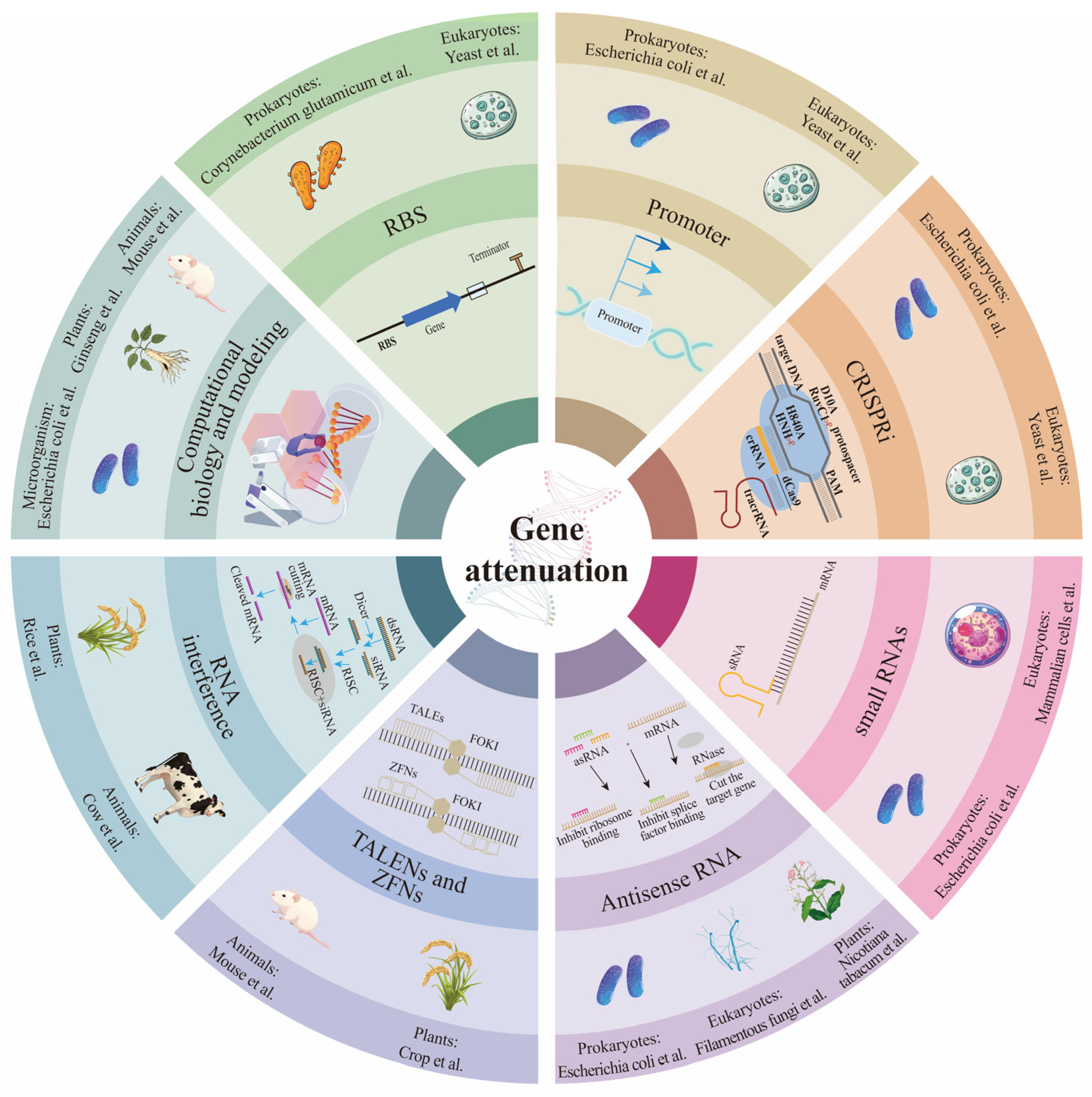
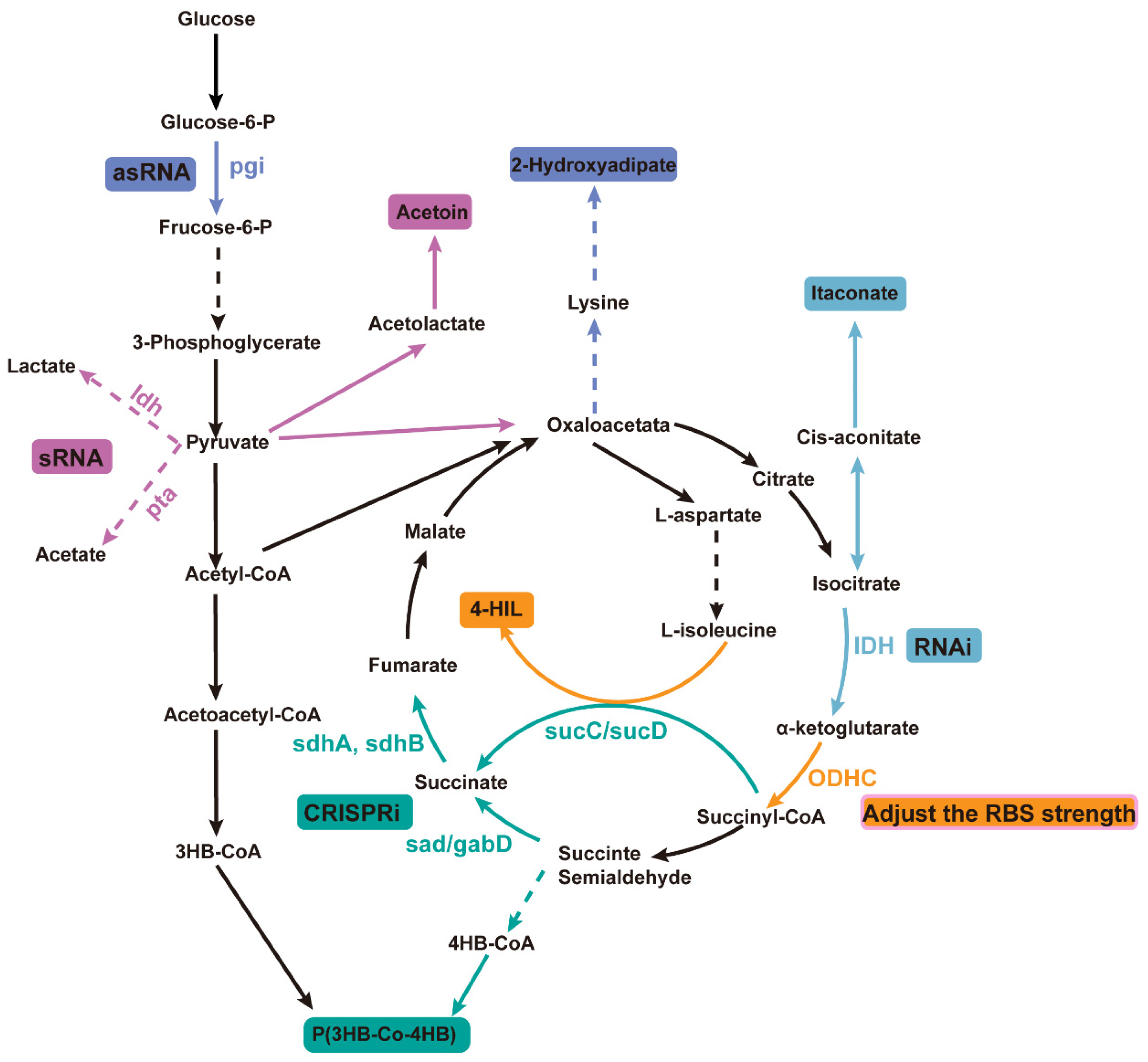
| Strategy | Description | Methods | Applications | Ref. |
|---|---|---|---|---|
| Gene attenuation | Reduces gene expression or lowers product activity to weaken function. | RNAi, CRISPRi, etc. | Studying the effects of reduced gene expression on an organism. | [30,31] |
| Gene knockout | Completely removes or deactivates a gene. | CRISPR-Cas9, homologous recombination. | Investigating a gene’s function in biological processes. | [32] |
| Gene overexpression | Increases gene expression to enhance product levels. | Promoters, introducing extra copies. | Boosting the synthesis of target products. | [33] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Jin, Z.; Liu, L.; Zhang, D. The Strategy and Application of Gene Attenuation in Metabolic Engineering. Microorganisms 2025, 13, 927. https://doi.org/10.3390/microorganisms13040927
Zhang Y, Jin Z, Liu L, Zhang D. The Strategy and Application of Gene Attenuation in Metabolic Engineering. Microorganisms. 2025; 13(4):927. https://doi.org/10.3390/microorganisms13040927
Chicago/Turabian StyleZhang, Yahui, Zhaoxia Jin, Linxia Liu, and Dawei Zhang. 2025. "The Strategy and Application of Gene Attenuation in Metabolic Engineering" Microorganisms 13, no. 4: 927. https://doi.org/10.3390/microorganisms13040927
APA StyleZhang, Y., Jin, Z., Liu, L., & Zhang, D. (2025). The Strategy and Application of Gene Attenuation in Metabolic Engineering. Microorganisms, 13(4), 927. https://doi.org/10.3390/microorganisms13040927