
Metagenomics Analysis of the Microbial Consortium in Samples from Lake Xochimilco, a World Cultural Heritage Site



Abstract
1. Introduction
2. Materials and Methods
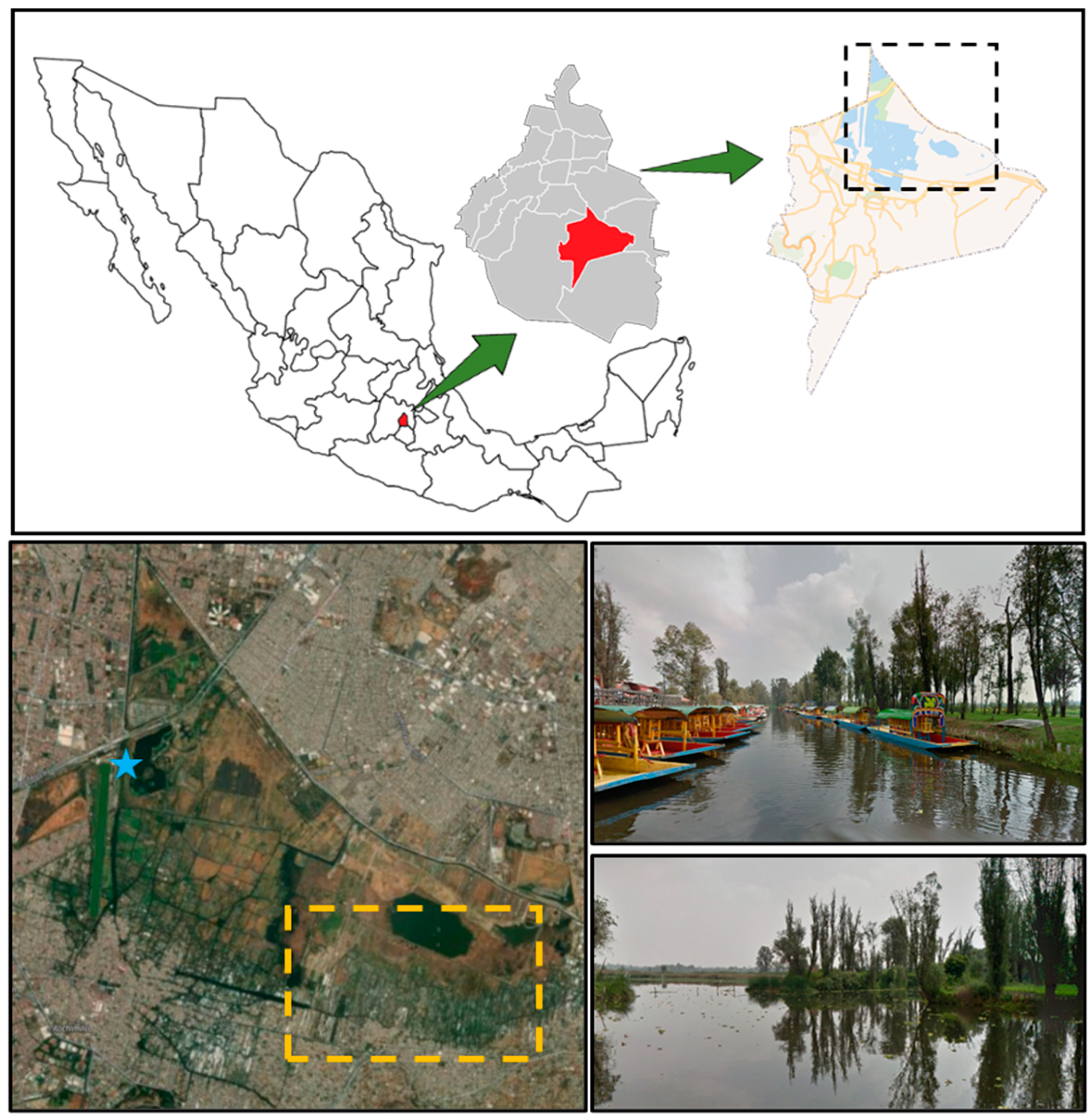
2.1. Sample Collection
2.2. Nucleic Acid Extraction
2.3. 16S rDNA Amplification and Data Analysis
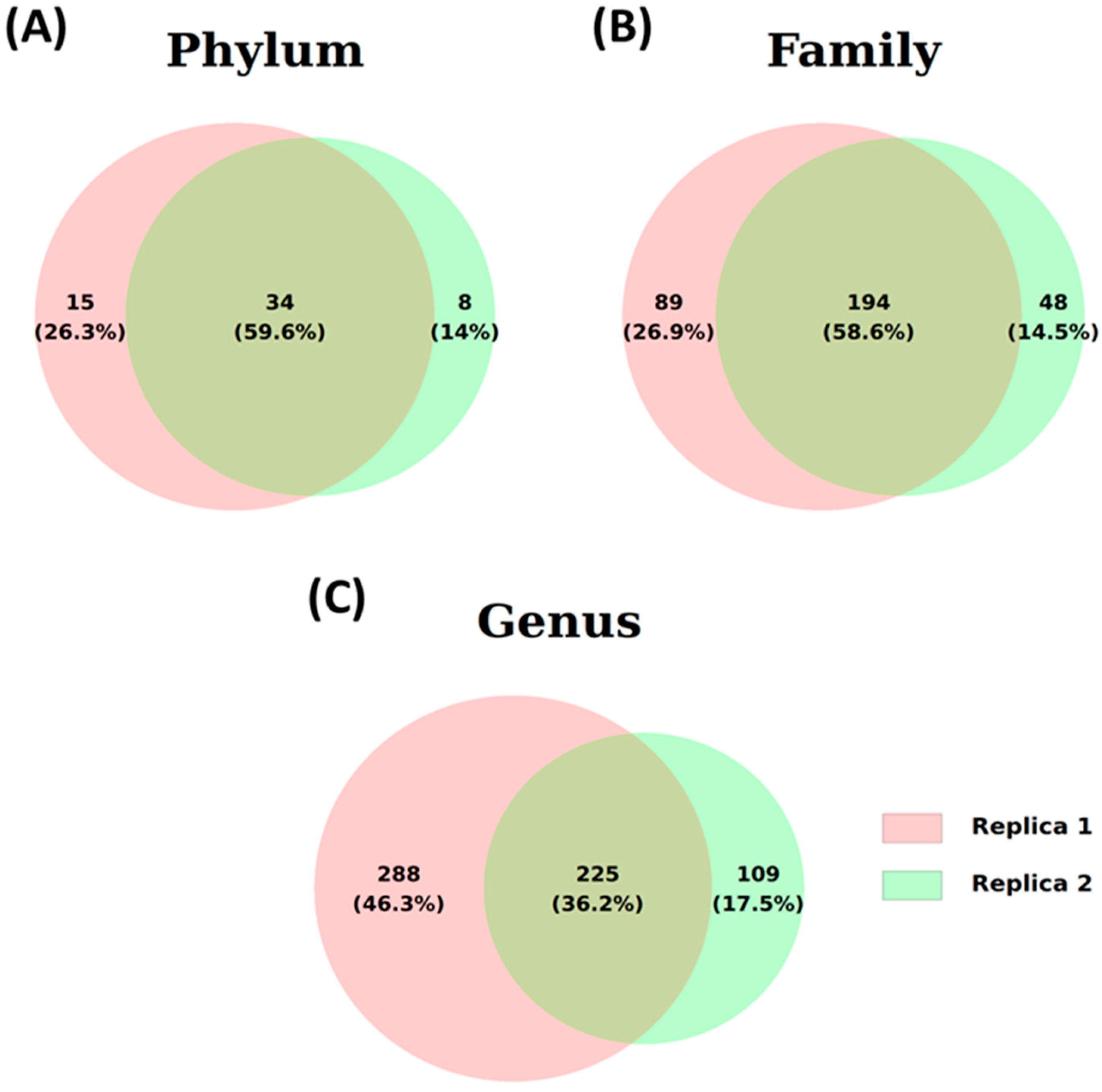
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ebel, R. Chinampas: An Urban Farming Model of the Aztecs and a Potential Solution for Modern Megalopolis. Hortte 2020, 30, 13–19. [Google Scholar] [CrossRef]
- Onofre, S.A. The floating gardens in México Xochimilco, world heritage risk site. City Time 2005, 1, 5. [Google Scholar]
- Rosas, I.; Báez, A.; Coutiño, M. Bacteriological quality of crops irrigated with wastewater in the Xochimilco plots, Mexico City, Mexico. Appl. Environ. Microbiol. 1984, 47, 1074–1079. [Google Scholar] [CrossRef]
- Pérez-Belmont, P.; Alvarado, J.; Vázquez-Salvador, N.; Rodríguez, E.; Valiente, E.; Díaz, J. Water quality monitoring in the Xochimilco peri-urban wetland: Experiences engaging in citizen science. Freshw. Sci. 2019, 38, 342–351. [Google Scholar] [CrossRef]
- Metch, J.W.; Burrows, N.D.; Murphy, C.J.; Pruden, A.; Vikesland, P.J. Metagenomic analysis of microbial communities yields insight into impacts of nanoparticle design. Nat. Nanotechnol. 2018, 13, 253–259. [Google Scholar] [CrossRef]
- Cuevas-Castilleja, J.; Martínez-Luz, A.; López-Arzate, M.A.; Ramírez-García, I.A.; Mora-Aguilera, G.; Ávila-Alistac, N. Perspectives of integrated pest management in CDMX urban agriculture and impacts of SARS-CoV-2 health emergency. Rev. Mex. Fitopatol. 2021, 39, 328–350. [Google Scholar] [CrossRef]
- Lima, P.T.; Sanchez, L.M.R.; García, B.I. Mexico City: The integration of urban agriculture to contain urban sprawl. In Growing Cities Growing Food: Urban Agriculture on the Policy Agenda; Academia: San Francisco, CA, USA, 2000; pp. 363–390. [Google Scholar]
- García-Gómez, R.; Chávez-Espinosa, J.; Mejía-Chávez, A.; Durán-de-Bazúa, C. Microbiological determinations of some vegetables from the Xochimilco zone in Mexico City, Mexico. Rev. Latinoam. Microbiol. 2002, 44, 24–30. [Google Scholar]
- Mazari-Hiriart, M.; Ponce-de-León, S.; López-Vidal, Y.; Islas-Macías, P.; Amieva-Fernández, R.I.; Quiñones-Falconi, F. Microbiological Implications of Periurban Agriculture and Water Reuse in Mexico City. PLoS ONE 2008, 3, e2305. [Google Scholar] [CrossRef]
- Embarcadero-Jiménez, S.; Rivera-Orduña, F.N.; Wang, E.T. Bacterial communities estimated by pyrosequencing in the soils of chinampa, a traditional sustainable agro-ecosystem in Mexico. J. Soils Sediments 2016, 16, 1001–1011. [Google Scholar] [CrossRef]
- Fang, W.; Fan, T.; Wang, S.; Yu, X.; Lu, A.; Wang, X.; Zhou, W.; Yuan, H.; Zhang, L. Seasonal changes driving shifts in microbial community assembly and species coexistence in an urban river. Sci. Total Environ. 2023, 905, 167027. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Villegas, P.; Ruiz-Cantisani, M.I.; Caratozzolo, P.; Lara-Prieto, V.; Ponce-López, R.; Martínez-Acosta, M.; Torres, A.; Sriraman, V.; Martínez-Ortiz, A.; Membrillo-Hernández, J. Preserving World Cultural Heritage: Social Justice and Sustainability Competencies via Socially-Oriented Interdisciplinary Education. J. Teach. Educ. Sustain. 2022, 24, 49–72. [Google Scholar] [CrossRef]
- Pérez-Belmont, P.; Lerner, A.M.; Mazari-Hiriart, M.; Valiente, E. The survival of agriculture on the edge: Perceptions of push and pull factors for the persistence of the ancient chinampas of Xochimilco, Mexico City. J. Rural Stud. 2021, 86, 452–462. [Google Scholar] [CrossRef]
- Carlsen, L.; Bruggemann, R. The 17 United Nations’ sustainable development goals: A status by 2020. Int. J. Sustain. Dev. World Ecol. 2022, 29, 219–229. [Google Scholar] [CrossRef]
- Illumina. 16S Metagenomic Sequencing Library Preparation. In Preparing 16S Ribosomal RNA Gene Amplicons for the Illumina MiSeq System; Illumina: San Diego, CA, USA, 2013; Volume 21, 28p. [Google Scholar]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef]
- Krueger, F.; James, F.; Ewels, P.; Afyounian, E.; Schuster-Boeckler, B. FelixKrueger/TrimGalore: v0.6.10, Zenodo. arXiv 2023, arXiv:1303.3997. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Pineda-Mendoza, R.M.; Briones-Roblero, C.I.; Gonzalez-Escobedo, R.; Rivera-Orduña, F.N.; Martínez-Jerónimo, F.; Zúñiga, G. Seasonal changes in the bacterial community structure of three eutrophicated urban lakes in Mexico city, with emphasis on Microcystis spp. Toxicon 2020, 179, 8–20. [Google Scholar] [CrossRef]
- Alotaibi, M.O.; Mohammed, A.E.; Eltom, K.H. Metagenomic analysis of bacterial communities of Wadi Namar Lake, Riyadh, Saudi Arabia. Saudi J. Biol. Sci. 2022, 29, 3749–3758. [Google Scholar] [CrossRef] [PubMed]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; Van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S. The phylogeny of proteobacteria: Relationships to other eubacterial phyla and eukaryotes. FEMS Microbiol. Rev. 2000, 24, 367–402. [Google Scholar] [CrossRef] [PubMed]
- Yanez-Montalvo, A.; Gómez-Acata, S.; Águila, B.; Hernández-Arana, H.; Falcón, L.I. The microbiome of modern microbialites in Bacalar Lagoon, Mexico. PLoS ONE 2020, 15, e0230071. [Google Scholar] [CrossRef]
- Kotzampassi, K.; Giamarellos-Bourboulis, E.J.; Stavrou, G. Obesity as a Consequence of Gut Bacteria and Diet Interactions. ISRN Obes. 2014, 2014, 651895. [Google Scholar]
- Tania Isaac, S. Bacterial Community Analysis of Seven Polluted Lakes in Bangalore, India: The Foam Stops Here, Tulane University. 2020. Available online: https://docslib.org/doc/4453776/bacterial-community-analysis-of-seven-polluted-lakes-in (accessed on 20 March 2025).
- Tinahones, F.J. La importancia de la microbiota en la obesidad. Rev. Española Endocrinol. Pediátrica 2017. [Google Scholar] [CrossRef]
- Brown, B.L.; LePrell, R.V.; Franklin, R.B.; Rivera, M.C.; Cabral, F.M.; Eaves, H.L.; Gardiakos, V.; Keegan, K.P.; King, T.L. Metagenomic analysis of planktonic microbial consortia from a non-tidal urban-impacted segment of James River. Stand. Genom. Sci. 2015, 10, 65. [Google Scholar] [CrossRef]
- Dastager, S.G.; Krishnamurthi, S.; Rameshkumar, N.; Dharne, M. The Family Micrococcaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 455–498. [Google Scholar] [CrossRef]
- García-Armisen, T.; İnceoğlu, Ö.; Ouattara, N.K.; Anzil, A.; Verbanck, M.A.; Brion, N.; Servais, P. Seasonal Variations and Resilience of Bacterial Communities in a Sewage Polluted Urban River. PLoS ONE 2014, 9, e92579. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Scheuner, C.; Göker, M.; Schumann, P. The Family Intrasporangiaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 397–424. [Google Scholar] [CrossRef]
- Shao, K.; Yao, X.; Wu, Z.; Jiang, X.; Hu, Y.; Tang, X.; Xu, Q.; Gao, G. The bacterial community composition and its environmental drivers in the rivers around eutrophic Chaohu Lake, China. BMC Microbiol. 2021, 21, 179. [Google Scholar] [CrossRef]
- Trujillo-Cabrera, Y.; Ponce-Mendoza, A.; Vásquez-Murrieta, M.S.; Rivera-Orduña, F.N.; Wang, E.T. Diverse cellulolytic bacteria isolated from the high humus, alkaline-saline chinampa soils. Ann. Microbiol. 2013, 63, 779–792. [Google Scholar] [CrossRef]
- Ting, A.S.Y.; Zoqratt, M.Z.H.M.; Tan, H.S.; Hermawan, A.A.; Talei, A.; Khu, S.T. Bacterial and eukaryotic microbial communities in urban water systems profiled via Illumina MiSeq platform. 3 Biotech 2021, 11, 40. [Google Scholar] [CrossRef]
- Lian, Y.; Zhen, L.; Chen, X.; Li, Y.; Li, X. Dynamics and Heterogeneity of Microbial Communities in Urban Water Environments. Res. Square 2020. [Google Scholar] [CrossRef]
- Beata, G. The use of -omics tools for assessing biodeterioration of cultural heritage: A review. J. Cult. Herit. 2020, 45, 351–361. [Google Scholar] [CrossRef]
- Piñar, G.; Sterflinger, K. Natural sciences at the service of art and cultural heritage: An interdisciplinary area in development and important challenges. Microb. Biotechnol. 2021, 14, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Marvasi, M.; Cavalieri, D.; Mastromei, G.; Casaccia, A.; Perito, B. Omics technologies for an in-depth investigation of biodeterioration of cultural heritage. Int. Biodeterior. Biodegrad. 2019, 144, 104736. [Google Scholar] [CrossRef]
- Lian, Y.; Zhen, L.; Chen, X.; Li, Y.; Li, X. Microbial biomarkers as indication of dynamic and heterogeneous urban water environments. Environ. Sci. Pollut. Res. 2022, 30, 107304–107316. [Google Scholar] [CrossRef]
- Zhao, D.; Huang, R.; Zeng, J.; Yan, W.; Wang, J.; Ma, T.; Wang, M.; Wu, Q.L. Diversity analysis of bacterial community compositions in sediments of urban lakes by terminal restriction fragment length polymorphism (T-RFLP). World J. Microbiol. Biotechnol. 2012, 28, 3159–3170. [Google Scholar] [CrossRef]
- Newton, R.J.; Jones, S.E.; Eiler, A.; McMahon, K.D.; Bertilsson, S. A Guide to the Natural History of Freshwater Lake Bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 14–49. [Google Scholar] [CrossRef]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Binda, C.; Lopetuso, L.R.; Rizzatti, G.; Gibiino, G.; Cennamo, V.; Gasbarrini, A. Actinobacteria: A relevant minority for the maintenance of gut homeostasis. Dig. Liver Dis. 2018, 50, 421–428. [Google Scholar] [CrossRef]
- Pan, X.; Raaijmakers, J.M.; Carrión, V.J. Importance of Bacteroidetes in host–microbe interactions and ecosystem functioning. Trends Microbiol. 2023, 31, 959–971. [Google Scholar] [CrossRef]
- Mehrshad, M.; Salcher, M.M.; Okazaki, Y.; Nakano, S.; Šimek, K.; Andrei, A.-S.; Ghai, R. Hidden in plain sight—Highly abundant and diverse planktonic freshwater Chloroflexi. Microbiome 2018, 6, 176. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample 1 | Sample 2 | ||||
|---|---|---|---|---|---|
| Phylum | Prevalence | Abundance (%) | Phylum | Prevalence | Abundance (%) |
| Actinobacteria | 41,983 | 42.69 | Actinobacteria | 27,381 | 46.68 |
| Proteobacteria | 18,919 | 19.24 | Proteobacteria | 15,224 | 25.96 |
| Firmicutes | 16,750 | 17.03 | Bacteroidota | 4467 | 7.62 |
| Chloroflexi | 4755 | 4.84 | Chloroflexi | 2586 | 4.41 |
| Bacteroidota | 4430 | 4.50 | Firmicutes | 2093 | 3.57 |
| Patescibacteria | 2114 | 2.15 | Fusobacteriota | 1286 | 2.19 |
| Cyanobacteria | 2006 | 2.04 | Patescibacteria | 1186 | 2.02 |
| Plantomycetota | 1164 | 1.18 | Cyanobacteria | 1011 | 1.72 |
| Gemmatimonadota | 956 | 0.97 | Desulfobacterota | 655 | 1.12 |
| Verrucomicrobiota | 807 | 0.82 | Verrucomicrobiota | 566 | 0.97 |
| Acidobacteriota | 692 | 0.70 | Spirochaetota | 452 | 0.77 |
| Desulfobacterota | 648 | 0.66 | Deinococcota | 422 | 0.72 |
| Myxococcota | 648 | 0.66 | Acidobacteriota | 249 | 0.42 |
| Spirochaetota | 623 | 0.63 | Fibrobacterota | 197 | 0.34 |
| Bdellovibrionota | 435 | 0.44 | Plantomycetota | 176 | 0.30 |
| Deinococcota | 326 | 0.33 | Campylobacterota | 153 | 0.26 |
| Fusobacteriota | 227 | 0.23 | Myxococcota | 109 | 0.19 |
| Armatimonadota | 80 | 0.08 | Halobacterota | 78 | 0.13 |
| Calditrichota | 75 | 0.08 | Nanoarchaeota | 39 | 0.07 |
| Unassigned | 25 | 0.03 | Unassigned | 27 | 0.05 |
| Sample 1 | Sample 2 | ||||
|---|---|---|---|---|---|
| Family | Prevalence | Abundance (%) | Family | Prevalence | Abundance (%) |
| Micrococcaceae | 12,505 | 12.72 | Micrococcaceae | 18,635 | 31.77 |
| Intrasporangiaceae | 11,322 | 11.51 | Rhodobacteraceae | 4608 | 7.86 |
| Bacillaceae | 6673 | 6.79 | Intrasporangiaceae | 3408 | 5.81 |
| Nocardioidaceae | 5006 | 5.09 | Azospirillaceae | 1602 | 2.73 |
| Rhodobacteraceae | 3742 | 3.81 | Rhodocyclaceae | 1480 | 2.52 |
| Planococcaceae | 3182 | 3.24 | Acetobacteraceae | 1372 | 2.34 |
| Geodermatophilaceae | 2758 | 2.80 | Leptotrichiaceae | 1286 | 2.19 |
| JG30-KF-CM45 | 2113 | 2.15 | JG30-KF-CM45 | 980 | 1.67 |
| Flavobacteriaceae | 1642 | 1.67 | Anaerolineaceae | 978 | 1.67 |
| Sphingomonadaceae | 1592 | 1.62 | Marinilabiliaceae | 799 | 1.36 |
| Cellulomonadaceae | 1495 | 1.52 | Propionibacteriaceae | 785 | 1.34 |
| Rhizobiaceae | 1103 | 1.12 | Sphingomonadaceae | 739 | 1.26 |
| Comamonadaceae | 1098 | 1.12 | Geodermatophilaceae | 707 | 1.21 |
| Clostridiaceae | 965 | 0.98 | Acidothiobacillaceae | 688 | 1.17 |
| Rhodocyclaceae | 942 | 0.96 | Prolixibacteraceae | 639 | 1.09 |
| Paenibacillaceae | 923 | 0.94 | Comamonadaceae | 455 | 0.78 |
| Xanthomonadaceae | 798 | 0.81 | Bacteroidota vadinHA17 | 441 | 0.75 |
| Micromonosporaceae | 795 | 0.81 | Dermabacteraceae | 440 | 0.75 |
| Anaerolineaceae | 792 | 0.81 | Aeromonadaceae | 435 | 0.74 |
| Unassigned | 10,782 | 10.96 | Unassigned | 5758 | 9.82 |
| Sample 1 | Sample 2 | ||||
|---|---|---|---|---|---|
| Genus | Prevalence | Abundance (%) | Genus | Prevalence | Abundance (%) |
| Kocuria * | 10,627 | 10.81 | Kocuria * | 18,464 | 31.48 |
| Ornithinimicrobium | 5597 | 5.69 | Paracoccus | 4181 | 7.13 |
| Bacillus | 3995 | 4.06 | Arsenicicoccus | 3080 | 5.25 |
| Nocardioides | 3017 | 3.07 | Skermanella | 1602 | 2.73 |
| Serinicoccus | 2388 | 2.43 | Roseomonas | 902 | 1.54 |
| Blastococcus * | 2227 | 2.26 | [Cytophaga] xylanolytica group | 710 | 1.21 |
| Paracoccus | 1999 | 2.03 | KCM-B-112 | 688 | 1.17 |
| Marmoricola | 1603 | 1.63 | Blastococcus * | 594 | 1.01 |
| Ornithinicoccus | 1394 | 1.42 | Luteococcus | 579 | 0.99 |
| Cellulomonas | 1105 | 1.12 | Dechloromonas | 472 | 0.80 |
| Skermanella | 779 | 0.79 | Brachybacterium | 438 | 0.75 |
| Planococcus | 718 | 0.73 | Deinococcus | 407 | 0.69 |
| Antarcticibacterium | 713 | 0.73 | Aeromonas | 391 | 0.67 |
| Pseudarthrobacter | 707 | 0.72 | Belnapia | 374 | 0.64 |
| Amaricoccus | 698 | 0.71 | Sphingomonas * | 374 | 0.64 |
| Sporosarcina | 646 | 0.66 | Paludibacter | 336 | 0.57 |
| Clostridium sensu stricto 1 | 624 | 0.63 | Chitinivorax | 326 | 0.56 |
| Sphingomonas * | 624 | 0.63 | Thauera | 294 | 0.50 |
| Paenibacillus | 597 | 0.61 | Rubellimicrobium | 288 | 0.49 |
| Unassigned | 25,534 | 25.97 | Unassigned | 12,232 | 20.86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Obeso Fernández del Valle, A.; Membrillo-Hernández, J. Metagenomics Analysis of the Microbial Consortium in Samples from Lake Xochimilco, a World Cultural Heritage Site. Microorganisms 2025, 13, 835. https://doi.org/10.3390/microorganisms13040835
de Obeso Fernández del Valle A, Membrillo-Hernández J. Metagenomics Analysis of the Microbial Consortium in Samples from Lake Xochimilco, a World Cultural Heritage Site. Microorganisms. 2025; 13(4):835. https://doi.org/10.3390/microorganisms13040835
Chicago/Turabian Stylede Obeso Fernández del Valle, Alvaro, and Jorge Membrillo-Hernández. 2025. "Metagenomics Analysis of the Microbial Consortium in Samples from Lake Xochimilco, a World Cultural Heritage Site" Microorganisms 13, no. 4: 835. https://doi.org/10.3390/microorganisms13040835
APA Stylede Obeso Fernández del Valle, A., & Membrillo-Hernández, J. (2025). Metagenomics Analysis of the Microbial Consortium in Samples from Lake Xochimilco, a World Cultural Heritage Site. Microorganisms, 13(4), 835. https://doi.org/10.3390/microorganisms13040835