

Amebicidal Activity of Escherichia coli Nissle 1917 Against Entamoeba histolytica

 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. E. histolytica Strain
2.2. Escherichia coli Nissle 1917
2.3. In Vitro Association
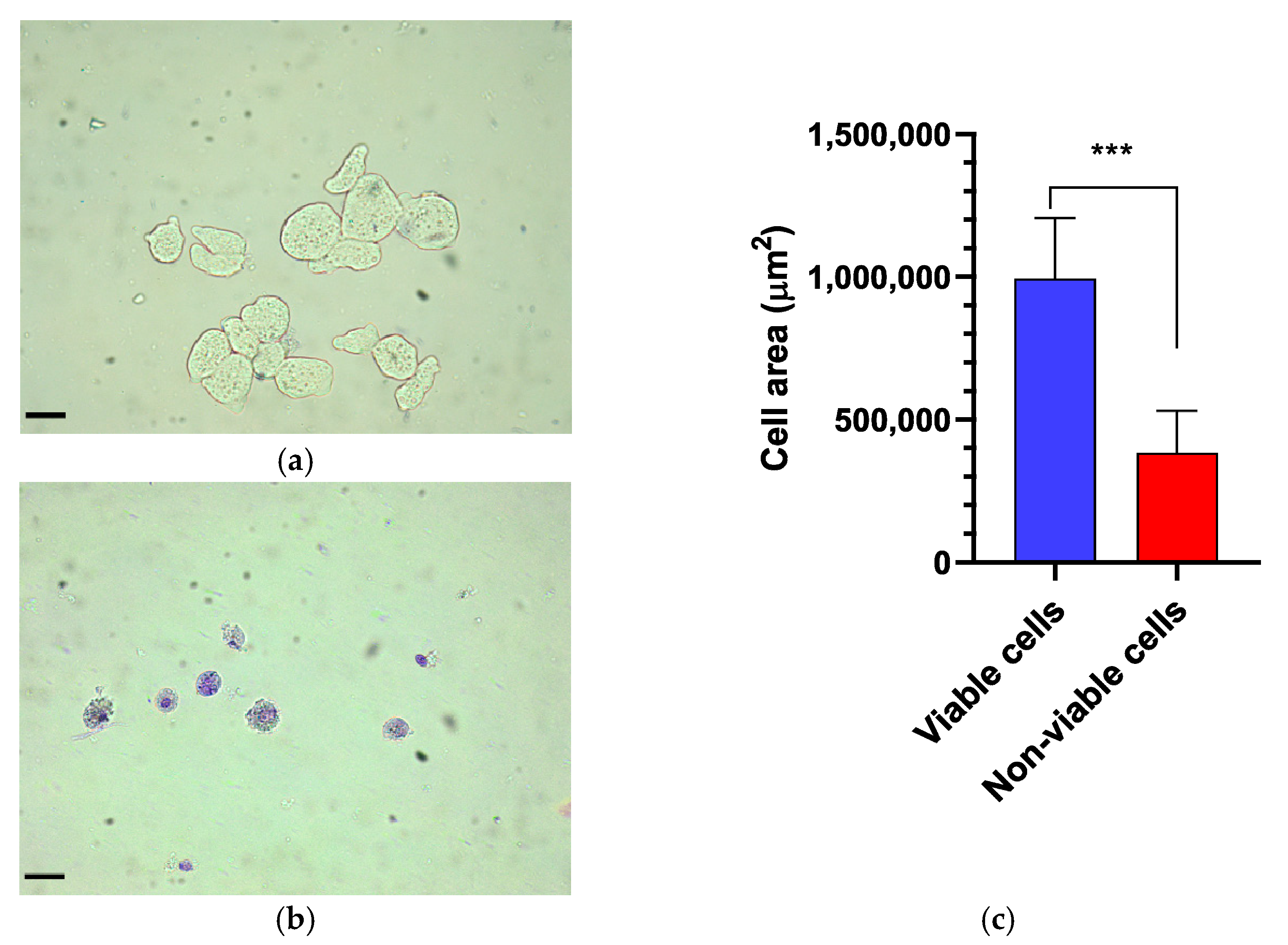
2.4. Morphometric Analysis of the Trophozoites
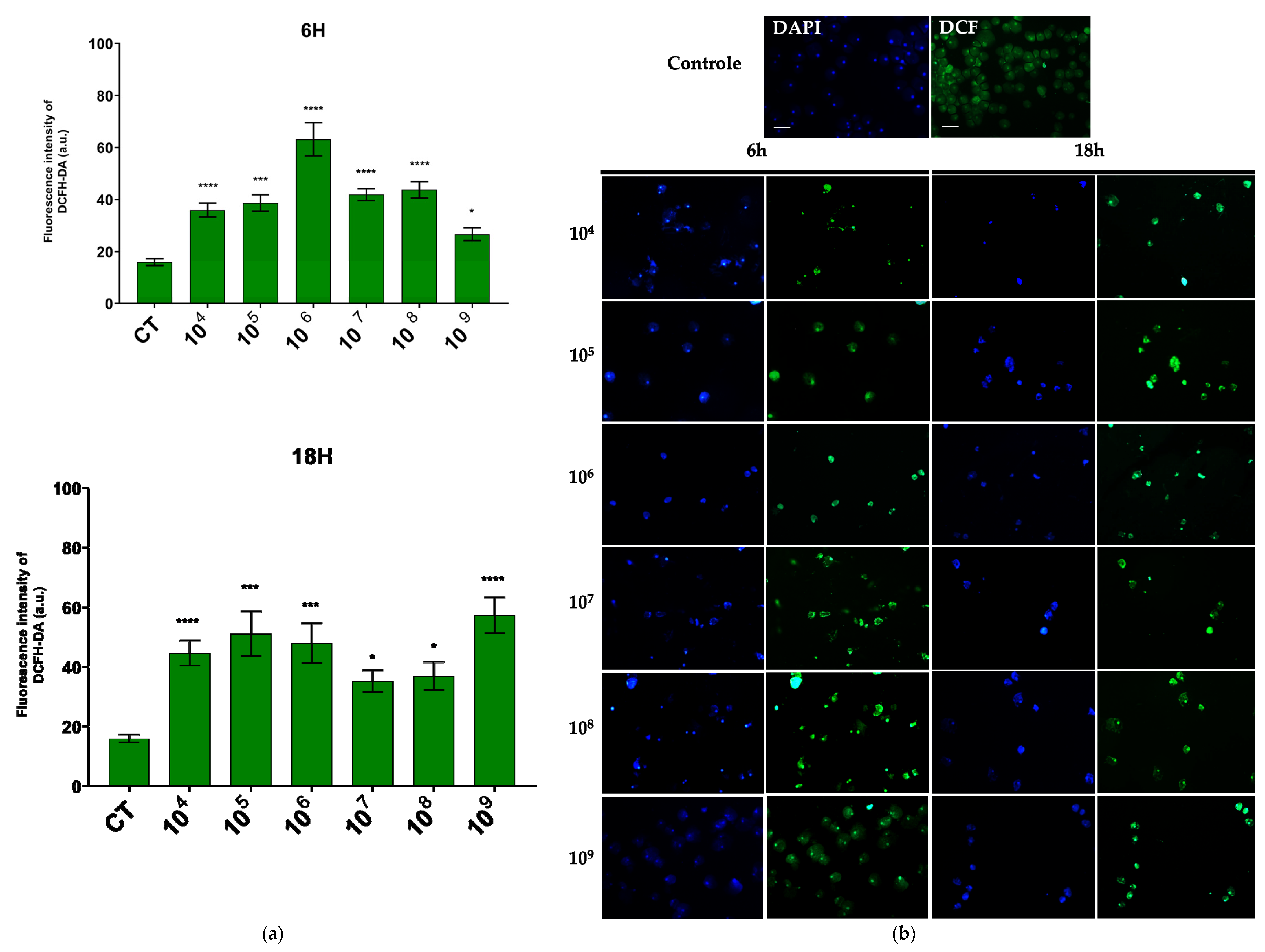
2.5. Determination of Reactive Oxygen Species
2.6. Fluorescence
2.7. Statistical Analysis
3. Results
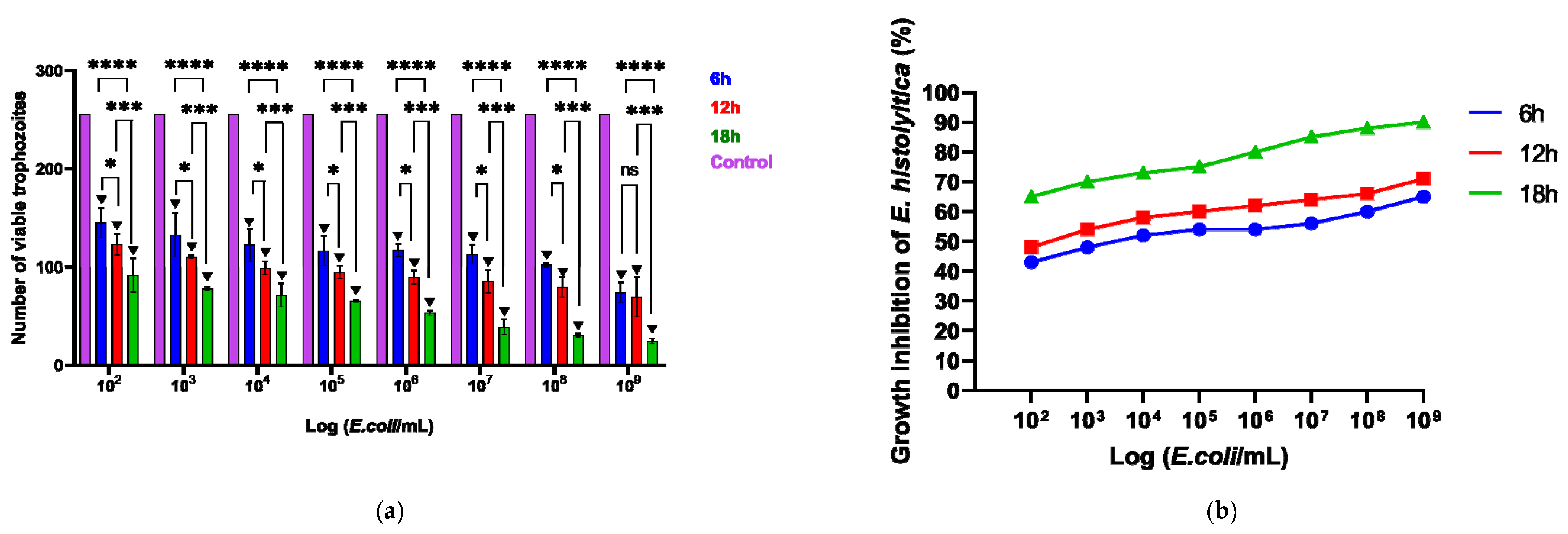
3.1. Effect of Probiotic EcN on E. histolytica Viability
3.2. ROS Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barlaam, A.; Sannella, A.R.; Ferrari, N.; Temesgen, T.T.; Rinaldi, L.; Normanno, G.; Cacciò, S.M.; Robertson, L.J.; Giangaspero, A. Ready-to-eat salads and berry fruits purchased in Italy contaminated by Cryptosporidium spp., Giardia duodenalis, and Entamoeba histolytica. Int. J. Food Microbiol. 2022, 370, 109634. [Google Scholar] [CrossRef]
- Saleh, F.E.R.; Gad, M.A.; Ashour, A.A.; Soliman, M.I.; El-Senousy, W.M.; Al-Herrawy, A.Z. Molecular detection of Entamoeba histolytica in fresh vegetables and irrigation. Egypt. J. Aquat. Biol. Fish. 2019, 22, 551–561. [Google Scholar] [CrossRef]
- Yanagawa, Y.; Singh, U. Diversity and Plasticity of Virulent Characteristics of Entamoeba histolytica. Trop. Med. Infect. Dis. 2023, 8, 255. [Google Scholar] [CrossRef] [PubMed]
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Shirley, D.-A.T.; Farr, L.; Watanabe, K.; Moonah, S. A Review of the Global Burden, New Diagnostics, and Current Therapeutics for Amebiasis. Open Forum Infect. Dis. 2018, 5, ofy161. [Google Scholar] [CrossRef]
- Nasrallah, J.; Akhoundi, M.; Haouchine, D.; Marteau, A.; Mantelet, S.; Wind, P.; Benamouzing, R.; Bouchaud, R.; Dhote, R.; Izri, A. Updates on the worldwide burden of amoebiasis: A case series and literature review. J. Infect. Public Health 2022, 15, 1134–1141. [Google Scholar] [CrossRef]
- Ximénez, C.; Morán, P.; Rojas, L.; Valadez, A.; Gómez, A. Reassessment of the epidemiology of amebiasis: State of the art. Infect. Genet. Evol. 2009, 9, 1023–1032. [Google Scholar] [CrossRef]
- Kumanan, T.; Sujanitha, V.; Sri Ranganathan, S. Metronidazole for Amoebiasis: A tale of more than half a century. Jaffna Med. J. 2021, 33, 6–13. [Google Scholar] [CrossRef]
- Van de Wijgert, J.H.H.M.; Verwijs, M.C. Lactobacilli-containing vaginal probiotics to cure or prevent bacterial or fungal vaginal dysbiosis: A systematic review and recommendations for future trial designs. BJOG Int. J. Obstet. Gynaecol. 2020, 127, 287–299. [Google Scholar] [CrossRef]
- Iqbal, Z.; Ahmed, S.; Tabassum, N.; Bhattacharya, R.; Bose, D. Role of probiotics in prevention and treatment of enteric infections: A comprehensive review. 3 Biotech 2021, 11, 242. [Google Scholar] [CrossRef]
- Lopez-Santamarina, A.; Gonzalez, E.G.; Lamas, A.; Mondragon, A.C.; Regal, P.; Miranda, J.M. Probiotics as a possible strategy for the prevention and treatment of allergies. A narrative review. Foods 2021, 10, 701. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Zhou, T.; Tang, H.; Feng, P.; Ali, G.; Liu, P.; Li, X. Genetically encoded probiotic EcN 1917 alleviates alcohol-induced acute liver injury and restore gut microbiota homeostasis. J. Funct. Foods 2021, 85, 104661. [Google Scholar] [CrossRef]
- Michael, H.; Miyazaki, A.; Langel, S.N.; Amimo, J.O.; Kick, M.K.; Chepngeno, J.; Paim, F.C.; Fischer, D.D.; Rajashekara, G.; Saif, L.J.; et al. Escherichia coli Nissle 1917 Enhances Efficacy of Oral Attenuated Human Rotavirus Vaccine in a Gnotobiotic Piglet Model. Vaccines 2022, 10, 83. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Hao, L.; Liu, X.; Borrás-Hidalgo, O.; Zhang, Y. Enhanced anti-proliferative efficacy of epothilone B loaded with Escherichia coli Nissle 1917 bacterial ghosts on the HeLa cells by mitochondrial pathway of apoptosis. Drug Dev. Ind. Pharm. 2018, 44, 1328–1335. [Google Scholar] [CrossRef]
- Blum, G.; Marre, R.; Hacker, J. Properties of Escherichia coli Strains of Serotype 06. Infection 1994, 23, 234–236. [Google Scholar] [CrossRef]
- Kruis, W. Maintaining remission of ulcerative colitis with the probiotic Escherichia coli Nissle 1917 is as effective as with standard mesalazine. Gut 2004, 53, 1617–1623. [Google Scholar] [CrossRef] [PubMed]
- Sonnenborn, U. Escherichia coli strain Nissle 1917—From bench to bedside and back: History of a special Escherichia coli strain with probiotic properties. FEMS Microbiol. Lett. 2016, 363, fnw212. [Google Scholar] [CrossRef]
- Storm, D.W.; Koff, S.A.; Horvath, D.J.; Li, B.; Justice, S.S. In Vitro Analysis of the Bactericidal Activity of Escherichia Coli Nissle 1917 Against Pediatric Uropathogens. J. Urol. 2011, 186, 1678–1683. [Google Scholar] [CrossRef]
- Boudeau, J.; Glasser, A.L.; Julien, S.; Colombel, J.F.; Darfeuille-Michaud, A. Inhibitory effect of probiotic Escherichia coli strain Nissle 1917 on adhesion to and invasion of intestinal epithelial cells by adherent–invasive E. coli strains isolated from patients with Crohn’s disease. Aliment. Pharmacol. Ther. 2003, 18, 45–56. [Google Scholar] [CrossRef]
- Gomes, M.A.; Melo, M.N.; Macedo, A.M.; Pena, G.P.M.; Caliari, M.V.; Silva, E.F. Characterization of Entamoeba histolytica and Entamoeba dispar by Biological, Biochemical, and Molecular Parameters. Arch. Med. Res. 2000, 31, S249–S250. [Google Scholar] [CrossRef]
- Clark, C.G.; Diamond, L.S. Methods for Cultivation of Luminal Parasitic Protists of Clinical Importance. Clin. Microbiol. Rev. 2002, 15, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Edgington, L.V. Fungitoxic Spectrum of Benzimidazole Compounds. Phytopathology 1971, 61, 42–44. [Google Scholar] [CrossRef]
- Mansour-Ghanaei, F.; Dehbashi, N.; Yazdanparast, K.; Shafaghi, A. Efficacy of saccharomyces boulardii with antibiotics in acute amoebiasis. World J. Gastroenterol. 2003, 9, 1832–1833. [Google Scholar] [CrossRef] [PubMed]
- Rigothier, M.C.; Maccario, J.; Gayral, P. Inhibitory activity of saccharomyces yeasts on the adhesion ofEntamoeba histolytica trophozoites to human erythrocytes in vitro. Parasitol. Res. 1994, 80, 10–15. [Google Scholar] [CrossRef]
- Sarid, L.; Zanditenas, E.; Ye, J.; Trebicz-Geffen, M.; Ankri, S. Insights into the Mechanisms of Lactobacillus acidophilus Activity against Entamoeba histolytica by Using Thiol Redox Proteomics. Antioxidants 2022, 11, 814. [Google Scholar] [CrossRef]
- Varet, H.; Shaulov, Y.; Sismeiro, O.; Trebicz-Geffen, M.; Legendre, R.; Coppée, J.-Y.; Ankri, S.; Guillen, N. Enteric bacteria boost defences against oxidative stress in Entamoeba histolytica. Sci. Rep. 2018, 8, 9042. [Google Scholar] [CrossRef]
- Sarjapuram, N.; Mekala, N.; Singh, M.; Tatu, U. The Potential of Lactobacillus casei and Entercoccus faecium Combination as a Preventive Probiotic Against Entamoeba. Probiotics Antimicrob. Proteins 2017, 9, 142–149. [Google Scholar] [CrossRef]
- Prado, G.K.S.; Torrinha, K.C.; Cruz, R.E.; Gonçalves, A.B.B.; Silva, C.A.V.; Oliveira, F.M.S.; Nunes, A.C.; Gomes, M.A.; Caliari, M.V. Weissella paramesenteroides WpK4 ameliorate the experimental amoebic colitis by increasing the expression of MUC-2 and the intestinal epithelial regeneration. J. Appl. Microbiol. 2020, 129, 1706–1719. [Google Scholar] [CrossRef]
- Van Baarlen, P.; Wells, J.M.; Kleerebezem, M. Regulation of intestinal homeostasis and immunity with probiotic lactobacilli. Trends Immunol. 2013, 34, 208–215. [Google Scholar] [CrossRef]
- Martens, K.; Pugin, B.; De Boeck, I.; Spacova, I.; Steelant, B.; Seys, S.F.; Lebeer, S.; Hellings, P.W. Probiotics for the airways: Potential to improve epithelial and immune homeostasis. Allergy 2018, 73, 1954–1963. [Google Scholar] [CrossRef]
- Shini, S.; Bryden, W.L. Probiotics and gut health: Linking gut homeostasis and poultry productivity. Anim. Prod. Sci. 2021, 62, 1090–1112. [Google Scholar] [CrossRef]
- Bai, J.C.; Ciacci, C. World Gastroenterology Organisation Global Guidelines: Celiac Disease February 2017. J. Clin. Gastroenterol. 2017, 51, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Shahi, P.; Trebicz-Geffen, M.; Nagaraja, S.; Alterzon-Baumel, S.; Hertz, R.; Methling, K.; Lalk, M.; Ankri, S. Proteomic Identification of Oxidized Proteins in Entamoeba histolytica by Resin-Assisted Capture: Insights into the Role of Arginase in Resistance to Oxidative Stress. PLoS Neglected Trop. Dis. 2016, 10, e0004340. [Google Scholar] [CrossRef]
- Collins, E.B.; Aramaki, K. Production of Hydrogen Peroxide by Lactobacillus acidophilus. J. Dairy Sci. 1980, 63, 353–357. [Google Scholar] [CrossRef]
- Hering, N.A.; Richter, J.F.; Fromm, A.; Wieser, A.; Hartmann, S.; Günzel, D.; Bucker, R.; Fromm, M.; Schulzke, J.D.; Troeger, H. TcpC protein from E. coli Nissle improves epithelial barrier function involving PKCζ and ERK1/2 signaling in HT-29/B6 cells. Mucosal Immunol. 2014, 7, 369–378. [Google Scholar] [CrossRef]
- Zyrek, A.A.; Cichon, C.; Helms, S.; Enders, C.; Sonnenborn, U.; Schmidt, M.A. Molecular mechanisms underlying the probiotic effects of Escherichia coli Nissle 1917 involve ZO-2 and PKC? redistribution resulting in tight junction and epithelial barrier repair. Cell. Microbiol. 2007, 9, 804–816. [Google Scholar] [CrossRef] [PubMed]
- Ukena, S.N.; Singh, A.; Dringenberg, U.; Engelhardt, R.; Seidler, U.; Hansen, W.; Bleich, A.; Bruder, D.; Franzke, A.; Rogler, G.; et al. Probiotic Escherichia coli Nissle 1917 Inhibits Leaky Gut by Enhancing Mucosal Integrity. PLoS ONE 2007, 2, e1308. [Google Scholar] [CrossRef]
- Schlee, M.; Wehkamp, J.; Altenhoefer, A.; Oelschlaeger, T.A.; Stange, E.F.; Fellermann, K. Induction of Human β-Defensin 2 by the Probiotic Escherichia coli Nissle 1917 Is Mediated through Flagellin. Infect. Immun. 2007, 75, 2399–2407. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moura-Oliveira, V.; Oliveira, F.M.S.; Moreno, O.L.M.; Ferreira, J.R.; Szawka, R.E.; Campideli-Santana, A.C.; Teles, J.; Capettini, L.S.A.; Martins, F.S.; Gomes, M.A. Amebicidal Activity of Escherichia coli Nissle 1917 Against Entamoeba histolytica. Microorganisms 2025, 13, 828. https://doi.org/10.3390/microorganisms13040828
Moura-Oliveira V, Oliveira FMS, Moreno OLM, Ferreira JR, Szawka RE, Campideli-Santana AC, Teles J, Capettini LSA, Martins FS, Gomes MA. Amebicidal Activity of Escherichia coli Nissle 1917 Against Entamoeba histolytica. Microorganisms. 2025; 13(4):828. https://doi.org/10.3390/microorganisms13040828
Chicago/Turabian StyleMoura-Oliveira, Vivian, Fabrício M. S. Oliveira, Olga L. M. Moreno, Julia R. Ferreira, Raphael E. Szawka, Ana C. Campideli-Santana, Jullia Teles, Luciano S. A. Capettini, Flaviano S. Martins, and Maria A. Gomes. 2025. "Amebicidal Activity of Escherichia coli Nissle 1917 Against Entamoeba histolytica" Microorganisms 13, no. 4: 828. https://doi.org/10.3390/microorganisms13040828
APA StyleMoura-Oliveira, V., Oliveira, F. M. S., Moreno, O. L. M., Ferreira, J. R., Szawka, R. E., Campideli-Santana, A. C., Teles, J., Capettini, L. S. A., Martins, F. S., & Gomes, M. A. (2025). Amebicidal Activity of Escherichia coli Nissle 1917 Against Entamoeba histolytica. Microorganisms, 13(4), 828. https://doi.org/10.3390/microorganisms13040828