
Development of an RPA-CRISPR/Cas12a-Based Rapid Diagnosis Strip for the Tangerine Pathotype of Alternaria alternata


Abstract
1. Introduction
2. Materials and Methods
2.1. The Source of Fungal Strains Used in This Study
2.2. Detection Target Screening, RPA Primer Design, and crRNA Design
2.3. Establishment of the Reaction System
2.4. Optimization of the RPA-CRISPR/Cas12a Detection System
2.5. Specificity, Sensitivity, and Applications Assay for RPA-CRISPR/Cas12a with the Test Strip Detection
3. Results
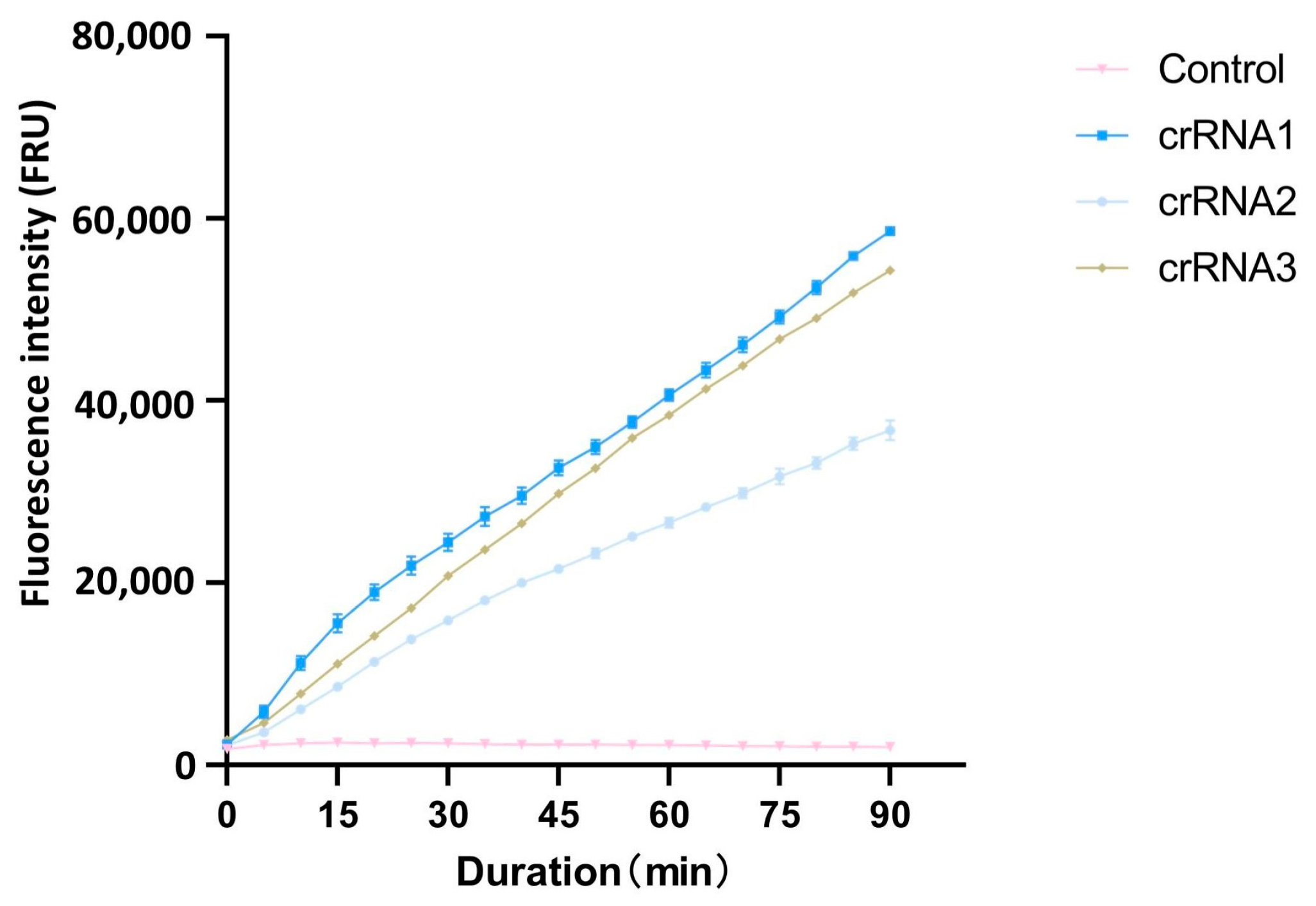
3.1. crRNA Screening and Cutting Efficiency
3.2. Optimization of Reaction Time and Temperature for RPA
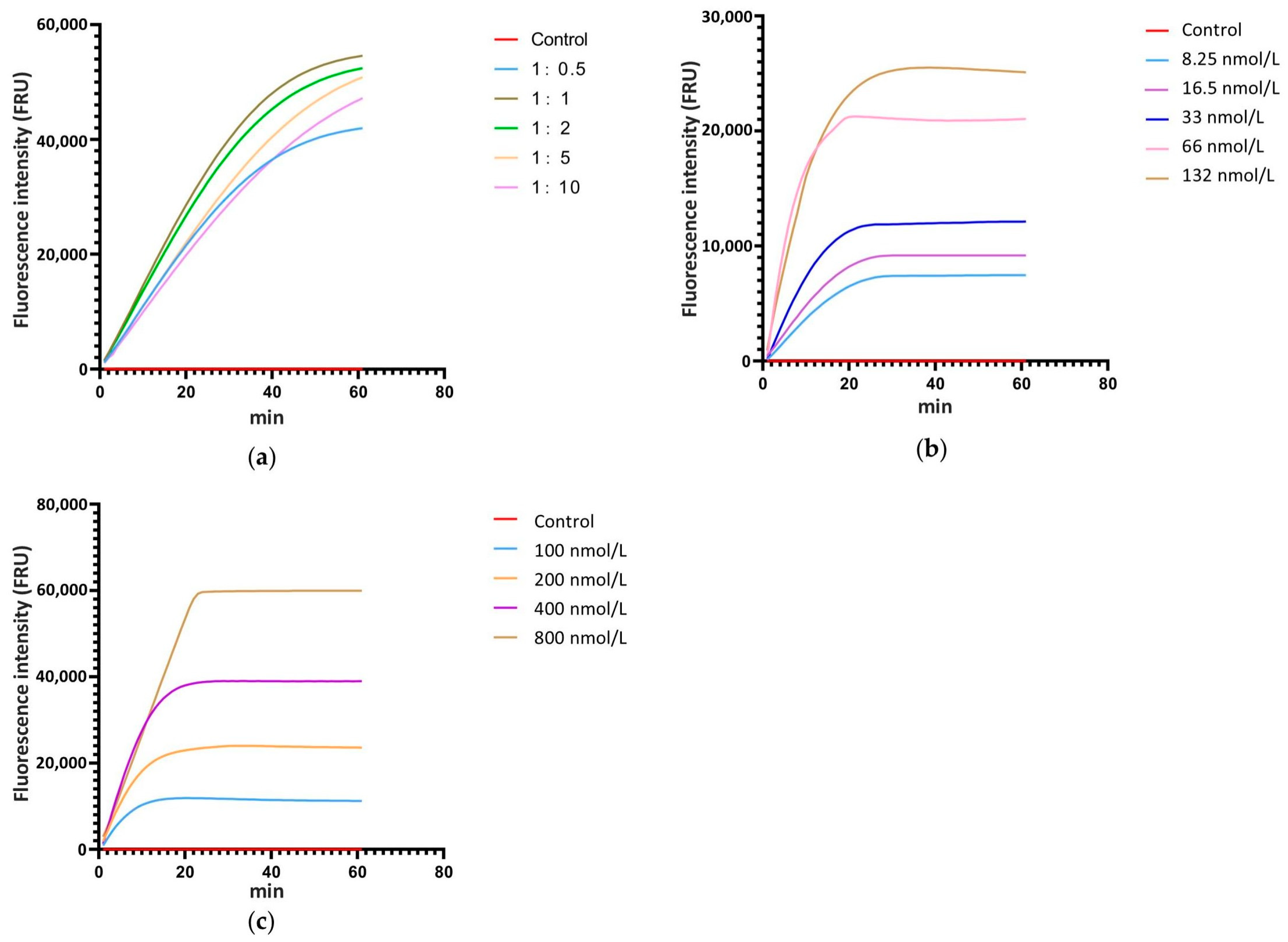
3.3. Optimization of the Ratio of Cas12a to crRNA, the Concentration of Cas12a and crRNA, and the Concentration of ssDNA in the CRISPR/Cas12a Reaction System
3.4. Screening of Reaction Time for CRISPR/Cas12a with the Test Strip Detection
3.5. Specificity and Sensitivity Testing of the RPA-CRISPR/Cas12a Test Strip Detection System
3.6. Applications Assay for RPA-CRISPR/Cas12a with the Test Strip Detection
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Y.X.; Dong, X.F.; Yang, A.L.; Zhang, H.B. Diversity and pathogenicity of Alternaria species associated with the invasive plant Ageratina adenophora and local plants. PeerJ 2022, 10, e13012. [Google Scholar] [CrossRef]
- Fu, H.; Chung, K.R.; Liu, X.; Li, H. Aaprb1, a subtilsin-like protease, required for autophagy and virulence of the tangerine pathotype of Alternaria alternata. Microbiol. Res. 2020, 240, 126537. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Fu, H.; Shen, X.X.; Ruan, R.; Rokas, A.; Li, H. Genomic features and evolution of the conditionally dispensable chromosome in the tangerine pathotype of Alternaria alternata. Mol. Plant Pathol. 2019, 20, 1425–1438. [Google Scholar] [CrossRef]
- Huang, F.; Fu, Y.; Nie, D.; Stewart, J.E.; Peever, T.L.; Li, H. Identification of a novel phylogenetic lineage of Alternaria alternata causing citrus brown spot in China. Fungal. Biol. 2015, 119, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Meena, M.; Samal, S. Alternaria host-specific (HSTs) toxins: An overview of chemical characterization, target sites, regulation and their toxic effects. Toxicol. Rep. 2019, 6, 745–758. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, Y.; Masunaka, A.; Tsuge, T.; Yamamoto, M.; Ohtani, K.; Fukumoto, T.; Gomi, K.; Peever, T.L.; Tada, Y.; Ichimura, K.; et al. ACTTS3 encoding a polyketide synthase is essential for the biosynthesis of ACT-toxin and pathogenicity in the tangerine pathotype of Alternaria alternata. Mol. Plant Microbe Interact. 2010, 23, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.D.; Johnson, L.; Kohomoto, K.; Otani, H.; Lane, C.R.; Kodama, M. A polymerase chain reaction-based method to specifically detect Alternaria alternata apple pathotype (A. mali), the causal agent of Alternaria blotch of apple. Phytopathology 2000, 90, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Moghimi, H.; Moradi, A.; Hamedi, J.; Basiri, M. Development of a loop-mediated isothermal amplification assay for rapid and specific identification of ACT producing Alternaria alternata, the agent of brown spot disease in tangerine. Appl. Biochem. Biotechnol. 2016, 178, 1207–1219. [Google Scholar] [CrossRef] [PubMed]
- Lobato, I.M.; O’Sullivan, C.K. Recombinase polymerase amplification: Basics, applications and recent advances. Trac-Trend Anal. Chem. 2018, 98, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Li, C.; Lan, H.; Pan, D.; Wu, Y. Comparison of four isothermal amplification techniques: LAMP, SEA, CPA, and RPA for the identification of chicken adulteration. Food Control 2024, 159, 110302. [Google Scholar] [CrossRef]
- Tan, M.; Liao, C.; Liang, L.; Yi, X.; Zhou, Z.; Wei, G. Recent advances in recombinase polymerase amplification: Principle, advantages, disadvantages and applications. Front. Cell Infect. Microbiol. 2022, 12, 1019071. [Google Scholar] [CrossRef]
- Chen, J.S.; Ma, E.; Harrington, L.B.; Da Costa, M.; Tian, X.; Palefsky, J.M.; Doudna, J.A. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science 2018, 360, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, R.; Wang, D.; Wu, J.; Li, J.; Wang, J.; Liu, H.; Wang, Y. Cas12aVDet: A CRISPR/Cas12a-based platform for rapid and visual nucleic acid detection. Anal. Chem. 2019, 91, 12156–12161. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, H.; Salehiabar, M.; Barsbay, M.; Ghaffarlou, M.; Kavetskyy, T.; Sharafi, A.; Davaran, S.; Chauhan, S.C.; Danafar, H.; Kaboli, S.; et al. CRISPR systems for COVID-19 diagnosis. ACS Sens. 2021, 6, 1430–1445. [Google Scholar] [CrossRef] [PubMed]
- Aslam, S.; Tahir, A.; Aslam, M.F.; Alam, M.W.; Shedayi, A.A.; Sadia, S. Recent advances in molecular techniques for the identification of phytopathogenic fungi–a mini review. J. Plant Interact. 2017, 12, 493–504. [Google Scholar] [CrossRef]
- Möller, E.M.; Bahnweg, G.; Sandermann, H.; Geiger, H.H. A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tissues. Nucleic Acids. Res. 1992, 20, 6115–6116. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Ding, Y.D.; Zhou, Y.Q.; Jin, W.Q.; Xie, K.B.; Chen, L.L. CRISPR-P 2.0: An improved CRISPR/Cas9 tool for genome editing in plants. Mol. Plant. 2017, 10, 530–532. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Sun, X.; Yu, D.; Xu, J.; Chung, K.; Li, H. Genomic and transcriptomic analyses of the tangerine pathotype of Alternaria alternata in response to oxidative stress. Sci Rep. 2016, 6, 32437. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Frisch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Woodbury, NY, USA, 1989. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Primer Name | Sequence (5′-3′) |
|---|---|---|
| RPA amplification | RPA-F | CCAAGGCGAAGCGATGTTAAAGTCTAGCACA |
| RPA-R | CATACCCTATCAGTCGCGGGAGATAGAAGCT | |
| crRNA for CRISPR/Cas12a | crRNA-1 | UAAUUUCUACUAAGUGUAGAUCUUUUGUUAGAAGAGGAAUC |
| crRNA-2 | UAAUUUCUACUAAGUGUAGAUCCACUGGCGCCUUGUAUCCG | |
| crRNA-3 | UAAUUUCUACUAAGUGUAGAUUAAAGCGAAGUGGCAAUACU | |
| Fluorescent detection probe for CRISPR/Cas12a | ssDNA-FQ | FAM-/TTATTATT/-BHQ1 |
| Lateral flow (strips) detection probe for CRISPR/Cas12a | ssDNA-FB | FAM-/TTATTATT/-Biotin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Tang, J.; Ma, Z.; Zhang, Y.; Ye, Z.; Fu, H. Development of an RPA-CRISPR/Cas12a-Based Rapid Diagnosis Strip for the Tangerine Pathotype of Alternaria alternata. Microorganisms 2025, 13, 826. https://doi.org/10.3390/microorganisms13040826
Li W, Tang J, Ma Z, Zhang Y, Ye Z, Fu H. Development of an RPA-CRISPR/Cas12a-Based Rapid Diagnosis Strip for the Tangerine Pathotype of Alternaria alternata. Microorganisms. 2025; 13(4):826. https://doi.org/10.3390/microorganisms13040826
Chicago/Turabian StyleLi, Wenge, Jintian Tang, Zhiya Ma, Yafen Zhang, Zihong Ye, and Huilan Fu. 2025. "Development of an RPA-CRISPR/Cas12a-Based Rapid Diagnosis Strip for the Tangerine Pathotype of Alternaria alternata" Microorganisms 13, no. 4: 826. https://doi.org/10.3390/microorganisms13040826
APA StyleLi, W., Tang, J., Ma, Z., Zhang, Y., Ye, Z., & Fu, H. (2025). Development of an RPA-CRISPR/Cas12a-Based Rapid Diagnosis Strip for the Tangerine Pathotype of Alternaria alternata. Microorganisms, 13(4), 826. https://doi.org/10.3390/microorganisms13040826