
Lab-Scale Experimental Study of Microbial Enhanced Oil Recovery on Low-Permeability Cores Using the Silicate Bacterium Paenibacillus mucilaginosus



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Microorganism Strains
2.1.2. Cores
2.1.3. Crude Oil
2.2. Methods
2.2.1. Microbial Cultivation
2.2.2. Measurement of Microbial Cell Concentration
2.2.3. Measurements of pH, Redox Potential, and Conductivity
2.2.4. Measurement of Surface Tension
2.2.5. MEOR Simulation Experiment
2.2.6. Measurement of Element Concentration
2.2.7. Fraction Analysis of Crude Oil
2.2.8. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis of Saturated and Aromatic Hydrocarbons
2.2.9. Scanning Electron Microscope and Energy Dispersive X-Ray Spectroscopy (SEM-EDX)
2.2.10. μCT Detection
3. Results and Discussion
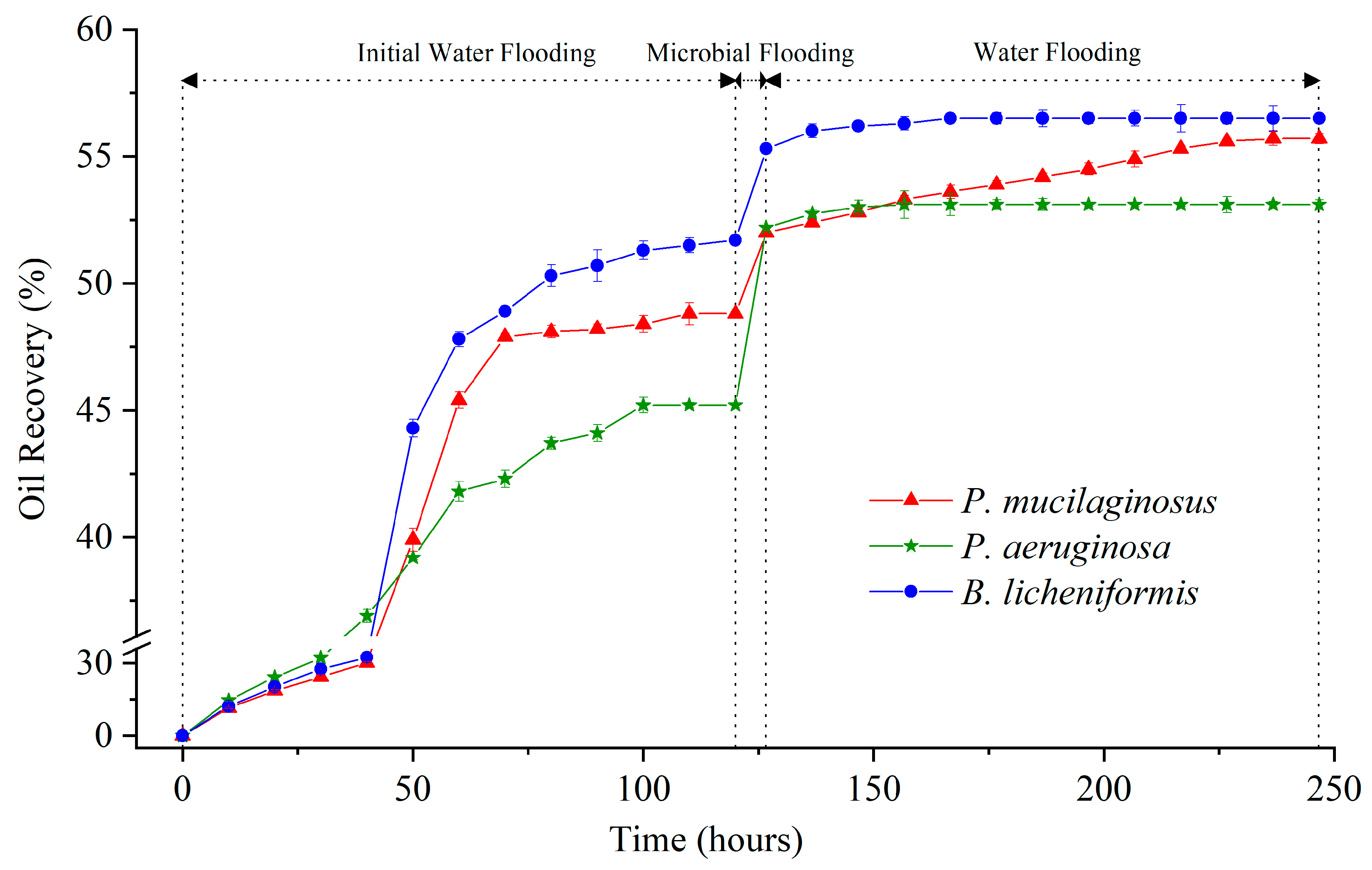
3.1. Initial Water Flooding Stage
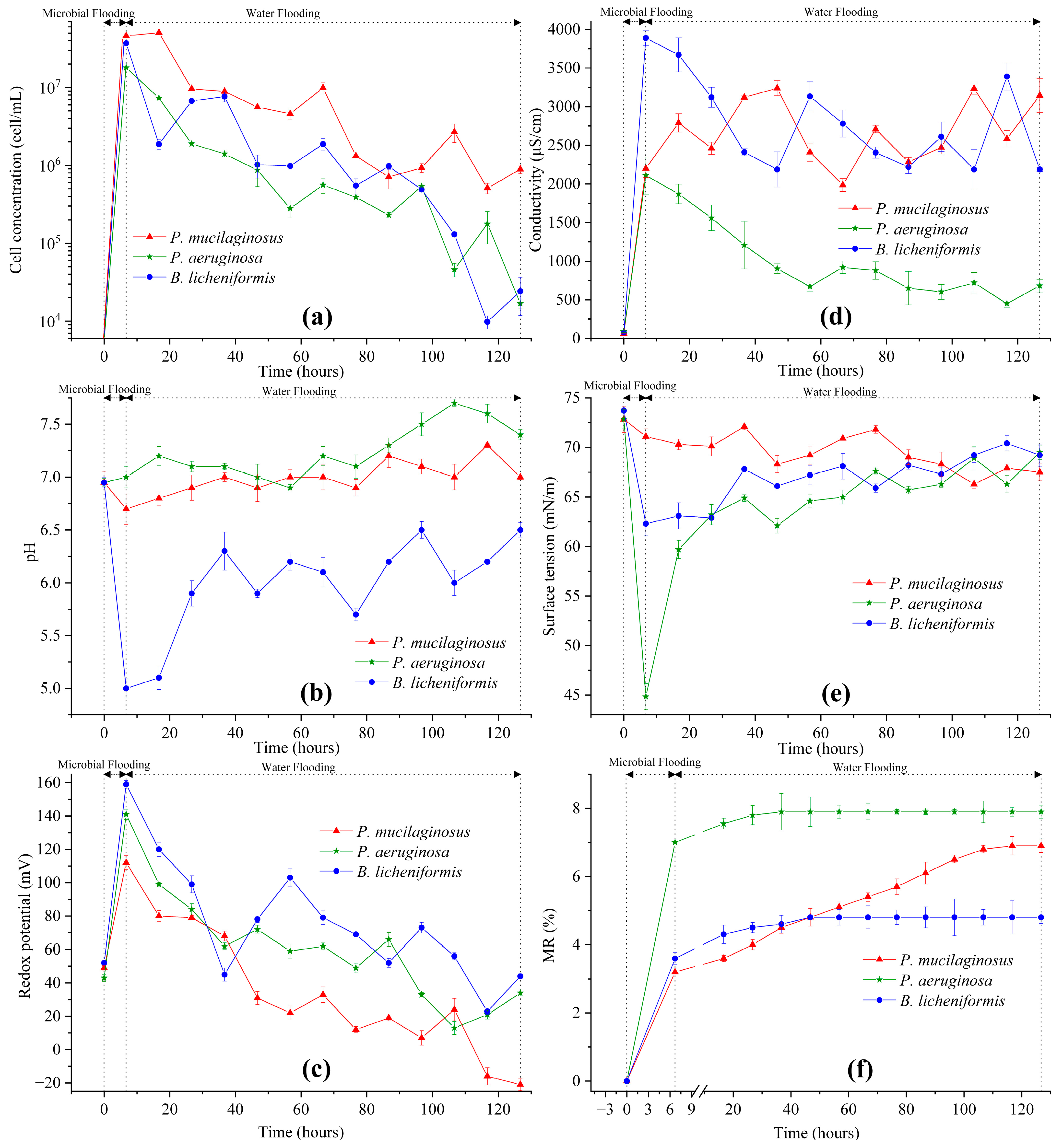
3.2. Microbial Flooding Stage
3.3. Water Flooding Stage
3.3.1. Microorganisms
3.3.2. Microbial Dissolution of Core Minerals
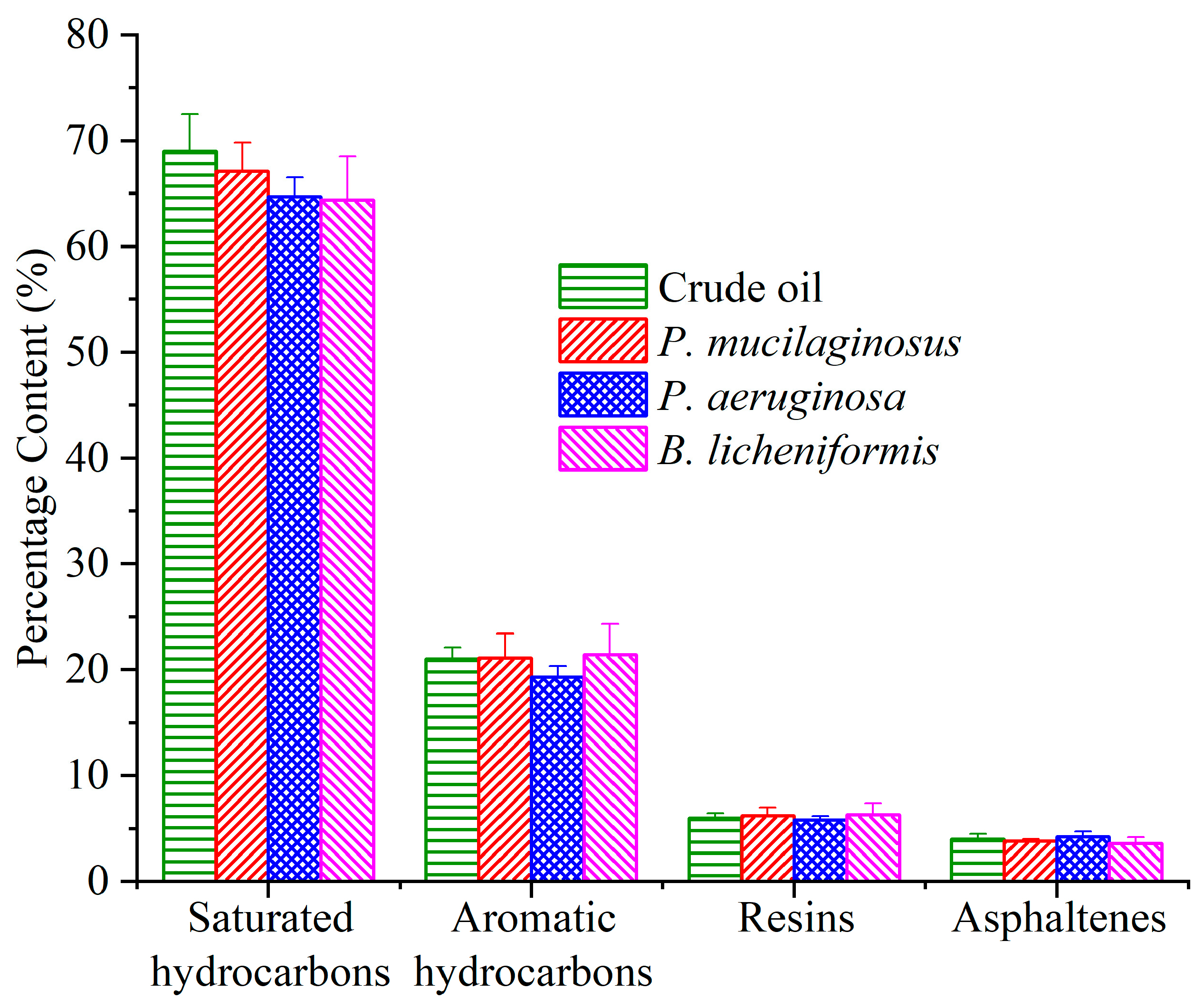
3.3.3. Microbial Degradation of Crude Oil
3.3.4. The Enhanced Oil Recovery of Water Flooding
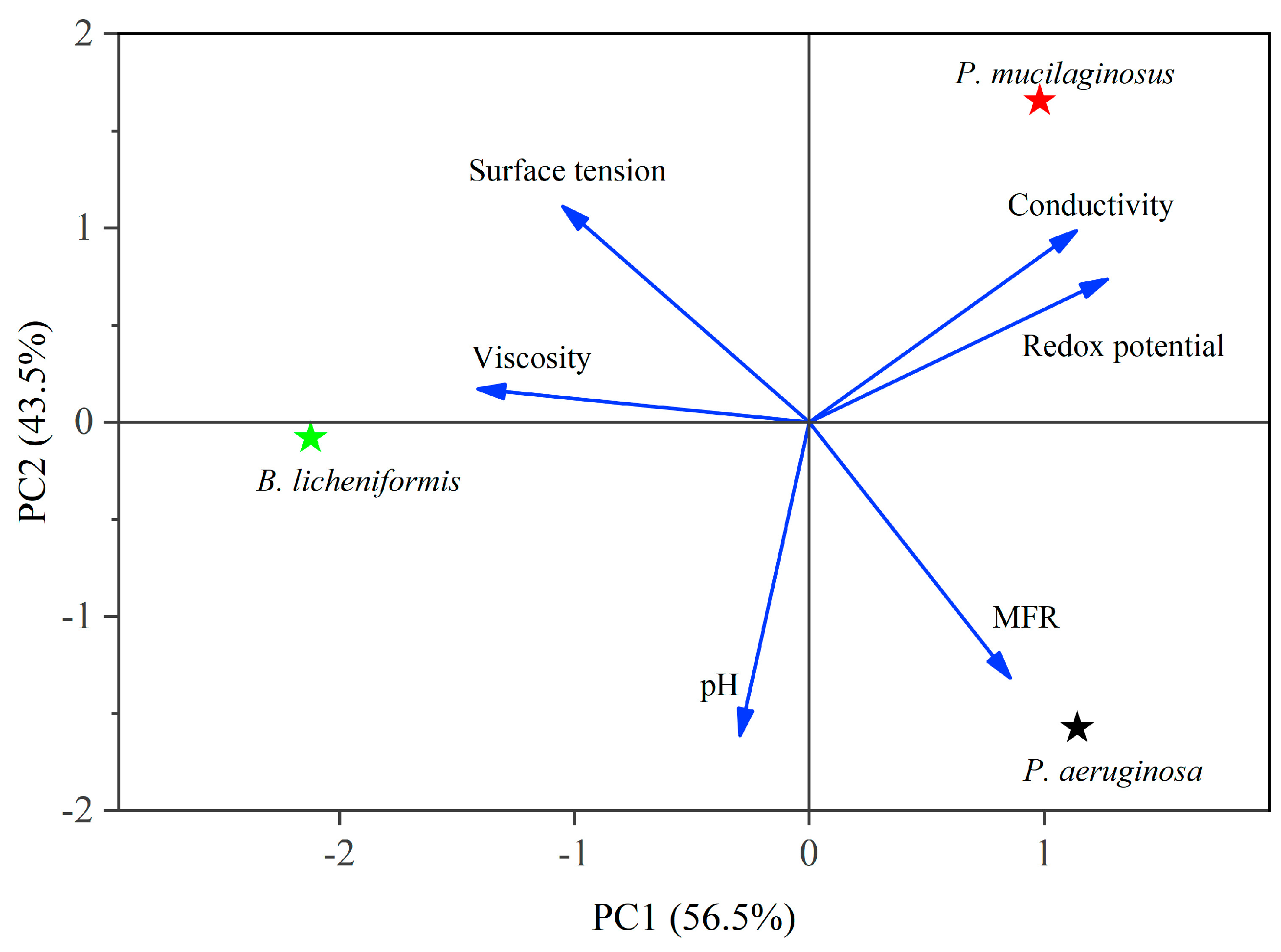
3.4. MEOR Characteristics of Silicate Bacterium P. mucilaginosus
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- SY/T 6169-2024; China National Energy Administration, 2021. Classification and Development Technical Specifications for Low-Permeability Oil and Gas Reservoirs. Petroleum Industry Press: Beijing, China, 2021. (In Chinese)
- Kokkinos, N.; Nkagbu, D.; Marmanis, D.; Konstantinos, D.; Maliaris, G. Evolution of unconventional hydrocarbons: Past, present, future and environmental footprint. J. Eng. Sci. Technol. Rev. 2022, 15, 15–24. [Google Scholar] [CrossRef]
- Kang, W.L.; Zhou, B.B.; Issakhov, M.; Gabdullin, M. Advances in enhanced oil recovery technologies for low permeability reservoirs. Pet. Sci. 2022, 19, 1622–1640. [Google Scholar] [CrossRef]
- Cui, K.; Zhang, Z.Y.; Zhang, Z.Z.; Sun, S.S.; Li, H.L.; Fu, P.C. Stimulation of indigenous microbes by optimizing the water cut in low permeability reservoirs for green and enhanced oil recovery. Sci. Rep. 2019, 9, 15772. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Zhang, Z.Z.; Wang, J.X.; Zhang, G.Q.; Luo, Y.J.; Song, Z.Z.; Zhang, J.Y. Bacterial community diversity in a low-permeability oil reservoir and its potential for enhancing oil recovery. Bioresour. Technol. 2013, 147, 110–116. [Google Scholar] [CrossRef]
- Bian, Z.W.; Wu, H.N.; Wei, L.S.; Zhi, Z.N.; Wu, Y.F. Isolation and characterization of viscosity-reducing and biosurfactant-producing bacteria in low-permeability reservoir. Int. J. Energy Res. 2023, 2023, 3223516. [Google Scholar]
- Wang, Y.; Cheng, S.Q.; Zhang, K.D.; He, Y.W.; Yu, H.Y. Pressure-transient analysis of water injectors considering the multiple closures of waterflood-induced fractures in tight reservoirs: Case studies in Changqing Oilfield, China. J. Pet. Sci. Eng. 2019, 172, 643–653. [Google Scholar] [CrossRef]
- Xiao, M.; Sun, S.S.; Zhang, Z.Z.; Wang, J.M.; Qiu, L.W.; Sun, H.Y.; Song, Z.Z.; Zhang, B.Y.; Gao, D.L.; Zhang, G.Q.; et al. Analysis of bacterial diversity in two oil blocks from two low-permeability reservoirs with high salinities. Sci. Rep. 2016, 6, 19600. [Google Scholar] [CrossRef]
- Wu, B.; Xiu, J.L.; Yu, L.; Huang, L.X.; Yi, L.N.; Ma, Y.D. Biosurfactant production by Bacillus subtilis SL and its potential for enhanced oil recovery in low permeability reservoirs. Sci. Rep. 2022, 12, 7785. [Google Scholar] [CrossRef]
- Yin, J.; Wei, X.X.; Hu, F.T.; Cheng, C.K.; Zhuang, X.L.; Song, M.Y.; Zhuang, G.Q.; Wang, F.; Ma, A.Z. Halotolerant Bacillus velezensis sustainably enhanced oil recovery of low permeability oil reservoirs by producing biosurfactant and modulating the oil microbiome. Chem. Eng. J. 2023, 453, 139912. [Google Scholar] [CrossRef]
- Li, J.; Xue, S.W.; He, C.Q.; Qi, H.X.; Chen, F.L.; Ma, Y.L. Effect of exogenous inoculants on enhancing oil recovery and indigenous bacterial community dynamics in long-term field pilot of low permeability reservoir. World J. Microbiol. Biotechnol. 2018, 34, 53. [Google Scholar] [CrossRef]
- Zhao, Y.; Guo, Z.H.; Wu, J.C.; Shi, H.Q. Construction and performance evaluation of dual-metabolite oil-producing engineering bacteria suitable for low-permeability reservoir. Lithosphere 2022, 2022, 8427896. [Google Scholar]
- Dong, H.; Zhang, Z.Z.; He, Y.L.; Luo, Y.J.; Xia, W.J.; Sun, S.S.; Zhang, G.Q.; Zhang, Z.Y.; Gao, D.L. Biostimulation of biogas producing microcosm for enhancing oil recovery in low-permeability oil reservoir. RSC Adv. 2015, 5, 91869–91877. [Google Scholar]
- Ghojavand, H.; Vahabzadeh, F.; Shahraki, A.K. Enhanced oil recovery from low permeability dolomite cores using biosurfactant produced by a Bacillus mojavensis (PTCC 1696) isolated from Masjed-I Soleyman field. J. Pet. Sci. Eng. 2012, 81, 24–30. [Google Scholar]
- Sen, R. Biotechnology in petroleum recovery: The microbial EOR. Prog. Energy Combust. Sci. 2008, 34, 714–724. [Google Scholar] [CrossRef]
- Dong, H.; Zheng, A.; He, Y.; Wang, X.; Li, Y.; Yu, G.; Gu, Y.; Banat, I.M.; Sun, S.; She, Y.; et al. Optimization and characterization of biosurfactant produced by indigenous Brevibacillus borstelensis isolated from a low permeability reservoir for application in MEOR. RSC Adv. 2022, 12, 2036–2047. [Google Scholar] [CrossRef]
- Li, Y.; Li, H.; Chen, S.; Mbia, E.; Wang, K.; Ren, H. Capillarity characters measurement and effects analysis in different permeability formations during waterflooding. Fuel 2017, 194, 129–143. [Google Scholar]
- Chakraborty, S.; Govindarajan, S.K.; Gummadi, S.N. Influence of crucial reservoir properties and microbial kinetic parameters on enhanced oil recovery by microbial flooding under nonisothermal conditions: Mathematical modelling and numerical simulation. J. Pet. Sci. Eng. 2020, 195, 107831. [Google Scholar]
- Chakraborty, S.; Govindarajan, S.K.; Gummadi, S.N. Numerical modeling on the influence of effective porosity, microbial kinetics, and operational parameters on enhanced oil recovery by microbial flooding within a sandstone formation. SPE J. 2020, 25, 2932–2961. [Google Scholar]
- Cui, K.; Wang, C.J.; Li, L.; Zou, J.G.; Huang, W.H.; Zhang, Z.Z.; Wang, H.M.; Guo, K. Controlling the hydro-swelling of smectite clay minerals by Fe(III) reducing bacteria for enhanced oil recovery from low-permeability reservoirs. Energies 2022, 15, 4393. [Google Scholar] [CrossRef]
- Ke, C.Y.; Sun, W.J.; Li, Y.B.; Hui, J.F.; Lu, G.M.; Zheng, X.Y.; Zhang, Q.Z.; Zhang, X.L. Polymer-assisted microbial-enhanced oil recovery. Energy Fuels 2018, 32, 5885–5892. [Google Scholar]
- Sun, S.S.; Dong, H.; Zhnag, Z.Z.; Cao, S.Y.; Wan, Y.Y.; Dong, H.P.; Yu, L. Study on application of gas and acid producing bacteria for enhanced oil recovery of low permeability reservoirs. Chem. Bioeng. 2013, 30, 54–57. (In Chinese) [Google Scholar]
- Li, H.; Ding, L.; Zang, Q.; Wu, Q.; Ma, Y.; Wang, Y.; Zhou, S.; Cheng, Q.; Tian, X.; Niu, J.; et al. Study on sensitivity mechanism of low-permeability sandstone reservoir in Huilu Area of Pearl River Mouth Basin. J. Mar. Sci. Eng. 2024, 12, 12060888. [Google Scholar] [CrossRef]
- Osman, A.G. Study of some characteristics of silicate bacteria. J. Sci. Technol. 2009, 10, 24–31. [Google Scholar]
- Chiang, Y.W.; Santos, R.M.; Van Audenaerde, A.; Monballiu, A.; Van Gerven, T.; Meesschaert, B. Chemoorganotrophic bioleaching of olivine for nickel recovery. Minerals 2014, 4, 553–564. [Google Scholar] [CrossRef]
- Lv, Y.; Li, J.; Ye, H.; Du, D.; Gan, C.; Wuri, L.; Sun, P.; Wen, J. Bioleaching of silicon in electrolytic manganese residue using single and mixed silicate bacteria. Bioprocess Biosyst. Eng. 2019, 42, 1819–1828. [Google Scholar] [CrossRef]
- Lv, Y.; Li, J.; Ye, H.P.; Du, D.Y.; Sun, P.; Ma, M.Y.; Zhang, T.C. Bioleaching of silicon in electrolytic manganese residue (EMR) by Paenibacillus mucilaginosus: Impact of silicate mineral structures. Chemosphere 2020, 256, 127043. [Google Scholar] [CrossRef]
- Capelli, I.A.; Scasso, R.A.; Spangenberg, J.E.; Kietzmann, D.A.; Cravero, F.; Duperron, M.; Adatte, T. Mineralogy and geochemistry of deeply-buried marine sediments of the Vaca Muerta-Quintuco system in the Neuquen Basin (Chacay Melehue section), Argentina: Paleoclimatic and paleoenvironmental implications for the global Tithonian-Valanginian reconstructions. J. South Am. Earth Sci. 2021, 107, 103103. [Google Scholar]
- Wang, D.; Poinsot, V.; Li, W.; Lu, Y.; Liu, C.; Li, Y.; Xie, K.; Sun, L.; Shi, C.; Peng, H.; et al. Genomic insights and functional analysis reveal plant growth promotion traits of Paenibacillus mucilaginosus G78. Genes 2023, 14, 14020392. [Google Scholar] [CrossRef]
- Mercl, F.; Tejnecky, V.; Sagova-Mareckova, M.; Dietel, K.; Kopecky, J.; Brendova, K.; Kulhanek, M.; Kosnar, Z.; Szakova, J.; Tlustos, P. Co-application of wood ash and Paenibacillus mucilaginosus to soil: The effect on maize nutritional status, root exudation and composition of soil solution. Plant Soil 2018, 428, 105–122. [Google Scholar] [CrossRef]
- Attar, A. Effects of azorhizophilus paspali and Paenibacillus mucilaginosus as biofertilizer and determination of nutritional efficiency by sensors. Arab. J. Sci. Eng. 2018, 43, 3477–3484. [Google Scholar] [CrossRef]
- Teng, Q.; Feng, Y.L.; Li, H.R. Effects of silicate-bacteria pretreatment on desiliconization of magnesite by reverse flotation. Colloids Surf. A Physicochem. Eng. Asp. 2018, 544, 60–67. [Google Scholar]
- Lv, Y.; Li, J.; Liu, X.; Chen, B.; Zhang, M.; Chen, Z.; Zhang, T.C. Screening of silicon-activating bacteria and the activation mechanism of silicon in electrolytic manganese residue. Environ. Res. 2021, 202, 111659. [Google Scholar] [PubMed]
- Camara, J.M.D.A.; Sousa, M.A.S.B.; Barros Neto, E.L.; Oliveira, M.C.A. Application of rhamnolipid biosurfactant produced by Pseudomonas aeruginosa in microbial-enhanced oil recovery (MEOR). J. Pet. Explor. Prod. Technol. 2019, 9, 2333–2341. [Google Scholar] [CrossRef]
- Amani, H.; Sarrafzadeh, M.H.; Haghighi, M.; Mehrnia, M.R. Comparative study of biosurfactant producing bacteria in MEOR applications. J. Pet. Sci. Eng. 2010, 75, 209–214. [Google Scholar]
- Khademolhosseini, R.; Jafari, A.; Mousavi, S.M.; Hajfarajollah, H.; Noghabi, K.A.; Manteghian, M. Physicochemical characterization and optimization of glycolipid biosurfactant production by a native strain of Pseudomonas aeruginosa HAK01 and its performance evaluation for the MEOR process. RSC Adv. 2019, 9, 7932–7947. [Google Scholar] [CrossRef]
- Lee, J.; Jaeseon, K.; Jin, H.B. Characterization of Bacillus licheniformis as a probiotic. Korean Soc. Biotechnol. Bioeng. J. 2005, 20, 359–362. [Google Scholar]
- GB/T 29617-2013; General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, 2013. Determination of Density, Relative Density, and API Gravity of Iquids by Digital Density Meter. China Quality Inspection press: Beijing, China, 2013. (In Chinese)
- GB/T 30515-2014; General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, 2014. Petroleum Products-Transparent and Opaque Liquids-Determination of Kinematic Viscosity and Calculation of Dynamic Viscosity. China Quality Inspection Press: Beijing, China, 2014. (In Chinese)
- Wan, Y.Y.; Dong, H.L. Environmental Geomicrobiology Experiments; Petroleum Industry Press: Beijing, China, 2014; pp. 100–105. (In Chinese) [Google Scholar]
- Zhao, J.Y.; Li, L.; Wan, Y.Y.; Li, Z.G.; Luo, N.; Mu, H.M.; Li, W.H.; Zhang, Y. Interaction between in-situ oil reservoir microorganisms and minerals. J. China Univ. Pet. (Ed. Nat. Sci.) 2021, 45, 121–130. (In Chinese) [Google Scholar]
- GB/T 22237-2008; General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, 2008. Surface Active Agents-Determination of Surface Tension. Standards Press of China: Beijing, China, 2008. (In Chinese)
- SY/T 6888-2012; China National Energy Administration, 2012. Microbial Flooding Technical Specifications. Petroleum Industry Press: Beijing, China, 2012. (In Chinese)
- GB/T 29172-2012; General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, 2012. Practices for Core Analysis. China Quality Inspection Press: Beijing, China, 2012. (In Chinese)
- SY/T 5345-2007; National Development and Reform Commission, 2007. Test Method for Two Phase Relative Permeability in Rock. Petroleum Industry Press: Beijing, China, 2007. (In Chinese)
- Fulazzaky, M.; Astuti, D.I.; Ali Fulazzaky, M. Laboratory simulation of microbial enhanced oil recovery using Geobacillus toebii R-32639 isolated from the Handil reservoir. RSC Adv. 2015, 5, 3908–3916. [Google Scholar]
- Karimi, M.; Mahmoodi, M.; Niazi, A.; Al-Wahaibi, Y.; Ayatollahi, S. Investigating wettability alteration during MEOR process, a micro/macro scale analysis. Colloids Surf. B Biointerfaces 2012, 95, 129–136. [Google Scholar]
- Soleimani, Y.; Mohammadi, M.; Schaffie, M.; Zabihi, R.; Ranjbar, M. An experimental study of the effects of bacteria on asphaltene adsorption and wettability alteration of dolomite and quartz. Sci. Rep. 2023, 13, 21497. [Google Scholar] [CrossRef]
- Li, X.; Du, Y.; Wu, G.; Li, Z.; Li, H.; Sui, H. Solvent extraction for heavy crude oil removal from contaminated soils. Chemosphere 2012, 88, 245–249. [Google Scholar] [CrossRef] [PubMed]
- HJ 700-2014; Ministry of Environmental Protection, 2014. Water Quality-Determination of 65 Elements-Inductively Coupled Plasma-Mass Spectrometry. China Environmental Science Press: Beijing, China, 2014. (In Chinese)
- Liu, P.; Shi, Q.; Chung, K.H.; Zhang, Y.H.; Pan, N.; Zhao, S.Q.; Xu, C.M. Molecular characterization of sulfur compounds in Venezuela crude oil and Its SARA fractions by electrospray ionization Fourier transform ion cyclotron resonance mass spectrometry. Energy Fuels 2010, 24, 5089–5096. [Google Scholar]
- SY/T 5119-2016; China National Energy Administration, 2016. Analysis Method for Family Composition of Rock Extracts and Crude Oil. Petroleum Industry Press: Beijing, China, 2016. (In Chinese)
- Liu, Y.; Shi, S.B.; Zeng, Q.; Li, Y.; Chen, Y.; Guo, D.Y.; Hu, D.F.; Dong, H.L. Coupled reduction of structural Fe(III) in nontronite and oxidation of petroleum hydrocarbons. Geochim. Et Cosmochim. Acta 2023, 344, 103–121. [Google Scholar]
- SY/T 5779-2024; China National Energy Administration, 2024. Petrology and Diagenetic Organic Matter Hydrocarbon Gas Chromatographic Analysis Method. Petroleum Industry Press: Beijing, China, 2024. (In Chinese)
- Li, Y.; Liu, Y.; Guo, D.Y.; Dong, H.L. Differential degradation of petroleum hydrocarbons by Shewanella putrefaciens under aerobic and anaerobic conditions. Front. Microbiol. 2024, 15, 1389954. [Google Scholar]
- Camelo, D.D.L.; Gilkes, R.J.; Leopold, M.; Silva, A.C.; Vidal-Torrado, P. The application of quartz grain morphology measurements to studying iron-rich duricrusts. Catena 2018, 170, 397–408. [Google Scholar]
- Al-Yaseri, A.Z.; Lebedev, M.; Vogt, S.J.; Johns, M.L.; Barifcani, A.; Iglauer, S. Pore-scale analysis of formation damage in Bentheimer sandstone with in-situ NMR and micro-computed tomography experiments. J. Pet. Sci. Eng. 2015, 129, 48–57. [Google Scholar]
- Cui, J.; Liu, D.M.; Cai, Y.D.; Pan, Z.J.; Zhou, Y.F. Insights into fractures and minerals in subbituminous and bituminous coals by FESEM-EDS and X-ray μ-CT. Fuel 2019, 237, 977–988. [Google Scholar]
- Bird, M.B.; Butler, S.L.; Hawkes, C.D.; Kotzer, T. Numerical modeling of fluid and electrical currents through geometries based on synchrotron X-ray tomographic images of reservoir rocks using Avizo and COMSOL. Comput. Geosci. 2014, 73, 6–16. [Google Scholar]
- Niu, Y.B.; Jing, C.H.; Zhang, L.J.; Jin, Y.; Liu, S.X.; Cheng, M.Y.; Shao, W.M.; Cheng, Y.G.; Pan, J.N. Quantitative characterization of 3D pore structures and percolation characteristics in bioturbated reservoir media based on X-ray micro-CT: A case study of the Neogene Sanya Formation in the Qiongdongnan Basin, Northern South China Sea. Ichnos-Int. J. Plant Anim. Traces 2024, 31, 199–219. [Google Scholar]
- Bao, M.; Kong, X.; Jiang, G.; Wang, X.; Li, X. Laboratory study on activating indigenous microorganisms to enhance oil recovery in Shengli Oilfield. J. Pet. Sci. Eng. 2009, 66, 42–46. [Google Scholar]
- Sharifipour, M.; Nakhaee, A.; Pourafshary, P. Model development of permeability impairment due to clay swelling in porous media using micromodels. J. Pet. Sci. Eng. 2019, 175, 728–742. [Google Scholar]
- Zhou, Y.; Yang, W.; Yin, D. Experimental investigation on reservoir damage caused by clay minerals after water injection in low permeability sandstone reservoirs. J. Pet. Explor. Prod. Technol. 2022, 12, 915–924. [Google Scholar]
- Zhang, L.; Zhou, F.; Zhang, S.; Wang, Y.; Wang, J.; Wang, J. Investigation of Water-Sensitivity Damage for Tight Low-Permeability Sandstone Reservoirs. ACS Omega 2019, 4, 11197–11204. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Jiang, H.; Li, J.; Zhao, L. Experimental study on the micro alkali sensitivity damage mechanism in low-permeability reservoirs using QEMSCAN. J. Nat. Gas Sci. Eng. 2016, 36, 1004–1017. [Google Scholar]
- Shi, X.; Wei, J.; Bo, H.; Zheng, Q.; Yi, F.; Yin, Y.; Chen, Y.; Dong, M.; Zhang, D.; Li, J.; et al. A novel model for oil recovery estimate in heterogeneous low-permeability and tight reservoirs with pseudo threshold pressure gradient. Energy Rep. 2021, 7, 1416–1423. [Google Scholar] [CrossRef]
- Cui, G.; Yang, L.; Fang, J.; Qiu, Z.; Wang, Y.; Ren, S. Geochemical reactions and their influence on petrophysical properties of ultra-low permeability oil reservoirs during water and CO2 flooding. J. Pet. Sci. Eng. 2021, 203, 108672. [Google Scholar]
- Wang, X.; Liu, Y.; Hou, J.; Wang, D.; Ji, L.; Sun, J.; Li, Y.; Yan, X. Impacts of water flooding on pore structure of sandstone reservoirs-case study of Wang Guantun oilfield, Bohai Bay Basin, China. Arab. J. Geosci. 2018, 11, 580. [Google Scholar]
- Leal, E.; Teixeira, J.A.; Gudina, E.J. Development of foam-free biosurfactant production processes using Bacillus licheniformis. Fermentation 2024, 10, 10070340. [Google Scholar] [CrossRef]
- Kaster, K.M.; Hiorth, A.; Kjeilen-Eilertsen, G.; Boccadoro, K.; Lohne, A.; Berland, H.; Stavland, A.; Brakstad, O.G. Mechanisms involved in microbially enhanced oil recovery. Transp. Porous Media 2012, 91, 59–79. [Google Scholar] [CrossRef]
- Raturi, G.; Sharma, Y.; Rana, V.; Thakral, V.; Myaka, B.; Salvi, P.; Singh, M.; Dhar, H.; Deshmukh, R. Exploration of silicate solubilizing bacteria for sustainable agriculture and silicon biogeochemical cycle. Plant Physiol. Biochem. 2021, 166, 827–838. [Google Scholar]
- Wang, L.; Zhang, H.; Peng, X.; Wang, P.; Zhao, N.; Chu, S.; Wang, X.; Kong, L. Water-sensitive damage mechanism and the injection water source optimization of low permeability sandy conglomerate reservoirs. Pet. Explor. Dev. 2019, 46, 1218–1230. [Google Scholar]
- He, W.; Liu, Z. Numerical simulation of formation damage by drilling fluid in low permeability sandstone reservoirs. J. Pet. Explor. Prod. Technol. 2021, 11, 1865–1871. [Google Scholar]
- Patel, J.; Borgohain, S.; Kumar, M.; Rangarajan, V.; Somasundaran, P.; Sen, R. Recent developments in microbial enhanced oil recovery. Renew. Sustain. Energy Rev. 2015, 52, 1539–1558. [Google Scholar]
- Bordoloi, N.K.; Konwar, B.K. Microbial surfactant-enhanced mineral oil recovery under laboratory conditions. Colloids Surf. B Biointerfaces 2008, 63, 73–82. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
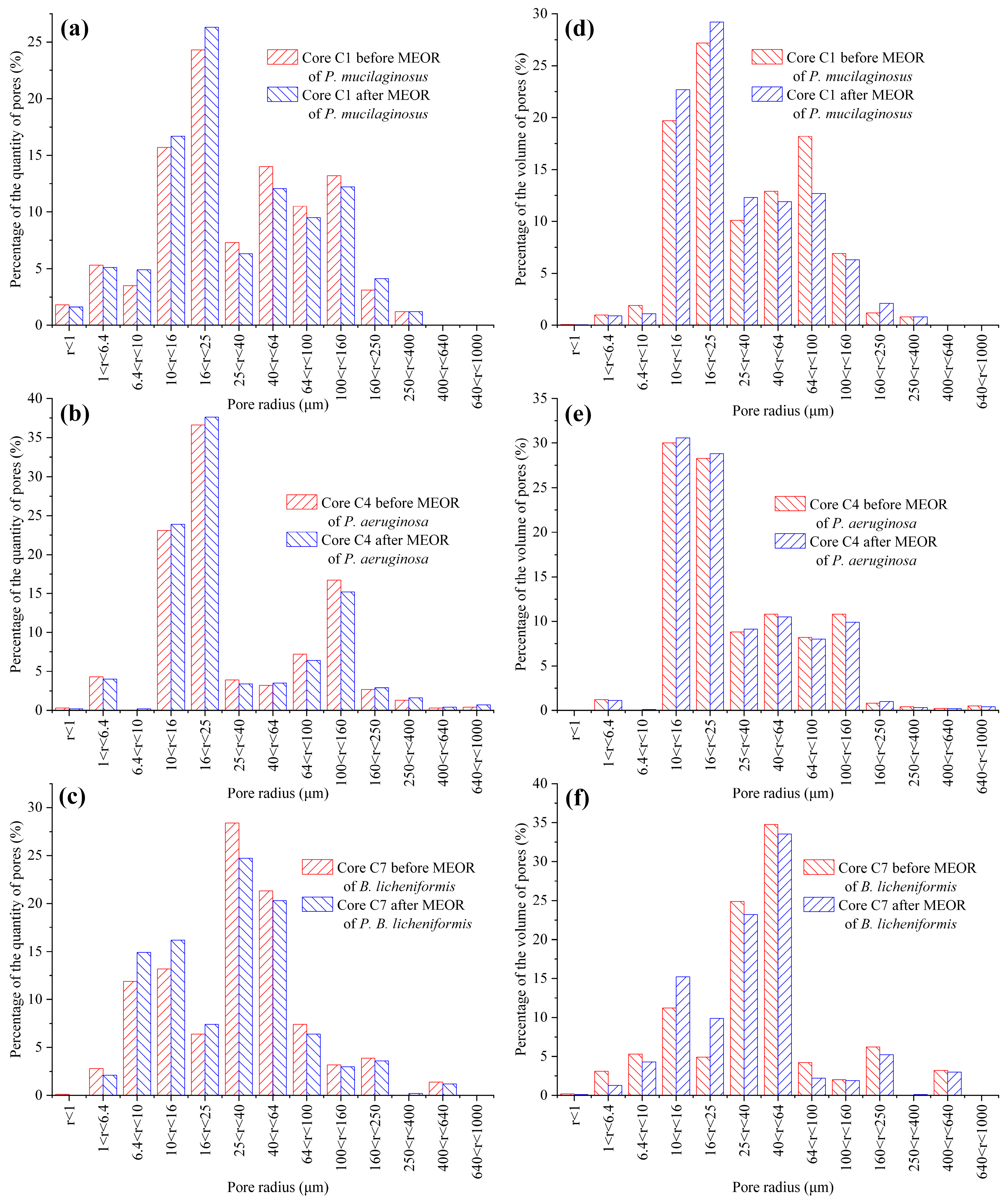
| Core No. | Total Pore Quantity | Total Pore Volume * | ||||
|---|---|---|---|---|---|---|
| Before (Pores) | After (Pores) | Difference (%) | Before (cm3) | After (cm3) | Difference (%) | |
| C1 | 507,593 | 519,129 | 2.27 | 16.61 | 18.33 | 4.33 |
| C4 | 539,832 | 510,014 | −5.52 | 17.49 | 17.01 | −2.74 |
| C7 | 510,987 | 529,926 | 3.71 | 18.92 | 19.08 | 0.85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Zhang, C.; Su, P.; Mu, H. Lab-Scale Experimental Study of Microbial Enhanced Oil Recovery on Low-Permeability Cores Using the Silicate Bacterium Paenibacillus mucilaginosus. Microorganisms 2025, 13, 738. https://doi.org/10.3390/microorganisms13040738
Li L, Zhang C, Su P, Mu H. Lab-Scale Experimental Study of Microbial Enhanced Oil Recovery on Low-Permeability Cores Using the Silicate Bacterium Paenibacillus mucilaginosus. Microorganisms. 2025; 13(4):738. https://doi.org/10.3390/microorganisms13040738
Chicago/Turabian StyleLi, Lei, Chunhui Zhang, Peidong Su, and Hongmei Mu. 2025. "Lab-Scale Experimental Study of Microbial Enhanced Oil Recovery on Low-Permeability Cores Using the Silicate Bacterium Paenibacillus mucilaginosus" Microorganisms 13, no. 4: 738. https://doi.org/10.3390/microorganisms13040738
APA StyleLi, L., Zhang, C., Su, P., & Mu, H. (2025). Lab-Scale Experimental Study of Microbial Enhanced Oil Recovery on Low-Permeability Cores Using the Silicate Bacterium Paenibacillus mucilaginosus. Microorganisms, 13(4), 738. https://doi.org/10.3390/microorganisms13040738