Abstract
Persistent apical periodontitis (PAP) of human teeth is related to Enterococcus faecalis (E. faecalis) with higher alkaline resistance. This study aimed to investigate how highly alkaline-resistant (HAR) E. faecalis modulates macrophage M1 polarization and phagocytosis via Z-DNA-binding protein 1 (ZBP1). HAR E. faecalis was generated through serial alkaline passaging. RAW264.7 macrophages were infected with standard or HAR E. faecalis. M1 polarization markers and Cd274 (Programmed death-ligand 1 (PD-L1)) were profiled by RT-qPCR. ZBP1 was detected by RT-qPCR and immunofluorescence staining, and was silenced using small interfering RNA (siRNA). iNOS, ZBP1 and PD-L1 proteins were analyzed by Western blotting. The phagocytosis of CFDA-SE-labeled bacteria was then quantified by confocal microscopy and flow cytometry. The results showed that HAR E. faecalis induced significantly lower expression of ZBP1, M1 polarization markers and Cd274 in macrophages than the standard strain. After ZBP1 knock-down, expression of these markers decreased. Macrophages phagocytosed much fewer HAR E. faecalis than the standard strain. After ZBP1 knock-down, the differences between the two strains disappeared. In conclusion, HAR E. faecalis induced compromised M1 polarization and phagocytosis of macrophage via ZBP1. These findings may provide new insights into the pathogenesis and treatment of PAP.
1. Introduction
Persistent apical periodontitis (PAP) of human teeth refers specifically to a chronic pathological condition characterized by persistent or recurrent inflammation following root canal treatment. Although modern root canal treatment techniques exhibit a high success rate, PAP still occurs in approximately 30–40% of cases [1]. The primary etiological factor of PAP is the residual microbial biofilm within the root canal system and around the root apex, particularly Enterococcus faecalis (E. faecalis), which is detected in 24–77% of PAP cases and is frequently associated with intracanal calcium hydroxide (CH) medication [2]. Notably, E. faecalis isolated from the root canals of PAP patients shows high alkaline resistance [3,4]. This resistance enables their survival in the alkaline environment of CH-treated root canals, which may contribute to the pathogenesis of PAP [5]. However, why the alkaline-resistant E. faecalis is related to PAP and the differences in inflammation-inducing or suppressing potential between highly alkaline-resistant (HAR) and normal E. faecalis remains unclear.
Macrophages serve as central regulators in the immune response during PAP [6]. Recent studies have demonstrated that Z-DNA binding protein 1 (ZBP1), as a critical nucleic acid sensor [7], primarily activates downstream signaling pathways (particularly RIPK3-MLKL-mediated necroptosis [8], NLRP3 inflammasome [9,10] and NF-κB/IRF [11,12,13]) by sensing Z-RNA/Z-DNA derived from pathogens or endogenous damage signals [14,15], thereby playing a pivotal regulatory role in macrophage function. Macrophage polarization state directly influences disease progression: pro-inflammatory M1 macrophages exert antimicrobial effects through cytokines such as TNF-α and IL-6 [16], yet exacerbate bone tissue destruction [17,18]. The phagocytic function of macrophages serves as a critical defense mechanism for the host to eliminate bacterial infections [19,20]. However, it is unclear whether ZBP1 is involved in the inflammatory responses during HAR E. faecalis infection through regulating macrophage M1 polarization and phagocytosis.
Programmed death-ligand 1 (PD-L1), as a pivotal immune checkpoint molecule [21], plays a critical role in tumor immune escape [22] and chronic infections [23] by binding to the PD-L1 receptor to suppress T-cell activity [24] and to remodel macrophage function [25,26]. In the context of oral infections, PD-L1 levels are significantly elevated in the periodontal tissues of periodontitis patients [27] and are closely associated with the virulence factors of Porphyromonas gingivalis [28,29,30], suggesting that PD-L1 may serve as a target for pathogen-mediated immune escape. However, in PAP, the association of PD-L1 with persistent E. faecalis infection remains unknown. A study showed that ZBP1 also contributed to Unfolded Protein Response (UPR) activation during infection [12]. The UPR process is initiated by the endoplasmic reticulum trans-membrane protein inositol-requiring enzyme 1α (IRE1α/ERN1) [31]. Promoting PD-L1 expression through the IRE1α-XBP1 axis is beneficial for eliminating anti-tumor immunity [32]. It is worth investigating whether HAR E. faecalis affects PD-L1-dependent immunosuppression via modulating ZBP1.
This study aimed to compare the differential inflammatory effects between HAR and normal E. faecalis and investigate whether HAR E. faecalis influences the M1 polarization and phagocytic ability of macrophage via ZBP1.
2. Materials and Methods
2.1. Cell Culture
The murine RAW264.7 macrophage cell line ATCC SC-6003 (ATCC, Manassas, VA, USA) was maintained in Dulbecco’s Modified Eagle Medium (DMEM, GIBCO, Waltham, MA, USA) containing 10% fetal bovine serum (GIBCO, Waltham, MA, USA). For experimental procedures, cells were plated at a density of 1 × 106 cells per well in six-well plates and allowed to adhere overnight prior to experimentation.
2.2. E. faecalis Culture
Prior to experimental applications, standard E. faecalis ATCC 29212 (ATCC, Manassas, VA, USA) was maintained in Brain Heart Infusion broth (BHI, Beijing Land Bridge Technology, Beijing, China) at 37 °C, 5% CO2, 1% O2.
2.3. Screening of HAR E. faecalis
Exponential-phase standard E. faecalis suspensions (100 μL, OD600 = 1.0) were transferred to an alkaline (pH = 10) BHI medium containing 3 mg/mL CH [4]. Sequential subculturing (3 generations, 37 °C/24–48 h per cycle) under sustained alkaline stress yielded alkali-adapted variants that were purified by single-colony streaking on agar plates and designated as HAR E. faecalis. For phenotypic validation, equal quantities of HAR and standard E. faecalis were cultured separately in the same alkaline BHI medium mentioned above. After 24 h of incubation, 50 μL of liquid was plated on BHI agar plates and incubated at 37 °C for 24 h. High alkaline resistance was confirmed when HAR E. faecalis demonstrated a ≥1 log10 increase in colony-forming units (CFUs) relative to the standard strain (Figure 1A).
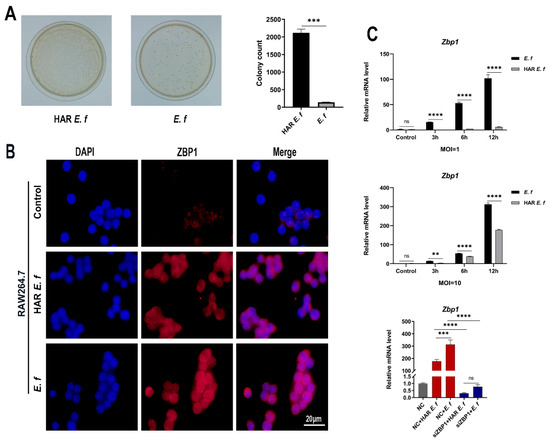
Figure 1.
Activation of ZBP1 in macrophages by different E. faecalis. (A) The difference in CFUs of HAR and standard E. faecalis after being cultured in alkaline BHI for 24 h (Welch’s t-test). (B) Immunofluorescence staining of ZBP1 in HAR and standard E. faecalis-treated (MOI = 10, 12 h) cells (Scale bar, 20 μm). (C) mRNA levels of Zbp1 in RAW264.7 cells infected with HAR and standard E. faecalis (MOI = 1, 10) for 3 h, 6 h and 12 h (Two-way ANOVA); and mRNA levels of Zbp1 in RAW264.7 cells infected with HAR and standard E. faecalis (MOI = 10) for 12 h after Zbp1 silencing (One-way ANOVA). NC, negative control. Data are expressed as the mean ±SD (** p < 0.01; *** p < 0.001; **** p < 0.0001; ns = no significant).
2.4. Cell Infection
Prior to microbial stimulation experiments, RAW264.7 cells were seeded in 6-well culture plates at a density of 1 × 106 cells/well and cultured for 12 h, with one well reserved for cell counting. Concurrently, cryopreserved HAR and standard E. faecalis were revived and adjusted to the OD600 of 1.0 (approximately 1 × 109 CFUs/mL). Bacterial suspensions were added into cells at the calculated volumes to achieve the target multiplicities of infection (MOI) of 1 and 10. Infection groups were cultured under 37 °C, 5% CO2 at three time points (3, 6, and 12 h). Cells without infection were used as control.
2.5. Cellular Immunofluorescence Staining
For immunofluorescence analysis, RAW264.7 macrophages were plated in 12-well chambers (5 × 105 cells/well) and exposed to standard and HAR E. faecalis at MOI = 10 for 12 h. Post-infection processing included: (i) Fixation: 4% paraformaldehyde (PFA) in Phosphate-buffered saline (PBS) (20 min, room temperature); (ii) Permeabilization: 0.2% Triton X-100 (Beyotime, Shanghai, China; 10 min); (iii) Blocking: 5% BSA in PBS (30 min, 37 °C). Primary antibody incubation was conducted overnight at 4 °C with anti-ZBP1 (1:500, AG-20B-0010, AdipoGen, Liestal, Switzerland). Samples were subsequently incubated with fluorescently labeled secondary antibodies for 1 h, nuclei were counterstained with DAPI (Beyotime, Shanghai, China) and mounted in anti-fade mounting medium. Images were acquired on a Zeiss LSM880 Fast microscope (Zeiss, Oberkochen, Germany) via ZEN Blue 3.9 software.
2.6. Small Interfering RNA (siRNA)-Mediated Gene Silencing
ZBP1-targeting siRNA (siZBP1; GenePharma, Suzhou, China) was reverse-transfected into RAW264.7 cells using siRNA-mate transfection reagent (GenePharma, Suzhou, China) according to the manufacturer’s protocol. Parallel control groups received validated scrambled siRNA (NC-siRNA; GenePharma, Suzhou, China) at an equivalent concentration, and the knockdown result was confirmed by RT-qPCR and Western blot. The siRNA sequences are listed in Table 1.

Table 1.
siRNA sequences for gene knockdown.
2.7. Reverse Transcription Quantitative Real-Time PCR (RT-qPCR)
Cellular RNA isolation was conducted with TRIzol reagent (Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA), followed by first-strand cDNA synthesis using ABScript III Reverse Transcriptase Master Mix (ABclonal, Wuhan, China). Quantitative amplification was performed in triplicate on a LightCycler 480 II platform (Roche Diagnostics, Basel, Switzerland) with SYBR Green master mix (Servicebio, Wuhan, China). Relative transcript quantification employed the comparative threshold cycle (2−ΔΔCT) method, with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) serving as the endogenous normalization control. Primer sequences used in this experiment are listed in Table 2.
Table 2.
Primer sequences used for the RT-qPCR.
2.8. Western Blot Analysis
Cellular lysates were prepared using RIPA lysis buffer supplemented with protease inhibitor cocktail (both from Beyotime, Shanghai, China). Protein samples underwent denaturation (95 °C, 5 min) prior to electrophoretic separation on 10% SDS-PAGE systems (Servicebio, Wuhan, China). Wet transfer (200 mA, 60 min) immobilized proteins onto PVDF membranes (0.22 µm, Merck Millipore, Billerica, MA, USA), followed by 1 h blocking in 5% non-fat milk in Tris-Buffered Saline with Tween 20 (TBST). Membranes were probed overnight at 4 °C with the following validated antibodies: anti-iNOS (1:1000, T55993, Abmart, Shanghai, China), anti-ZBP1 (1:1000, AG-20B-0010, AdipoGen, Liestal, Switzerland), anti-PD-L1 (1:1000, M033179, Abmart, Shanghai, China) and anti-β-actin (1:1000, PMK058, BIOPRIMACY, Wuhan, China). After TBST washes (3 × 10 min), membranes were incubated with HRP-conjugated secondary antibody (1:5000, SA00001-1 or SA00001-2, Proteintech, Chicago, IL, USA) for 1 h at room temperature. Chemiluminescent detection was performed using ECL Prime (EpiZyme, Shanghai, China) with signal capture on an Odyssey System (LI-COR Biosciences, Lincoln, NE, USA). Densitometric analysis utilized ImageJ 1.53t (NIH, Bethesda, MD, USA).
2.9. Confocal Microscopy Imaging Analysis
Log-phase cultures of both standard and HAR E. faecalis were fluorescently labeled with CFDA-SE (Elabscience, Wuhan, China). The labeled bacteria were used to infect both control and Zbp1-knockdown murine macrophages in the log phase (grown on coverslips) at an MOI of 20 [33]. Infection was conducted at 37 °C for 40 min in the dark. Phagocytosis was terminated by incubating samples on ice for 10 min. A 0.04% trypan blue solution was applied for 1 min to quench fluorescence from extracellular bacteria. Samples were rinsed three times with PBS. Cells were fixed with 4% paraformaldehyde at room temperature for 20 min, followed by three PBS washes. Cells were incubated with TRITC-phalloidin (OriLeaf, Shanghai, China) for 30 min at room temperature in the dark, followed by PBS washes. Samples were mounted using anti-fade mounting medium containing DAPI (Beyotime, Shanghai, China). Processed samples were imaged using a confocal laser scanning microscope (CLSM, Zeiss LSM880, Carl Zeiss, Oberkochen, Germany).
2.10. Phagocytosis Assay
Both control and Zbp1-knockdown RAW264.7 cells were infected with CFDA-SE (Elabscience, Wuhan, China)-labeled standard or HAR E. faecalis (MOI = 20 [33]) at 37 °C for 40 min in the dark. Extracellular bacteria were quenched with 0.04% trypan blue. After three PBS washes, the cells were pelleted (200 × g, 5 min), resuspended in PBS, and analyzed on a CytoFLEX flow cytometer (Beckman Coulter Life Sciences, Indianapolis, IN, USA) using the FITC-A channel.
2.11. Statistical Analysis
All experiments were conducted in triplicate and data were presented as mean ± standard deviation (SD). Data analysis was performed using GraphPad Prism 8.0 software. Comparisons were made using Welch’s t-test, one-way analysis of variance (ANOVA), or two-way ANOVA. Statistical significance was set at p < 0.05 (* p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001).
3. Results
3.1. HAR E. faecalis Induced Lower ZBP1 Expression in Macrophages than Standard Strain
Immunofluorescence results demonstrated that ZBP1 protein expression in RAW264.7 cells was upregulated upon both standard and HAR E. faecalis infection (Figure 1B). RT-qPCR and Western blot analyses confirmed that both standard and HAR E. faecalis infection upregulated ZBP1 expression when compared with the control group, while HAR E. faecalis induced significantly lower ZBP1 expression in macrophages than the standard E. faecalis (Figure 1C and Figure 2A, Figure 2B, Figure 2C and Figure 2D). Additionally, ZBP1 expression was higher at the MOI of 10 relative to the MOI of 1 (Figure 1C). Upon ZBP1 knockdown using siRNA, the ZBP1 levels activated by both standard and HAR E. faecalis were markedly reduced without difference between the two strains (Figure 1C and Figure 2E,F).
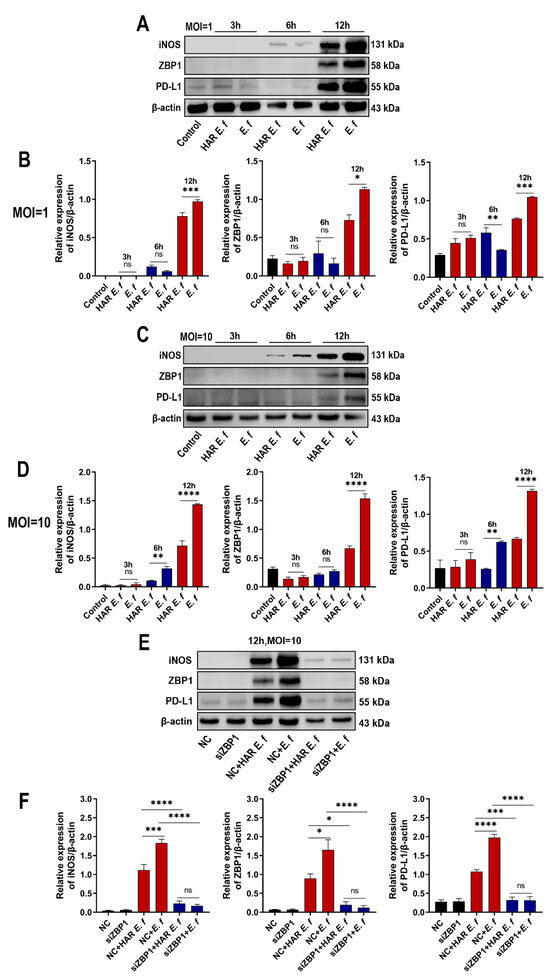
Figure 2.
Macrophages stimulated with HAR E. faecalis showed compromised protein expression of iNOS and PD-L1 via ZBP1, relative to those stimulated with the standard strain. (A–D) Protein levels of iNOS, ZBP1 and PD-L1 in RAW264.7 cells infected with HAR and standard E. faecalis (MOI = 1, 10) for 3 h, 6 h and 12 h. The quantitative data represent the relative ratio of the target protein to β-actin (Two-way ANOVA). (E,F) Protein levels of iNOS, ZBP1 and PD-L1 in RAW264.7 cells infected with HAR and standard E. faecalis (MOI = 10) for 12 h after ZBP1 knockdown (NC, negative control). The quantitative data represent the relative ratio of the target protein to β-actin (One-way ANOVA). Data are expressed as the mean ±SD (* p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001; ns = no significant).
3.2. HAR E. faecalis Showed Compromised M1 Polarization Expression and Cd274(PD-L1) in Macrophages Through ZBP1
RT-qPCR and Western blot analyses revealed that in RAW264.7 macrophages, both standard and HAR E. faecalis induced dose- and time-dependent upregulation of the M1 markers Nos2 (iNOS), Il6, Tnf and the immune checkpoint gene Cd274 (PD-L1), yet HAR E. faecalis showed significantly lower expressions of these genes than the standard strain ( Figure 2A–D and Figure 3). siRNA-mediated ZBP1 silencing eliminated the difference in expression of M1 markers and Cd274 (PD-L1) induced by both HAR and standard E. faecalis and significantly reduced their overall expression levels (Figure 2E,F and Figure 3).
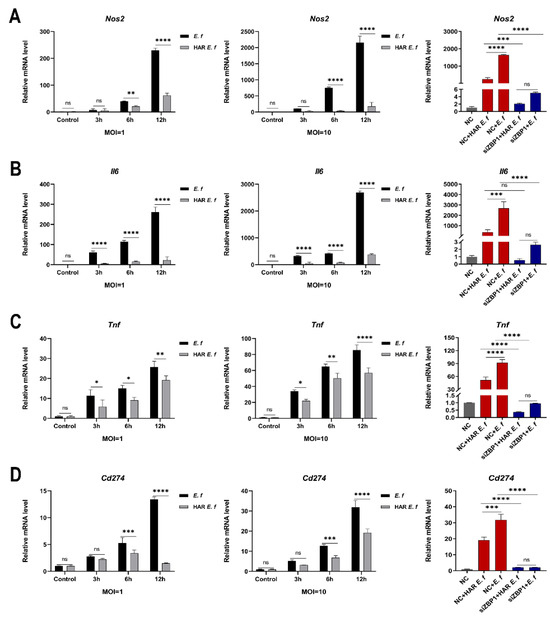
Figure 3.
Macrophages stimulated with HAR E. faecalis showed compromised gene expression of M1 polarization markers and Cd274 via Zbp1, relative to those stimulated with the standard strain. (A–D) mRNA levels of Nos2, Il6, Tnf and Cd274 in RAW264.7 cells infected with HAR and standard E. faecalis (MOI = 1, 10) for 3 h, 6 h and 12 h (Two-way ANOVA); and mRNA levels of Nos2, Il6, Tnf and Cd274 in RAW264.7 cells infected with HAR and standard E. faecalis (MOI = 10) for 12 h after Zbp1 silencing (One-way ANOVA). NC, negative control. Data are expressed as the mean ±SD (* p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001; ns = no significant).
3.3. HAR E. faecalis Compromised Macrophage Phagocytosis
Confocal microscopy and flow cytometric analysis revealed that both strains induced a typical M1-polarized morphology in macrophages, characterized by cell enlargement, extensive membrane ruffling, and pseudopodia formation after 40 min of infection at an MOI of 20 (Figure 4). The number of HAR E. faecalis internalized by RAW264.7 macrophages was significantly lower than that of the standard E. faecalis. Furthermore, following ZBP1 knockdown, phagocytosis of macrophage cells on both E. faecalis strains was attenuated without difference between the strains, and intracellular bacteria were significantly reduced (Figure 5).
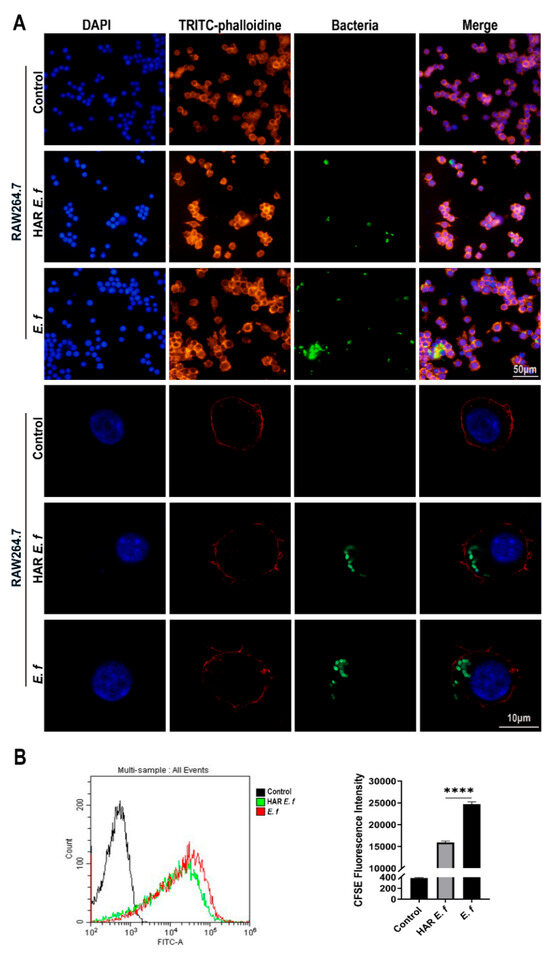
Figure 4.
Phagocytosis of macrophages on standard and HAR E. faecalis. (A) Confocal microscopy images of macrophages infected with CFDA-SE-labeled HAR and standard E. faecalis (MOI = 20). (B) Flow cytometry plots of macrophages infected with CFDA-SE-labeled HAR and standard E. faecalis (MOI = 20; one-way ANOVA). Data are expressed as the mean ±SD (**** p < 0.0001).
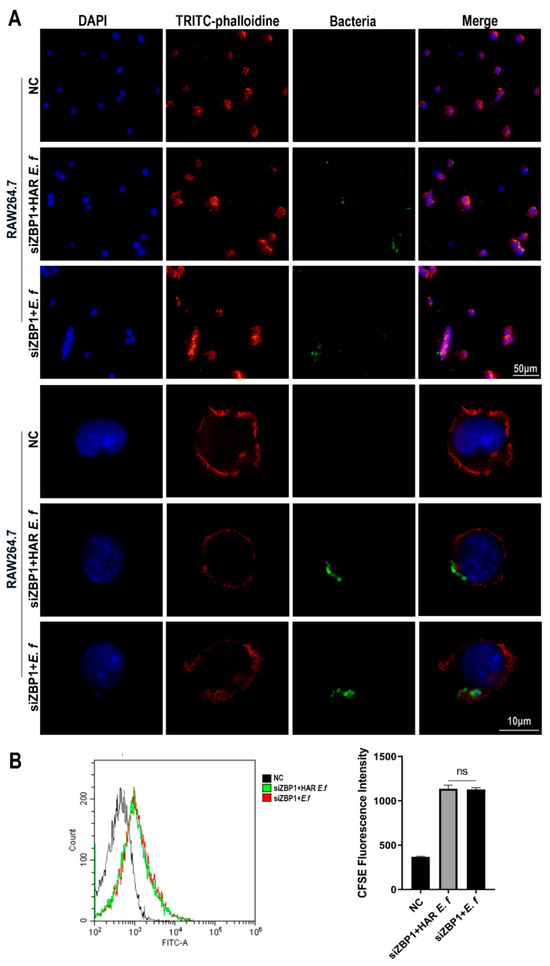
Figure 5.
Phagocytosis of macrophages on standard and HAR E. faecalis after ZBP1 knockdown. (A) Confocal microscopy images of macrophages infected with CFDA-SE-labeled HAR and standard E. faecalis after ZBP1 knockdown (MOI = 20). (B) Flow cytometry plots of macrophages infected with CFDA-SE-labeled HAR and standard E. faecalis after ZBP1 knockdown (MOI = 20; one-way ANOVA). Data are expressed as the mean ±SD (ns = no significant).
4. Discussion
In PAP, E. faecalis has been identified as a related pathogen [34]. E. faecalis, especially those that survive root canal disinfection and medication procedures, often show alkaline resistance to CH [35], but the correlation between these alkaline-resistant E. faecalis and immune status in the apical tissues of PAP remains unclear. Macrophages serve as the primary cells responsible for recognizing and eliminating bacteria in periapical tissues [36], and their pro-inflammatory M1 polarization could regulate the progression of inflammation and promote pathogen removal [37]. The phagocytic ability of macrophages is also crucial for eliminating bacteria and can be influenced by bacteria [38]. Their innate immune sensor ZBP1 can influence pathogen clearance and tissue damage [39]. However, ZBP1’s role in PAP related to HAR E. faecalis is also unclear. Therefore, this study aimed to investigate the potential mechanism by which HAR E. faecalis affects macrophage M1 polarization and phagocytosis through regulating ZBP1 in macrophages.
Previous studies identified F. nucleatum-infected RAW264.7 cells polarized to the M1 phenotype, and this was accompanied by inflammatory cytokine production [40]. ZBP1 inhibition reduced inflammatory cytokine secretion and the occurrence of PANoptosis [41]. This study indicated that both HAR and standard E. faecalis can up-regulate ZBP1 in macrophages, exhibiting time-dependent and MOI-dependent patterns. However, the HAR strain showed a weaker ability to activate ZBP1 expression compared to the standard strain. Potential explanations for this phenomenon include unique genetic characteristics of the HAR strain which may encode specific proteins that interfere with ZBP1 transcription or signal transduction [9]. At the mechanistic level, ZBP1 can activate NF-κB and MAPK signaling pathways [42], promoting the massive release of inflammatory cytokines such as TNF-α and IL-6. These cytokines, in turn, drive macrophage polarization toward the M1 phenotype via autocrine or paracrine signaling, establishing a pro-inflammatory state [16]. In this study, the expression trends of M1 polarization markers were consistent with those of ZBP1, indicating that ZBP1 plays an important role in regulating macrophage polarization.
Recent studies have revealed that PD-L1 expression is not only regulated by the host microenvironment but can also be directly hijacked by pathogens [43]. E. faecalis, a pathogen related to post-endodontic treatment recurrence [44], may promote chronic infection through PD-L1-mediated suppression of both antimicrobial immunity and tissue repair [45], though the underlying molecular pathways remain unresolved. In this study, compared to the standard strain, HAR E. faecalis exhibited a unique “low immune activation-low immunosuppression” phenotype. The low level of immune activation compromises antigen presentation and co-stimulatory signals, thereby inhibiting the activation of antigen-specific T and B cells and ultimately preventing the host from developing robust, long-lasting immunity [46], which can lead to chronic or recurrent periapical infections. This study confirms that ZBP1 is a key molecule in inducing PD-L1 expression by HAR and standard strains, as its knockout leads to a significant reduction in PD-L1 expression. It is noteworthy that the HAR strain exhibits significantly weaker activation of the ZBP1-PD-L1 axis compared to the standard strain, yet they retain some PD-L1 induction capability even after ZBP1 knockout. This phenomenon may suggest that HAR strain could evade host surveillance by activating alternative pathways such as TLR2/MyD88 [47] or cGAS-STING [48] as a compensatory mechanism to the PD-L1.
Previous reports have demonstrated that bacterial surface modifications (e.g., capsular polysaccharides [49,50], protein conformation [51], or charge alterations [52]) can inhibit phagocytosis. The anti-phagocytic characteristics of E. faecalis involve synergistic actions of multiple mechanisms [53,54]. However, the specific molecular mechanisms linking its alkaline resistance to phagocytosis inhibition remain to be fully elucidated. In this study, comparative analysis of macrophage phagocytic capacity revealed a significantly lower efficiency in engulfing HAR E. faecalis compared to standard E. faecalis. During the early stage of infection, ZBP1 activates the NF-κB and MAPK signaling pathways [42], thereby enhancing phagocytic capacity [55,56]. In this study, ZBP1 knock down resulted in reduced phagocytic capacity of macrophages, further confirming its critical role in early immunity. Furthermore, HAR E. faecalis induced compromised expression of PD-L1 and phagocytosis via ZBP1, thereby facilitating immune evasion. Similar mechanisms have been reported in methicillin-resistant Staphylococcus aureus [57,58], suggesting that different pathogens may employ analogous strategies to achieve immune evasion.
Although this study indicated the mechanistic role of ZBP1 in macrophage-mediated inflammatory responses to HAR E. faecalis infection, several limitations remain to be addressed. First, the mechanisms by which HAR E. faecalis regulates ZBP1 expression, and how this differs from the standard E. faecalis strain, remain incompletely understood. It is unclear whether this regulation involves Toll-like receptor signaling or alterations in bacterial surface proteins. Second, systematic comparative analyses are needed to determine whether there are intrinsic differences in surface protein profiles between HAR and standard E. faecalis strains. Third, the molecular basis underlying the interaction between ZBP1 and macrophage phagocytic capacity needs further investigations. In particular, it remains to be elucidated whether ZBP1-mediated regulation is directly controlled by specific transcription factors or mediated through intermediate signaling molecules. Future studies should prioritize deciphering the molecular “inflammatory switch” mechanism governed by ZBP1 and explore the development of therapeutic strategies targeting specific inflammatory pathways to control refractory infections involving HAR E. faecalis.
5. Conclusions
HAR E. faecalis induced compromised M1 polarization and significantly impaired the phagocytic capacity of macrophages compared with the standard strain, thereby likely enhancing its capacity for immune evasion. These findings may provide new insights into the pathogenesis and treatment of PAP of human teeth.
Author Contributions
Y.X.: Writing—review and editing, Writing—original draft, Visualization, Methodology, Investigation, Formal analysis, Data curation. R.L.: Writing—review and editing, Validation, Methodology, Formal analysis, Data curation, Conceptualization. Y.Y. and X.L.: Methodology, Investigation, Formal analysis, Data curation. Y.M. and W.F.: Writing—review and editing, Writing—original draft, Supervision, Resources, Project administration, Funding acquisition, Conceptualization. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 82270968) and Natural Science Foundation of Hubei Province of China (2024AFB721).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| ANOVA | Analysis of Variance |
| ATCC | American Type Culture Collection |
| BSA | Bovine Serum Albumin |
| CFDA-SE | Carboxyfluorescein Diacetate Succinimidyl Ester |
| CFU | Colony-Forming Unit |
| CH | Calcium Hydroxide |
| CLSM | Confocal Laser Scanning Microscopy |
| DAPI | 4′,6-Diamidino-2-Phenylindole |
| DMEM | Dulbecco’s Modified Eagle Medium |
| E. faecalis | Enterococcus faecalis |
| FBS | Fetal Bovine Serum |
| GAPDH | Glyceraldehyde-3-Phosphate Dehydrogenase |
| HAR | Highly Alkaline-Resistant |
| HRP | Horseradish Peroxidase |
| iNOS | Inducible Nitric Oxide Synthase |
| IRE1α/ERN1 | Inositol-Requiring Enzyme 1α |
| MOI | Multiplicity of Infection |
| PAP | Persistent Apical Periodontitis |
| PBS | Phosphate-Buffered Saline |
| PD-L1 | Programmed Death-Ligand 1 |
| PFA | Paraformaldehyde |
| PVDF | Polyvinylidene Fluoride |
| RT-qPCR | Reverse Transcription Quantitative real-time Polymerase Chain Reaction |
| SD | Standard Deviation |
| SDS-PAGE | Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis |
| siRNA | Small Interfering RNA |
| TBST | Tris-Buffered Saline with Tween 20 |
| UPR | Unfolded Protein Response |
| ZBP1 | Z-DNA Binding Protein 1 |
References
- Bergenholtz, G. Assessment of treatment failure in endodontic therapy. J. Oral Rehabil. 2016, 43, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, G.; Ren, B.; Gao, Y.; Peng, X.; Li, M.; Xu, H.H.K.; Han, Q.; Li, J.; Zhou, X.; et al. Effect of Antibacterial Root Canal Sealer on Persistent Apical Periodontitis. Antibiotics 2021, 10, 741. [Google Scholar] [CrossRef] [PubMed]
- Salem, A.S.; Tompkins, G.R.; Cathro, P.R. Alkaline Tolerance and Biofilm Formation of Root Canal Isolates of Enterococcus faecalis: An In Vitro Study. J. Endod. 2022, 48, 542–547.e4. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Liu, P.; Luo, Y.; Fan, W.; Fan, B. Metformin reduced the alkaline resistance of Enterococcus faecalis against calcium hydroxide via Man-PTS EII: In vitro and in vivo studies. Clin. Oral Investig. 2024, 28, 520. [Google Scholar] [CrossRef] [PubMed]
- Pascale, C.; Geaman, J.; Mendoza, C.; Gao, F.; Kaminski, A.; Cuevas-Nunez, M.; Darvishan, B.; Mitchell, J.C.; Carrilho, M.R.; Sigar, I. In vitro assessment of antimicrobial potential of low molecular weight chitosan and its ability to mechanically reinforce and control endogenous proteolytic activity of dentine. Int. Endod. J. 2023, 56, 1337–1349. [Google Scholar] [CrossRef]
- Hernández-Sandoval, E.M.; Sánchez-Gutiérrez, R.; Torres-Monjarás, A.P.; Alvarado-Hernández, D.L.; Méndez-González, V.; Hernández-Castro, B.; Bernal-Silva, S.; Comas-García, A.; Martínez-Rider, R.; González-Amaro, R.; et al. α-IRAK-4 Suppresses the Activation of RANK/RANKL Pathway on Macrophages Exposed to Endodontic Microorganisms. Int. J. Mol. Sci. 2024, 25, 8434. [Google Scholar] [CrossRef]
- Mishra, S.; Dey, A.A.; Kesavardhana, S. Z-Nucleic Acid Sensing and Activation of ZBP1 in Cellular Physiology and Disease Pathogenesis. Immunol. Rev. 2025, 329, e13437. [Google Scholar] [CrossRef]
- Chen, C.; Xie, J.; Chen, Z.; Ye, K.; Wu, C.; Dai, X.; Yuan, Y.; Lin, Y.; Wang, Y.; Chen, H.; et al. Role of Z-DNA Binding Protein 1 Sensing Mitochondrial Z-DNA and Triggering Necroptosis in Oxalate-Induced Acute Kidney Injury. J. Am. Soc. Nephrol. 2025, 36, 361–377. [Google Scholar] [CrossRef]
- Ma, Z.; Liu, D.; Cao, W.; Guo, L.; Liu, K.; Bai, J.; Li, X.; Jiang, P.; Liu, X. Suppression of ZBP1-mediated NLRP3 inflammasome by the tegument protein VP22 facilitates pseudorabies virus infection. mBio 2024, 15, e0194524. [Google Scholar] [CrossRef]
- Yan, S.; Yu, L.; Chen, Z.; Xie, D.; Huang, Z.; Ouyang, S. ZBP1 promotes hepatocyte pyroptosis in acute liver injury by regulating the PGAM5/ROS pathway. Ann. Hepatol. 2024, 29, 101475. [Google Scholar] [CrossRef]
- Sun, K.; Lu, F.; Hou, L.; Zhang, X.; Pan, C.; Liu, H.; Zheng, Z.; Guo, Z.; Ruan, Z.; Hou, Y.; et al. IRF1 regulation of ZBP1 links mitochondrial DNA and chondrocyte damage in osteoarthritis. Cell Commun. Signal. 2024, 22, 366. [Google Scholar] [CrossRef]
- Gomes, M.T.R.; Guimarães, E.S.; Oliveira, S.C. ZBP1 senses Brucella abortus DNA triggering type I interferon signaling pathway and unfolded protein response activation. Front. Immunol. 2024, 15, 1511949. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Ortega-Ribera, M.; Thevkar Nagesh, P.; Joshi, R.; Huang, H.; Wang, Y.; Zivny, A.; Mehta, J.; Parikh, S.M.; Szabo, G. Bile acid-induced IRF3 phosphorylation mediates cell death, inflammatory responses, and fibrosis in cholestasis-induced liver and kidney injury via regulation of ZBP1. Hepatology 2024, 79, 752–767. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhu, Y.; Tian, Z.; Liu, M.; Gao, A.; Fu, W.; Lu, F.; Sun, Y.; Guo, Y.; Pan, R.; et al. ZBP1 senses spliceosome stress through Z-RNA:DNA hybrid recognition. Mol. Cell 2025, 85, 1790–1805.e1797. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.H.; Wu, P.; Zhang, B.X.; Yang, C.R.; Huang, J.; Wu, L.; Guo, S.H.; Zhou, Y.; Mao, Y.; Yin, Y.; et al. ZBP1 senses splicing aberration through Z-RNA to promote cell death. Mol. Cell 2025, 85, 1775–1789.e7. [Google Scholar] [CrossRef]
- Kong, X.; Zhang, T.; Zhang, Y.; Gao, L.; Wang, W.; Wang, M.; Wang, G.; Lü, K. Overexpression of lncRNA HEM2M alleviates liver injury in mice with non-alcoholic fatty liver disease. Nan Fang Yi Ke Da Xue Xue Bao 2024, 44, 1–8. [Google Scholar]
- Dou, J.; Chen, X.; Zhang, J.; Yang, L.; Lin, J.; Zhu, W.; Huang, D.; Tan, X.P. Gingivalis induce macrophage polarization by regulating hepcidin expression in chronic apical periodontitis. Int. Immunopharmacol. 2024, 142, 113139. [Google Scholar] [CrossRef]
- Bi, J.; Mo, C.; Li, S.; Zeng, J.; Chai, Y.; Yao, M.; Liu, Z.; Yuan, P.; Ni, J.; Xu, S. High concentrations of NaF aggravate periodontitis by promoting M1 polarization in macrophages. Int. Immunopharmacol. 2024, 140, 112830. [Google Scholar] [CrossRef]
- Lesbats, J.; Brillac, A.; Reisz, J.A.; Mukherjee, P.; Lhuissier, C.; Fernández-Monreal, M.; Dupuy, J.W.; Sequeira, A.; Tioli, G.; De La Calle Arregui, C.; et al. Macrophages recycle phagocytosed bacteria to fuel immunometabolic responses. Nature 2025, 640, 524–533. [Google Scholar] [CrossRef]
- Li, T.; Qin, K.; Li, N.; Han, C.; Cao, X. An endosomal LAPF is required for macrophage endocytosis and elimination of bacteria. Proc. Natl. Acad. Sci. USA 2019, 116, 12958–12963. [Google Scholar] [CrossRef]
- Cappelli, L.C. Immune checkpoint inhibitors and rheumatoid arthritis: All roads lead to PD-1? Semin. Arthritis Rheum. 2025, 70s, 152582. [Google Scholar] [CrossRef]
- Simonds, E.F.; Lu, E.D.; Badillo, O.; Karimi, S.; Liu, E.V.; Tamaki, W.; Rancan, C.; Downey, K.M.; Stultz, J.; Sinha, M.; et al. Deep immune profiling reveals targetable mechanisms of immune evasion in immune checkpoint inhibitor-refractory glioblastoma. J. Immunother. Cancer 2021, 9, e002181. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Wang, J.; Du, L.; Hu, X.; Liu, Y.; Zhang, P.; Wang, J.; Dong, Q. PD-L1 in plasmacytoid dendritic cells promote HBV persistence through disrupting humoral immune response. Front. Immunol. 2025, 16, 1545667. [Google Scholar] [CrossRef]
- Zhou, X.; Wu, C.; Wang, X.; Pan, N.; Sun, X.; Chen, B.; Zheng, S.; Wei, Y.; Chen, J.; Wu, Y.; et al. Tumor cell-released autophagosomes (TRAPs) induce PD-L1-decorated NETs that suppress T-cell function to promote breast cancer pulmonary metastasis. J. Immunother. Cancer 2024, 12, e009082. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-de la Cruz, M.L.; Salinas-Carmona, M.C. The immune exhaustion paradox: Activated functionality during chronic bacterial infections. J. Infect. Dev. Ctries. 2024, 18, 1824–1836. [Google Scholar] [CrossRef] [PubMed]
- Xie, P.; Guo, L.; Yu, Q.; Zhao, Y.; Yu, M.; Wang, H.; Wu, M.; Xu, W.; Xu, M.; Zhu, X.D.; et al. ACE2 Enhances Sensitivity to PD-L1 Blockade by Inhibiting Macrophage-Induced Immunosuppression and Angiogenesis. Cancer Res. 2025, 85, 299–313. [Google Scholar] [CrossRef]
- Wang, S.; Nie, F.; Yin, Q.; Tian, H.; Gong, P.; Ju, J.; Liu, J.; Yang, P.; Yang, C. Periodontitis promotes tumor growth and immune evasion via PD-1/PD-L1. Cancer Immunol. Immunother. 2024, 74, 22. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, Z.; Weng, Y.; Sitosari, H.; He, Y.; Zhang, X.; Shiotsu, N.; Fukuhara, Y.; Ikegame, M.; Okamura, H. Gingipain regulates isoform switches of PD-L1 in macrophages infected with Porphyromonas gingivalis. Sci. Rep. 2025, 15, 10462. [Google Scholar] [CrossRef]
- Groeger, S.; Wu, F.; Wagenlehner, F.; Dansranjav, T.; Ruf, S.; Denter, F.; Meyle, J. PD-L1 Up-Regulation in Prostate Cancer Cells by Porphyromonas gingivalis. Front. Cell. Infect. Microbiol. 2022, 12, 935806. [Google Scholar] [CrossRef]
- Adel-Khattab, D.; Groeger, S.; Domann, E.; Chakraborty, T.; Lochnit, G.; Meyle, J. Porphyromonas gingivalis induced up-regulation of PD-L1 in colon carcinoma cells. Mol. Oral Microbiol. 2021, 36, 172–181. [Google Scholar] [CrossRef]
- Oakes, S.A.; Papa, F.R. The role of endoplasmic reticulum stress in human pathology. Annu. Rev. Pathol. 2015, 10, 173–194. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Wang, H.; Yang, Y.; Wang, H.; Zhang, H.; Guo, S.; Chen, J.; Du, J.; Tian, Y.; Ma, J.; et al. SIRT7 orchestrates melanoma progression by simultaneously promoting cell survival and immune evasion via UPR activation. Signal Transduct. Target. Ther. 2023, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.; Cohen, J.M.; Reglinski, M.; Jose, R.J.; Chan, W.Y.; Marshall, H.; de Vogel, C.; Gordon, S.; Goldblatt, D.; Petersen, F.C.; et al. Naturally Acquired Human Immunity to Pneumococcus Is Dependent on Antibody to Protein Antigens. PLoS Pathog. 2017, 13, e1006137, Erratum in PLOS Pathog. 2017, 13, e1006259. [Google Scholar]
- Nair, P.N. On the causes of persistent apical periodontitis: A review. Int. Endod. J. 2006, 39, 249–281. [Google Scholar] [CrossRef]
- Liu, R.; Luo, Y.; Liu, P.; Xiao, S.; Fan, W.; Fan, B. The potential regulatory role of mannose phosphotransferase system EII in alkaline resistance of Enterococcus faecalis. J. Oral Microbiol. 2025, 17, 2487944. [Google Scholar] [CrossRef]
- Gong, Q.; Lv, X.; Liao, C.; Liang, A.; Luo, C.; Wu, J.; Zhou, Y.; Huang, Y.; Tong, Z. Single-cell RNA sequencing combined with proteomics of infected macrophages reveals prothymosin-α as a target for treatment of apical periodontitis. J. Adv. Res. 2024, 66, 349–361. [Google Scholar] [CrossRef]
- Ma, R.Y.; Deng, Z.L.; Du, Q.Y.; Dai, M.Q.; Luo, Y.Y.; Liang, Y.E.; Dai, X.Z.; Guo, S.M.; Zhao, W.H. Enterococcus faecalis Extracellular Vesicles Promote Apical Periodontitis. J. Dent. Res. 2024, 103, 672–682. [Google Scholar] [CrossRef]
- Weißelberg, S.; Both, A.; Failla, A.V.; Huang, J.; Linder, S.; Ohnezeit, D.; Bartsch, P.; Aepfelbacher, M.; Rohde, H. Staphylococcus epidermidis alters macrophage polarization and phagocytic uptake by extracellular DNA release in vitro. NPJ Biofilms Microbiomes 2024, 10, 131. [Google Scholar] [CrossRef]
- Song, Q.; Fan, Y.; Zhang, H.; Wang, N. Z-DNA binding protein 1 orchestrates innate immunity and inflammatory cell death. Cytokine Growth Factor Rev. 2024, 77, 15–29. [Google Scholar] [CrossRef]
- Liu, H.; Liu, Y.; Fan, W.; Fan, B. Fusobacterium nucleatum triggers proinflammatory cell death via Z-DNA binding protein 1 in apical periodontitis. Cell Commun. Signal. 2022, 20, 196. [Google Scholar] [CrossRef]
- Karki, R.; Lee, S.; Mall, R.; Pandian, N.; Wang, Y.; Sharma, B.R.; Malireddi, R.S.; Yang, D.; Trifkovic, S.; Steele, J.A.; et al. ZBP1-dependent inflammatory cell death, PANoptosis, and cytokine storm disrupt IFN therapeutic efficacy during coronavirus infection. Sci. Immunol. 2022, 7, eabo6294. [Google Scholar] [CrossRef]
- Udawatte, D.J.; Rothman, A.L. Viral Suppression of RIPK1-Mediated Signaling. mBio 2021, 12, e0172321. [Google Scholar] [CrossRef]
- Chopra, U.; Sabu, M.K.; Rajmani, R.S.; Chaudhary, A.D.; Gupta, S.K.; Chakravortty, D. Salmonella Effector SseL Induces Programmed Death Ligand 1 Upregulation and T-Cell Inactivation via the β-Catenin Signaling Axis. J. Infect. Dis. 2025, 232, 401–411. [Google Scholar] [CrossRef]
- Liu, R.Z.; Bai, L.A.; Luo, Y.; Liu, P.; Hua, F.; Fan, W.; Fan, B. The presence of Enterococcus in root canal infections based on next-generation sequencing: A systematic review and meta-analysis. Int. Endod. J. 2025, 58, 1331–1353. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, R.; Sun, Q.; Chen, Z.; Fan, W. Enterococcus faecalis and lipoteichoic acid up-regulated PD-L1 of macrophage through TLR and IRE1 α/XBP1 signaling axis. Odontology 2025. [Google Scholar] [CrossRef]
- Wang, X.Q.; Danenberg, E.; Huang, C.S.; Egle, D.; Callari, M.; Bermejo, B.; Dugo, M.; Zamagni, C.; Thill, M.; Anton, A.; et al. Spatial predictors of immunotherapy response in triple-negative breast cancer. Nature 2023, 621, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, W.; Zhang, W.; Wu, S.; Yan, Z. Candida albicans induces upregulation of programmed death ligand 1 in oral squamous cell carcinoma. J. Oral Pathol. Med. 2022, 51, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Leuzzi, G.; Vasciaveo, A.; Taglialatela, A.; Chen, X.; Firestone, T.M.; Hickman, A.R.; Mao, W.; Thakar, T.; Vaitsiankova, A.; Huang, J.W.; et al. SMARCAL1 is a dual regulator of innate immune signaling and PD-L1 expression that promotes tumor immune evasion. Cell 2024, 187, 861–881.e832. [Google Scholar] [CrossRef]
- Liu, X.; Xu, Q.; Yang, X.; Heng, H.; Yang, C.; Yang, G.; Peng, M.; Chan, E.-C.; Chen, S. Capsular polysaccharide enables Klebsiella pneumoniae to evade phagocytosis by blocking host-bacteria interactions. mBio 2025, 16, e0383824. [Google Scholar] [CrossRef] [PubMed]
- Zierke, L.; Mourad, R.; Kohler, T.P.; Müsken, M.; Hammerschmidt, S. Influence of the polysaccharide capsule on virulence and fitness of Klebsiella pneumoniae. Front. Microbiol. 2025, 16, 1450984. [Google Scholar] [CrossRef]
- Hou, W.; Liu, Y.; Wu, S.; Zhang, H.; Guo, B.; Zhang, B.; Qin, X.J.; Li, H. Preadsorption of Serum Proteins Regulates Bacterial Infections and Subsequent Macrophage Phagocytosis on Biomaterial Surfaces. ACS Appl. Bio Mater. 2019, 2, 5957–5964. [Google Scholar] [CrossRef] [PubMed]
- Maschalidi, S.; Ravichandran, K.S. Phagocytosis: Sweet Repulsions via the Glycocalyx. Curr. Biol. 2021, 31, R20–R22. [Google Scholar] [CrossRef]
- Nunez, N.; Derré-Bobillot, A.; Gaubert, S.; Herry, J.M.; Deschamps, J.; Wei, Y.; Baranek, T.; Si-Tahar, M.; Briandet, R.; Serror, P.; et al. Exploration of the role of the virulence factor ElrA during Enterococcus faecalis cell infection. Sci. Rep. 2018, 8, 1749. [Google Scholar] [CrossRef]
- Amuasi, G.R.; Dsani, E.; Owusu-Nyantakyi, C.; Owusu, F.A.; Mohktar, Q.; Nilsson, P.; Adu, B.; Hendriksen, R.S.; Egyir, B. Enterococcus species: Insights into antimicrobial resistance and whole-genome features of isolates recovered from livestock and raw meat in Ghana. Front. Microbiol. 2023, 14, 1254896. [Google Scholar] [CrossRef]
- Marquardt, V.; Theruvath, J.; Pauck, D.; Picard, D.; Qin, N.; Blümel, L.; Maue, M.; Bartl, J.; Ahmadov, U.; Langini, M.; et al. Tacedinaline (CI-994), a class I HDAC inhibitor, targets intrinsic tumor growth and leptomeningeal dissemination in MYC-driven medulloblastoma while making them susceptible to anti-CD47-induced macrophage phagocytosis via NF-kB-TGM2 driven tumor inflammation. J. Immunother. Cancer 2023, 11, e005871. [Google Scholar] [PubMed]
- Yang, Y.; Huang, Y.; Li, J.; Li, K.; Mao, C.; Liu, G.; Li, S.; Wen, L. Chemerin 15 enhances microglial phagocytosis to attenuate cerebral ischemia-reperfusion injury through the ChemR23/p38 MAPK pathway. iScience 2025, 28, 113396. [Google Scholar] [CrossRef]
- Bai, S.; Wen, X.; Li, B.; Hu, R.; Yang, J.; Yu, Q.; Zeng, X.; Feng, H.; Zhu, F.; Cai, Z.; et al. Extracellular vesicles from alveolar macrophages harboring phagocytosed methicillin-resistant Staphylococcus aureus induce necroptosis. Cell Rep. 2024, 43, 114453. [Google Scholar] [CrossRef]
- Terasaki, A.; Ahmed, F.; Okuno, A.; Peng, Z.; Cao, D.Y.; Saito, S. Neutrophils Expressing Programmed Death-Ligand 1 Play an Indispensable Role in Effective Bacterial Elimination and Resolving Inflammation in Methicillin-Resistant Staphylococcus aureus Infection. Pathogens 2024, 13, 401. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).