Abstract
Clonostachys rosea, an entomopathogenic fungus that infects Cephalcia chuxiongica, is highly pathogenic and has significant potential for controlling the damage this pest causes to pine forests. To investigate the role of C. rosea secondary metabolites in fungal pathogenicity, we conducted toxicity assays using crude metabolite extracts. These assays evaluated the effects of different concentrations, larval developmental stages, and exposure methods on larval mortality. Gas chromatography–mass spectrometry (GC–MS) was subsequently employed to identify the chemical constituents of the crude extracts, and the toxicity of the identified compounds was assessed. The results showed that the crude extract at a concentration of 7.5 μg/mL exhibited the highest toxicity. Two hours post-treatment, the mortality rate of non-diapause larvae reached 65%, which was significantly higher than that of the diapause group. Moreover, contact toxicity was more lethal to C. chuxiongica larvae than oral exposure. A total of 23 compounds were identified from the crude extract, of which nine exhibited toxicity: 2-piperidone, hydrocinnamic acid, phenethyl alcohol, oleic acid, tryptophol, stearic acid methyl ester, myristic acid, dodecanoic acid, and benzeneacetic acid. Except for 2-piperidone, which showed low toxicity, the other eight compounds demonstrated notable contact toxicity against C. chuxiongica larvae. These findings confirm the insecticidal potential of C. rosea secondary metabolites and provide a valuable reference for the biological control of C. chuxiongica and other chewing insect pests.
1. Introduction
Because of its adaptability and wide distribution, pine has become a foundational species in China’s forest system. It not only constitutes an irreplaceable component of the national economy, but also plays a vital role in various sectors, including pharmaceutical research and development, forest chemical production, ecotourism, and construction engineering [1,2,3]. However, the larvae of Cephalcia chuxiongica (Hymenoptera: Pamphiliidae) pose a significant threat to pine forests, particularly to the needles of Pinaceae species. These larvae exhibit high host specificity, with a marked preference for species such as Pinus yunnanensis, P. armandii, and Keteleeria evelyniana (Gymnospermae: Pinaceae) [4,5]. The larvae initially spin silk to form webs at the base of pine needles, establishing protective shelters. They then sever needles, draw them into the webs for feeding, and continue to spin silk, constructing dense nests [6]. In severe infestations, the larvae can completely strip the trees of their needles, leaving only the petioles behind. This process results in large areas of scorched forest and significantly hinders pine growth [7]. The ecological damage caused by C. chuxiongica not only threatens forest health but also undermines the coordinated achievement of ecological, economic, and social benefits, which are central to China’s sustainable forestry development goals. Consequently, this species is a major target in China’s forestry pest management and poses a serious threat to forest sustainability [8,9].
Biological control has emerged as a leading strategy in modern pest management due to its ecological sustainability and effectiveness. By restoring natural ecological balance, it minimizes harm to non-target organisms and avoids the environmental and resistance-related issues commonly associated with chemical pesticides, thereby positioning itself as a preferred alternative. Currently, biological control efforts often focus on exploiting the insecticidal potential of entomopathogenic fungi (EPF), which exhibit strong virulence against a wide range of pest species. Under artificial culture conditions, many EPF strains are capable of producing diverse secondary metabolites with potent insecticidal properties [4,5,10]. These toxic metabolites play a critical role in the infection process and overall pathogenicity of EPF [11]. After penetrating the host cuticle, assisted by cuticle-degrading enzymes, EPF establish infection, proliferate by absorbing nutrients from the insect, and release toxic secondary metabolites that disrupt the host’s secretory and metabolic systems [5,12,13,14]. Numerous studies have shown that insect death during EPF infection is closely linked to these metabolic toxins. Pathological examinations of infected hosts often reveal that their death is primarily caused by the secondary toxic metabolites produced by the fungi [15,16,17]. Thus, toxic secondary metabolites are considered a major factor in the insecticidal activity of EPF.
Clonostachys rosea (Hipocreales: Bionectriaceae) is a widely used entomopathogenic fungus, capable of effectively controlling various pests by parasitizing the larvae, eggs, and female adults of nematodes as well as insects such as the mango hopper Amritodus atkinsoni (Hemiptera: Cicadelidae) and Xylosandrus germanus (Curculionidae: Scolytinae) [18,19,20,21]. This parasitism causes disease and mortality in the pests, thereby effectively preventing damage to plants. Consequently, C. rosea has become one of the primary research subjects in the study of biocontrol agents against plant pests. To date, at least 229 secondary metabolites have been isolated from C. rosea, mainly comprising 84 nitrogen-containing metabolites, 85 polyketides, 40 terpenoids, and 20 other compounds. These secondary metabolites exhibit significant cytotoxic and antibacterial activities, showing remarkable inhibitory effects on plant pathogens [22].
Given that high-virulence strains of C. rosea exhibit a rapid and potent lethal effect on C. chuxiongica larvae [23], this study aimed to investigate the insecticidal mechanism of its secondary metabolites. Fermentation broth of C. rosea was extracted and concentrated to obtain a crude metabolite extract. The effects of different concentrations, exposure durations, larval developmental stages, and administration methods on the mortality of C. chuxiongica larvae were systematically evaluated. In parallel, GC–MS was employed to identify the chemical constituents of the crude extracts. The insecticidal activities of the identified compounds were then assessed to determine their virulence against C. chuxiongica larvae. This integrated approach was designed to elucidate the mode of action of C. rosea secondary metabolites, identify the key pathogenic compounds involved, and provide a theoretical foundation for the development of biological control strategies against C. chuxiongica.
2. Materials and Methods
2.1. Experimental Materials
The entomopathogenic fungus Clonostachys rosea was isolated from the surface of naturally diseased and dead larvae of the sawfly Cephalcia chuxiongica. Detailed procedures for the isolation of this fungal strain, as well as preliminary research, have been previously published in our prior study [7,24]. Currently, C. rosea strain is preserved at the Laboratory of Forest Disaster Warning and Control, Yunnan Province.
The tested Cephalcia chuxiongica larvae used in this study were collected from the soil beneath pine forests in Beidaying Village, Qixing Township, Xundian County, Kunming City, Yunnan Province, China (25.30° N, 103.21° E; altitude: 1970 m). In this study, mature diapausing larvae and non-diapausing larvae at the 3rd–4th instars were collected in batches from March to November 2016, with approximately 500 larvae collected each batch. After collection, the larvae were quickly transferred to sterile soil under controlled laboratory conditions, with the relative humidity of the soil maintained at 30%, and fed pine needles regularly. Only healthy larvae that remained viable after 5 days of cultivation were selected for subsequent experiments. Larvae exhibiting damaged body surfaces, abnormal coloration (e.g., yellowing or darkening), fluid exudation, refusal to feed, sluggish movement, or curled immobility were excluded.
The culture medium used for fungal growth was Czapek-Dox Agar, composed of NaNO3 (2 g), K2HPO4 (1 g), KCl (0.5 g), MgSO4 (0.5 g), FeSO4 (0.01 g), and sucrose (30 g) per liter of distilled water. The medium was sterilized at 121 °C for 20 min.
2.2. Determination of Optimal Preparation Time for High-Virulence Fermentation Broth
A 0.1% (v/v) sterile Tween-80 solution was prepared by adding 5 mL of Tween 80 to 5000 mL of deionized water, followed by autoclaving at 115 °C for 30 min. After C.s rosea was cultured on solid medium plates for 20 days, aerial conidia produced on the medium surface were scraped into 50 mL sterile centrifuge tubes. An appropriate volume of sterile 0.1% (v/v) Tween-80 solution was added to each tube, and the mixture was vortexed for 3 min to form a conidial suspension. The suspension was filtered through six layers of sterile lens paper using a sterile funnel, and the filtrate was transferred to new sterile 50 mL centrifuge tubes. The concentration of the conidial suspension was standardized to 1 × 1010 conidia/mL and stored at 4 °C until use.
A conidial suspension of C. rosea at a concentration of 1.0 × 1010 conidia/mL was inoculated into the fermentation medium at a volume ratio of 5% (v/v) and incubated in a shaker at 25 °C and 200 rpm [24]. Fermentation broths were collected at 12, 24, 36, 48, 60, and 72 h post-inoculation. After centrifugation at 5000 rpm for 20 min, the supernatant was collected for bioassays. Non-diapause larvae were immersed for 1 min in fermentation broths maintained at 15 °C. Sterile water served as the control. Each treatment group consisted of 20 healthy larvae, with three biological replicates per group [25]. Treated larvae were transferred to 90-mm Petri dishes with controlled humidity and were regularly fed fresh Pinus yunnanensis needles. Larval mortality was recorded every 2 h.
2.3. Extraction of Metabolites from the Strain
C. rosea was cultured according to the method described in Section 2.2. Cultures were incubated until the optimal time point for producing high-virulence fermentation broth, as determined in Section 2.2. At this point, shaking incubation was terminated, and the fermentation broths were centrifuged at 5000 rpm for 20 min. The supernatant was reduced to half its original volume by rotary evaporation at 60 °C. Subsequently, twice the volume of 99% ethanol was added to the concentrated extract for alcohol precipitation, and the mixture was allowed to stand at room temperature for 24 h. After centrifugation at 4000 rpm for 20 min, the supernatant was collected, and ethanol was removed via rotary evaporation. Liquid–liquid extraction was performed with ethyl acetate (organic:aqueous = 2:1, v/v), repeated three times. The combined organic phase was concentrated and dried to yield the crude extract of fungal secondary metabolites.
2.4. Toxicity Determination of Crude Extracts from Strain Metabolites
2.4.1. Toxicity Determination at Different Concentrations
The obtained crude extract was weighed, and the extraction yield was calculated using the following formula:
Aqueous solutions of the crude metabolite extract were prepared at concentrations of 2.5 μg/mL, 5 μg/mL, and 7.5 μg/mL using sterile water. Non-diapause larvae were immersed in crude extract solutions at different concentrations for 1 min. Each treatment group contained 20 healthy larvae, with sterile water treatment serving as the control. Three biological replicates were performed for each treatment group. The post-treatment cultivation methods followed the procedures described in Section 2.2, and larval mortality was recorded every 2 h.
2.4.2. Toxicity Determination on Larvae at Different Developmental Stages
The crude extract of fungal metabolites was diluted to the concentration exhibiting the highest virulence, as determined in Section 2.4.1. C. chuxiongica larvae in both diapause and non-diapause stages were immersed in the crude extract solution for 1 min, with sterile water used as the control treatment. For feeding, twenty Pinus yunnanensis needles (each approximately 3 cm in length) were placed in each Petri dish at the beginning of the experiment. Additional needles were provided based on the number of surviving larvae, at a rate of one needle per larva per supplementation.
2.4.3. Determination of Virulence Under Different Toxicant Administration Methods
Group A (contact toxicity): Non-diapause C. chuxiongica larvae were starved for 3 h. The crude extract of fungal metabolites was diluted to the optimal virulence concentration determined in Section 2.4.1. At 15 °C, larvae were immersed in the aqueous crude extract solution for 1 min, with sterile water-treated larvae serving as controls. Each treatment consisted of 20 healthy larvae with three replicates. After treatment, larvae were transferred to humidity-controlled Petri dishes (30% relative humidity) and provided with 20 P. yunnanensis needles (3 cm in length) for feeding. Additional needles were supplemented based on the number of surviving larvae at a rate of one needle per larva per feeding. All larvae were maintained at room temperature and observed at 2-h intervals.
Group B (stomach toxicity): Non-diapause C. chuxiongica larvae were starved for 3 h. The crude extract was diluted to the optimal virulence concentration determined in Section 2.4.1. At 15 °C, P. yunnanensis needles were immersed in the extract for 30 min before being fed to the larvae, while needles soaked in sterile water for 30 min served as controls. Each treatment consisted of 20 healthy larvae with three biological replicates. After treatment, larvae were transferred to humidity-controlled Petri dishes and provided with additional needles based on survival rates (one needle per larva per feeding). All larvae were maintained at room temperature and observed at 2-h intervals.
2.5. Identification and Analysis of Chemical Components of Strain-Derived Metabolic Toxins
2.5.1. Component Identification
The crude metabolite extract was filtered through a membrane filter and dissolved in chromatographic-grade methanol. Chemical composition analysis was performed using GC–MS with an Agilent 7890B-5977A system (Agilent Technologies, Santa Clara, CA, USA) equipped with an HP-5MS capillary column (30 m × 0.32 mm i.d. × 0.25 μm film thickness). The GC temperature program was as follows: initial temperature of 120 °C held for 2 min, ramped to 250 °C at 20 °C/min, and held for 20 min. Injection was performed in split mode with a split ratio of 10:1, and the inlet temperature was set at 250 °C. Mass spectrometry was performed using an electron impact (EI) ion source at 70 eV. The ion source temperature was set at 250 °C, the interface temperature at 280 °C, the quadrupole temperature at 150 °C, and the solvent delay was 3 min. Full-scan mode was used with a mass range of 20–650 m/z.
2.5.2. Determination of the Insecticidal Activity of Toxic Compounds
The purified toxic compounds identified by GC-MS were dissolved as follows: except for phenethyl alcohol and 2-piperidone reference standards purchased from Tianjin Chemical Reagent Third Factory (Tianjin, China), which were directly dissolved in sterile water, all other reference compounds were first dissolved in dimethyl sulfoxide (DMSO) and then diluted with sterile water to concentrations of 0.05 mg/mL, 0.5 mg/mL, and 5 mg/mL. Non-diapause C. chuxiongica larvae were immersed in organic solutions at each concentration gradient for 1 min, with sterile water-treated larvae serving as controls. Each treatment consisted of 20 healthy larvae with three replicates. After treatment, larvae were transferred to humidity-controlled Petri dishes and provided with 20 P. yunnanensis needles (3 cm in length) for feeding. Observations were conducted at 2-h intervals, and needle supplementation was adjusted based on consumption rates. All larvae were maintained at room temperature, and mortality was recorded every 12 h.
The C. rosea conidial suspension (1.0 × 108 conidia/mL) was compared with the optimal lethal concentration of toxic compounds to evaluate the lethal efficacy of the conidial suspension against C. chuxiongica. Each treatment included three biological replicates with 20 larvae per replicate (following the same rearing protocol as above).
2.6. Statistical Analysis
Statistical analysis was performed using GraphPad Prism 9.5 and SPSS 26.0. After verifying that all datasets met the assumptions of normality (Shapiro–Wilk test) and homoscedasticity (Levene’s test) with p > 0.05, a two-way analysis of variance (ANOVA) with a significance threshold of p < 0.05 was employed to evaluate inter-sample differences, followed by Tukey’s HSD test for pairwise comparisons. To further dissect significant interaction effects between specific factor levels, Šidák’s and Tukey’s multiple comparisons tests were systematically applied. Figures were prepared using GraphPad Prism 9.5.
3. Results and Analysis
3.1. Comparison of the Toxicity of Crude Extracts from Strain Metabolites
3.1.1. Determination of Optimal Preparation Time for High-Virulence Fermentation Broth Analysis
Bioassay results evaluating the insecticidal activity of Clonostachys rosea fermentation broth against Cephalcia chuxiongica larvae indicated that the toxic potency of the broth peaked between 48 and 60 h post-inoculation. Based on this observation, the shaking incubation of the C. rosea fermentation culture was terminated after 60 h of liquid cultivation at 25 °C.
Following 60 h of submerged cultivation (25 °C, 200 rpm) of C. rosea in 6 L of medium, the mycelia were separated by filtration to obtain ~5.4 L of fermentation broth. The crude extract of C. rosea secondary metabolites, obtained through centrifugation, concentration, ethanol precipitation, liquid–liquid extraction, and drying, yielded 0.72 g of a dark brown, highly water-soluble paste, corresponding to an extraction efficiency of 133 mg/L.
3.1.2. Toxicity Assay of Crude Extracts from the Strain’s Metabolites
Comparative Analysis of Toxic Potency at Different Concentrations
Analysis of covariance revealed that both the crude extract concentration and treatment time had extremely significant effects on the mortality of Cephalcia chuxiongica larvae (p < 0.0001). Furthermore, the interaction between the crude extract concentration and treatment time also exerted an extremely significant influence on larval mortality (p < 0.0001). Throughout the experimental observation period, the control group consistently showed 0% cumulative mortality.
Analysis of cumulative larval mortality across different treatment durations and concentrations revealed distinct patterns. At a concentration of 2.5 μg/mL, C. chuxiongica larvae exhibited 0% cumulative mortality at all assessed time points. At 5 μg/mL, treatment duration had a significant effect on mortality, showing a progressive increase over time. No mortality was observed at 2 h post-treatment, while cumulative mortality reached 51.7% at 4 h and 100% by 10 h. Statistically significant differences were observed between all pairwise comparisons of treatment durations with 2.5 μg/mL, except between the 6-h and 8-h groups and between the 8-h and 10-h groups. At 7.5 μg/mL, a similar time-dependent increase in cumulative mortality was observed. Mortality reached 65% at 2 h and 90% at 4 h post-treatment. Compared with 2.5 μg/mL, no notable differences in cumulative mortality of larvae were detected among the 4-h, 6-h, 8-h, and 10-h groups at 7.5 μg/mL; however, all these time points showed markedly higher mortality than the 2-h group. Complete mortality was achieved by 8 h post-treatment (Figure 1).
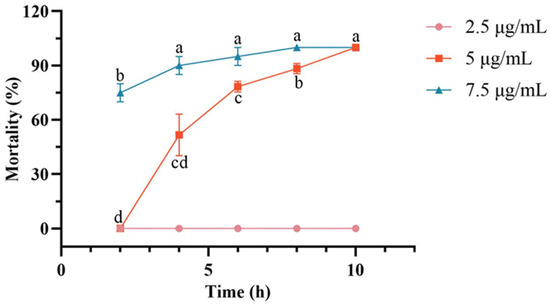
Figure 1.
Changes in cumulative mortality of Cephalcia chuxiongica larvae over the treatment time gradient of 2–10 h under different concentrations: 2.5 μg/mL, 5 μg/mL, and 7.5 μg/mL. Different letters (a, b, c, d) above the data points indicate significant differences at the p < 0.05 level, where distinct letters represent statistically significant differences between groups.
Comparative Analysis of Toxic Potency Against Larvae at Different Developmental Stages
Following a 1-min immersion in an aqueous solution of C. rosea crude extract at a concentration of 7.5 μg/mL, cumulative mortality of both non-diapause and diapause C. chuxiongica larvae was recorded. No mortality occurred in the control groups treated with sterile water, regardless of diapause status. In contrast, larvae exposed to the crude extract exhibited time-dependent increases in mortality.
In non-diapause larvae, exposure to the crude extract resulted in a statistically significant increase in cumulative mortality over time. While no significant differences were observed between the 4-h group and other time points, mortality at 6 h post-treatment was significantly higher than that at 2 h (65%), reaching 90% by 10 h post-treatment (Figure 2).
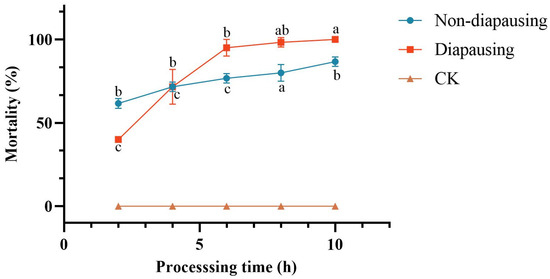
Figure 2.
Changes in mortality of Cephalcia chuxiongica larvae over processing time across different developmental stages: Different letters (a, b, c, d) above the data points indicate significant differences at the p < 0.05 level, where distinct letters represent statistically significant differences between groups.
In diapause larvae, exposure to the crude extract of C. rosea metabolites produced a cumulative mortality trend similar to that observed in the non-diapause group, characterized by a progressive increase with prolonged exposure duration. No significant differences in cumulative mortality were detected among the 6-h, 8-h, and 10-h treatment groups; however, mortality at these time points was significantly higher than at 4 h post-treatment. Cumulative mortality reached 95% at 6 h and 100% at 10 h. Additionally, mortality at 4 h post-treatment (76.67%) was significantly higher than at 2 h (41.67%).
For both non-diapause and diapause larvae, cumulative mortality increased progressively with treatment duration, although the rate and extent of change differed between the two groups). At 2 h post-treatment, the cumulative mortality of the non-diapause group (65%) was significantly higher than that of the diapause group (41.67%). By 4 h, mortality rates were comparable between the two groups. However, with extended exposure, the diapause group exhibited a more rapid increase in cumulative mortality, with significantly higher mortality than the non-diapause group at both 6 and 8 h post-treatment. By 10 h, although the cumulative mortality of the diapause group exceeded that of the non-diapause group, the difference was not statistically significant.
Determination of Toxic Potency Under Different Toxicant Administration Methods
Non-diapause C. chuxiongica larvae were selected as test subjects and exposed to two treatment modalities: contact toxicity and oral toxicity. Throughout the experimental period, both treatments resulted in a progressive increase in cumulative larval mortality, while no mortality was observed in the control group.
In C. chuxiongica larvae treated with the crude extract via the contact toxicity method, cumulative mortality increased significantly over time. Although no significant differences were observed between the 4-h group and other time points, cumulative mortality at 6 h post-treatment was significantly higher than at 2 h (65%), reaching 90% by 10 h post-treatment (Figure 3).
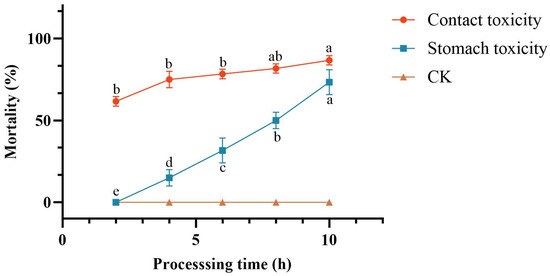
Figure 3.
Comparison of lethality rates between contact and oral toxicity treatments. Different letters (a, b, c, d) above the data points indicate significant differences at the p < 0.05 level, where distinct letters represent statistically significant differences between groups.
In C. chuxiongica larvae treated with the crude extract via the oral toxicity method, cumulative mortality also exhibited a significant upward trend with increasing exposure duration. No mortality was observed at 2 h post-treatment; however, larval mortality increased progressively over time, with significant differences in cumulative mortality recorded at each subsequent 2-h interval.
When comparing larval mortality under the two treatment methods (contact and oral toxicity), cumulative mortality exhibited a time-dependent increase in both groups; however, the rate and magnitude of mortality differed. Across all time points prior to 10 h, cumulative mortality under contact toxicity treatment was consistently and significantly higher than that under oral toxicity treatment. At 10 h post-treatment, although the cumulative mortality in the contact toxicity group (90%) remained higher than that in the oral toxicity group (73.3%), the difference was not statistically significant.
3.2. Identification of Chemical Constituents in Crude Extracts from Strain Metabolites
The chemical constituents of the crude extract derived from C. rosea secondary metabolites were analyzed using GC–MS. Chromatographic peaks were primarily detected within the retention time range of 5.348–23.980 min, yielding a total of 65 peaks. Based on GC–MS analysis and mass spectral library matching, 65 components were identified, of which 23 exhibited a match score ≥ 30% (Table 1). These 23 components were further screened using the Chemical Substances Toxicity Database (accessed on 10 December 2024, http://www.drugfuture.com/toxic/search.aspx), resulting in the identification of 9 major toxic compounds: phenethyl alcohol (7.27% of total toxic content), 2-piperidone (5.13%), benzeneacetic acid (3.13%), hydrocinnamic acid (5.6%), dodecanoic acid (1.43%), myristic acid (5.5%), tryptophol (4.2%), stearic acid methyl ester (1.1%), and oleic acid (66.63%).

Table 1.
Compounds detected in the crude extract of Clonostachys rosea using Gas Chromatography-Mass Spectrometry (GC-MS) analysis.
3.3. Insecticidal Activity Assay of Toxic Compounds
3.3.1. Determination of the Insecticidal Activity of Oleic Acid
Oleic acid at a concentration of 0.05 mg/mL exhibited no lethal effect on C. chuxiongica larvae. However, larval mortality increased with higher concentrations of oleic acid. At 0.5 mg/mL, more than 50% of larvae died within 108 h post-treatment, with cumulative mortality reaching 81.67% by 180 h and a calculated median lethal time (LT50) of 77 h. At 5 mg/mL, nearly 50% mortality was observed within the first 12 h, and complete mortality (100%) was achieved within 96 h of treatment (Table 2).
Table 2.
Mortality of Non-diapause Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Oleic Acid.
3.3.2. Determination of the Insecticidal Activity of Tryptophol
As shown in Table 3, tryptophol at a concentration of 0.05 mg/mL exhibited no toxic effect on C. chuxiongica larvae. However, mortality increased with higher tryptophol concentrations. At 0.5 mg/mL, cumulative mortality reached 75.00% after 180 h of treatment, with an LT50 of 83.3 h. At 5 mg/mL, 50% of the larvae died within 12 h, and mortality rose to 98.33% by 180 h.
Table 3.
Mortality of Non-diapause Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Tryptophol.
3.3.3. Determination of the Insecticidal Activity of Stearic Acid Methyl Ester
As shown in Table 4, stearic acid methyl ester at a concentration of 0.05 mg/mL exhibited negligible toxicity against C. chuxiongica larvae. In contrast, higher concentrations showed clear toxic effects, with mortality increasing in a concentration-dependent manner. At 0.5 mg/mL, cumulative mortality reached 75.00% after 180 h of treatment, with an LT50 of 110 h. At 5 mg/mL, more than 50% mortality was observed within 48 h, and cumulative mortality reached 98.33% by 180 h.
Table 4.
Mortality of Non-diapause Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Stearic Acid.
3.3.4. Determination of the Insecticidal Activity of Myristic Acid
As shown in Table 5, myristic acid at a concentration of 0.05 mg/mL exhibited negligible toxicity against C. chuxiongica larvae. In contrast, higher concentrations demonstrated significant toxicity, with mortality increasing in a concentration-dependent manner. At 0.5 mg/mL, cumulative mortality reached 73.33% after 180 h of treatment, with an LT50 of 80 h. At 5 mg/mL, over 50% mortality was observed within 60 h, and complete mortality (100%) was achieved by 144 h.
Table 5.
Mortality of Non-diapause Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Myristic Acid.
3.3.5. Determination of the Insecticidal Activity of Dodecanoic Acid
As shown in Table 6, dodecanoic acid at a concentration of 0.05 mg/mL exhibited negligible toxicity against C. chuxiongica larvae. In contrast, higher concentrations displayed clear toxic effects, with mortality increasing in a concentration-dependent manner. At 0.5 mg/mL, cumulative mortality reached 60.00% after 180 h of treatment, with an LT50 of 99.3 h. At 5 mg/mL, over 50% of the larvae died within 48 h, and complete mortality (100%) was achieved by 144 h.
Table 6.
Mortality of Non-diapause Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Dodecanoic Acid.
3.3.6. Determination of the Insecticidal Activity of 2-Piperidone
As shown in Table 7, 2-piperidone exhibited no detectable toxicity against C. chuxiongica larvae at any of the tested concentrations.
Table 7.
Mortality of Non-diapause Cephalcia chuxiongica Larvae Exposed to Different Concentrations of 2-Piperidinone.
3.3.7. Determination of the Insecticidal Activity of Phenethyl Alcohol
Phenethyl alcohol at a concentration of 0.05 mg/mL exhibited no toxic effect on C. chuxiongica larvae. However, higher concentrations displayed clear toxicity, with mortality increasing in a concentration-dependent manner. At 0.5 mg/mL, cumulative mortality reached 76.67% after 180 h of treatment, with an LT50 of 114 h. At 5 mg/mL, more than 50% of larvae died within 84 h of exposure (Table 8).
Table 8.
Mortality of Non-diapause Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Phenylethyl Alcohol.
3.3.8. Determination of the Insecticidal Activity of Benzeneacetic Acid
Benzeneacetic acid at a concentration of 0.05 mg/mL exhibited no toxicity against C. chuxiongica larvae. In contrast, higher concentrations showed clear toxic effects, with mortality increasing in a concentration-dependent manner. At 0.5 mg/mL, cumulative mortality reached 70.00% after 180 h of treatment, with an LT50 of 130.7 h. At 5 mg/mL, more than 50% mortality was observed within 60 h, and complete mortality (100%) was achieved by 84 h (Table 9).
Table 9.
Mortality of Non-diapause Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Benzeneacetic Acid.
3.3.9. Determination of the Insecticidal Activity of Hydrocinnamic Acid
Hydrocinnamic acid at a concentration of 0.05 mg/mL exhibited no toxicity against C. chuxiongica larvae. However, higher concentrations demonstrated clear toxic effects, with mortality increasing in a concentration-dependent manner. At 0.5 mg/mL, cumulative mortality reached 80.00% after 180 h of treatment, with an LT50 of 92 h (Table 10).
Table 10.
Mortality of Non-diapause Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Hydrocinnamic Acid.
3.3.10. Comparison of the Toxicity Between Identified Compounds and Conidial Suspensions
A comparative analysis of insecticidal activity was conducted between each identified toxic compound at a concentration of 0.5 mg/mL and a C. rosea conidial suspension at 1.0 × 108 conidia/mL, as shown in Figure 4. The results indicated that the 2-piperidone treatment group exhibited no significant difference from the control group, confirming its lack of toxicity. In contrast, the remaining eight compounds—hydrocinnamic acid, phenethyl alcohol, oleic acid, tryptophol, stearic acid methyl ester, myristic acid, dodecanoic acid, and benzeneacetic acid—demonstrated insecticidal activity against C. chuxiongica larvae. However, their toxic effects were markedly lower than those obs erved with the C. rosea conidial suspension. Notably, the difference in lethality between the conidial suspension and each of the toxic compounds became highly significant after 60 h of treatment. For the analysis of the lethal ability of C. rosea against C. chuxiongica, please refer to Supplementary Materials Table S1.
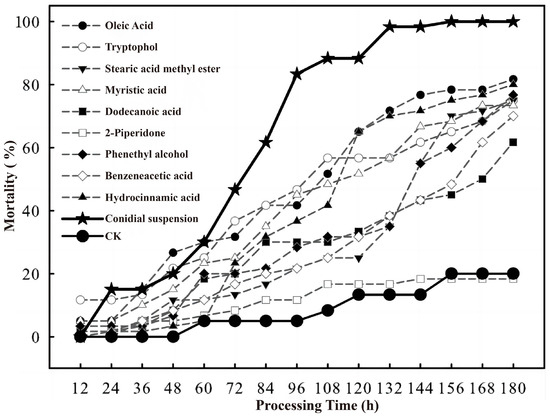
Figure 4.
Comparison of mortality of Cephalcia chuxiongica larvae exposed to different toxic compounds (oleic acid, tryptophol, stearic acid methyl ester, myristic acid, dodecanoic acid, 2-piperidone, phenethyl alcohol, benzeneacetic acid and hydrocinnamic acid), conidial suspension of Clonostachys rosea, and sterile water treatment (CK) at different treatment times (12–180 h).
4. Discussion
Analysis of Cephalcia chuxiongica larvae mortality exposed to Clonostachys rosea crude extracts showed that a 7.5 μg/mL aqueous solution was most toxic: 65% died within 2 h, and 100% after 8 h. A previous study on Beauveria brongniartii (Ascomycota: Cordycipitaceae) against Dendrolimus tabulaeformis (Lepidoptera: Lasiocampidae) reported that the cumulative mortality rate of D. tabulaeformis was 71.3% after 7 days of treatment with the highest toxicity concentration (550 μg/mL) in the toxicity assay of crude extracts of metabolic toxins from B. brongniartii against D. tabulaeformis [26]. In contrast, C. rosea’s crude secondary metabolite extract was more toxic, showing advantages in low effective concentration, rapid action, and high efficacy, supporting its potential as a better alternative to B. brongniartii-based bioinsecticides.
Furthermore, the larvae exhibited pronounced twisting behavior when exposed to the crude extract, which is consistent with the discoveries of Baggio-Deible [27] and Haiyang Wang [28]. These findings are consistent with previous studies. A comparison of the toxicity of the crude extract of the strain’s metabolites against larvae at various developmental stages revealed that it is toxic to both diapausing and non-diapausing C. chuxiongica larvae. The toxicity of the substance was more potent against non-diapause larvae within four hours of treatment than it was against diapause larvae. However, the toxicity of the substance against diapause larvae became more potent after four hours of treatment. The physiological status of the larvae (metabolic activity, barrier function, detoxification ability) and the mechanism of action of the toxin are the primary factors contributing to the time-dependent difference in toxicity. The toxin’s absorption and action are expedited by the high metabolic activity of non-diapause larvae in the early stage, whereas the low metabolic state of diapause larvae results in long-term accumulation and slow toxin clearance, ultimately leading to stronger toxicity in the later stage [28,29,30,31]. This phenomenon also implies that application timing and dosage of fungal crude extracts should be tailored to different insect developmental stages, to enhance biological control efficiency.
In addition, the results showed that contact toxicity caused more larval deaths than oral toxicity in C. chuxiongica. The main reason for the higher efficiency of the contact toxicity method may be that it bypasses the defensive barrier of the larval digestive tract, utilizing the permeability of the cuticle to achieve rapid invasion and action of toxins. In contrast, the oral toxicity method is limited by feeding behavior, digestive barriers, and detoxification mechanisms, reducing toxin efficiency [32,33]. This suggests that in biological control of this pest, prioritizing contact-type fungal preparations, such as conidial suspensions and metabolites capable of penetrating the body wall, may more effectively enhance control efficacy.
During the experiment, it was observed that C. chuxiongica larvae in Petri dishes died and exhibited body stiffening even without feeding on pine needles treated with the crude extract of C. rosea metabolites. This phenomenon may be attributed to the volatility of certain C. rosea metabolites, which can exert toxic effects through fumigation. In the present study, GC–MS analysis of the crude extract of C. rosea secondary metabolites identified nine toxic compounds. Although these compounds exhibited lethal effects on C. chuxiongica larvae at a concentration of 0.5 mg/mL, their toxicity was lower than that of the C. rosea conidial suspension (1.0 × 108 conidia/mL). Shichuang Ma [34] evaluated the insecticidal activity of toxic compounds against Myzus persicae, Aphis pomi, Brevicoryne brassicae (Hemiptera: Aphididae), and Bemisia tabaci (Hemiptera: Aleyrodidae), and found that the derivatives exhibited notable contact and systemic effects. Similarly, Cecilia Labbé [35] isolated 2-phenylethyl acetate and 2-phenylethyl benzoate from the extract of Balantiopsis cancellata (Bryophyta: Balantiopsaceae), a Chilean plant species, and identified their antifeedant activity against Spodoptera frugiperda (Lepidoptera: Noctuidae). These previous findings collectively support the conclusion that toxic compounds induce rapid insect mortality through disruption of insect-specific metabolic pathways and physiological mechanisms. Building on this foundation, the toxin action mechanism elucidated in the present study provides a critical theoretical basis for developing novel green pesticides with high target specificity and superior environmental compatibility, thereby offering substantial practical value for advancing integrated pest management (IPM) and promoting sustainable agricultural practices.
Previous studies have demonstrated that toxic compounds can rapidly induce insect mortality by disrupting insect-specific metabolic pathways and other physiological mechanisms. However, Moraes [36] compared the efficacy of Bacillus thuringiensis (Bacillota: Bacillaceae) formulations with that of chemical insecticides, such as diphenylurea and deltamethrin, against Plutella xylostella (Lepidoptera: Plutellidae). The findings indicated that B. thuringiensis exhibited superior control over third-instar larvae compared to the chemical agents. Similarly, Jiang Shuai et al. [37] conducted toxicity assays using crude toxin extracts from Amanita muscaria (Basidiomycota: Amanitaceae) and Amanita verna (Basidiomycota: Amanitaceae) and confirmed their insecticidal activity against Gryllotalpa unispina (Orthoptera: Gryllotalpidae). These studies highlight the promising potential of microbial and natural compound-based agents in pest management strategies. Parajuli [38] also reported that fungal biopesticides, including Beauveria bassiana (Ascomycota: Cordycipitaceae) and Metarhizium anisopliae (Ascomycota: Clavicipitaceae), are highly effective in controlling a wide range of insect pests, such as aphids, whiteflies, and locusts. These fungal agents function through mechanisms such as cuticle penetration and the production of toxic secondary metabolites. Compared to conventional chemical pesticides, fungal biopesticides are more environmentally friendly and represent a sustainable alternative for pest management. Thus, microbial formulations offer broader modes of action and present a promising strategy for advancing sustainable agricultural practices.
Investigating the crude extract of insect-derived secondary metabolites from C. rosea fermentation broth facilitates the elucidation of its insecticidal pathways and mechanisms of action. Such studies provide a theoretical basis for the application of C. rosea in biological pest control and contribute to assessing its safety for humans and animals. Among the identified metabolites, benzeneacetic acid—an important organic intermediate—holds broad application potential, particularly in the pharmaceutical and agrochemical industries [39,40,41]. However, due to the complexity of its synthetic routes, it remains a relatively scarce resource in the domestic pharmaceutical market. Additionally, stearic acid methyl ester is widely used in the synthesis of surfactants, lubricants, and other industrial chemicals [42], while tryptophol serves as a valuable reagent in organic synthesis [43]. Given that C. rosea can stably produce these metabolites through a fermentation process that is both scalable and easily regulated, it presents a sustainable and renewable source of raw materials for industrial applications, offering new opportunities for green biomanufacturing in related sectors.
Based on the above findings, a canopy spraying system is recommended for forest application. This system utilizes the fumigation effect of volatile compounds to penetrate pine needle layers, enabling three-dimensional control of concealed pests. Subsequent research will focus on optimizing large-scale fermentation processes to increase target compound yields, and verifying sustained control efficacy at different concentration gradients through field trials. This green pest control system not only provides a new solution for pine forest IPM, but also pioneers a sustainable development model integrating biological control with green manufacturing.
5. Conclusions
By extracting and purifying the crude metabolites of Clonostachys rosea, toxicity assays were conducted on Cephalcia chuxiongica larvae across different concentrations, developmental stages, and exposure methods. The results indicated that the crude extract at a concentration of 7.5 μg/mL exhibited the highest insecticidal activity, achieving 100% larval mortality within 8 h post-treatment. When larvae at different developmental stages were exposed to the same concentration, the non-diapause group showed a significantly higher mortality rate (65%) at 2 h compared to the diapause group (41.67%). However, after 10 h of exposure, cumulative mortality reached 90% in the non-diapause group, while 100% mortality was observed in the diapause group, indicating a temporal difference in susceptibility. In terms of exposure methods, contact toxicity was found to be more effective than oral toxicity in inducing larval death. The metabolic components of the C. rosea crude extract were identified using GC-MS, resulting in the detection of 23 compounds, among which 9 exhibited significant toxicity. Except for 2-piperidone, which showed low toxicity, eight compounds—hydrocinnamic acid, phenethyl alcohol, oleic acid, tryptophol, stearic acid methyl ester, myristic acid, dodecanoic acid, and benzeneacetic acid—demonstrated contact toxicity against C. chuxiongica larvae. Among these, myristic acid exhibited the strongest insecticidal activity, although its efficacy was still inferior to that of the C. rosea conidial suspension at 1.0 × 108 conidia/mL.
This study confirms that the secondary metabolites of C. rosea exert lethal effects on C. chuxiongica, highlighting their considerable potential for pest management. These findings provide a robust theoretical foundation for the development of novel biocontrol strategies and the enhancement of pest control precision. In subsequent investigations, we aim to identify the specific molecular targets of C. rosea secondary metabolites in C. chuxiongica, with the goal of elucidating the underlying molecular and physiological mechanisms governing their insecticidal activity. Additionally, we will optimize the formulation and application parameters of this biocontrol agent to minimize unintended ecological impacts on non-target organisms, ensuring its sustainable integration into forest pest management strategies.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/microorganisms13102289/s1, Table S1: Mortality of Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Oleic Acid; Table S2: Mortality of Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Tryptophol; Table S3: Mortality of Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Stearic Acid; Table S4: Mortality of Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Myristic Acid; Table S5: Mortality of Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Dodecanoic Acid; Table S6: Mortality of Cephalcia chuxiongica Larvae Exposed to Different Concentrations of 2-Piperidinone; Table S7: Mortality of Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Phenylethyl Alcohol; Table S8: Mortality of Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Benzeneacetic Acid; Table S9: Mortality of Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Hydrocinnamic Acid; Table S10: Mortality of Cephalcia chuxiongica Larvae Exposed to Different Concentrations of Conidial Suspension of Clonostachys rosea.
Author Contributions
Conceptualization, J.L. (Jian Liu) and H.L.; Data curation, J.L. (Jian Liu); Formal analysis, J.L. (Jian Liu) and Y.S.; Funding acquisition, J.L. (Junjia Lu), Y.M. and Y.L.; Investigation, J.L. (Junjia Lu) and J.L. (Jian Liu); Methodology, Y.S.; Project administration, Y.L.; Resources, J.L. (Jian Liu) and H.L.; Software, J.L. (Junjia Lu) and J.L. (Jian Liu); Validation, J.L. (Jian Liu); Visualization, J.L. (Jian Liu); Writing—original draft, J.L. (Jian Liu) and Y.S.; Writing—review & editing, J.L. (Jian Liu). All authors have read and agreed to the published version of the manuscript.
Funding
This work is funded by the National Natural Science Foundation of China (32060695), the Yunnan Province Agricultural Joint Program (202301BD070001-091).
Institutional Review Board Statement
“The animal study protocol was approved by the Ethics Committee of Southwest Forestry University (6 June 2023 of approval).” for studies involving animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data from this trial can be found in the document and additional information package. Reagents, larvae and microbial materials, as well as data sets used, created, and analysed during this work, are available from the reporting author upon request.
Acknowledgments
We would like to express our gratitude to the Chinese National Natural Science Foundation and Yunnan Province Agricultural Joint Program for their support in this trial. I would like to thank my fellow students Huali Li and Yajiao Sun for their assistance during the experimental process; thank Junjia Lu and Yonghe Li for their support in terms of experimental materials and equipment, as well as their efforts in revising the manuscript; and also thank Yunqiang Ma for providing financial support.
Conflicts of Interest
The authors declare no conflict of interest. All authors hereby certify that they are not affiliated with, nor involved in, any organization or entity that holds either a financial or non-financial interest in the subject matter and materials presented in this manuscript.
References
- Moctezuma López, G.; Flores, A. Economic importance of pine (Pinus spp.) as a natural resource in Mexico. Rev. Mex. De Cienc. For. 2020, 11, 161–185. [Google Scholar] [CrossRef]
- Mullin, T.; Andersson, B.; Bastien, J.-C.; Beaulieu, J.; Burdon, R.; Dvorak, W.; King, J.; Kondo, T.; Krakowski, J.; Lee, S. Economic Importance, Breeding Objectives and Achievements; Taylor: Boca Raton, FL, USA, 2011; pp. 40–127. [Google Scholar] [CrossRef]
- Peter, G.F. Southern Pines: A Resource for Bioenergy; Springer: New York, NY, USA, 2008; pp. 421–449. [Google Scholar] [CrossRef]
- Qu, S.-L.; Li, S.-S.; Li, D.; Zhao, P.-J. Metabolites and their bioactivities from the genus Cordyceps. Microorganisms 2022, 10, 1489. [Google Scholar] [CrossRef]
- San Juan-Maldonado, N.; Rodrí, S.; Angel-Cuapio, A.; Vá, N.; Barranco-Florido, J.E. Secondary Metabolites of Entomopathogenic Fungi, Biological Alternative for the Control of Agricultural Pests and Disease: Present and Perspectives. Adv. Enzyme Res. 2024, 12, 1–15. [Google Scholar] [CrossRef]
- Zeng, T.; Lu, J.; Xu, R.; Fan, J.; Li, Y. The Culture Conditions and Sporulation Characteristics of 2 Entomopathogenic Fungi with Cephlica chuxiongica. J. Southwest For. Univ. 2018, 38, 126–133. [Google Scholar] [CrossRef]
- Lu, J.; Liu, J.; Li, H.; Sun, Y.; Liu, S.; Wang, M.; Li, Y. Toxicity Assay and Pathogenic Process Analysis of Clonostachys rogersoniana Infecting Cephalcia chuxiongica. Microorganisms 2025, 13, 709. [Google Scholar] [CrossRef]
- Qin, Y.; Liu, X.; Peng, G.; Xia, Y.; Cao, Y. Recent advancements in pathogenic mechanisms, applications and strategies for entomopathogenic fungi in mosquito biocontrol. J. Fungi. 2023, 9, 746. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Keyhani, N.O.; Xia, Y.; Xie, J. The potential and limitations of entomopathogenic fungi as biocontrol agents for insect pest management. Entomol. Gen. 2024, 44, 797–811. [Google Scholar] [CrossRef]
- Zhang, L.; Fasoyin, O.E.; Molnár, I.; Xu, Y. Secondary metabolites from hypocrealean entomopathogenic fungi: Novel bioactive compounds. Nat. Prod. Rep. 2020, 37, 1181–1206. [Google Scholar] [CrossRef]
- Wei, G.; Lai, Y.; Wang, G.; Chen, H.; Li, F.; Wang, S. Insect pathogenic fungus interacts with the gut microbiota to accelerate mosquito mortality. Proc. Natl. Acad. Sci. USA 2017, 114, 5994–5999. [Google Scholar] [CrossRef]
- Bava, R.; Castagna, F.; Piras, C.; Musolino, V.; Lupia, C.; Palma, E.; Britti, D.; Musella, V. Entomopathogenic fungi for pests and predators control in beekeeping. Vet. Sci. 2022, 9, 95. [Google Scholar] [CrossRef]
- De Carolina Sánchez-Pérez, L.; Barranco-Florido, J.E.; Rodríguez-Navarro, S.; Cervantes-Mayagoitia, J.F.; Ramos-López, M.Á. Enzymes of entomopathogenic fungi, advances and insights. Adv. Enzyme Res. 2014, 2, 65–76. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, X.; Eleftherianos, I.; Mohamed, A.; Bastin, A.; Keyhani, N.O. Cross-talk between immunity and behavior: Insights from entomopathogenic fungi and their insect hosts. FEMS Microbiol. Rev. 2024, 48, fuae003. [Google Scholar] [CrossRef]
- Ma, M.; Luo, J.; Li, C.; Eleftherianos, I.; Zhang, W.; Xu, L. A life-and-death struggle: Interaction of insects with entomopathogenic fungi across various infection stages. Front. Immunol. 2024, 14, 1329843. [Google Scholar] [CrossRef]
- Proctor, R.H.; McCormick, S.P.; Kim, H.-S.; Cardoza, R.E.; Stanley, A.M.; Lindo, L.; Kelly, A.; Brown, D.W.; Lee, T.; Vaughan, M.M. Evolution of structural diversity of trichothecenes, a family of toxins produced by plant pathogenic and entomopathogenic fungi. PLoS Pathog. 2018, 14, e1006946. [Google Scholar] [CrossRef]
- Rustiguel, C.B.; Fernández-Bravo, M.; Guimarães, L.H.S.; Quesada-Moraga, E. Different strategies to kill the host presented by Metarhizium anisopliae and Beauveria bassiana. Can. J. Microbiol. 2018, 64, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, M.E.; Braga, F.R.; Gives, P.M.d.; Uriostegui, M.A.M.; Reyes, M.; Soares, F.E.d.F.; Carvalho, L.M.d.; Rodrigues, F.B.; Araújo, J.V.d. Efficacy of Clonostachys rosea and Duddingtonia flagrans in reducing the Haemonchus contortus infective larvae. BioMed Res. Int. 2015, 2015, 474879. [Google Scholar] [CrossRef] [PubMed]
- Tamta, A.K.; Pandey, R.; Sharma, J.R.; Rai, R.; Barman, M.; MG, D.; Mitra, D.; Mohapatra, P.K.D.; Sami, R.; Al-Mushhin, A.A. First record of Clonostachys rosea (Ascomycota: Hypocreales) Entomopathogenic fungus in the mango hopper Amritodus atkinsoni (Hemiptera: Cicadellidae). Pathogens 2022, 11, 1447. [Google Scholar] [CrossRef] [PubMed]
- Pastirčáková, K.; Baková, K.; Adamčíková, K.; Barta, M.; Lalík, M.; Pavlík, M.; Kunca, A.; Galko, J.; Pastirčák, M. Fungi associated with ambrosia beetle Xylosandrus germanus in Slovakia. Biologia 2024, 79, 2387–2400. [Google Scholar] [CrossRef]
- Ahmed, M.; Laing, M.; Nsahlai, I. A new control strategy for nematodes of sheep using chlamydospores of a fungus, Clonostachys rosea f. rosea, and an ethanolic extract of a plant, Ananas comosus. Biocontrol Sci. Technol. 2014, 24, 860–871. [Google Scholar] [CrossRef]
- Han, P.; Zhang, X.; Xu, D.; Zhang, B.; Lai, D.; Zhou, L. Metabolites from Clonostachys fungi and their biological activities. J. Fungi 2020, 6, 229. [Google Scholar] [CrossRef]
- Lu, J.; Xia, J.; Zhuang, F.; Xu, R.; Zeng, T.; Li, Y. Identification and biological characterization of mycoparasitic fungus Clonostachys rosea isolated from sawfly Cephalcia chuxiongica. J. Plant Prot. 2018, 45, 622–631. [Google Scholar]
- Idrees, A.; Qadir, Z.A.; Akutse, K.S.; Afzal, A.; Hussain, M.; Islam, W.; Waqas, M.S.; Bamisile, B.S.; Li, J. Effectiveness of entomopathogenic fungi on immature stages and feeding performance of fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae. Insects 2021, 12, 1044. [Google Scholar] [CrossRef] [PubMed]
- Mani, K.; Vitenberg, T.; Khatib, S.; Opatovsky, I. Effect of entomopathogenic fungus Beauveria bassiana on the growth characteristics and metabolism of black soldier fly larvae. Pestic. Biochem. Physiol. 2023, 197, 105684. [Google Scholar] [CrossRef] [PubMed]
- Fan JinHua, F.J.; Xie YingPing, X.Y.; Xue JiaoLiang, X.J.; Liu Rui, L.R. The effect of Beauveria brongniartii and its secondary metabolites on the detoxification enzymes of the pine caterpillar, Dendrolimus tabulaeformis. J. Insect Sci. 2013, 13, 44. [Google Scholar] [CrossRef] [PubMed]
- Baggio-Deibler, M.V.; da Costa Ferreira, M.; Monteiro, A.C.; de Souza-Pollo, A.; Lemos, M.V.F. Management of the American cockroach’s oothecae: The potential of entomopathogenic fungi control. J. Invertebr. Pathol. 2018, 153, 30–34. [Google Scholar] [CrossRef]
- Wang, H.; Peng, H.; Li, W.; Cheng, P.; Gong, M. The toxins of Beauveria bassiana and the strategies to improve their virulence to insects. Front. Microbiol. 2021, 12, 705343. [Google Scholar] [CrossRef]
- Chakraborty, S.; Schuster, S. How plant toxins cause early larval mortality in herbivorous insects: An explanation by modeling the net energy curve. Toxins 2024, 16, 72. [Google Scholar] [CrossRef]
- Genersch, E.; Ashiralieva, A.; Fries, I. Strain-and genotype-specific differences in virulence of Paenibacillus larvae subsp. larvae, a bacterial pathogen causing American foulbrood disease in honeybees. Appl. Environ. Microbiol. 2005, 71, 7551–7555. [Google Scholar] [CrossRef]
- Skrobek, A.; Butt, T.M. Toxicity testing of destruxins and crude extracts from the insect-pathogenic fungus Metarhizium anisopliae. FEMS Microbiol. Lett. 2005, 251, 23–28. [Google Scholar] [CrossRef]
- Sakulpanich, A.; Phayakkaphon, A.; Ounklong, K.; Sommanat, J.; Samung, Y.; Srisawat, R.; Ruangsittichai, J. Evaluation of Stemona collinsiae root extracts for topical cockroach control: Adulticidal, nymphicidal, and chemical distribution analysis. Toxicon X 2025, 26, 100225. [Google Scholar] [CrossRef]
- Yang, Y.; Jiang, Q.; Peng, M.; Zhou, Z.; Du, X.; Yin, M.; Shen, J.; Yan, S. A star polyamine-based nanocarrier delivery system for enhanced avermectin contact and stomach toxicity against green peach aphids. Nanomaterials 2022, 12, 1445. [Google Scholar] [CrossRef]
- Ma, S.; Jiang, W.; Hu, Y.; Wang, Q.; Wu, W.; Shi, B. Synthesis, crystal structure, and insecticidal activity of steroidal N-piperidone. J. Agric. Food Chem. 2022, 70, 1467–1476. [Google Scholar] [CrossRef]
- Labbé, C.; Faini, F.; Villagrán, C.; Coll, J.; Rycroft, D.S. Antifungal and insect antifeedant 2-phenylethanol esters from the liverwort Balantiopsis cancellata from Chile. J. Agric. Food Chem. 2005, 53, 247–249. [Google Scholar] [CrossRef]
- Moraes, C.P.d.; Foerster, L.A. Toxicity and residual control of Plutella xylostella L. (Lepidoptera: Plutellidae) with Bacillus thuringiensis Berliner and insecticides. Cienc. Rural 2012, 42, 1335–1340. [Google Scholar] [CrossRef]
- Jiang ShuaiZhang, J.S.; Yang ShaoBin, Y.S. Toxicity of crude toxin from two kinds of Amanita to Gryllotalpa unispina. Guizhou Agric. Sci. 2014, 42, 120–122. [Google Scholar]
- Parajuli, S.; Shrestha, J.; Subedi, S.; Pandey, M. Biopesticides: A sustainable approach for pest management: Biopesticides in sustainable pest management. SAARC J. Agric. 2022, 20, 1–13. [Google Scholar] [CrossRef]
- Cook, S.D. An historical review of phenylacetic acid. Plant Cell Physiol. 2019, 60, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Jiao, M.; He, W.; Ouyang, Z.; Shi, Q.; Wen, Y. Progress in structural and functional study of the bacterial phenylacetic acid catabolic pathway, its role in pathogenicity and antibiotic resistance. Front. Microbiol. 2022, 13, 964019. [Google Scholar] [CrossRef]
- Perez, V.C.; Zhao, H.; Lin, M.; Kim, J. Occurrence, function, and biosynthesis of the natural auxin phenylacetic acid (PAA) in plants. Plants 2023, 12, 266. [Google Scholar] [CrossRef]
- Lu, Q.; Liu, T.; Wang, N.; Dou, Z.; Wang, K.; Zuo, Y. Nematicidal effect of methyl palmitate and methyl stearate against Meloidogyne incognita in bananas. J. Agric. Food Chem. 2020, 68, 6502–6510. [Google Scholar] [CrossRef]
- Palmieri, A.; Petrini, M. Tryptophol and derivatives: Natural occurrence and applications to the synthesis of bioactive compounds. Nat. Prod. Rep. 2019, 36, 490–530. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).