Abstract
Epichloë endophytes are mutualistic associates with grasses, conferring host plants with enhanced competitiveness, improved stress tolerance, and increased ecological dominance. Epichloë can produce any of several classes of bioactive alkaloids, of which indole-diterpenes and ergot alkaloids are toxic to invertebrate and mammalian herbivores; peramine acts as an insect-feeding deterrent; and loline alkaloids possess potent insecticidal activity. Here, it was characterized as Epichloë endophytes inhibiting the Elymus species, El. cylindricus, and El. tangutorum from the Qinghai–Tibet Plateau, China. Based on the results of four types of alkaloid synthesis genes, the 30 isolates were divided into five types; they exhibited distinct alkaloid synthesis capabilities, highlighting intraspecific diversity within E. bromicola regarding its alkaloid-producing potential. Considering the toxicity of these isolates to the safety of herbivorous livestock, the above five types of isolates can be divided into two categories. Category I includes five animal-safe strains of type V, which do not produce alkaloids. Category II includes the remaining 25 strains, which could produce indole-diterpene (paspaline) and/or ergot alkaloids (chanoclavine I, D-lysergic acid, ergovaline) that are toxic to herbivorous livestock. Morphology and phylogenetic analysis confirmed all 30 isolates were Epichloë bromicola; mating type gene detection shows that all belonged to mating type A. Overall, this study has laid a solid foundation for the scientific and rational utilization of Epichloë endophyte resources. Furthermore, the presence of ergovaline in El. cylindricus and El. tangutorum poses a potential concern for livestock managers who conduct grazing.
1. Introduction
The Epichloë genus (Clavicipitaceae, Ascomycota) are a class of fungal endophytes that typically establish mutualistic symbiotic relationships with cool-season grasses, and they can complete all or part of their life without causing obvious disease symptoms in the host [1,2]. Many cool-season grasses often have symbiotic relationships with Epichloë. The Poaceae family is the only recorded host of Epichloë endophytes. Up to now, Epichloë endophytes have been found in more than 300 species of grasses in 80 genera worldwide [3,4,5,6,7]. These endophytes are found in the intercellular spaces of aerial tissues of hosts [3]. Species of the Epichloë genus that sexually reproduce are capable of forming fruiting bodies known as stromata, which envelop the developing inflorescences and inhibit seed formation—resulting in “choke” diseases in host plants, and consequently facilitating horizontal transmission. However, some can colonize the host in an asymptomatic state and spread vertically; the asexual reproductive form previously classified as a species of Neotyphodium spp. is still benign and grows into the developing ovules, spreading through the seeds of the mother plant lineage. Some Epichloë species can be facultative in their dispersal, using both seeds and ascospores for transmission [8,9]. However, vertical transmission remains the main mode of dispersal for Epichloë species [10]. To date, based on morphological, host specificity and phylogenetic analyses, more than 50 species have been described. The phylogenetic analysis of the intron-rich sequences from the translation elongation factor 1-a gene (tefA) and the β-tubulin gene (tubB) has been employed to substantiate the evolutionary relationship among Epichloë species.
Epichloë form reciprocal symbionts with host plants, host plants provide the living environment and nutrients for Epichloë, and in return, some Epichloë are known to provide fitness benefits to their host plants including drought, salinity, low temperature, flooding, heavy metals tolerance, disease resistance, enhance competitiveness, improve survival ability [2,11], and protection against pests and mammalian herbivory through the production of alkaloid compound [12,13,14]. The interaction between symbionts is mainly played by secondary metabolites [15]. Hence, these secondary metabolites play an important role in the symbiotic relationship between host plants and Epichloë endophyte, of which alkaloids account for nearly half [16]. As early as 1997, after Bacon et al. found that alkaloids of Epichloë could cause livestock poisoning, researchers paid extensive attention to the alkaloids and thus conducted a series of studies [17]. Common alkaloids can be divided into four main groups: ergot alkaloids (such as ergovaline), indole-diterpenes (such as lolitrems B), bispyrrolidines (loline), and pyrrolipyrazines (peramine). Peramine is an insect-feeding deterrent, whereas loline has potent insecticidal activity. Indole-diterpenes (lolitrems B) and ergot are most commonly documented for being toxic to vertebrates, causing ryegrass staggers and fescue toxicosis, respectively [18,19]. Based on the completion of whole genome sequencing, four major classes of alkaloid-synthesis-related genes have been elucidated. The ppzA (formerly known as perA) gene located at the PER locus is essential for the biosynthesis of peramine [20,21]. However, the loci required for the production of loline, indole-diterpenoids, and ergot alkaloids are all complex gene clusters containing large stretches of related repetitive elements [22,23,24,25]. The diversity of alkaloids produced by metabolic pathways depends on the genes present in the genome, and the presence of functional domains in biosynthetic enzymes. Therefore, the presence or absence of certain genes and the sequences of the proteins they encode can be used to predict the ability of Epichloë to produce alkaloids or related intermediates [26,27,28]. This prediction represents a cost-effective and rapid method for screening the individuals of large Epichloë–grass symbiotic populations likely to be useful in forage agriculture [28]. This provides a theoretical basis for fully utilizing the alkaloid characteristics of Epichloë endophytes [29].
The diversity of alkaloids has promoted the development of novel Epichloë, which do not produce toxic alkaloids harmful to vertebrates. These novel Epichloë have been integrated into the biological breeding program to inoculate Epichloë-free plants with isolates that only synthesize insect-resistant alkaloids, without synthesizing alkaloids that cause toxicity to livestock, thereby establishing a new animal-safe Epichloë–host symbiotic association [30,31]. The goal is to accelerate germplasm innovation by leveraging multiple stress-resistance enhancement traits of Epichloë for their host and their vertical transmission characteristics, while with nil or much reduced (acceptable and manageable) adverse impacts on livestock [32]. This method aims to transfer the beneficial traits conferred by Epichloë from the natural host to the new host. However, the success of this inoculation process depends on various factors, including the inoculation technique, the Epichloë isolate, host genotype, and host specificity [33]. Consequently, the identification and exploration of Epichloë isolates’ characteristics is crucial for advancing this research. Through this process, a number of novel Epichloë isolates have been delivered and are now commercially used in USA and New Zealand, such as AR1TM, AR37TM, Endo5TM, and NEATM endophyte for ryegrass, E34TM, AR542TM, MaxQTM, and MaxPTM for tall fescue. Effective delivery of novel Epichloë-infected cultivars requires care with management of seed and quality control systems, the monitoring of Epichloë viability is required through the distribution chain, and the seed must be stored at relative low humidity and low temperature until ready to be sown.
Elymus spp. plants are the largest genus in the Poaceae family. Among these plants, Elymus cylindricus and Elymus tangutorum are excellent herbage in western China; they possess remarkable nutritional value and palatable taste, making them an important choice for animal feed. Their root systems are well-developed, enabling them to have a strong soil and water conservation ability and they have become one of the important native grasses for the restoration and reconstruction of degraded grassland on the Qinghai–Tibet Plateau [34,35]. El. tangutorum is known to harbor Epichloë endophyte [35,36], and many other Elymus species have been reported to be associated with Epichloë elymi, Epichloë bromicola, and Epichloë canadensis. The genes responsible for the biosynthesis of ergot alkaloids, indole-diterpenes, bispyrrolidines (loline), and pyrrolipyrazines (peramine) in Epichloë endophyte symbiotic with El. cylindricus and El. tangutorum from the Qinghai–Tibet Plateau have not yet been characterized. In this study, El. cylindricus and El. tangutorum were collected from four different locations in the Qinghai–Tibet Plateau, and 30 Epichloë strains were isolated. Morphology and phylogenetic methods were used to characterize the asexual Epichloë of El. cylindricus and El. tangutorum. Further, alkaloid production potential was conducted based on PCR profiling of genes required for the four kinds of alkaloid biosynthetic pathways. The results contribute to the knowledge about grass–Epichloë symbioses and lay a solid foundation for further utilization of Epichloë in germplasm innovation.
2. Materials and Methods
2.1. Collection and Preservation of Plant Materials and Endophytes
The El. tangutorm and El. cylindrical specimens were collected between August and September 2023, with mature seeds obtained from Haixi, Hainan, Guoluo, and Yushu prefectures in Qinghai Province, China (Table 1). The total number of plants sampled at each location is presented in Table 1. To ensure the independence and representativeness of the samples, only one tiller was collected per plant. Leaf sheaths of samples were microscopically examined using the aniline blue staining method described by Li (2008); healthy stem tissue was ruptured gently, stained with 0.05% aniline blue solution, and then viewed under an optical microscope [37]. Half of the seeds from endophyte-infected (E+) plants were used for Epichloë endophyte isolation. The rest of the plants were stored at 4 °C to maintain the viability of the Epichloë endophyte and seeds.

Table 1.
The seed lots of Elymus cylindricus and Elymus tangutorum used in this study.
2.2. Isolation of Endophytes and Morphological Examination
Infected seeds were surface sterilized with 70% ethanol for 3 min, placed into 5% sodium hypochlorite for another 3 min, then rinsed three consecutive times with sterile water, and left on sterile filter paper to dry. A group of 3–5 seeds was placed on each PDA plate supplemented with 100 μg/mL ampicillin and 50 μg/mL streptomycin [38]. The plates were sealed and incubated at 22 °C in the dark, with daily checks for contamination. After the mycelium grew out, it was purified three times to obtain the pure culture isolate. Mycelial disks (0.4 cm diameter) were placed in the center of the PDA medium and incubated at 22 °C in the dark for 32 days. Colony morphology was observed and photographed [39]. Growth rate experiments were conducted by placing mycelial disks on a PDA medium with six replicates per strain and incubating at 25 °C in the dark, colony diameter was measured weekly over a 56 d period using the cross-crossing method, and the daily growth rate for each strain was calculated [37,40,41]. The results presented are expressed as the mean ± standard error (Table 2). A drop of sterile water was placed in the center of the slide, a cover slide with mycelium was placed on it, and the morphology of conidia and conidiogenous cells was observed and photographed using a fluorescence microscope (Olympus BX63, Olympus, Tokyo, Japan). Images were captured with an Olympus BX51 camera supported by Cellsens Entry 1.8 software (Olympus Corp., Tokyo, Japan). Images were used to measure the length and width of conidia (30 conidia were measured for each isolate) and the length of conidiogenous cells (20 conidiogenous cells were measured for each isolate). Results are presented as mean ± standard error (Table 2). The morphologies of the isolates were compared with those of other Epichloë species, including species isolated from Elymus spp.
Table 2.
Morphological characteristics of Epichloë bromicola isolated from Elymus cylindricus, Elymus tangutorm in this study, and E. bromicola endophytes isolated from Elymus spp. as references.
2.3. DNA Extraction, PCR Analyses, and Phylogenetic Analysis
Total DNA was extracted from Epichloë endophytes. After each strain was purified and cultured on PDA medium for four weeks, mycelia on the surface of the medium were scraped using a sterile slide, and the total DNA of Epichloë endophyte was extracted using a fungal kit with the instructions of the manufacturer (Omega, Beijing, China). Following Moon et al. (2002) [52], PCR amplification was carried out with two sets of housekeeping genes: tefA, which is an elongation factor gene, and tubB, which codes for tubulin B. For tubB, the primers were tub2-exon 4u-2 and tub2-exon 1d-1 [52]. For tefA, the primers were tef1-exon 5u-1 and tef1-exon 1d-1 [53]. The 25 µL PCR amplification mixture consisted of 12.5 µL of 2 × San Taq Fast PCR Mix, 1 µL (10 µM) of each of the target-specific primer, 9.5 µL of ddH2O, and 1 µL of DNA at a concentration of 40 ng/µL. The PCR cycling conditions used were one cycle of initial denaturation at 94 °C for 5 min, then 34 cycles of denaturation at 94 °C for 30 s, annealing at 52 °C (for tubB) or 55 °C (for tefA) for 30 s, extension at 72 °C for 1 min, and finally one cycle for synthesis at 72 °C for 10 min [54,55]. The amplified products were analyzed by electrophoresis using 1.5% agarose gel in 1 × TAE buffer. DNA fragments were stained with Gold View (Solarbio Corp., Beijing, China) and viewed by UV transillumination. The products were sent to Shanghai BioEngineering Company (Shanghai, China) for sequencing. The sequences were submitted to GenBank (Accession numbers: tubB: PQ157737~PQ157766; tefA: PQ119943~PQ119947 and PQ157712~PQ157736). The sequences were compared with known Epichloë spp. using the online tool MAFFT (v. 7.526) (https://mafft.cbrc.jp/alignment/server/index.html (accessed on 13 September 2024)) [56]. Bioedit v. 7.20 software checked and optimized the comparison results [57]. IQtree (v. 2.3.6) software was used to select the most suitable tree-building model and construct the maximum likelihood (ML) phylogenetic tree [58]. Use Interactive Tree of Life (ITOL) (v. 7.2) (https://itol.embl.de/ (accessed on 15 September 2024)) to view and beautify evolutionary trees [59]. The housekeeping genes tubB and tefA were used to construct a phylogenetic tree. tefA sequence of Claviceps purpurea (GenBank accession number AF276508) and tubB sequence of C. purpurea (GenBank accession number AF062646) were used as the outgroup of the tree [50,52].
2.4. Alkaloid Gene Detection
Based on PCR amplification, the endophytes were tested for the presence of mating type and alkaloid synthesis genes using specific primers, as described previously [51,60]. About 46 genes related to Epichloë endophyte were detected. In total, 2 mating type genes, 8 segments of the ppzA gene, which is involved in peramine biosynthesis, 14 EAS cluster genes involved in ergot alkaloid biosynthesis, 11 IDT/LTM cluster genes required for indole-diterpene production, and 11 LOL cluster genes required for loline biosynthesis were identified. Using the total DNA of Epichloë endophyte, specific primers for each of the 46 genes were selected for PCR amplification. The PCR cycling parameters were set to 94 °C for 1 min, followed by 30 cycles of 94 °C for 15 s, 56 °C for 30 s, 72 °C for 1 min, and then final annealing at 72 °C for 10 min [51,60]. This cycle was used for all the primers. The PCR products were detected by 1.5% agarose gel electrophoresis and viewed by UV transillumination. E. inebrians strain E818 was used as a positive control for EAS gene cluster genes [10], and E. festucae var. lolii strain AR1 was used as a positive control for ppzA and some genes at the IDT locus [24,61]. The endophytic fungus Epichloë sp. FS001 isolated from Festuca sinensis was also used as a positive control for the ppzA and IDT gene cluster genes [62].
3. Results
3.1. Isolation of Endophytes
Table 1 shows that El. cylindricus and El. tangutorum were collected from Qinghai Province, China. There were 514 individual plants of the two grasses, 83 of which harbored endophytic fungi of the Epichloë genus, resulting in a total Epichloë infection rate of 16.15% (Table 1). For El. cylindricus, a total of 372 individual plants were collected, of which 61 were found to harbor Epichloë endophyte, resulting in a total infection rate of 16.4%. The Epichloë infection rate among plants from different regions ranged from 5.88% to 60%, with the highest rate found in the Haixi prefecture. For El. tangutorum, a total of 142 individual plants were collected from Haixi, Hainan, and Yushu prefectures, of which only 22 were found to carry Epichloë endophyte, yielding a total infection rate of 15.5%. The Epichloë infection rate for this species ranged between 4.34% and 40%, with the Haixi prefecture again exhibiting the highest rate. A total of 30 Epichloë endophytes strains were isolated from surface-sterilized Epichloë endophyte-infected samples, including 17 strains from El. cylindricus and 13 strains from El. tangutorum (Table 1).
3.2. Morphological Examination
All 30 strains exhibited typical characteristics of Epichloë endophyte in the PDA medium. Due to different environmental conditions such as sampling host, altitude, latitude, and longitude, 30 strains showed some differences in colony morphology, conidia size, conidiogenous length, growth rate, and other morphological characteristics. In culture on PDA after 30d at 25 °C, the colonies’ characteristics of the strains can be summarized into two categories: in the first type, the middle of the mycelia is white, the edge is light yellow, the center has a spherical bulge, and the colony edge is regular and cotton-like at the top; in the second type, the mycelium texture is dense, the cotton is white, and the colony edge is regular (Figure 1). The reverse of colonies is light brown centrally to cream at the margin, and the color of the isolates MB6, MB8, YZ8, YZ12, and YZ14 gradually changed from dark brown in the middle to pale yellow on both sides. The mature conidia could be seen after 40 days of cultivation on PDA medium. The conidia were boat-shaped to oval-shaped and had single terminals. For 30 isolates, the length of the conidia ranged from 3.45 to 5.47 μm, the width from 1.27 to 3.37 μm, and the length of the conidiogenous cell from 9.09 to 16.55 μm. The 30-day colonies range in diameter from 1.38 to 3.67 cm, with an average growth rate ranging from 0.63 to 1.18 mm/day (Table 2). The isolates from two grasses differ in conidia size, conidiogenous cell size, and growth rate. For El. cylindricus, the length of the conidia ranged from 3.45 to 4.64 μm, the width from 1.27 to 2.79 μm, the length of the conidiogenous cell from 9.09 to 25.48 μm, and the growth rate from 0.63 to 1.18 mm/day. For El. tangutorum, the length of the conidia ranged from 3.5 to 5.47 μm, the width from 1.66 to 3.37 μm, the length of the conidiogenous cell from 9.56 to 16.55 μm, and the growth rate from 0.78 to 1.11 mm/day. Significant differences in conidia size, conidiogenous cell size, and growth rate existed between isolates from two hosts and between isolates from the same host.

Figure 1.
Colony morphology, conidia, and conidiogenous cells of selected Epichloë endophyte isolated from Elymus tangutorum and Elymus cylindricus. (The colony is from cultures grown on PDA at 25 °C for 32 d; MB5 and MB13 were isolated from El. tangutorum, YZ5 and YZ12 were isolated from El. cylindricus. The surface view of colonies (A,D,G,J). The reverse view of colonies (B,E,H,K). The micrographs of conidiogenous cells (black arrow) and conidia (red arrow) (C,F,I,L), black scale bars = 5 μm, white scale bars = 1 cm).
The morphological characteristics of the other E. bromicola strains listed in Table 2 included E. bromicola from Bromus ramosus, B. benekenii, B. erectum, Hordeum brevisubulatum, Hordelymus europaeus, H. bogdanii, Leymus chinensis, Psathyrostachys lanuginosa, and Roegneria kamoji. When compared to the previously reported E. bromicola endophytes, the morphological features (conidia size, length of conidiogenous cells, and growth rate) of the 30 isolates examined in this study were slightly different but still fell within the normal range. However, the morphological indexes could be easily affected by culture conditions, and the Epichloë identity could not be accurately identified from morphological characteristics. In the morphology of Epichloë species in different host plants in the Elymus genus, at least four different taxa, E. bromicola, Epichloë elymi, Epichloë canadensis, and Epichloë glyceriae, have been identified from different Elymus species.
3.3. Phylogenetic Analysis
The phylogenetic tree obtained from the maximum likelihood analysis based on the tubB gene sequence showed that the 30 strains from El. cylindricus and El. tangutorum fell into a cluster with E. bromicola isolated from El. tangutorum, El. cylindricus, El. repens, and L. chinensis. All these strains constituted a branch with a 100 bootstrap support value, as shown in Figure 2. According to the maximum likelihood phylogenetic tree based on the tefA housekeeping gene sequences of Epichloë endophytes, 30 strains isolated from El. cylindricus and El. tangutorum clustered with E. bromicola isolated from El. tangutorum, which had a bootstrap support value of 84 (Figure 3). Maximum likelihood phylogenetic trees constructed with housekeeping gene sequences, tefA and tubB, were congruent. Based on these results, all 30 Epichloë endophytes from El. cylindricus and El. tangutorum were identified as E. bromicola.
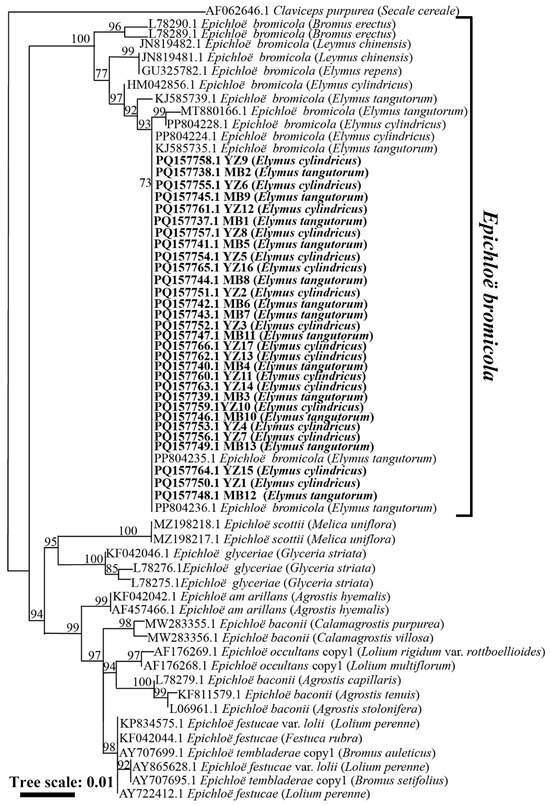
Figure 2.
Phylogeny derived from a maximum likelihood (substitution model K2P + I) analysis of the tubB gene from representative Epichloë spp. The tree was rooted with Claviceps purpurea as the outgroup.
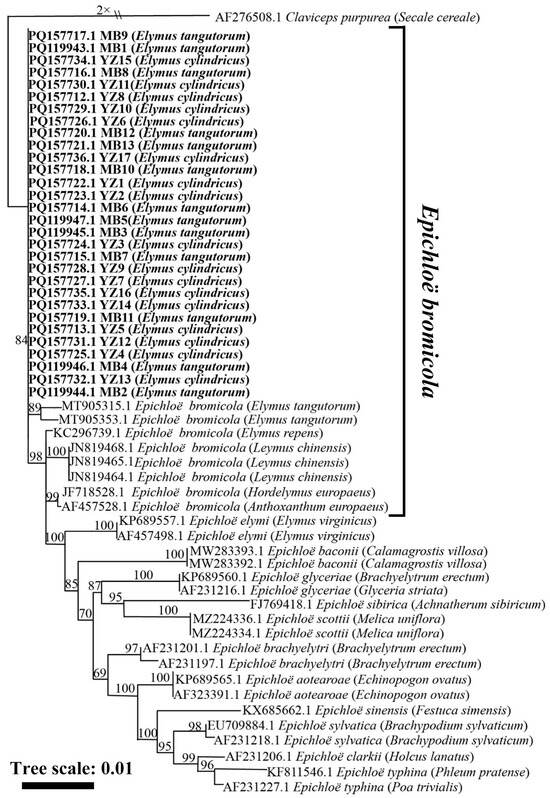
Figure 3.
Phylogeny derived from a maximum likelihood (substitution model K2P + R2) analysis of the tefA gene from representative Epichloë spp. The tree was rooted with Claviceps purpurea as the outgroup.
3.4. Alkaloid Gene Detection
During the amplification of mating type genes, 30 isolates in this study exhibited identical amplification profiles, being positive exclusively for the mtAC marker, which is indicative of mating type A (Table 3). Within the eight domain structures of the peramine synthetase-encoding gene, ppzA (formerly known as the perA gene), five isolates harbored all eight domains; in contrast, three strains were negative for all eight domains, while other strains exclusively contained one or several domains among the ppzA gene. The ppzA-∆R (representing allele ppzA-2) refers to the ppzA gene from which the R-domain has been deleted, the functional implication of this deletion is the absence of the final enzymatic step required to convert diketopiperazine into peramine in the ∆R variant, resulting in the production of pyrrolopyrazine-1,4-diones instead of peramine. Previous studies have indicated that this variant retains the capacity to encode different metabolites and may confer protective effects on their host [21]. Among the strains tested, 24 were positive for ppzA-∆R, suggesting their inability to produce peramine. In contrast, strains YZ8, MB8, MB11, YZ11, YZ13, and MB13 were negative for ppzA-∆R, but likely incapable of synthesizing peramine due to the absence of key functional domains within the ppzA gene.
Table 3.
Mating type and segments of peramine biosynthesis genes (ppzA) in the genome of Epichloë.
Among the 14 genes involved in ergot alkaloid synthesis, 25 out of 30 isolates were found to harbor the genes dmaW, easF, easC, and easE, indicating their potential for synthesizing chanolavine I (CC). Seven isolates, MB2, MB3, MB6, MB7, MB8, MB11, and YZ11, contained 8 of the 14 genes associated with ergot alkaloids’ biosynthesis, including dmaW, easF, easC, easE, easD, easA, easG, and cloA, suggesting their potential to produce chanolavine I (CC) and D-lysergic acid (D-LA). Furthermore, 11 out of 14 genes at the EAS locus were identified in 11 isolates, implying that these strains may be capable of synthesizing chanolavine I (CC), D-lysergic acid (D-LA), and ergovaline (ERV), but no ergonovine and lysergic acid, based on the current understanding of the biosynthetic pathway. However, five isolates, YZ5, YZ7, YZ8, YZ9, and YZ14, appeared to lack a functional easF gene, which has been shown to be required for the biosynthesis of early intermediates in the ergot alkaloid pathway, indicating that the five isolates may not produce even early intermediates in the ergot alkaloid pathway (Table 4). Of the 11 genes within the LTM/IDT clusters, strains YZ16, YZ17, and MB10 contained five of them, including idtG, idtB, idtM, idtC, and idtS, suggesting their potential to synthesize paspaline, but not terpendole, paxilline, paspalinine, and lolitrm, theoretically. Other strains appeared to lack some functional genes such as idtG, idtB, idtM, and idtC, which have been shown to be required for the biosynthesis of early intermediates in the lolitrem B pathway, probably rendering them incapable of synthesizing any type of indole-diterpene alkaloids (Table 5). Regarding the 11 genes within the genes responsible for loline alkaloid synthesis, all 30 strains contained only the lolC gene and theoretically had no potential to the production of 1-acetamido-pyrrolizidine (ACAP) or other LOL intermediates.
Table 4.
Ergot alkaloid biosynthesis genes (EAS) in the genome of Epichloë endophytes.
Table 5.
Indole-diterpene alkaloids biosynthesis genes (IDT/LTM) in the genome of Epichloë endophytes.
Phylogenetic analysis confirmed that all 30 isolates were E. bromicola; mating type gene detection shows that all 30 strains belonged to mating type A. Based on the results of four types of alkaloid synthesis genes, 30 isolates were divided into five types (Table 6). Type I included three strains (YZ16, YZ17, MB10) which would have the ability to produce paspaline (PAS), chanoclavinel (CC), D-lysergicacid (D-LC), and ergovaline (ERV). This type is also the most abundant type of alkaloid production among the isolates. Type II included nine strains (YZ1, YZ2, YZ3, YZ4, YZ6, YZ10, YZ15, MB1, MB9) which were predicted to possess the capability to synthesize chanoclavinel (CC), D-lysergicacid (D-LA), and ergovaline (ERV). Type III comprised seven strains (YZ11, MB2, MB3, MB6, MB7, MB8, MB11), which were predicted to possess the potential to synthesize chanoclavinel (CC) and D-lysergicacid (D-LA). Type IV included six strains (YZ12, YZ13, MB4, MB5, MB12, MB13), which had the potential to synthesize chanoclavinel (CC). Type V included five strains (YZ5, YZ7, YZ8, YZ9, YZ14), which had no theoretical potential to synthesize any kind of alkaloids.
Table 6.
The types of Epichloë endophytes isolated from Elymus cylindricus and Elymus tangutorum.
4. Discussion
It was demonstrated that E. bromicola is a common seed-transmitted endophyte of El. cylindricus and El. tangutorum, which are important rangeland forage grasses in north China, and that E. bromicola is vertically transmitted in El. cylindricus and El. tangutorum seeds but very rarely causes the chock disease. Furthermore, variation was found in the genes for biosynthesis of peramine, ergot, and indole-diterpene alkaloids, and it was shown that some isolates may have the ability to produce complex ergot alkaloids such as ergovaline, which are known to be toxic to insects and livestock [26], while 5 isolates out of 30 were found to lack genes responsible for the biosynthesis of any alkaloids.
In previous studies, it has also been found that E. bromicola endophytes can infect many other host plants, for example, H. brevisubulatum [42], L. chinensis [45,63], P. lanuginosa [63], and are further found in a large number of Elymus spp. plants, including El. dahuricus, El. excelsus, El. nutans, and El. tibeticus [16]. In a study by Du et al. (2024), all 20 E. bromicola strains associated with five Elymus spp. from five regions in northwest China were identified as mating type A (MTA) [36]. Yi et al. (2018) conducted a study analyzing E. bromicola from six Hordeum seed accessions, all of which were classified as MTA [44]. E. bromicola isolates obtained from symbiotic associations with H. brevisubulatum were also found to belong exclusively to MTA [42]. In another study, E. bromicola isolates derived from El. dahuricus revealed that all but one of the ten isolates belonged to mating type A (MTA) [49]. Among the eight E. bromicola isolates from El. kamoji, six were classified as mating type B (MTB), while the remaining two were categorized as MTA [47]. Similarly, Chen et al. (2024) found two MTA and two MTB isolates from P. lanuginosa [46]. In the present study, all 30 isolates were categorized as MTA. The above studies collectively suggested that mating type diversity in E. bromicola is relatively low. However, the simultaneous presence of both mating type A (MTA) and mating type B (MTB) in certain E. bromicola populations suggests the potential existence of a sexual stage in specific cases, despite the absence of observed stromata on El. cylindricus and El. tangutorum under natural field conditions. Surveys of Epichloë endophyte infection in El. dahuricus across its natural distribution range in Xinjiang, Shanxi, and Beijing, China, have revealed highly variable infection frequencies, ranging from 0% to 100%. E. bromicola is the predominant, and often sole, Epichloë species associated with El. dahuricus; however, its sexual stage has not been observed in this host [49]. Similarly, the sexual reproductive stage of E. bromicola strains isolated from five Elymus species collected in various regions of northwest China remains unobserved [36]. Originally identified from Bromus spp. grasses, E. bromicola was first described as a common choke disease pathogen of B. erectus and as a strictly seed-transmitted endophyte in B. benekenii and B. ramosus within the grass tribe Bromeae [36]. The formation of stromata in El. cylindricus and El. tangutorum warrants further investigation.
In this study, the morphological characteristics of Epichloë endophyte isolated from different grass species and the same grass species often differed in the PDA medium, mainly in conidia size, conidiogenous cell size, and growth rates. This has been seen in previous studies; Epichloë endophytes carried by the same species of grass and the morphological characteristics of the Epichloë endophyte of the same species vary depending on the geographical region [44]. For example, Epichloë endophytes exhibiting distinct morphological characteristics were isolated from Festuca arizonica, a dominant perennial bunchgrass found in ponderosa pine–grassland communities in the southwestern United States [64]. In addition, three isolates from surface-sterilized seeds of H. bogdanii showed different morphological characteristics in culture [65].
The salient feature of Epichloë endophytes is that they provide a direct defense to their host by producing protective alkaloids that prevent the host from being consumed by herbivorous insects or herbivores [39,66]. This study comprehensively examines the Epichloë endophytes associated with seeds of El. cylindricus and El. tangutorum. The 30 strains exhibited distinct alkaloid biosynthetic capabilities. Alkaloid biosynthesis potential differed significantly among strains from host plants and habitats [42,49]. This phenomenon is consistent with the results of this study where 30 isolates highlight intraspecific diversity within E. bromicola regarding their alkaloid-producing potential; examples of this phenomenon are seen in E. bromicola isolated from El. dahuricus [49], El. tangutorum, El. cylindricus, El. sibiricus, El. nutans [36], and P. lanuginosa [46]. Studies on the biosynthetic pathway of alkaloids based on genomics and gene silencing technology show that the presence or absence of genes in the middle or at the end of the biosynthetic pathway greatly increases the diversity of alkaloids [26,67]. The diversity of indole-diterpenoid–alkaloid profiles is related to IDT cluster genes [15], which contain many transposon-derived repeats, which may promote rearrangement and partial or complete deletion of genes [26]. In this study, 27 strains lacked the potential to synthesize lolitrem B and other intermediate metabolites due to the lack of one or more genes required to synthesize the first intermediate product. The diversity of ergot alkaloid profiles is associated with changes in the organization, gene content, and gene sequence of EAS clusters [15]. Due to the lack of lpsC, easO, and easP genes, all 30 Epichloë endophytes were probably unable to produce ergonovine (EN) and lysergic acid α-hydroxyethylamide (LAH). Five strains lacked easF, easC, and easE gene fragments and did not have the potential to synthesize any ergot alkaloids. This phenomenon has been previously observed in E. festucae. Genome sequencing of two isolated Epichloë endophytes, F11 and E2368, from E. festucae revealed that isolate F11 was capable of synthesizing the ergot alkaloid end products ergovaline and peramine, as well as the indole-diterpene end product lolitrem B. In contrast, E2368 produced only ergot alkaloid end products [26]. This phenomenon is also found in the endophyte of F. rubra subsp. commutate [24,68]. In summary, there is considerable variation in alkaloids between different Epichloë species and even within species [68,69]. This intraspecific alkaloid diversity appeared to be influenced by both genetic polymorphism within the population and host species.
The identification of ergovaline-producing endophyte isolates YZ16, YZ17, and MB10 in El. cylindricus and El. tangutorum was unexpected since the ergovaline-producing endophyte could pose risks to livestock. No study has reported that livestock were poisoned by El. cylindricus and El. tangutorum, yet. However, the effects of ergot alkaloids are dependent on multiple factors, such as livestock genetics, stress, and dosage. Further investigations are required to evaluate whether El. Cylindricus and El. tangutorum with ergovaline-producing E. bromicola could affect livestock reproduction and health in current conditions. Ergovaline-producing Epichloë coenophiala isolates in much of more than 14 million ha of Schedonorus arundinaceus have caused hundreds of millions of dollars in lost productivity annually due to fescue toxicosis in United States [70,71]. The presence of ergonovine and ergine in El. tangutorum was documented by Shi et al.; ergovaline, terpendoles, and peramine were confirmed in El. dahuricus plants with E. bromicola in China [72]. Although the risk of toxicosis may be much less in rangelands of China due to relative high diversity of forage and other grasses, the significance of ergovaline, ergonovine, and ergine production by Epichloë of important forage grasses such as El. cylindricus and El. tangutorum warrants consideration for livestock that graze them and livestock management in rangelands of China. No livestock poisoning incidents were observed in the corresponding collection area of this study. This is primarily attributed to the insufficient concentration of toxic alkaloids to exceed the threshold required for livestock toxicity, or to the low biological activity of these alkaloids, which is inadequate to induce poisoning. However, further experimental investigation is necessary to assess their alkaloid production capacity in host grasses, with potential implications for applications in artificial inoculation studies.
The utilization of Epichloë in plant breeding has been gradually adopted due to the demonstrable benefits they contribute to the host and the ease of detecting and screening the alkaloids they produce. One strategy is to inoculate Epichloë species that do not synthesize harmful alkaloids into other grass species or food grain species, thereby generating novel varieties with desirable properties [73]. However, the actual results are often suboptimal, as some materials that have been successfully inoculated with Epichloë strains may exhibit poorer performance, such as severe stunted growth [74]. Researchers have introduced E. bromicola from El. dahuricus and El. mutabilis into Triticum aestivum and Secale cereale, respectively, with variable effects on host phenotype [74]. Nevertheless, it is noteworthy that although some of the materials inoculated with Epichloë showed successful performance, there are potential intergenerational transmission barriers due to the incompatibility between certain Epichloë species and specific hosts. Many successful cases have demonstrated the practicality of the Epichloë species in grass germplasm innovation. For instance, researchers utilized the well-characterized Epichloë strains NEA2, AR1, and AR37 to develop multiple commercially valuable grass cultivars, which comprised over 70% of proprietary seed sales in New Zealand a decade ago [75]. All isolates examined in this study provide the necessary materials for utilizing Epichloë for plant germplasm innovation. However, further experiments are needed to assess their ability to produce alkaloids in hosts, with the potential for application in artificial inoculation studies.
So far, there have been no reports of natural infection of cereal crops by the Epichloë species. Given the significance of cereal crops, there is an increasing interest in exploring the Epichloë species to accelerate the breeding of new cereal crop varieties with specific traits. The prerequisites for this are that the Epichloë provide an enhanced adaptability potential for their non-natural hosts [74,76], although obtaining and optimizing this benefit in these grains will require subsequent breeding or even genetic engineering to address the variability and compatibility of the Epichloë effect. Researchers have investigated the inoculation of E. bromicola strains isolated from wild barley to cultivated barley, which show a close genetic relatedness to wild barley. The report highlights the significant achievements, such as an increase in above-ground biomass, seed production per plant, and an earlier onset of the growing period [76]. The experimental operation involving fungal cultivation and inoculation of the Epichloë species indicates that the degree of genetic similarity between the natural and new host plants is positively correlated with the possibility of establishing a mutually symbiotic relationship [74]. It is imperative to explore Epichloë species in the wild relatives of cultivated plants. While challenges remain in optimizing Epichloë species inoculation and understanding the mechanisms underlying host compatibility, successful applications in cereal crops’ breeding underscore the utility of these endophytes in generating novel germplasm with desirable attributes. The Epichloë analyzed in this study were isolated from Elymus species (tribe Triticeae), which have a relatively close genetic relationship with barley, rye, and wheat, highlighting their potential in the genetic improvement of barley, rye, wheat, and their specific traits. The finding of ergovaline, paspaline, chanoclavine I, and D-lysergic produced by some El. dahuricus and El. mutabilis isolates illustrates the need to caution that these fungal toxins might also be introduced into crops through this novel, seed-transmitted symbiotic relationship.
Although alkaloids are generally regarded as the direct mediators of host defense, new evidence suggests that the Epichloë species may also activate the indirect defense pathways of grasses. Previous studies mainly focused on alkaloid-producing Epichloë spp. that simultaneously activate both direct and indirect defense mechanisms of host grasses, thereby complicating the unraveling of their mechanisms. In the current research, it was found that five isolates had no theoretical potential to synthesize any kinds of alkaloids; should these isolates be unable to synthesize alkaloids from all four main categories, it may serve as a valuable model for exploring the mechanism of how Epichloë species activate host defense and for underling alkaloid gene loss during evolution. This system can also provide a comprehensive framework for studying the Epichloë-mediated indirect plant defense mechanisms, thereby reducing the confounding factors in metabolite analysis.
5. Conclusions
In conclusion, El. cylindricus and El. tangutorum were identified as previously unreported hosts of Epichloë endophytes in China. Morphological examination and phylogenetic analyses confirmed the identity of 30 strains as E. bromicola. The alkaloid biosynthesis gene profiles in El. cylindricus and El. tangutorum were first investigated and it was found that the isolates only have the potential to produce some early pathway intermediates, and intermediates within the indole-diterpene alkaloids and ergot alkaloids pathway, such as paspaline, chanoclavine I, D-lysergic acid, and ergovaline, while no peramine and loline would be expected. Considering the toxicity of these isolates to the safety of herbivorous livestock, the five types of strains can be divided into two categories. Category I includes five animal-safe strains of type V, which do not produce any kinds of alkaloids, provide excellent basic materials for artificial inoculation in the future, and can be fully utilized in the resistance breeding of Poaceae plants. However, further experimental studies are required to evaluate the alkaloid-producing capabilities of these endophytes in cultivated hosts, with potential implications for applications in artificial inoculation programs. Category II includes the remaining 25 strains which could produce indole-diterpene (paspaline) and/or ergot alkaloids (chanoclavine I, D-lysergic acid, ergovaline) that are toxic to herbivorous livestock. This research advances the knowledge of Epichloë endophytes by identifying the potential for specific alkaloid production and documenting two new symbiotic relationships in China. Based on successful cases, it is advisable to continue exploring the diversity of Epichloë endophytes species and their diversity of alkaloid profiles to further investigate their interactions with different pasture species and their potential to enhance host stress resistance.
Author Contributions
Conceptualization, C.L.; Resources, C.L., K.H., G.B. and T.C.; Data curation, W.L. and T.C.; Writing—original draft preparation, W.L. and T.C.; Writing—review and editing, T.C., G.B. and C.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Foundation of China (U21A20239 and 32471779); the Science and Technology Planning Project of Gansu Province, China (Natural Science Foundation) (22JR5RA458); Gansu Provincial Science and Technology Major Projects (22ZD6NA007); the Science and Technology Planning Project of Dingxi County (DX2024BZ020).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
We wish to thank the editor and anonymous reviewers for their valuable comments.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Siegel, M.R.; Schardl, C.L.; Johnson, M.C. Fungal Endophytes of Grasses 1987. Annu. Rev. Phytopathol. Previous Issues 1987, 25, 293–315. [Google Scholar] [CrossRef]
- Schardl, C.L. The Epichloë symbionts of the grass subfamily Poöideae. Ann. Mo. Bot. Gard. 2010, 97, 646–665. [Google Scholar] [CrossRef]
- Christensen, M.J.; Bennett, R.J.; Ansari, H.A.; Koga, H.; Johnson, R.D.; Bryan, G.T.; Simpson, W.R.; Koolaard, J.P.; Nickless, E.M.; Voisey, C.R. Epichloë endophytes grow by intercalary hyphal extension in elongating grass leaves. Fungal Genet. Biol. 2008, 45, 84–93. [Google Scholar] [CrossRef]
- Tanaka, A.; Takemoto, D.; Chujo, T.; Scott, B. Fungal endophytes of grasses. Curr. Opin. Plant Biol. 2012, 15, 462–468. [Google Scholar] [CrossRef]
- Leuchtmann, A.; Bacon, C.W.; Schardl, C.L.; White, J.F.; Tadych, M. Nomenclatural realignment of Neotyphodium species with genus Epichloë. Mycologia 2014, 106, 202–215. [Google Scholar] [CrossRef]
- Campbell, M.A.; Tapper, B.A.; Simpson, W.R.; Johnson, R.D.; Mace, W.; Ram, A.; Lukito, Y.; Dupont, P.Y.; Johnson, L.J.; Scott, D.B.; et al. Epichloë hybrida, sp. nov., an emerging model system for investigating fungal allopolyploidy. Mycologia 2017, 109, 715–729. [Google Scholar] [CrossRef]
- Leuchtmann, A.; Young, C.A.; Stewart, A.V.; Simpson, W.R.; Hume, D.E.; Scott, B. Epichloë novae-zelandiae, a new endophyte from the endemic New Zealand grass Poa matthewsii. New Zealand J. Bot. 2019, 57, 271–288. [Google Scholar] [CrossRef]
- Tintjer, T.; Leuchtmann, A.; Clay, K. Variation in horizontal and vertical transmission of the endophyte Epichloë elymi infecting the grass Elymus hystrix. New Phytol. 2008, 179, 236–246. [Google Scholar] [CrossRef]
- Gundel, P.E.; Rudgers, J.A.; Whitney, K.D. Vertically transmitted symbionts as mechanisms of transgenerational effects. Am. J. Bot. 2017, 104, 787–792. [Google Scholar] [CrossRef]
- Chen, L.; Li, X.; Li, C.; Swoboda, G.A.; Young, C.A.; Sugawara, K.; Leuchtmann, A.; Schardl, C.L. Two distinct Epichloë species symbiotic with Achnatherum inebrians, drunken horse grass. Mycologia 2015, 107, 863–873. [Google Scholar] [CrossRef]
- Kendall, L.; Ali, M.; Kishan, M.; Holly, P.; Marin, L. Interaction between grasses and Epichloë endophytes and its significance to biotic and abiotic stress tolerance and the rhizosphere. Microorganisms 2021, 9, 2186. [Google Scholar] [CrossRef]
- Krauss, J.; Vikuk, V.; Young, C.A.; Krischke, M.; Mueller, M.J.; Baerenfaller, K. Epichloë endophyte infection rates and alkaloid content in commercially available grass seed mixtures in europe. Microorganisms 2020, 8, 498. [Google Scholar] [CrossRef]
- Bastias, D.A.; Ueno, A.C.; Gundel, P.E. Global change factors influence plant-Epichloë associations. J. Fungi 2023, 9, 446. [Google Scholar] [CrossRef]
- Wang, J.; Hou, W.; Christensen, M.J.; Li, X.; Xia, C.; Li, C.; Nan, Z. Role of Epichloë endophytes in improving host grass resistance ability and soil properties. J. Agric. Food Chem. 2020, 68, 6944–6955. [Google Scholar] [CrossRef]
- Schardl, C.L.; Florea, S.; Pan, J.; Nagabhyru, P.; Bec, S.; Calie, P.J. The epichloae: Alkaloid diversity and roles in symbiosis with grasses. Curr. Opin. Plant Biol. 2013, 16, 480–488. [Google Scholar] [CrossRef]
- Song, H.; Nan, Z.B.; Song, Q.Y.; Xia, C.; Li, X.Z.; Yao, X.; Xu, W.B.; Kuang, Y.; Tian, P.; Zhang, Q.P. Advances in research on Epichloë endophytes in Chinese native grasses. Front. Microbiol. 2016, 7, 1399. [Google Scholar] [CrossRef]
- Bacon, C.W.; Porter, J.K.; Robbins, J.D.; Luttrell, E.S. Epichloë typhina from toxic tall fescue grasses. Appl. Environ. Microbiol. 1977, 34, 576–581. [Google Scholar] [CrossRef]
- Rowan, D.D.; Gaynor, D.L. Isolation of feeding deterrents against argentine stem weevil from ryegrass infected with the endophyte Acremonium loliae. J. Chem. Ecol. 1986, 12, 647–658. [Google Scholar] [CrossRef]
- Schardl, C.L.; Grossman, R.B.; Nagabhyru, P.; Faulkner, J.R.; Mallik, U.P. Loline alkaloids: Currencies of mutualism. Phytochemistry 2007, 68, 980–996. [Google Scholar] [CrossRef]
- Tanaka, A.; Tapper, B.A.; Popay, A.; Parker, E.J.; Scott, B. A symbiosis expressed non-ribosomal peptide synthetase from a mutualistic fungal endophyte of perennial ryegrass confers protection to the symbiotum from insect herbivory. Mol. Microbiol. 2005, 57, 1036–1050. [Google Scholar] [CrossRef]
- Berry, D.; Mace, W.; Grage, K.; Wesche, F.; Gore, S.; Schardl, C.L.; Young, C.A.; Dijkwel, P.P.; Leuchtmann, A.; Bode, H.B.; et al. Efficient nonenzymatic cyclization and domain shuffling drive pyrrolopyrazine diversity from truncated variants of a fungal NRPS. Proc. Natl. Acad. Sci. USA 2019, 116, 25614–25623. [Google Scholar] [CrossRef]
- Young, C.A.; Bryant, M.K.; Christensen, M.J.; Tapper, B.A.; Bryan, G.T.; Scott, B. Molecular cloning and genetic analysis of a symbiosis-expressed gene cluster for lolitrem biosynthesis from a mutualistic endophyte of perennial ryegrass. Mol. Genet. Genom. 2005, 274, 13–29. [Google Scholar] [CrossRef]
- Young, C.A.; Felitti, S.; Shields, K.; Spangenberg, G.; Johnson, R.D.; Bryan, G.T.; Saikia, S.; Scott, B. A complex gene cluster for indole-diterpene biosynthesis in the grass endophyte Neotyphodium lolii. Fungal Genet. Biol. 2006, 43, 679–693. [Google Scholar] [CrossRef]
- Young, C.A.; Tapper, B.A.; May, K.; Moon, C.D.; Schardl, C.L.; Scott, B. Indole-diterpene biosynthetic capability of Epichloë endophytes as predicted by ltm gene analysis. Appl. Environ. Microbiol. 2009, 75, 2200–2211. [Google Scholar] [CrossRef]
- Fleetwood, D.J.; Scott, B.; Lane, G.A.; Tanaka, A.; Johnson, R.D. A complex ergovaline gene cluster in Epichloë endophytes of grasses. Appl. Environ. Microbiol. 2007, 73, 2571–2579. [Google Scholar]
- Schardl, C.L.; Young, C.A.; Hesse, U.; Amyotte, S.G.; Andreeva, K.; Calie, P.J.; Fleetwood, D.J.; Haws, D.C.; Moore, N.; Oeser, B.; et al. Plant-symbiotic fungi as chemical engineers: Multi-genome analysis of the clavicipitaceae reveals dynamics of alkaloid loci. PLoS Genet. 2013, 9, e1003323. [Google Scholar] [CrossRef]
- Charlton, N.D.; Craven, K.D.; Afkhami, M.E.; Hall, B.A.; Ghimire, S.R.; Young, C.A. Interspecific hybridization and bioactive alkaloid variation increases diversity in endophytic Epichloë species of Bromus laevipes. FEMS Microbiol. Ecol. 2014, 90, 276–289. [Google Scholar] [CrossRef]
- Takach, J.E.; Young, C.A. Alkaloid genotype diversity of tall fescue endophytes. Crop Sci. 2014, 54, 667–678. [Google Scholar] [CrossRef]
- Easton, H.S. Grasses and Neotyphodium endophytes: Co-adaptation and adaptive breeding. Euphytica 2007, 154, 295–306. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Dash, C.K.; Akutse, K.S.; Keppanan, R.; Afolabi, O.G.; Hussain, M.; Qasim, M.; Wang, L. Prospects of endophytic fungal entomopathogens as biocontrol and plant growth promoting agents: An insight on how artificial inoculation methods affect endophytic colonization of host plants. Microbiol. Res. 2018, 217, 34–50. [Google Scholar] [CrossRef]
- Saikkonen, K.; Phillips, T.D.; Faeth, S.H.; McCulley, R.L.; Saloniemi, I.; Helander, M. Performance of endophyte infected tall fescue in Europe and north America. PLoS ONE 2016, 11, e0157382. [Google Scholar] [CrossRef]
- Cagnano, G.; Vazquez-de-Aldana, B.R.; Asp, T.; Roulund, N.; Jensen, C.S.; Soto-Barajas, M.C. Determination of loline alkaloids and mycelial biomass in endophyte-infected Schedonorus pratensis by Near-Infrared Spectroscopy and Chemometrics. Microorganisms 2020, 8, 776. [Google Scholar] [CrossRef]
- Becker, Y.; Green, K.A.; Scott, B.; Becker, A.M. Artificial inoculation of Epichloë festucae into Lolium perenne, and visualisation of endophytic and epiphyllous fungal growth. Bio-Protoc. 2018, 8, e2990. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, L.; Liu, Y.; Huang, Z.; Shi, J.; Wang, Y.; Ma, Y.; Esteban Lucas-Borja, M.; Lopez-Vicente, M.; Wu, G.L. Restoration of a hillslope grassland with an ecological grass species (Elymus tangutorum) favors rainfall interception and water infiltration and reduces soil loss on the Qinghai-Tibetan Plateau. Catena 2022, 219, 106632. [Google Scholar] [CrossRef]
- Song, H.; Nan, Z. Origin, divergence, and phylogeny of asexual Epichloë endophyte in Elymus Species from Western China. PLoS ONE 2015, 10, e0127096. [Google Scholar] [CrossRef]
- Du, M.X.; Wang, T.; Li, C.J.; Chen, T.X. Discovery and characterization of Epichloë fungal endophytes from Elymus spp. in Northwest China. Microorganisms 2024, 12, 1497. [Google Scholar] [CrossRef]
- Li, C.J.; Nan, Z.B.; Li, F. Biological and physiological characteristics of Neotyphodium gansuense symbiotic with Achnatherum inebrians. Microbiol. Res. 2008, 163, 431–440. [Google Scholar] [CrossRef]
- Latch, G.C.M.; Christensen, M.J. Artificial infection of grasses with endophytes. Ann. Appl. Biol. 1985, 107, 17–24. [Google Scholar] [CrossRef]
- Leuchtmann, A.; Schardl, C.L. Mating compatibility and phylogenetic relationships among two new species of Epichloë and other congeneric European species. Mycol. Res. 1998, 102, 1169–1182. [Google Scholar] [CrossRef]
- White, J.F. Widespread distribution of endophytes in the Poaceae. Plant Dis. 1987, 71, 340–342. [Google Scholar] [CrossRef]
- Craven, K.D.; Blankenship, J.D.; Leuchtmann, A.; Hignight, K.; Schardl, C.L. Hybrid fungal endophytes symbiotic with the grass Lolium pratense. Sydowia 2001, 53, 44–73. [Google Scholar]
- Chen, T.X.; Simpson, W.R.; Song, Q.Y.; Chen, S.H.; Li, C.J.; Ahmad, R.Z. Identification of Epichloë endophytes associated with wild barley (Hordeum brevisubulatum) and characterisation of their alkaloid biosynthesis. New Zealand J. Agric. Res. 2019, 62, 131–149. [Google Scholar] [CrossRef]
- Leuchtmann, A.; Oberhofer, M. The Epichloë endophytes associated with the woodland grass Hordelymus europaeus including four new taxa. Mycologia 2013, 105, 1315–1324. [Google Scholar] [CrossRef]
- Yi, M.; Hendricks, W.Q.; Kaste, J.; Charlton, N.D.; Nagabhyru, P.; Panaccione, D.G.; Young, C.A. Molecular identification and characterization of endophytes from uncultivated barley. Mycologia 2018, 110, 453–472. [Google Scholar] [CrossRef]
- Zhu, M.J.; Ren, A.Z.; Wen, W.; Gao, Y.B. Diversity and taxonomy of endophytes from Leymus chinensis in the Inner Mongolia steppe of China. FEMS Microbiol. Lett. 2013, 340, 135–145. [Google Scholar] [CrossRef]
- Chen, T.X.; Wang, T.; Du, M.X.; Malik, K.; Li, C.J.; Bao, G.S. Discovery of Epichloë as novel endophytes of Psathyrostachys lanuginosa in China and their alkaloid profiling. Front. Microbiol. 2024, 15, 1383923. [Google Scholar] [CrossRef]
- Li, W.; Ji, Y.L.; Yu, H.S.; Wang, Z.W. A new species of Epichloë symbiotic with Chinese grasses. Mycologia 2006, 98, 560–570. [Google Scholar] [CrossRef]
- Kang, Y.; Ji, Y.; Sun, X.; Zhan, L.; Li, W.; Yu, H.; Wang, Z. Taxonomy of Neotyphodium endophytes of Chinese native Roegneria plants. Mycologia 2009, 101, 211–219. [Google Scholar] [CrossRef]
- Shi, C.; An, S.; Yao, Z.; Young, C.A.; Panaccione, D.G.; Lee, S.T.; Schardl, C.L.; Li, C. Toxin-producing Epichloë bromicola strains symbiotic with the forage grass Elymus dahuricus in China. Mycologia 2017, 109, 847–859. [Google Scholar] [CrossRef]
- Schardl, C.L.; Leuchtmann, A. Three new species of Epichloë symbiotic with North American grasses. Mycologia 1999, 91, 95–107. [Google Scholar] [CrossRef]
- Charlton, N.D.; Craven, K.D.; Mittal, S.; Hopkins, A.A.; Young, C.A. Epichloë canadensis, a new interspecific epichloid hybrid symbiotic with Canada wildrye (Elymus canadensis). Mycologia 2012, 104, 1187–1199. [Google Scholar] [CrossRef]
- Moon, C.D.; Miles, C.O.; Järlfors, U.; Schardl, C.L. The evolutionary origins of three new Neotyphodium endophyte species from grasses indigenous to the Southern Hemisphere. Mycologia 2002, 94, 694–711. [Google Scholar] [CrossRef] [PubMed]
- Moon, C.D.; Guillaumin, J.J.; Ravel, C.; Li, C.; Craven, K.D.; Schardl, C.L. New Neotyphodium endophyte species from the grass tribes Stipeae and Meliceae. Mycologia 2007, 99, 895–905. [Google Scholar] [CrossRef]
- Gentile, A.; Rossi, M.S.; Cabral, D.; Craven, K.D.; Schardl, C.L. Origin, divergence, and phylogeny of Epichloë endophytes of native Argentine grasses. Mol. Phylogenetics Evol. 2005, 35, 196–208. [Google Scholar] [CrossRef]
- Zhang, D.X.; Nagabhyru, P.; Schardl, C.L. Regulation of a chemical defense against herbivory produced by symbiotic fungi in grass plants. Plant Physiol. 2009, 150, 1072–1082. [Google Scholar] [CrossRef]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Tippmann, H.F. Analysis for free: Comparing programs for sequence analysis. Brief. Bioinform. 2004, 5, 82–87. [Google Scholar] [CrossRef]
- Bui Quang, M.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 2461. [Google Scholar] [CrossRef]
- Zhou, T.; Xu, K.; Zhao, F.; Liu, W.; Li, L.; Hua, Z.; Zhou, X. Itol. toolkit accelerates working with iTOL (Interactive Tree of Life) by an automated generation of annotation files. Bioinformatics 2023, 39, btad339. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.; Takach, J.E.; Schardl, C.L.; Charlton, N.D.; Scott, B.; Young, C.A. Disparate independent genetic events disrupt the secondary metabolism gene perA in certain symbiotic Epichloë species. Appl. Environ. Microbiol. 2015, 81, 2797–2807. [Google Scholar] [CrossRef] [PubMed]
- Young, C.A.; Hume, D.E.; Mcculley, R.L. Forages and pastures symposium: Fungal endophytes of tall fescue and perennial ryegrass: Pasture friend or foe? J. Anim. Sci. 2013, 91, 2379–2394. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Y. Characteristics of Epichloë Endophyte–Festuca sinensis Symbiote. Master’s Thesis, Lanzhou University, Lanzhou, China, 2016. [Google Scholar]
- Wei, Y.K.; Gao, Y.B.; Xu, H.; Su, D.; Zhang, X.; Wang, Y.H.; Lin, F.; Chen, L.; Nie, L.Y.; Ren, A.Z. Occurrence of endophytes in grasses native to northern China. Grass Forage Sci. 2006, 61, 422–429. [Google Scholar] [CrossRef]
- Schulthess, F.M.; Faeth, S.H. Distribution, abundances, and associations of the endophytic fungal community of Arizona fescue (Festuca arizonica). Mycologia 1998, 90, 569–578. [Google Scholar] [CrossRef]
- Wang, T.; Chen, T.X.; White, J.F.; Li, C.J. Identification of three Epichloë endophytes from Hordeum bogdanii Wilensky in China. J. Fungi 2022, 8, 928. [Google Scholar] [CrossRef]
- Li, C.J.; Nan, Z.B.; Paul, V.H.; Dapprich, P.D.; Liu, Y. A new Neotyphodium species symbiotic with drunken horse grass (Achnatherum inebrians) in China. Mycotaxon 2004, 90, 141–147. [Google Scholar]
- Saikkonen, K.; Gundel, P.E.; Helander, M. Chemical ecology mediated by fungal endophytes in grasses. J. Chem. Ecol. 2013, 39, 962–968. [Google Scholar] [CrossRef]
- Schardl, C.L.; Young, C.A.; Faulkner, J.R.; Florea, S.; Pan, J. Chemotypic diversity of epichloae, fungal symbionts of grasses. Fungal Ecol. 2012, 5, 331–344. [Google Scholar] [CrossRef]
- Christensen, M.J. Variation in the ability of Acremonium endophytes of Lolium perenne, Festuca arundinacea and F. pratensis to form compatible associations in the 3 grasses. Mycol. Res. 1995, 99, 466–470. [Google Scholar] [CrossRef]
- Hopkins, A.A.; Young, C.A.; Panaccione, D.G.; Simpson, W.R.; Mittal, S.; Bouton, J.H. Agronomic performance and lamb health among several tall fescue novel endophyte combinations in the South-Central USA. Crop Sci. 2010, 50, 1552–1561. [Google Scholar] [CrossRef]
- Klotz, J.L. Activities and effects of ergot alkaloids on livestock physiology and production. Toxins 2015, 7, 2801–2821. [Google Scholar] [CrossRef]
- Shi, Q.; Matthew, C.; Liu, W.H.; Nan, Z.B. Alkaloid contents in Epichloë endophyte-infected Elymus tangutorum sampled along an elevation gradient on the Qinghai-Tibetan Plateau. Agronomy 2020, 10, 1812. [Google Scholar] [CrossRef]
- White, J.F.; Kingsley, K.L.; Zhang, Q.W.; Verma, R.; Obi, N.; Dvinskikh, S.; Elmore, M.T.; Verma, S.K.; Gond, S.K.; Kowalski, K. Review: Endophytic microbes and their potential applications in crop management. Pest Manag. Sci. 2019, 75, 2543–2548. [Google Scholar] [CrossRef]
- Simpson, W.R.; Faville, M.J.; Moraga, R.A.; Williams, W.M.; McManus, M.T.; Johnson, R.D. Epichloë fungal endophytes and the formation of synthetic symbioses in Hordeeae (=Triticeae) grasses. J. Syst. Evol. 2014, 52, 794–806. [Google Scholar] [CrossRef]
- Caradus, J.R.; Card, S.D.; Hewitt, K.G.; Hume, D.E.; Johnson, L.J. Asexual Epichloë fungi—Obligate mutualists. Encyclopedia 2021, 1, 1084–1100. [Google Scholar] [CrossRef]
- Li, C.J.; Wang, Z.F.; Chen, T.X.; and Nan, Z.B. Creation of novel barley germplasm using an Epichloë endophyte. Chin. Sci. Bull. 2021, 66, 2608–2617. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).