Abstract
This study aims to identify and evaluate the phosphate-solubilizing ability of endophytic bacteria isolated from roots of Chinese Cymbidium and to assess their impact on phosphorus uptake and plant growth. Thirty strains of endophytic bacteria were isolated from six orchid varieties. Molecular identification based on 16S rRNA gene sequencing revealed that the most frequently isolated strains belonged to the genera Pseudomonas and Burkholderia. Among them, 10 bacterial strains exhibited the capacity to solubilize inorganic and organic phosphorus. Two strains, designated X1 (Paraburkholderia sp. Beta-32) and X13 (Rhizobium freirei PRF81 (X13), were identified as the most effective phosphate-solubilizing bacteria (PSB). Gluconic acid was the dominant organic acid secreted, driving inorganic phosphorus solubilization, while alkaline phosphatase activities facilitated organic phosphorus mineralization. Inoculation with phosphate-solubilizing bacteria (PSB) resulted in increased plant growth and phosphorus content in both leaves and roots as compared to the control plants. PSB treatments also increased available phosphorus content in soil, reduced total phosphorus content, and increased exopolysaccharide and alkaline phosphatase activities. Real-time q-PCR analysis showed that PSB inoculation significantly upregulated the expression of phosphorus transport-related genes, including PDR2, PHF1, PHR1, PHT1;9, and PHT4;4, thereby enhancing phosphorus absorption. Moreover, strains X1 and X13 not only exhibited strong phosphate-solubilizing capacity but also demonstrated stable colonization in both roots and root rhizosphere soil of orchids over extended periods. In conclusion, the endophytic PSB identified with phosphate-solubilizing abilities increased phosphorus availability and its uptake in Chinese Cymbidium, thereby promoting plant growth and development. This is the first attempt to characterize endophytic PSB from roots of Chinese Cymbidium orchids. These findings provide a basis for selection of PSB that are efficient in P uptake for application in microbial fertilizers for orchid cultivation.
1. Introduction
Chinese Cymbidium (Cymbidium spp.), also known as Chinese orchid, has an extensive cultivation history in China. It ranks among the top commercial flowers in the country due to its distinctive aroma and cultural significance [1]. Given the multifaceted importance of Chinese Cymbidium, enhancing its growth, flower yield, and quality—including the number, size, and longevity of blooms—could substantially benefit growers financially. Floriculture production often relies heavily on synthetic fertilizers containing nitrogen (N), phosphorus (P), and potassium (K). Among these nutrients, phosphorus plays a crucial role as a precursor of sugar phosphates during respiration and photosynthesis and as a precursor of phospholipids that make up plant membranes [2,3,4,5,6]. Phosphorus is also vital for cell division, development of meristematic tissues, and stimulation of bud and bulb formation [7]. The primary goal of phosphorus fertilization in ornamental plants is to enhance growth and cultivate healthy, robust development. However, the high cost of phosphorus fertilizers limits their widespread use [8]. Moreover, prolonged application of these fertilizers can degrade soil structure and ultimately have a negative impact on flower growth and yield [8,9]. Consequently, there is a need to identify cost-effective and environmentally friendly alternatives to the use of chemicals in floriculture practices. This has prompted us to focus on investigating the use of phosphate-solubilizing bacteria (PSB) to promote growth of Chinese Cymbidium.
PSB has the potential to convert insoluble phosphates available to plants through solubilization and mineralization, increasing crop yields, while maintaining environmental sustainability [10]. Genera such as Pseudomonas, Burkholderia, Bacillus, Rhizobium, Sphingobacterium, and Stenotrophomonas have been shown to exhibit phosphate-solubilizing ability along with plant growth-promoting traits, rendering them promising candidates for biofertilizer development [8,10,11,12]. PSB converts both insoluble inorganic and organic phosphorus into soluble active phosphorus, a process associated with the secretion of various organic acids, exopolysaccharide, and extracellular enzymes such as phosphatases [11,12,13,14,15,16,17,18,19]. The secretion of various organic acids by PSB is considered the primary mechanism of phosphorous solubilization, with gluconic acid being the main component during this process [11]. PSB affects the expression of genes related to phosphorus starvation responses and high-affinity phosphorus transporters in plants. Several genes, including PDR, PHF, PHO, PHT, and SPXs, are involved in the absorption, transport, and redistribution of inorganic phosphorus, improving host nutrient absorption and promoting growth and development [20,21,22].
The use of PSB has been shown to increase plant phosphorus uptake and yield in numerous studies across a variety of crops, including rice, wheat, and maize [23,24,25,26]. The majority of these PSB originate from the soil environment surrounding crop roots, known as the rhizosphere. Endophytic bacteria share a more intimate relationship with plants than soil bacteria. As endophytic PSB are isolated from root tissues, they may possess a superior ability to colonize the host plant and potentially facilitate the mobilization of internal phosphorus pools in addition to solubilizing phosphorus in the soil. Orchidaceous plants, including the Chinese Cymbidium, are a special group of plants that have a symbiotic relationship with endophytes [27,28]. Because the seeds of orchids are small, the embryos are not fully differentiated, have little endosperm, and therefore rely on endophytic microorganisms to provide nutrients to promote germination and subsequent protocorm growth [27,28,29,30].
The unique habitat of endophytic PSB in Chinese Cymbidium is an important but underdeveloped phosphate-soluble microbial resource. The endophytic bacteria with phosphate-solubilizing functions in Chinese Cymbidium have not been studied systematically. The aim of this research work is to screen for endophytic bacteria that can dissolve inorganic phosphorus and mineralize organic phosphorous from roots of Chinese Cymbidium. These findings provide a comparison of efficient PSB in P uptake as microbial fertilizers which can be used in production of Chinese Cymbidium.
2. Materials and Methods
2.1. Sample Collection and Treatment
Roots from six orchid plantlets of different hybrids were collected from the orchid germplasm resources of the Environmental Horticulture Research Institute, Guangdong Academy of Agricultural Sciences (23°8′ N113°20′ E) (Table S1), in February 2024. Three root samples were obtained from each plantlet in sterile plastic bags, washed with sterilized water, and dried at room temperature. Each sample was surface-sterilized with 75% ethanol for 2 min, with 10% (v/v) sodium hypochlorite solution for 10 min, and then was washed with sterilized water three times [31]. The washed root tissues were gently placed on nutrient agar (NA) (containing (L−1): peptone, 10 g; beef extract, 3 g; NaCl, 5 g; 20 g of agar; 1000 mL of distilled water (pH 7.0)) solid medium and incubated at 30 °C to test for effectiveness of surface sterilization. If no colonies grew on the surface of the NA solid medium within 3 d, it indicated that the surface sterilization was successful. Simultaneously, the roots were homogenized in a sterile grinder and suspended in 1 mL of sterile water, which served as the suspension medium, followed by thorough vortexing to form a uniform suspension. Serial dilutions were then prepared to obtain 10−1, 10−2, 10−3, 10−4, and 10−5 gradient dilutions. A 50 μL aliquot from each dilution was aseptically spread onto NA solid medium and incubated at 30 °C for 3 d. Colonies grown on the NA solid medium were purified and stored at 4 °C.
2.2. DNA Extraction and PCR Amplification
The extraction of endophytic bacterial DNA and sequencing were performed according to a previously described method [32]. The 16S ribosomal RNA gene of each isolate was amplified using the universal primers 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-CTACGGCTACCTTGTTACGA-3′) [33] under the following PCR conditions: 94 °C for 5 min, followed by 35 cycles of 94 °C for 30 s, 50 °C for 30 s, 72 °C for 1 min, and a final elongation step at 72 °C for 10 min. Initial isolation yielded over 70 bacterial strains from Chinese Cymbidium roots. All isolates were characterized by amplifying 16S rDNA using universal bacterial primers. PCR products of expected size were electrophoresed, excised, purified, and sequenced. The resulting sequences were analyzed using BLAST (http://www.ncbi.nlm.nih.gov/, accessed on 23 May 2024), against the NCBI database [34]. Redundant strains were eliminated based on sequence comparisons, yielding 30 phylogenetically distinct endophytes. These selected strains were preserved in 50% glycerol at −80 °C and are listed in Supplementary Table S1. A phylogenetic tree was constructed using the maximum likelihood method with the MEGA 7.0 software [35].
2.3. Selection of PSB Strains in Solid and Liquid Media
To measure the phosphate-solubilizing ability of 30 strains of PSB, they were inoculated on inorganic phosphorus (IP) and organic phosphorus (OP) medium. IP media contained (L−1), which was composed of the following: C6H12O6, 10.0 g; (NH4)2SO4 0.5 g; FeSO4·7H2O, 0.03 g; yeast extract, 0.5 g; NaCl, 0.3 g; KCl, 0.3 g; MnSO4·4H2O, 0.03 g; MgSO4·7H2O, 0.3 g; Ca3(PO4)2, 5.0 g; 20 g of agar; 1000 mL of distilled water (pH 7.0). OP was similar to OP medium except that 2 g lecithin and 5 g CaCO3 were added instead of calcium phosphate. The strains that were able to form halo zones on both IP and OP solid medium were considered positive strains. The accession numbers of these positive strains in the NCBI database are listed in Supplementary Table S2. Initially, positive strains were grown on IP and OP solid medium plates and incubated at 30 °C for 5 d. Measurement of the phosphate solubilization halo zone diameter (D) and colony diameter (d), followed by calculation of the ratio between halo zone diameter and colony diameter (D/d) was carried out [36]. Subsequently, the phosphate-solubilizing ability of the strains was evaluated using a liquid media method [37]. First, each bacterial strain was cultured to a standardized concentration of OD600 = 0.8–1.0; then, 200 uL of each standardized culture was inoculated into 100 mL of IP or OP liquid medium and incubated at 30 °C for 5 d on a rotary shaker at 180 rpm. Uninoculated media (with 200 uL of sterile water added) served as negative controls (initial pH, 7.0; final pH, 7.0). Experiments were repeated three times for each strain. The IP and OP liquid medium were centrifuged at 10,000 rpm for 10 min. The IP medium was analyzed for pH, soluble phosphorus, organic acids, and exopolysaccharides, while the OP medium was analyzed for soluble phosphorus and phosphatase activities.
2.4. Measurement of Plant Growth Parameters
To study the effects of selected endophytic PSB on the growth and development of orchids, plant experiments were carried out in the greenhouse. After washing and centrifugation steps to remove the culture medium, the bacterial pellet was resuspended in sterile water to an OD600 of 0.8–1.0. This bacterial suspension was then diluted 10-fold with sterile water to achieve the working concentration. In the subsequent pot experiments, we employed a completely randomized design to evaluate ten different PSB strains, with sterile water serving as the negative control treatment. The tissue culture plantlets were dipped and incubated in endophytic PSB for 12 h and then planted in the pot. Each pot contained 250 g of pine bark (pH 6.5) and was planted with one orchid plantlet. There were 15 pots for each treatment, and 5 mL of the bacterial suspension (working concentration) was added to the pine bark. To meet the nutritional requirements of the plantlets, they were irrigated with modified 1/2 phosphorus-deficient Hoagland’s nutrient solution. Additionally, 2.5 g·L−1 tricalcium phosphate and 1 g·L−1 lecithin were supplemented per liter of phosphorus-deficient Hoagland’s nutrient solution (KNO3, 607 g·L−1; MgSO4, 241 mg·L−1; NH4NO3, 40 mg·L−1; FeNaEDTA, 36.7 m·L−1; KI, 0.83 mg·L−1; H3BO3, 6.2 mg·L−1; MnSO4·H2O, 22.3 mg·L−1; ZnSO4·7H2O, 8.66 mg·L−1; Na2MOO4·10H2O, 0.25 mg·L−1; CuSO4·7H2O, 0.025 mg·L−1; CoCl2, 0.025 mg·L−1; Ca(NO3)2, 945 mg·L−1; 20 g of agar; 1000 mL of distilled water (pH 6.0) as a phosphorus source substitute for potassium dihydrogen phosphate). Watering (250 mL) was carried out once every two weeks. At 30 d of co-cultivation, the orchid plantlets in the treatment groups were harvested to measure leaf and root lengths. Leaf length was measured from the lamina base to the apex in fully expanded leaves, and root length was calculated as primary root length from the hypocotyl junction to the distal tip. Subsequently, part of the leaves and roots were washed several times with deionized water, and the dry weight was recorded after drying at 80 °C for 7 d. Rhizosphere soils were sampled at 30 d after treatment for measurements of total phosphorus, available phosphorus, and pH. Roots were sampled at 30 d after treatment for measurements of phosphorus content, exopolysaccharides, and phosphatase activities.
2.5. Measurements of Soluble Phosphorus in Liquid Medium and Total Available Phosphorus in Rhizosphere Soils
The analysis of soluble phosphorus concentrations in IP and OP liquid medium was performed using the previous method [37]. After centrifugation of the bacterial strain culture, 1 mL of supernatant was transferred to a test tube. Then, 20 μL of a 10 mM 2,4-dinitrophenol indicator (Product No. D835613, Macklin Biochemical, Shanghai, China; CAS 51-28-5) was added. Subsequently, 5 mL of molybdate/antimony reagent was accurately added, the mixture was shaken well, and distilled water was added to bring the volume up to 7 mL. Following this, the test tubes were incubated for 30 min at room temperature. The appearance and intensity of the blue color indicated the total concentration of phosphorus, and the absorbance was measured at 700 nm. The phosphorous concentrations in the test plants were determined using the molybdenum blue method [38]. Total phosphorus (TP) and available phosphorus (AP) in rhizosphere soil samples were determined by sulfuric acid–perchloric acid decoction and NaHCO3 extraction was carried out using the molybdenum antimony colorimetric method [39]. The pH variations were measured using a pH meter (Mettler-Toledo GmbH, Greifensee, Switzerland).
2.6. Measurements of Exopolysaccharides
Analyses of exopolysaccharides in IP liquid medium and rhizospheric soils were performed using Gauri’s method [40]. Exopolysaccharides (EPSs) were precipitated in the supernatant by adding two volumes of isopropanol and centrifuged at 10,000 rpm for 10 min at 4 °C. After removing the supernatant, the sediment was dissolved in 2 mL of distilled water, and 1 mL of 6% phenol solution was added, followed by vigorous shaking and rapid addition of 5 mL of concentrated sulfuric acid. The mixtures were immediately shaken again and left to stand at room temperature for 30 min, and the absorbance was measured at 490 nm.
2.7. Analysis of Organic Acid Production in Liquid Cultures by HPLC
The production of organic acids in bacterial cultures carried out in IP liquid medium was performed with the methodology used by Fiori and his co-workers [41]. After centrifugation of the bacterial strain culture, 1 mL of supernatant was transferred to a test tube. Then, 500 μL of 30% methanol aqueous solution containing 0.1% formic acid was added; the mixture was shaken, mixed vigorously for 60 s, and then centrifuged at 12,000 rpm at 4 °C for 10 min. The sample was filtered through 0.22 μm membranes, and the filtrate was added to the LC-MS bottle. An amount of 5 uL was injected into a high-performance liquid chromatography (HPLC) system (Agilent 1100 series; Agilent Technologies, Santa Clara, CA, USA) equipped with an ACQUITY UPLC® BEH C18 column (2.1 × 100 mm, 1.7 μm, Waters Corporation, Milford, MA, USA). The solution of 0.1% formic acid in water (A) and methanol (B) was used as the mobile phase at 0.4 mL·min−1 flow rate. The organic acid secretion of bacterial strains was identified through a comparison of the retention times, and the quantification was performed by the comparison of the peak areas using external standards.
2.8. Measurements of Alkaline Phosphatase Activities
Alkaline phosphatase activities in both OP liquid medium and rhizosphere soils were analyzed according to Qvirist et al. [42]. After 5 d incubation in OP liquid medium, 10 mL of bacterial culture was transferred to a 15 mL conical centrifuge tube. For soil samples, 2 g was homogenized with 10 mL of sterile distilled water in a 15 mL tube and incubated statically for 1 h. Both sample types were then centrifuged at 10,000 rpm for 10 min. Supernatant of extract (0.1 mL) was mixed in 2 mL of modified universal buffer (pH = 11) and 0.5 mL of p-nitrophenyl phosphate (p-NP) substrate solution (50 mM). The change in the alkaline phosphatase solution due to p-nitrophenol (p-NP) production was measured at 400 nm, and the amount of p-NP was calculated from a p-NP calibration curve. Solution without soil served as control. One unit of phosphomonoesterase activities was defined as the amount of enzyme required to liberate 1 mM of p-NP (product) from 1 kg of dried soil/h at 37 °C. Protein concentration was determined using Coomassie Brilliant Blue G-250 solution as previously described [43].
2.9. Colonization Dynamics of PSB in Orchid Roots and Rhizosphere Soil
The rifampicin-resistant marker method [44] was employed to screen for the marker strains grown on NA solid medium containing 300 μg·mL−1 rifampicin, with colony morphology consistent with that of the original strains. Following the methods of previous studies for strain isolation and recovery, the target strains were isolated from groups of inoculated and non-inoculated orchids after 30 d. A sample of 1 g each of orchid roots and rhizosphere soil was weighed and added to 9 mL of sterile water for dilution. An amount of 100 μL of the soil was diluted at 10−2, 10−3, and 10−4 and evenly spread on NA agar plates containing 300 μg·mL−1 rifampicin. After incubation at 30 °C for 3 d, the number of colonies on each plate was counted and converted into number of colony-forming units per gram of plant roots and dry soil (cfu·g−1). Each treatment was replicated three times.
2.10. Quantitative RT-PCR Analysis and Statistical Analysis
Total RNA was extracted using RNAprep pure Plant Kit (Tiangen Inc., Beijing, China). cDNA was synthesized using HiScript III RT SuperMix for qPCR Kit (Vazyme, Nanjing, China) according to the manufacturer’s protocol. Quantitative RT-PCR was conducted on the Thermal Cycler DiceTM Real-Time System (Takara, Otsu, Japan) using the diluted cDNA as templates. The primers were designed using Primer5 software. The β-actin (Mol013347) was used as an internal reference gene to normalize the amount of template in Cymbidium sinense as described, and Actin was also used as control [45]. All primers and their sequences are listed in Supplemental Table S2. Relative expression was calculated based on the 2−ΔΔCt method [46]. Each experiment contained two biological replicates and three technical replicates.
2.11. Statistical Analysis
The physiological measurements were repeated three times from different samples. A full random design model on the SPSS software (v28.0.1.1; SPSS Inc., Chicago, IL, USA) was used to analyze the variance of data. Differences among means of samples were evaluated using Duncan’s test at p < 0.05 level.
3. Results
3.1. Bacterial Identification
Thirty strains of endophytic bacteria were isolated from roots of six Chinese orchids. The nucleotide sequences of the 16S rRNA genes of the 30 strains were amplified by PCR and sequenced. These sequences were compared by BLAST with sequences in the NCBI database, and the results are shown in Figure S1. These results showed that the 30 strains of bacteria belonged to six genera, including 11 strains of Pseudomonas, 8 strains of Burkholderia, 7 strains of Bacillus, 1 strain of Sphingobacterium, 1 strain of Rhizobium, and 1 strain of Stenotrophomonas maltophilia. Two additional isolates were only broadly classified as Bacterium sp. (Figure S1).
The IP and OP were chosen as selective culture mediums to screen for phosphate-solubilizing ability of 30 strains of endophytic bacteria. Out of these, 17 strains of endophytic bacteria could form clear phosphate-solubilizing halos on IP solid medium, and 10 strains of endophytic bacteria could form clear phosphate-solubilizing halos on OP solid medium. Therefore, 10 strains of endophytic bacteria were identified as being capable of decomposing both inorganic phosphorus and mineralizing organic phosphorus. These strains included Paraburkholderia sp. strain Beta-32 (X1), Pseudomonas sp. strain SWUSTb-72 (X2), Burkholderia ambifaria strain AU0212 (X9), Rhizobium freirei PRF81 (X13), Burkholderia fungorum (X16), Pseudomonas brenneri strain EH-G3 (X20), Paraburkholderia phytofirmans PsJN (X21), Bacillus thuringiensis strain EI-17 (X27), Bacillus wiedmannii strain ASS-1 (X29), and Burkholderia tropica strain CACua-41 (X30) (Table S1).
3.2. Evaluation of Phosphate-Solubilizing Ability
3.2.1. Ability to Dissolve Inorganic Phosphorus
Ten isolates representing endophytic PSB were analyzed over a period of 5 d in terms of their phosphate-solubilized halo zone diameters, colony diameters, D/d, pH, exopolysaccharide, and soluble phosphorus using Ca3(PO4)2 as the insoluble phosphorous sourced (IP solid medium) (Figure 1). Phosphate-solubilized halo diameters ranging from 13.68 to 25.2 mm and colony diameters ranging from 5.1 to 8.7 mm were observed 5 d after inoculation, which resulted in D/d values of 1.9 to 3.48. The differences in phosphate-solubilizing halos and colony diameters among the 10 endophytic PSB were significant. Strains X1, X13, X9, X21, and X30 had a higher D/d than X27, X20, and X29, and all PSB exhibited more than 1.5 in D/d (Figure 1a–c). There were significant differences in pH, exopolysaccharide, and soluble phosphorus in the 10 endophytic PSB. Exopolysaccharide and soluble phosphorus were increased after inoculation. Higher levels of increase were observed in X1, X13, and X9 than in X2, X6, X27, X20, and X29 (Figure 1e,f). The pH decreased after inoculation. Lower levels were observed in X1, X13, and X9 than in X30, X2, and X6, which in turn were lower than those in X27, X20, and X29 (Figure 1d).
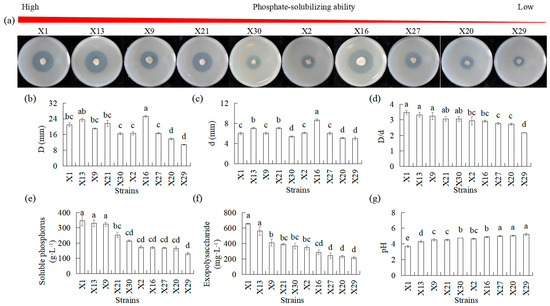
Figure 1.
Comparison of inorganic phosphate-solubilizing capacities of different endophytic PSB in Chinese Cymbidium. (a) PSB phosphate-solubilizing ability was visualized on IP solid medium through halo zone formation, followed by measurements of (b) halo zone diameter D and (c) colony diameter d. (d) The ratio between halo zone diameter and colony diameter D/d was calculated. For IP liquid medium, (e) measurements of soluble phosphorus, (f) exopolysaccharide, and (g) pH. Means of three replicates and standard errors are presented; the same letter above the column indicates no significant difference at p < 0.05.
Since the pH values of all strains decreased (Figure 1), we then measured the content of organic acids. Several organic acids were identified in the culture supernatants of endophytic PSB, most of which were glucuronic acid, lactic acid, citric acid, tartaric acid, and pantothenic acid in descending amounts (Table 1). The types of organic acids secreted by different bacteria were different, and the ability to secrete the same acid varies, but the total organic acid secretion capacities were X1 > X13 > X9 > X21 > X30 > X2 > X16 > X27 > X20 > X29. Higher levels of fumaric acid, succinic acid, pyroglutamic acid, glutaric acid, malic acid, 5-hydroxymethyl-2-furoic acid, tartaric acid, pyridoxine, citric acid, and glucuronic acid were observed in X1 as compared to X27, X20, and X29. Higher levels of lactic acid, fumaric acid, succinic acid, malic acid, phenylpyruvic acid, phenyllactic acid, glucuronic acid, and pantothenic acid were observed in X13 compared to X27, X20, and X29. Among them, fumaric acid, succinic acid, malic acid, and glucuronic acid were higher in X1 and X13 than in X27, X20, and X29.

Table 1.
Comparison of organic acid production of different endophytic PSB isolated from Chinese Cymbidium (ug·mL−1).
3.2.2. Ability to Mineralize Organic Phosphorus
Ten isolates representing endophytic PSB were studied over a period of 5 d in terms of their phosphate-solubilized halo zone diameters, colony diameters, D/d, alkaline phosphatase activities, and soluble phosphorus concentration. An amount of 2 g of lecithin and 5 g of CaCO3 was added instead of calcium phosphate (OP solid medium) as the organic phosphorous sourced (Figure 2). Phosphate-solubilized halo diameters ranging from 17.6 to 28.9 mm and colony diameters ranging from 7.3 to 11.0 mm were observed after inoculation, which resulted in D/d values of 2.0 to 2.82. The differences in phosphate-solubilizing halos and colony diameters among the 10 strains were significant. Strains X1, X13, and X9 had a higher D/d than X27, X20, and X29, and all PSB exhibited more than 2.0 in D/d (Figure 2b–d).
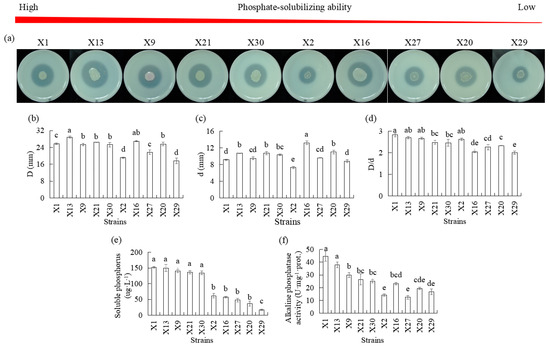
Figure 2.
Comparison of organic phosphate-solubilizing capacities of different endophytic PSB in Chinese Cymbidium. (a) PSB phosphate-solubilizing ability was visualized on OP solid medium through halo zone formation, followed by measurements of (b) halo zone diameter D and (c) colony diameter d. (d) The ratio between halo zone diameter and colony diameter D/d was calculated. For OP liquid medium, measurements of (e) soluble phosphorus and (f) alkaline phosphatase activities. Means of three replicates and standard errors are presented; the same letter above the column indicates no significant difference at p < 0.05.
We determined the content of alkaline phosphatase activities, and higher levels were observed in X1, X13, and X9 than in X2, X6, X27, X20, and X29, and the alkaline phosphatase activities secreted by X2 and X27 were the lowest (Figure 2f). There were significant differences in soluble phosphorous concentration levels among the 10 endophytic PSB strains. The soluble phosphorous concentration increased after inoculation, with higher levels observed in the X1, X13, and X9 strains as compared to the X2, X6, X27, X20, and X29 strains (Figure 2e).
3.3. Effects of Endophytic PSB Inoculation on P Uptake and Orchid Growth
Plant growth was measured to evaluate the phosphate-solubilizing ability of PSB after plantlets were inoculated with PSB for 30 d. Plants inoculated with endophytic PSB showed significantly higher growth in leaf length, root length, leaf dry weight, and root dry weight compared to the uninoculated endophytic PSB (CK) (Figure 3a). Plants inoculated with the X1, X13, and X9 strains had higher growth than plants inoculated with the X20 and X29 strains (Figure 3b,c,e,f). The increase in phosphorus content was observed in the treatment of endophytic PSB, which resulted in an increase in phosphorus content in leaf and root compared to plants not inoculated with endophytic PSB (CK). Plants inoculated with the X1, X13, and X9 strains had higher phosphorus content in leaves and in roots compared to plants inoculated with the X20 and X29 strains (Figure 3d,g).
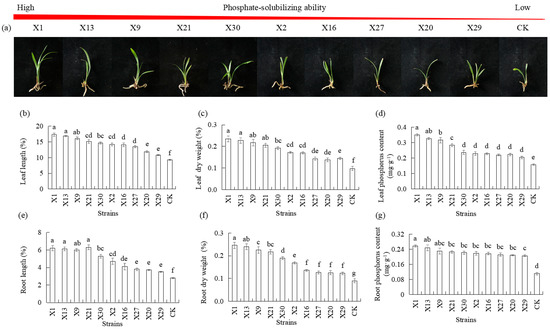
Figure 3.
Growth response of Chinese Cymbidium seedlings inoculated with different endophytic PSB. (a) Morphological characteristics photographs of Chinese Cymbidium seedlings after 30 d of PSB inoculation, followed by measurements of (b) leaf length, (c) leaf dry weight, (d) leaf phosphorus content, (e) root length, (f) root dry weight, and (g) root phosphorus content. CK represents the non-inoculated treatment (control group). Means of three replicates and standard errors are presented; the same letter above the column indicates no significant difference at p < 0.05.
3.4. Effects of Endophytic PSB Inoculation on the Properties and Phosphorus Availability in the Rhizosphere Soil of Orchids
The available phosphorus content, total phosphorus content, pH, exopolysaccharide, and alkaline phosphatase activities in rhizosphere soils were measured to evaluate phosphate-solubilizing ability in plants inoculated and not inoculated with PSB (Figure 4). Plants’ inoculation with PSB resulted in increased available phosphorus content, as well as in decreased total phosphorus content in rhizosphere soils, compared to plants not inoculated with PSB (CK). PSB treatments of orchids produced an increase of 381–619% in available phosphorus content, and total P content in rhizosphere soils decreased 11.8–25.8% compared to uninoculated plants (CK). The X1 and X13 strains demonstrated the maximum increase in available phosphorus content in rhizosphere soils, and the X1 and X13 strains demonstrated the maximum decrease in total phosphorus content in soil (Figure 4d,e). Plants’ inoculation with PSB resulted in decreased pH in soil compared to plants not inoculated with PSB (CK). The maximum decrease in pH in soil was recorded in plants inoculated with the X1 and X13 strains compared to the X27, X20, and X29 strains (Figure 4a). Inoculation with PSB significantly affected the exopolysaccharide and alkaline phosphatase activities of orchids compared to uninoculated orchids (CK). Higher levels were observed in the X1, X13, and X9 strains compared to the X2, X6, X27, X20, and X29 strains (Figure 4b,c).
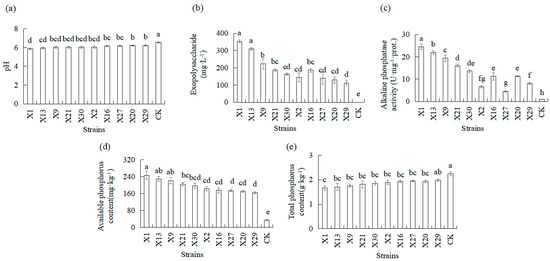
Figure 4.
Physical and chemical properties response of Chinese Cymbidium rhizosphere soil inoculated with different endophytic PSB. Rhizosphere soil properties were measured in terms (a) pH, (b) exopolysaccharide, (c) alkaline phosphatase activities, (d) available phosphorus, and (e) total phosphorus. CK represents the non-inoculated treatment (control group). Means of three replicates and standard errors are presented; the same letter above the column indicates no significant difference at p < 0.05.
3.5. Gene Transcript Levels Involved in Phosphorus Transport in Chinese Cymbidium Inoculated with Different PSB Strains
qPCR analysis showed that Chinese Cymbidium plants inoculated with endophytic PSB demonstrated significantly upregulated relative expression of PDR2, PHF1, PHR1, PHT1;9, and PHT4;4 in their roots. PDR2, PHF1, PHR1, PHT1;9, and PHT4;4 transcript levels were induced after the plants were inoculated with PSB. Higher transcript levels of PDR2, PHF1, PHR1, PHT1;9, and PHT4;4 were observed in plants inoculated with PSB compared to uninoculated plants (CK) (Figure 5a–e). SPX1, SPX3, and SPX4 transcripts were downregulated in all plants inoculated with PSB. Higher transcript levels of SPX1, SPX3, and SPX4 were observed in uninoculated plants compared to plants inoculated with PSB (Figure 5f–h). No significant difference in PHO1, PHOS32, and THO1 transcript levels was observed between plants that were uninoculated (CK) and those that were inoculated with PSB (Figure S2a,b,d). No significant difference between the PSB strains was observed. PS3 transcript levels in PSB showed no significant difference between X9, X21, X2, X16, X27, and X20 strains, on the one hand, and uninoculated PSB (CK), while higher levels were shown in X1, X13, X30, and 29 strains (Figure S2c).
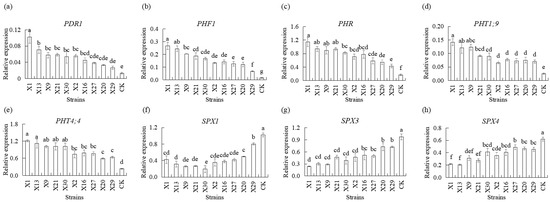
Figure 5.
Transcript levels of the genes involved in phosphorus transport of different endophytic PSB in Chinese Cymbidium. Relative expressions of (a) PDR1, (b) PHF1, (c) PHR, (d) PHT1;9, (e) PHT4;4, (f) SPX1, (g) SPX3, and (h) SPX4 in roots were determined 24 h after treatment with different endophytic PSB using qPCR. β-actin (Mol013347) was used as an internal reference gene to normalize the amount of template. Means of three replicates and standard errors are presented. The same letter above the column indicates no significant difference among the data in roots, respectively, at p < 0.05.
Results indicated that inoculation with endophytic PSB led to higher levels of PDR2, PHF1, PHR1, PHT1;9, PHT4;4, SPX1, SPX3, and SPX4 transcripts inoculated with endophytic PSB in orchid roots, suggesting their participation in phosphorus metabolism.
3.6. Colonization of PSB in Orchid Roots and Rhizosphere Soil
To track inoculated strains in plant tissues and soil, we utilized the rifampicin-resistant strains X1 and X13 (selected at 300 μg·mL−1) as biological markers. After 30 d of co-cultivation, these antibiotic-resistant strains were successfully re-isolated from both orchid roots and rhizosphere soil using rifampicin-supplemented medium. The colonization densities of X1 reached 2.8 × 104 cfu·g−1 in roots and 1.3 × 105 cfu·g−1 in rhizosphere soil for X1, with corresponding values of 2.1 × 104 cfu·g−1 and 1.0 × 105 cfu·g−1 for X13 (Figure 6).
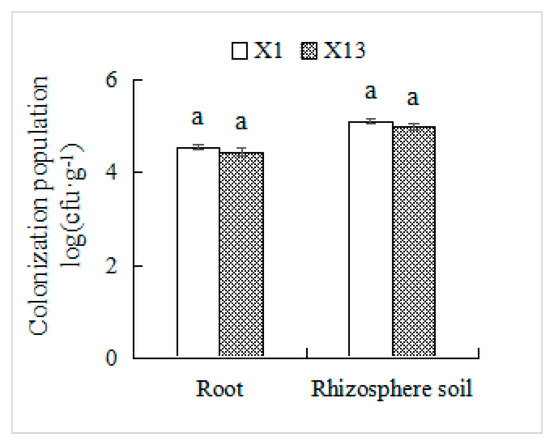
Figure 6.
Colonization population of endophytic PSB in Chinese Cymbidium root and rhizosphere soil. The root and rhizosphere soil samples of the orchid were diluted and uniformly spread on NA solid medium containing 300 µg·mL−1 rifampicin. After incubation at 30 °C for 72 h, the number of colonies on each plate was counted. Means of three replicates and standard errors are presented. Columns with the same letter indicate no significant difference at p < 0.05.
4. Discussion
Phosphorus is one of the most critical nutrients for plant growth. Phosphorus deficiency results in reduced plant growth [47]. Although chemical fertilizers can be added to the soil, plants can only utilize low amounts of phosphorus fertilizers [48,49]. As a result, choosing an effective PSB can increase the amount of phosphorus in the plant rhizosphere. In this study, the 30 isolates’ endophytic strains were proven to belong to Pseudomonas, Burkholderia, Bacillus, Rhizobium, Sphingobacterium, and Stenotrophomonas (Table S1). Members of these genera have been previously described as phosphorus solubilizers [8,12,50,51]. Ten endophytic strains showed clear halos on IP and OP solid medium, indicating their ability to dissolve inorganic phosphorus and mineralize organic phosphorus (Table S1). Ten endophytic strains revealed significant differences in phosphate-solubilizing halo zone diameter, colony diameters, and D/d among the strains, suggesting that each strain possesses a distinct capability to solubilize phosphorus (Figure 1 and Figure 2). The D/d produced by the 10 isolates varied from 2.2 to 3.5 mm (Figure 1d and Figure 2e). According to Marra et al. [52], PSB demonstrated a D/d exceeding 2.0, which indicates a general proficiency in phosphate solubilization. The dissolved inorganic phosphorus and mineralized organic phosphorus abilities of the endophytic strains isolated from Chinese Cymbidium in this study ranged from 130 to 346 mg·L−1 and 7.9 to 151.1 mg·L−1, respectively. More importantly, strains X13 and X1 demonstrated a distinctive capacity for simultaneously dissolving inorganic phosphorus and mineralizing organic phosphorus, as evidenced by their superior performance in both liquid and solid media (Figure 1 and Figure 2). This dual functionality is a notable advantage, as many reported PSB strains exhibit proficiency in only one type of phosphorus solubilization [53,54]. The robust and versatile P-solubilizing performance of these orchid-derived endophytic strains, coupled with their innate ability to colonize host plants, provides a strong theoretical basis for their application in the efficient production of orchids and the development of novel microbial fertilizers tailored for phosphorus-limited cultivation systems.
Previous studies have confirmed that distinct mechanisms between inorganic phosphorus and organic phosphorus solubilization by PSB strains are different, and there are also differences in the solubilization of different PSB strains [12]. Previous studies have demonstrated that the phosphorus-solubilizing ability of PSB is primarily attributed to their capacity to produce organic acids and extracellular polysaccharides, which mobilize inorganic phosphorus by reducing soil pH and chelating metal ions [12,17,18,24,55,56]. The negative correlation observed between the levels of soluble phosphorus released by bacteria and the pH of the supernatants in IP liquid medium containing tricalcium phosphate suggests that medium acidification due to organic acid production facilitates bacterial phosphate solubilization. This observation aligns with earlier reports indicating that decomposition of inorganic phosphorus is accompanied by a decrease in pH [18]. Consequently, we measured the content of organic acids and found that inoculation of PSB resulted in production of significant amounts of organic acids, particularly glucuronic acid, lactic acid, citric acid, tartaric acid, and pantothenic acid (Table 1). Numerous studies have demonstrated that one of the mechanisms by which microorganisms solubilize phosphorous and liberate phosphorus from insoluble phosphorous minerals is through production of gluconic acid by periplasmic cell-membrane-bound NADP-dependent glucose dehydrogenase [18,24]. The secreted organic acids lower soil pH, which increases phosphate solubility [17]. Moreover, PSB-produced exopolysaccharides can chelate metal ions, thereby preventing their recombination with phosphate ions [55]. Importantly, these EPS work in coordination with organic acids to effectively solubilize phosphates through combined acidification and chelation effects [56]. Therefore, the higher phosphate-solubilizing ability of strains X13 and X1, as compared to the lower phosphate-solubilizing ability strains X20 and X29, was associated with lower pH levels and higher organic acids and exopolysaccharides concentrations. Furthermore, PSB exhibit a complementary mechanism for enhancing phosphorus availability through the mineralization of organic phosphorus, a process predominantly mediated by extracellular enzymes such as phosphatases. All tested PSB strains demonstrated phosphatase activities, confirming their capacity for enzymatic hydrolysis of organic phosphorus. The higher phosphate-solubilizing ability of strains X13 and X1, when compared to the lower phosphate-solubilizing ability of strains X20 and X29, is associated with higher solubilized organic phosphorus and phosphatase activities (Figure 2). Phosphatases catalyze dephosphorylation of phosphoester bonds in organic compounds, releasing soluble phosphate [57]. This enzymatic system enables PSB to simultaneously mineralize organic phosphorus and solubilize inorganic phosphates, optimizing phosphorus cycling in soil ecosystems.
Given the observed phosphate-solubilizing capabilities of the selected strains in both solid and liquid media, we analyzed their efficacy in solubilizing phosphorus for incorporation into orchids grown in pots under greenhouse conditions. Studying the effects of PSB on the growth of orchids plantlets and unraveling the growth-promoting properties of these bacteria prove to be useful to orchids. Plant height, biomass, and other morphological growth indicators are direct manifestations of the efficiency of plantlets’ growth. As expected, orchids grown in a substrate with tricalcium phosphate and lecithin (CK) as the only source of phosphorus showed clear symptoms of phosphorus deficiency. A significant reduction in leaf length, root length, leaf dry weight, and root dry weight of orchids, as compared to orchids grown from plantlets of different PSB, was observed. Notably, All PSB treatments resulted in significantly greater leaf and root dry weight than the CK treatment (Figure 3). These data showed that most PSB promote the growth of orchid plantlets in terms of plant height and biomass. There were no adverse effects after inoculation of endophytic PSB in orchid plantlets. Soil is the primary reservoir of phosphorus in terrestrial ecosystems, directly influencing the availability of phosphorus for crops [58]. Soil phosphorus content is an important indicator of soil fertility, which reflects the storage and supply of nutrients in the soil. This study revealed that the effects of PSB on soil phosphorus content and microbial metabolites were significantly different (Figure 4). Specifically, the available phosphorus content increased, while total phosphorus decreased significantly (Figure 4). Additionally, the secretion of exopolysaccharides and phosphatase activity of microorganisms in the soil increased significantly, which, in turn, increased the phosphorus content in the orchid leaves and roots (Figure 3 and Figure 4). These results suggest that PSB strains convert insoluble soil phosphorus into a form that is utilizable by plants through the secretion of organic acids, exopolysaccharides, and phosphatases activities. Orchid roots can positively respond to soil phosphorus under phosphorus-deficient conditions so that orchid roots can actively absorb and utilize available phosphorus in soil to promote growth of orchid plantlets.
The uptake and transportation of phosphorus in plants are mediated by PDR, PHF, PHO, PHT, and SPXs, which play pivotal roles in the regulation of the absorption, utilization, and redistribution of phosphorus by plants [20,21,22]. When plants are subjected to phosphorus deficiency stress, PHR1 rapidly dissociates from the complex of regulatory genes and negative regulatory factors such as SPX1 and SPX2, thereby regulating the expression of phosphorus starvation response genes [59]. Additionally, regulatory factors involved in plant phosphorus starvation response (PSR) may affect the competition for phosphorus nutrients between plants and microbes, influence the expression levels of PSR transcription factors, and simultaneously stimulate the occurrence of PSR in plants [60]. Phosphate transporters also affect the phosphorous solubilization efficiency of the strain. Most bacteria have phosphate transport systems, which are activated when phosphorous levels are limited [61,62]. In this work, we evaluated the effect of PSB on the transcript levels of phosphorus-related genes in orchid root. The relative transcript levels of PDR2, PHF1, PHR, PHT1;9, and PHT4;4 were higher in inoculated orchid roots than in uninoculated roots (Figure 5). These results suggest that these strains’ PSB may solubilize insoluble phosphorus in the soil into soluble phosphorus for plant absorption and utilization, increase the phosphorus content in the environment, and then upregulate the transcript levels of PDR2, PHF1, PHR, PHT1;9, and PHT4;4 in orchid roots. One of the mechanisms by which PSB promotes orchid growth might be the strain-induced release of phosphorus from the soil, providing sufficient phosphorus for the growth of orchids. On the other hand, inoculation of a strain with PSB can upregulate or downregulate the transcript levels of the phosphorus-related genes in orchid roots, thereby enhancing phosphorus uptake from the soil solution and transporting it inside the plant, inside cells, between cells, and between plant organs. These results lay a solid foundation for additional research on the relationships between endophytic PSBs and their host plants. Furthermore, the results of the present work suggested that the endophytic PSBs X1 and X13 can colonize the orchid rhizosphere, as highly efficient phosphate-solubilizing endophytes would probably possess a natural affinity to their host plants This sustained colonization ensures persistent phosphate-solubilizing activity, demonstrating significant potential for promoting growth in Chinese Cymbidium orchids.
5. Conclusions
This study successfully isolated 10 endophytic bacterial strains with dual phosphate-solubilizing capabilities from the roots of Chinese Cymbidium. All strains demonstrated the ability to solubilize IP and mineralize OP through the secretion of organic acids and phosphatases. Among them, strains X1 (Paraburkholderia sp. Beta-32) and X13 (Rhizobium freirei PRF81) exhibited significantly stronger phosphate-solubilizing capacities compared to other strains, showing higher organic acid production, phosphatase activity, and phosphorus solubilization efficiency. Inoculation with these strains enhanced phosphorus uptake and promoted growth in orchids while upregulating the expression of phosphorus transporter-related genes. The most notable effects were observed in plants treated with X1 and X13. This study provides valuable microbial resources and a theoretical foundation for developing specialized microbial phosphorus fertilizers for orchids. Strains X1 and X13 show strong potential for practical applications, and further efforts should focus on field validation and large-scale application research.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms13102229/s1, Figure S1: Maximum likelihood phylogenetic tree of endophytic bacterial strains isolated from Chinese Cymbidium based on 16S rDNA gene sequences.; Figure S2: Transcript levels of the genes involved in phosphorus transport of different endophytic PSB in Chinese Cymbidium; Table S1: Preliminary identification of phosphate solubilization ability of bacterial strains isolated from Cymbidium orchids; Table S2: Strain names and their NCBI database accession numbers; Table S3: Primer sequences used for RT-qPCR and the accession numbers of the analyzed genes.
Author Contributions
Y.S.: writing—original draft, software, methodology, investigation, formal analysis, data curation. J.J.: methodology, investigation. X.W.: writing—review and editing, validation. W.Z.: writing—review and editing, validation. J.G.: project administration. J.L.: validation. Q.X.: validation. Y.W.: software, data curation. C.L.: resources. G.Z.: writing—review and editing. F.Y.: writing—review and editing, conceptualization, funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by National Key RD Program (2023YFD2300904), the Innovation Team of Modern Agriculture Industry Technology System in Guangdong Province (2024CXTD12), Guangdong Provincial Science and Technology Program Project (2024B1212060012), Hing-end Foreign Experts Recruitment plan of China (G2023030036L), Operation and Maintenance of Guangdong Famous and Excellent Flowers Germplasm Resource Nursery (2023-NBH-00-009), and the Guangdong Academy of Agricultural Sciences Discipline Team Construction Project (R2023PY-JG023).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Materials, further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yang, F.; Gao, J.; Wei, Y.; Ren, R.; Zhang, G.; Lu, C.; Jin, J.; Ai, Y.; Wang, Y.; Chen, L.; et al. The genome of Cymbidium sinense revealed the evolution of orchid traits. Plant Biotechnol. J. 2021, 19, 2501–2516. [Google Scholar] [CrossRef]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef]
- Shenoy, V.; Kalagudi, G.M. Enhancing plant phosphorus use efficiency for sustainable cropping. Biotechnol. Adv. 2005, 23, 501–513. [Google Scholar] [CrossRef]
- Herrera-Estrella, L.; López-Arredondo, D. Phosphorus: The underrated element for feeding the world. Trends Plant Sci. 2016, 21, 461–463. [Google Scholar] [CrossRef]
- Zhang, A.; Zhang, M.; Zhu, J.; Mei, Y.; Xu, F.; Bai, H.; Sun, K.; Zhang, W.; Dai, C.; Jia, Y. Endofungal bacterial microbiota promotes the absorption of chelated inorganic phosphorus by host pine through the ectomycorrhizal system. Microbiol. Spectr. 2023, 11, e00162-23. [Google Scholar] [CrossRef]
- Mei, Y.; Zhang, M.; Cao, G.; Zhu, J.; Zhang, A.; Bai, H.; Dai, C.; Jia, Y. Endofungal bacteria and ectomycorrhizal fungi synergistically promote the absorption of organic phosphorus in Pinus massoniana. Plant Cell Environ. 2024, 47, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Ahemad, M.; Zaidi, A.; Khan, M.S.; Oves, M. Biological importance of phosphorus and phosphate solubilizing microorganisms-an overview. In Phosphate Solubilizing Microbes for Crop Improvement; Khan, M.S., Zaidi, A., Eds.; Nova Science Publishers Inc.: New York, NY, USA, 2009; pp. 1–4. [Google Scholar]
- Zaidi, A.; Khan, M.S.; Ahmad, E.; Saif, S.; Rizvi, A.; Shahid, M. Growth stimulation and management of diseases of ornamental plants using phosphate solubilizing microorganisms: Current perspective. Acta Physiol. Plant 2016, 38, 117. [Google Scholar] [CrossRef]
- Singh, N.; Pandey, P.; Dubey, R.C.; Maheshwari, D.K. Biological control of root rot fungus Macrophomina phaseolina and growth enhancement of Pinus roxburghii (Sarg.) by rhizosphere competent Bacillus subtilis BN1. World J. Microb. Biot. 2008, 24, 1669–1679. [Google Scholar] [CrossRef]
- Bargaz, A.; Elhaissoufi, W.; Khourchi, S.; Benmrid, B.; Borden, K.A.; Rchiad, Z. Benefits of phosphate solubilizing bacteria on belowground crop performance for improved crop acquisition of phosphorus. Microbiol. Res. 2021, 252, 126842. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef]
- Cheng, Y.; Narayanan, M.; Shi, X.; Chen, X.; Li, Z.; Ma, Y. Phosphate-solubilizing bacteria: Their agroecological function and optimistic application for enhancing agro-productivity. Sci. Total Environ. 2023, 901, 166468. [Google Scholar] [CrossRef]
- Nico, M.; Ribaudo, C.M.; Gori, J.I.; Cantore, M.; Curá, J.A. Uptake of phosphate and promotion of vegetative growth in glucose-exuding rice plants (Oryza sativa) inoculated with plant growth-promoting bacteria. Appl. Soil Ecol. 2012, 61, 190–195. [Google Scholar] [CrossRef]
- Costa, E.M.; Lima, W.; Oliveira-Longatti, S.M.; Souza, F.M. Phosphate-solubilising bacteria enhance Oryza sativa growth and nutrient accumulation in an oxisol fertilized with rock phosphate. Ecol. Eng. 2015, 83, 380–385. [Google Scholar] [CrossRef]
- Pradhan, M.; Sahoo, R.K.; Pradhan, C.; Tuteja, N.; Mohanty, S. Contribution of native phosphorous-solubilizing bacteria of acid soils on phosphorous acquisition in peanut (Arachis hypogaea L.). Protoplasma 2017, 254, 2225–2236. [Google Scholar] [CrossRef]
- Li, H.; Ding, X.; Chen, C.; Zheng, X.; Han, H.; Li, C.; Gong, J.; Xu, T.; Li, Q.; Ding, G.; et al. Enrichment of phosphate solubilizing bacteria during late developmental stages of eggplant (Solanum melongena L.). FEMS Microbiol. Ecol. 2019, 95, fiz023. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Liu, J.; Jia, P.; Yang, T.; Zeng, Q.; Zhang, S.; Liao, B.; Shu, W.; Li, J. Novel phosphate-solubilizing bacteria enhance soil phosphorus cycling following ecological restoration of land degraded by mining. ISME J. 2020, 14, 1600–1613. [Google Scholar] [CrossRef]
- Khan, I.; Zada, S.; Rafiq, M.; Sajjad, W.; Zaman, S.; Hasan, F. Phosphate solubilizing epilithic and endolithic bacteria isolated from clastic sedimentary rocks, Murree lower Himalaya, Pakistan. Arch. Microbiol. 2022, 204, 332. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, Z.; Ahmad, M.; Raza, M.A.; Hilger, T.; Rasche, F. Phosphate-solubilizing bacillus sp. modulate soil exoenzyme activities and improve wheat growth. Microb. Ecol. 2024, 87, 31. [Google Scholar] [CrossRef]
- Kochian, L.V. Molecular physiology of mineral nutrient acquisition, transport, and utilization. In Biochemistry and Molecular Biology of Plants; Buchanan, B., Gruissem, W., Jones, R., Eds.; American Society of Plant Physiologists: Rockville, MD, USA, 2000; pp. 1222–1230. [Google Scholar]
- Cameron, D.D.; Johnson, I.; Leake, J.R.; Read, D.J.; Ren, Y.F. Mycorrhizal acquisition of inorganic phosphorus by the green-leaved terrestrial orchid Goodyera repens. Ann. Bot. 2007, 99, 831–834. [Google Scholar] [CrossRef]
- Paz-Ares, J.; Puga, M.I.; Rojas-Triana, M.; Martinez-Hevia, I.; Diaz, S.; Poza-Carrión, C.; Miñambres, M.; Leyva, A. Plant adaptation to low phosphorus availability: Core signaling, crosstalks, and applied implications. Mol. Plant 2022, 15, 104–124. [Google Scholar] [CrossRef]
- Othman, R.; Panhwar, Q.A. Phosphate-solubilizing bacteria improves nutrient uptake in Aerobic Rice. In Phosphate Solubilizing Microorganisms; Khan, M., Zaidi, A., Musarrat, J., Eds.; Springer: Cham, Switzerland, 2014. [Google Scholar] [CrossRef]
- Pande, A.; Pandey, P.; Mehra, S.; Singh, M.; Kaushik, S. Phenotypic and genotypic characterization of phosphate solubilizing bacteria and their efficiency on the growth of maize. J. Genet. Eng. Biotechnol. 2017, 15, 379–391. [Google Scholar] [CrossRef]
- Liu, X.; Jiang, X.; He, X.; Cao, Y.; Guo, T.; Li, T.; Ni, H.; Tang, X. Phosphate-solubilizing Pseudomonas sp. strain P34-L promotes wheat growth by colonizing the wheat rhizosphere and improving the wheat root system and soil phosphorus nutritional status. J. Plant Growth Regul. 2019, 38, 1314–1324. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, H.; Liu, L.; Li, S.; Xie, J.; Xue, X.; Jiang, Y. Screening of phosphate-solubilizing bacteria and their abilities of phosphorus solubilization and wheat growth promotion. BMC Microbiol. 2022, 22, 296. [Google Scholar] [CrossRef]
- McCormick, M.K.; Jacquemyn, H. What constrains the distribution of orchid populations? New Phytol. 2015, 202, 392–400. [Google Scholar] [CrossRef]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef]
- Fochi, V.; Chitarra, W.; Kohler, A.; Voyron, S.; Singan, V.R.; Lindquist, E.A.; Barry, K.W.; Girlanda, M.; Grigoriev, I.V.; Martin, F.; et al. Fungal and plant gene expression in the Tulasnella calospora-Serapias vomeracea symbiosis provides clues about nitrogen pathways in orchid mycorrhizas. New Phytol. 2017, 213, 365–379. [Google Scholar] [CrossRef]
- Selosse, M.A.; Petrolli, R.; Mujica, M.I.; Laurent, L.; Perez-Lamarque, B.; Figura, T.; Bourceret, A.; Jacquemyn, H.; Li, T.; Gao, J.; et al. The waiting room hypothesis revisited by orchids: Were orchid mycorrhizal fungi recruited among root endophytes? Ann. Bot. 2022, 129, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Zhang, P.; Qin, J.; Guo, J.; Deng, J. High-throughput sequencing-based analysis of the composition and diversity of endophytic bacteria community in tubers of Gastrodia elata f.glauca. Front. Microbiol. 2023, 13, 1092552. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, A.; Diez-Galán, A.; Cobos, R.; Calvo-Peña, C.; Barreiro, C.; Medina-Turienzo, J.; Sánchez-García, M.; Coque, J.J.R. Using rhizosphere phosphate solubilizing bacteria to improve barley (Hordeum vulgare) plant Productivity. Microorganisms 2021, 9, 1619. [Google Scholar] [CrossRef]
- Edwards, U.; Rogall, T.; Blöcker, H.; Emde, M.; Böttger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef]
- Altschul, S.; Modden, T.; Schafer, A.; Zhang, J.; Miller, W.; Lipman, D. Gapped BLAST and PSIBLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3404. [Google Scholar] [CrossRef]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Li, Z.; Li, J.; Liu, G.; Li, Y.; Wu, X.; Liang, J.; Wang, Z.; Chen, Q.; Peng, F. Isolation, Characterization and Growth-Promoting Properties of Phosphate-Solubilizing Bacteria (PSBs) Derived from Peach Tree Rhizosphere. Microorganisms 2025, 13, 718. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.; Dong, X.; Yang, H.; Chang, Y.S.; Xu, Z.; Che, F.; Wang, S.; Huang, W. Characterization of phosphate solubilizing bacteria in the sediments of eutrophic lakes and their potential for cyanobacterial recruitment. Chemosphere 2024, 352, 141276. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plan. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Gauri, S.S.; Mandal, S.S.; Gauri, S.M.; Mondal, K.C.; Dey, S.; Pati, B.R. Enhanced production and partial characterization of an extracellular polysaccharide from newly isolated Azotobacter sp. SSB81. Bioresour. Technol. 2009, 100, 4240–4243. [Google Scholar] [CrossRef]
- Fiori, J.; Amadesi, E.; Fanelli, F.; Tropeano, C.V.; Rugolo, M.; Gotti, R. Cellular and mitochondrial determination of low molecular mass organic acids by LC-MS/MS. J. Pharm. Biom. 2018, 150, 33–38. [Google Scholar] [CrossRef]
- Qvirist, L.; Carlsson, N.G.; Andlid, T. Assessing phytase activity-methods, definitions and pitfalls. J. Biol. Methods 2015, 2, e16. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Coy, R.M.; Held, D.W.; Kloepper, J.W. Rhizobacterial colonization of bermudagrass by Bacillus spp. in a Marvyn loamy sand soil. Appl. Soil Ecol. 2019, 141, 10–17. [Google Scholar] [CrossRef]
- Lin, Z.; Zhu, G.; Lu, C.Q.; Gao, J.; Li, J.; Xie, Q.; Wei, Y.; Jin, J.; Wang, F.; Yang, F. Functional conservation and divergence of SEPALLATA-like genes in floral development in Cymbidium sinense. Front. Plant Sci. 2023, 14, 1209834. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D.L. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Lin, W.; Hsiao, Y.; Chiou, T. Milestones in understanding transport, sensing, and signaling of the plant nutrient phosphorus. Plant Cell 2024, 36, 1504–1523. [Google Scholar] [CrossRef]
- Raymond, N.S.; Gómez-Muñoz, B.; van der Bom, F.J.T.; Nybroe, O.; Jensen, L.S.; Müller-Stöver, D.S.; Oberson, A.; Richardson, A.E. Phosphate-solubilising microorganisms for improved crop productivity: A critical assessment. New Phytol. 2021, 229, 1268–1277. [Google Scholar] [CrossRef]
- Lambers, H. Phosphorus acquisition and utilization in plants. Annu. Rev. Plant Biol. 2022, 73, 17–42. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Cai, B. Phosphate-solubilizing bacteria: Advances in their physiology, molecular mechanisms and microbial community effects. Microorganisms 2023, 11, 2904. [Google Scholar] [CrossRef]
- Fan, Y.; Lin, F.; Yang, L.; Zhong, X.; Wang, M.H.; Zhou, J.; Chen, Y.M.; Yang, Y. Decreased soil organic P fraction associated with ectomycorrhizal fungal activity to meet increased P demand under N application in a subtropical forest ecosystem. Biol. Fertil. Soils 2018, 54, 149–161. [Google Scholar] [CrossRef]
- Marra, L.M.; Sousa Soares, C.R.F.; de Oliveira, S.M.; Ferreira, P.A.A.; Soares, B.L.; de Carvalho, R.F.; de Moreira, F.M.S. Biological nitrogen fixation and phosphate solubilization by bacteria isolated from tropical soils. Plant Soil 2012, 357, 289–307. [Google Scholar] [CrossRef]
- Nautiyal, C.S.; Bhadauria, S.; Kumar, P.; Lal, H.; Mondal, R.; Verma, D. Stress induced phosphate solubilization in bacteria isolated from alkaline soils. FEMS Microbiol. Lett. 2000, 182, 291–296. [Google Scholar] [CrossRef]
- Chen, Y.P.; Rekha, P.D.; Arun, A.B.; Shen, F.T.; La, W.-A.; Young, C.C. Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities. Appl. Soil Ecol. 2006, 34, 33–41. [Google Scholar] [CrossRef]
- Yi, Y.; Huang, W.; Ge, Y. Exopolysaccharide: A novel important factor in the microbial dissolution of tricalcium phosphate. World J. Microb. Biot. 2008, 24, 1059–1065. [Google Scholar] [CrossRef]
- Fatima, T.; Mishra, I.; Verma, R.; Arora, N.K. Mechanisms of halotolerant plant growth promoting Alcaligenes sp. involved in salt tolerance and enhancement of the growth of rice under salinity stress. Biotech 2020, 10, 361. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Han, Q.; Liu, Q.; Gan, Y.; Rensing, C.; Rivera, W.; Zhao, Q.; Zhang, J. Roles of phosphate-solubilizing bacteria in mediating soil legacy phosphorus availability. Microbiol. Res. 2023, 272, 127375. [Google Scholar] [CrossRef]
- Chen, J.; Zhao, G.; Wei, Y.; Dong, Y.; Hou, L.; Jiao, R. Isolation and screening of multifunctional phosphate solubilizing bacteria and its growth-promoting effect on Chinese fir seedling. Sci. Rep. 2021, 11, 9081. [Google Scholar] [CrossRef]
- González, E.; Solano, R.; Rubio, V.; Leyva, A.; Paz-Ares, J. Phosphate transporter traffic facilitator is a plant-specific SEC12-related protein that enables the endoplasmic reticulum exit of a high-affinity phosphate transporter in Arabidopsis. Plant Cell 2005, 17, 3500–3512. [Google Scholar] [CrossRef]
- Castrillo, G.; Teixeira, P.J.P.L.; Paredes, S.H.; Theresa, F.; Law, T.F.; de Lorenzo, L.; Feltcher, M.E.; Finkel, O.M.; Breakfield, N.W.; Mieczkowski, P.; et al. Root microbiota drive direct integration of phosphate stress and immunity. Nature 2017, 543, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Anawar, H.M.; Rengel, Z.; Damon, P.; Tibbett, M. Arsenic-phosphorus interactions in the soil-plant-microbe system: Dynamics of uptake, suppression and toxicity to plants. Environ. Pollut. 2018, 233, 1003–1012. [Google Scholar] [CrossRef]
- Dissanayaka, D.M.S.B.; Ghahremani, M.; Siebers, M.; Wasaki, J.; Plaxton, W.C. Recent insights into the metabolic adaptations of phosphorus-deprived plants. J. Exp. Bot. 2020, 72, 199–223. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).