
Influence of Arbuscular Mycorrhizal Fungi on Nitrogen Dynamics During Cinnamomum camphora Litter Decomposition

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site Description and Litter Preparation
2.2. Experimental Design
2.3. Measurement of Mass Loss, Mycorrhiazl Colonization, and Protein Degradation
2.4. Measurement of Extracellular Enzyme Activities and N Mineralization
2.5. Statistical Analyses
3. Results
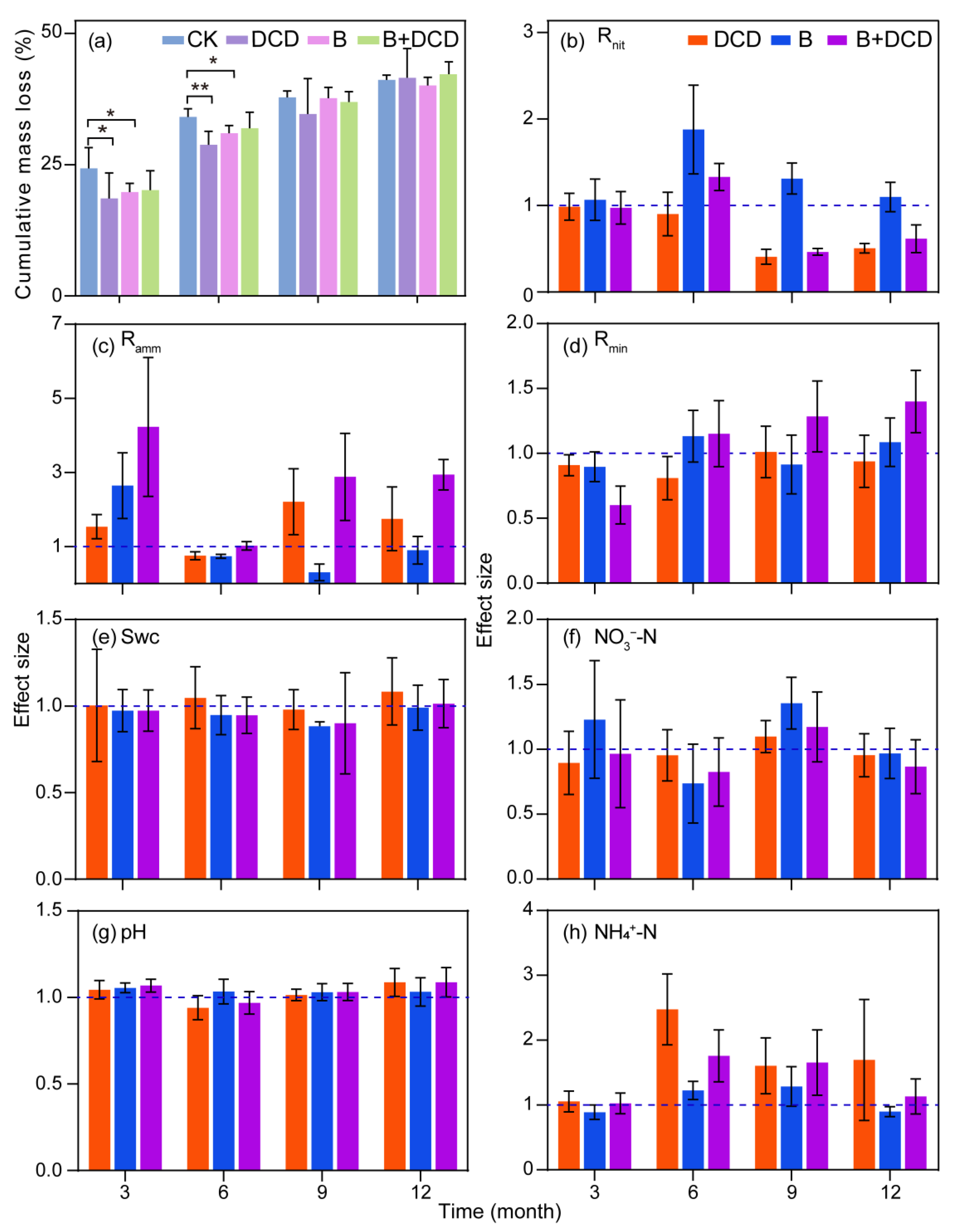
3.1. Decomposition Dynamics of C. camphora Litter
3.2. Soil Nitrogen Mineralization Dynamic
3.3. Dynamics in Extracellular Enzyme Activity
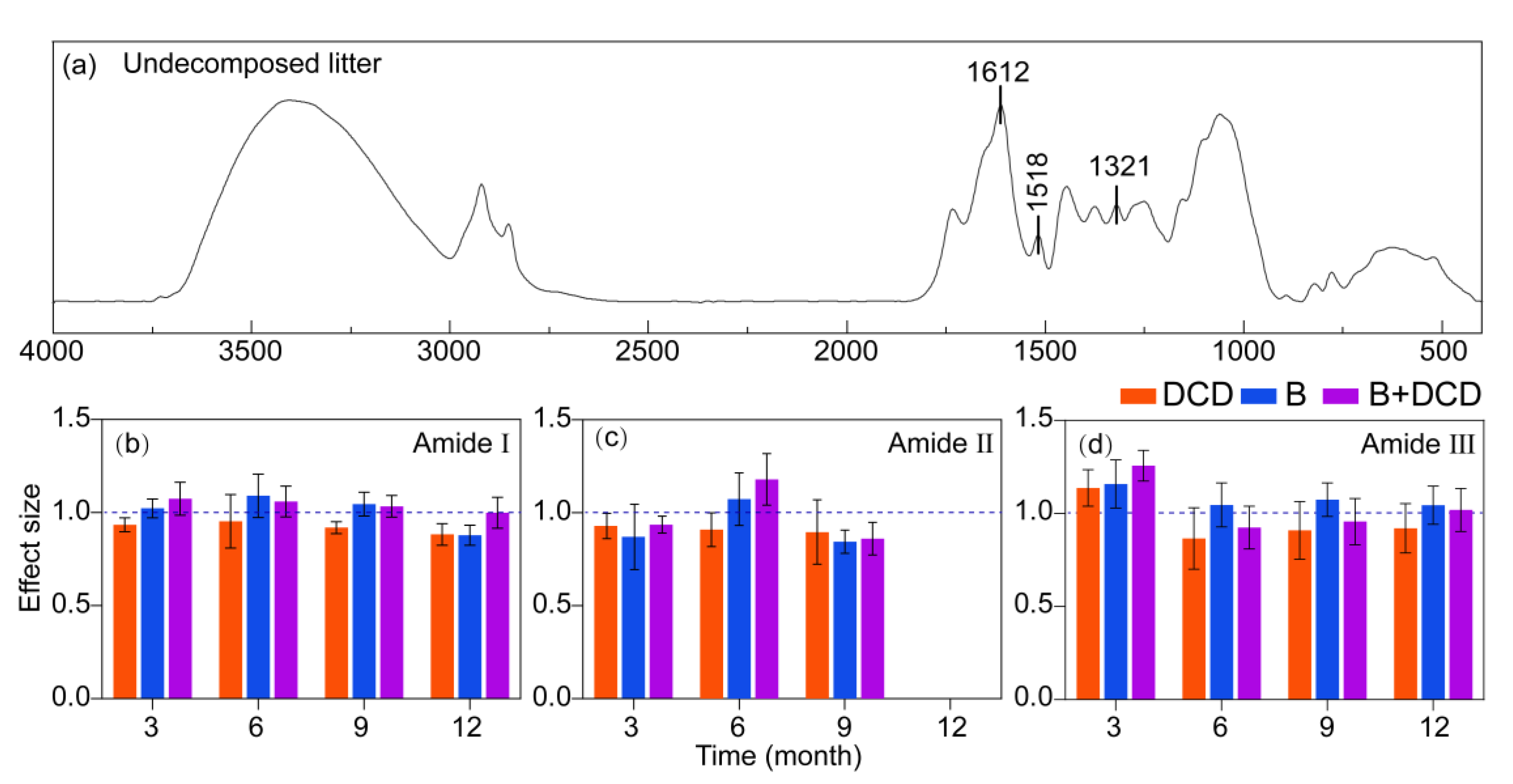
3.4. Dynamics in Litter Protein Degradation
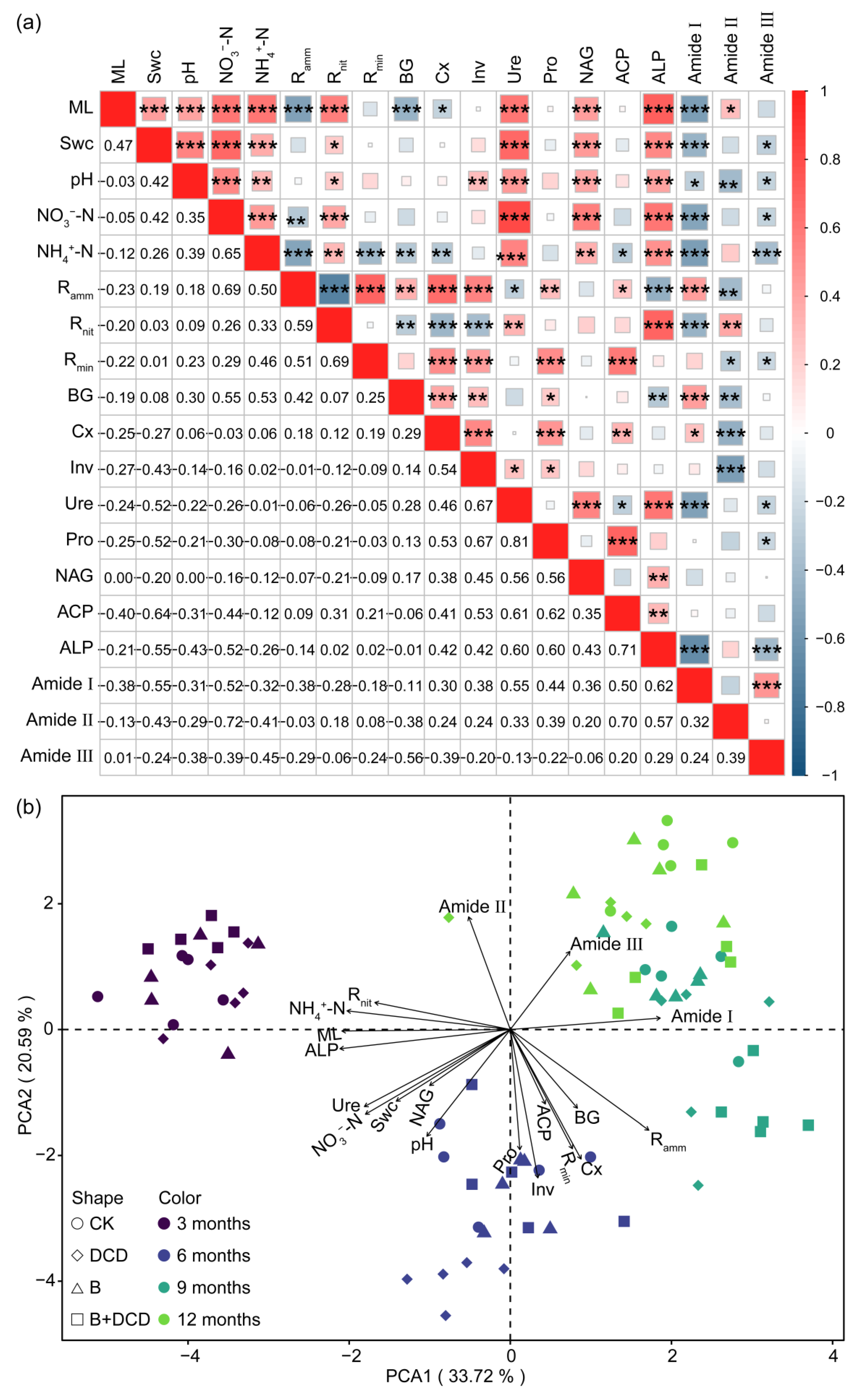
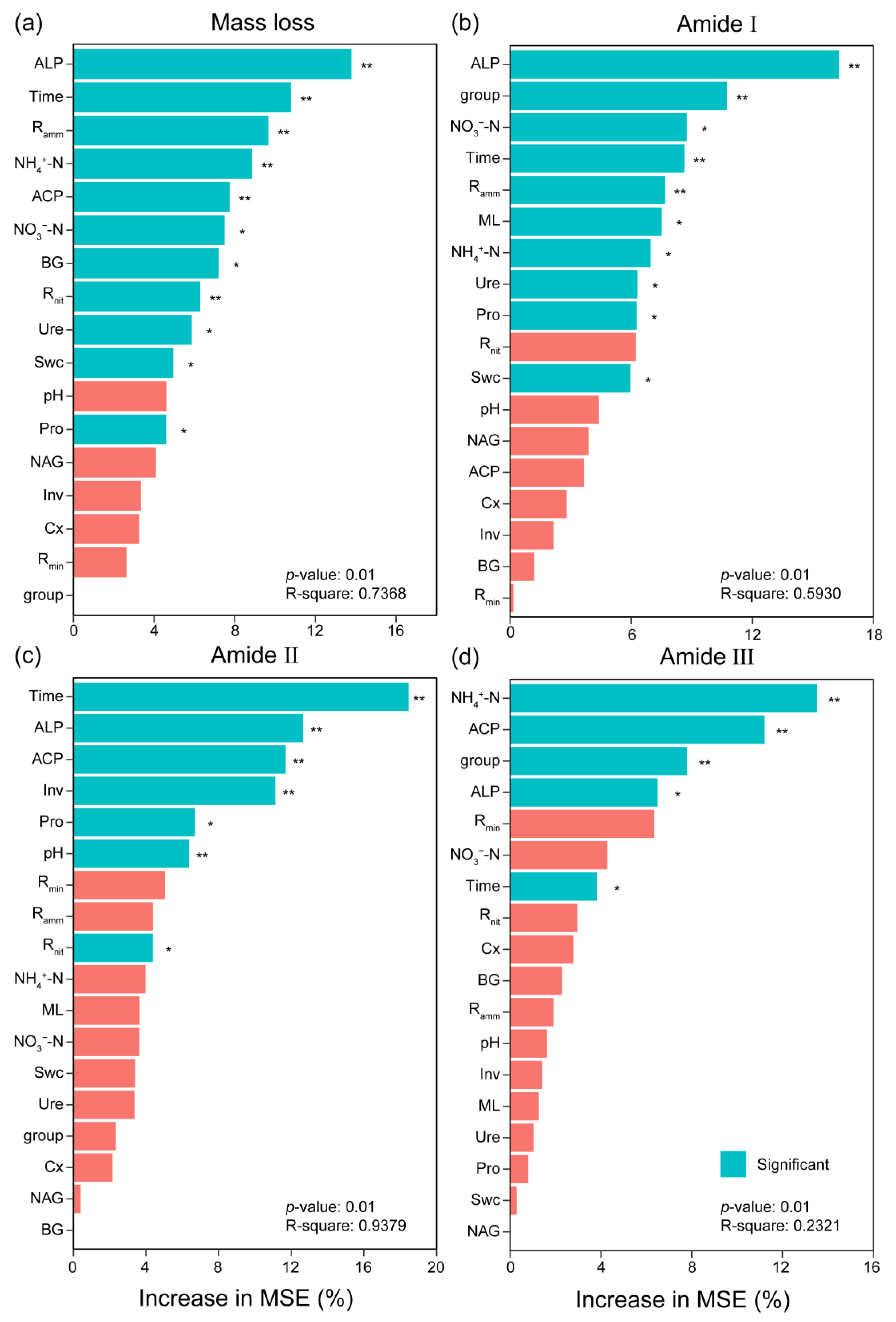
3.5. The Relationships Among Litter and Soil Physicochemical Properties
4. Discussion
4.1. Litter Decomposition
4.2. Soil N Mineralization Dynamics
4.3. Extracellular Enzyme Activities
4.4. Litter Protein Degradation
4.5. Limitiations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xi, L.; Chen, S.; Bian, H.; Peng, Z.; Niu, Y.; Li, Y. Organic carbon release from litter decomposition of woody and herbaceous plants in the Dongting Lake wetlands: A comparative study. Ecohydrol. Hydrobiol. 2023, 23, 408–419. [Google Scholar] [CrossRef]
- Prescott, C.E.; Vesterdal, L. Decomposition and transformations along the continuum from litter to soil organic matter in forest soils. For. Ecol. Manag. 2021, 498, 119522. [Google Scholar] [CrossRef]
- Mason, R.E.; Craine, J.M.; Lany, N.K.; Jonard, M.; Ollinger, S.V.; Groffman, P.M.; Fulweiler, R.W.; Angerer, J.; Read, Q.D.; Reich, P.B.; et al. Evidence, causes, and consequences of declining nitrogen availability in terrestrial ecosystems. Science 2022, 376, eabh3767. [Google Scholar] [CrossRef] [PubMed]
- Noll, L.; Zhang, S.; Wanek, W. Novel high-throughput approach to determine key processes of soil organic nitrogen cycling: Gross protein depolymerization and microbial amino acid uptake. Soil Biol. Biochem. 2019, 130, 73–81. [Google Scholar] [CrossRef]
- Feng, J.; Turner, B.L.; Wei, K.; Tian, J.; Chen, Z.; Lü, X.; Wang, C.; Chen, L. Divergent composition and turnover of soil organic nitrogen along a climate gradient in arid and semiarid grasslands. Geoderma 2018, 327, 36–44. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bennett, J. Nitrogen Mineralization: Challenges of a Changing Paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Khan, M.F.; Chowdhary, S.; Koksch, B.; Murphy, C.D. Biodegradation of Amphipathic Fluorinated Peptides Reveals a New Bacterial Defluorinating Activity and a New Source of Natural Organofluorine Compounds. Environ. Sci. Technol. 2023, 57, 9762–9772. [Google Scholar] [CrossRef]
- Geisseler, D.; Horwath, W.R.; Joergensen, R.G.; Ludwig, B. Pathways of nitrogen utilization by soil microorganisms—A review. Soil Biol. Biochem. 2010, 42, 2058–2067. [Google Scholar] [CrossRef]
- Phillips, R.P.; Edward, B.; Midgley, M.G. The mycorrhizal-associated nutrient economy: A new framework for predicting carbon-nutrient couplings in temperate forests. New Phytol. 2013, 199, 41–51. [Google Scholar] [CrossRef]
- Baldrian, P.; López-Mondéjar, R.; Kohout, P. Forest microbiome and global change. Nat. Rev. Microbiol. 2023, 21, 487–501. [Google Scholar] [CrossRef]
- Brundrett, M.C.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.; Hodge, A.; Fitter, A.H. Arbuscular mycorrhizal fungi can transfer substantial amounts of nitrogen to their host plant from organic material. New Phytol. 2009, 181, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Koller, R.; Rodriguez, A.; Robin, C.; Scheu, S.; Bonkowski, M. Protozoa enhance foraging efficiency of arbuscular mycorrhizal fungi for mineral nitrogen from organic matter in soil to the benefit of host plants. New Phytol. 2013, 199, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Bunn, R.A.; Simpson, D.T.; Bullington, L.S.; Lekberg, Y.; Janos, D.P. Revisiting the ‘direct mineral cycling’ hypothesis: Arbuscular mycorrhizal fungi colonize leaf litter, but why? ISME J. 2019, 13, 1891–1898. [Google Scholar] [CrossRef]
- Nottingham, A.T.; Turner, B.L.; Winter, K.; Chamberlain, P.M.; Stott, A.; Tanner, E.V. Root and arbuscular mycorrhizal mycelial interactions with soil microorganisms in lowland tropical forest. FEMS Microbiol. Ecol. 2013, 85, 37–50. [Google Scholar] [CrossRef]
- Averill, C.; Turner, B.L.; Finzi, A.C. Mycorrhiza-mediated competition between plants and decomposers drives soil carbon storage. Nature 2014, 505, 543–545. [Google Scholar] [CrossRef]
- Gui, H.; Hyde, K.; Xu, J.; Mortimer, P. Arbuscular mycorrhiza enhance the rate of litter decomposition while inhibiting soil microbial community development. Sci. Rep. 2017, 7, 42184. [Google Scholar] [CrossRef]
- Hodge, A.; Campbell, C.D.; Fitter, A.H. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature 2001, 413, 297–299. [Google Scholar] [CrossRef]
- Lindahl, B.D.; Ihrmark, K.; Boberg, J.; Trumbore, S.E.; Högberg, P.; Stenlid, J.; Finlay, R.D. Spatial separation of litter decomposition and mycorrhizal nitrogen uptake in a boreal forest. New Phytol. 2007, 173, 611–620. [Google Scholar] [CrossRef]
- Bodeker, I.T.; Clemmensen, K.E.; de Boer, W.; Martin, F.; Olson, A.; Lindahl, B.D. Ectomycorrhizal Cortinarius species participate in enzymatic oxidation of humus in northern forest ecosystems. New Phytol. 2014, 203, 245–256. [Google Scholar] [CrossRef]
- Geisseler, D.; Horwath, W.R. Regulation of extracellular protease activity in soil in response to different sources and concentrations of nitrogen and carbon. Soil Biol. Biochem. 2008, 40, 3040–3048. [Google Scholar] [CrossRef]
- Allison, S.D.; Vitousek, P.M. Responses of extracellular enzymes to simple and complex nutrient inputs. Soil Biol. Biochem. 2005, 37, 937–944. [Google Scholar] [CrossRef]
- Sayed, F.; Abdelaleem, F.; Goda, S. Protein utilization and release of extracellular proteinase by two root-rot fungi. Biol. Plant. 1994, 36, 613. [Google Scholar] [CrossRef]
- McCarty, G.; Shogren, D.; Bremner, J. Regulation of urease production in soil by microbial assimilation of nitrogen. Biol. Fertil. Soils 1992, 12, 261–264. [Google Scholar] [CrossRef]
- Vallon, O.; Bulté, L.; Kuras, R.; Olive, J.; Wollman, F.A. Extensive accumulation of an extracellular l-amino-acid oxidase during gametogenesis of Chlamydomonas reinhardtii. Eur. J. Biochem. 1993, 215, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Yang, L.; Kong, D.; Zhao, H. Differential effects of ammonium and nitrate addition on soil microbial biomass, enzymatic activities, and organic carbon in a temperate forest in North China. Plant Soil 2022, 481, 595–606. [Google Scholar] [CrossRef]
- Dudáš, M.; Pjevac, P.; Kotianová, M.; Gančarčíková, K.; Rozmoš, M.; Hršelová, H.; Bukovská, P.; Jansa, J. Arbuscular Mycorrhiza and Nitrification: Disentangling Processes and Players by Using Synthetic Nitrification Inhibitors. Appl. Environ. Microbiol. 2022, 88, e0136922. [Google Scholar] [CrossRef]
- Cao, T.; Fang, Y.; Chen, Y.; Kong, X.; Yang, J.; Alharbi, H.; Kuzyakov, Y.; Tian, X. Synergy of saprotrophs with mycorrhiza for litter decomposition and hotspot formation depends on nutrient availability in the rhizosphere. Geoderma 2022, 410, 115662. [Google Scholar] [CrossRef]
- Xu, H.; Shao, H.; Lu, Y. Arbuscular mycorrhiza fungi and related soil microbial activity drive carbon mineralization in the maize rhizosphere. Ecotoxicol. Environ. Saf. 2019, 182, 109476. [Google Scholar] [CrossRef]
- Kong, X.; Jia, Y.; Song, F.; Tian, K.; Lin, H.; Bei, Z.; Jia, X.; Yao, B.; Guo, P.; Tian, X. Insight into litter decomposition driven by nutrient demands of symbiosis system through the hypha bridge of arbuscular mycorrhizal fungi. Environ. Sci. Pollut. Res. 2018, 25, 5369–5378. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, X.; Li, G.; Qin, P. Interactions between arbuscular mycorrhizal fungi and phosphate-solubilizing fungus (Mortierella sp.) and their effects on Kostelelzkya virginica growth and enzyme activities of rhizosphere and bulk soils at different salinities. Biol. Fertil. Soils 2011, 47, 543–554. [Google Scholar] [CrossRef]
- Cheng, L.; Booker, F.L.; Tu, C.; Burkey, K.O.; Zhou, L.; Shew, H.D.; Rufty, T.W.; Hu, S. Arbuscular Mycorrhizal Fungi Increase Organic Carbon Decomposition Under Elevated CO2. Science 2012, 337, 1084–1087. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Liu, N.; Lu, W.; Wang, S.; Kan, H.; Zhang, Y.; Xu, L.; Chen, Y. The interaction between arbuscular mycorrhizal fungi and soil phosphorus availability influences plant community productivity and ecosystem stability. J. Ecol. 2014, 102, 1072–1082. [Google Scholar] [CrossRef]
- Wang, X.X.; Wang, X.; Sun, Y.; Cheng, Y.; Liu, S.; Chen, X.; Feng, G.; Kuyper, T.W. Arbuscular Mycorrhizal Fungi Negatively Affect Nitrogen Acquisition and Grain Yield of Maize in a N Deficient Soil. Front. Microbiol. 2018, 9, 418. [Google Scholar] [CrossRef]
- Di, H.J.; Cameron, K.C.; Shen, J.P.; Winefield, C.S.; O’Callaghan, M.; Bowatte, S.; He, J.Z. Nitrification driven by bacteria and not archaea in nitrogen-rich grassland soils. Nat. Geosci. 2009, 2, 621–624. [Google Scholar] [CrossRef]
- O’Callaghan, M.; Gerard, E.M.; Carter, P.E.; Lardner, R.; Sarathchandra, U.; Burch, G.; Ghani, A.; Bell, N. Effect of the nitrification inhibitor dicyandiamide (DCD) on microbial communities in a pasture soil amended with bovine urine. Soil Biol. Biochem. 2010, 42, 1425–1436. [Google Scholar] [CrossRef]
- Wu, C.; Kong, X.; He, X.; Lin, Y.; He, Z.; Gao, Y.; Kong, Q. Effects of arbuscular mycorrhizal fungi on microbial activity and nutrient release are sensitive to acid deposition during litter decomposition in a subtropical Cinnamomum camphora forest. iForest—Biogeosci. For. 2023, 16, 314–324. [Google Scholar] [CrossRef]
- Lin, H.; He, Z.; Hao, J.; Tian, K.; Jia, X.; Kong, X.; Akbar, S.; Bei, Z.; Tian, X. Effect of N addition on home-field advantage of litter decomposition in subtropical forests. For. Ecol. Manag. 2017, 398, 216–225. [Google Scholar] [CrossRef]
- Li, L.; Zhang, Q.; Yuan, X.; Yang, H.; Qin, S.; Hong, L.; Pu, L.; Li, L.; Zhang, P.; Zhang, J. Study of the molecular structure of proteins in fermented Maize-Soybean meal-based rations based on FTIR spectroscopy. Food Chem. 2024, 441, 138310. [Google Scholar] [CrossRef]
- Cai, S.; Singh, B.R. Identification of β-turn and random coil amide III infrared bands for secondary structure estimation of proteins. Biophys. Chem. 1999, 80, 7–20. [Google Scholar] [CrossRef]
- Sadat, A.; Joye, I.J. Peak Fitting Applied to Fourier Transform Infrared and Raman Spectroscopic Analysis of Proteins. Appl. Sci. 2020, 10, 5918. [Google Scholar] [CrossRef]
- Demyan, M.S.; Rasche, F.; Schulz, E.; Breulmann, M.; Müller, T.; Cadisch, G. Use of specific peaks obtained by diffuse reflectance Fourier transform mid-infrared spectroscopy to study the composition of organic matter in a Haplic Chernozem. Eur. J. Soil Sci. 2012, 63, 189–199. [Google Scholar] [CrossRef]
- Trap, J.; Bureau, F.; Akpa-Vinceslas, M.; Decaens, T.; Aubert, M. Changes in humus forms and soil N pathways along a 130-year-old pure beech forest chronosequence. Ann. For. Sci. 2011, 68, 595–606. [Google Scholar] [CrossRef]
- Zheng, H.; Wang, Z.; Deng, X.; Herbert, S.; Xing, B. Impacts of adding biochar on nitrogen retention and bioavailability in agricultural Soil. Geoderma 2013, 206, 32–39. [Google Scholar] [CrossRef]
- Olson, J.S. Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Surewicz, W.K.; Mantsch, H.H. New insight into protein secondary structure from resolution-enhanced infrared spectra. Biochim. Biophys. Acta 1988, 952, 115–130. [Google Scholar] [CrossRef]
- Ning, J.; Ai, S.; Cui, L. Dicyandiamide has more inhibitory activities on nitrification than thiosulfate. PLoS ONE 2018, 13, e0200598. [Google Scholar] [CrossRef]
- Meng, Y.; Wang, J.J.; Wei, Z.; Dodla, S.K.; Fultz, L.M.; Gaston, L.A.; Xiao, R.; Park, J.-h.; Scaglia, G. Nitrification inhibitors reduce nitrogen losses and improve soil health in a subtropical pastureland. Geoderma 2021, 388, 114947. [Google Scholar] [CrossRef]
- Storer, K.; Coggan, A.; Ineson, P.; Hodge, A. Arbuscular mycorrhizal fungi reduce nitrous oxide emissions from N2O hotspots. New Phytol. 2018, 220, 1285–1295. [Google Scholar] [CrossRef]
- Boot, C.M.; Hall, E.K.; Denef, K.; Baron, J.S. Long-term reactive nitrogen loading alters soil carbon and microbial community properties in a subalpine forest ecosystem. Soil Biol. Biochem. 2016, 92, 211–220. [Google Scholar] [CrossRef]
- Moore, J.A.M.; Anthony, M.A.; Pec, G.J.; Trocha, L.K.; Trzebny, A.; Geyer, K.M.; van Diepen, L.T.A.; Frey, S.D. Fungal community structure and function shifts with atmospheric nitrogen deposition. Glob. Chang. Biol. 2021, 27, 1349–1364. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.J.; Di, H.J.; Cameron, K.C.; Li, B.; Podolyan, A.; Moir, J.L.; Monaghan, R.M.; Smith, L.C.; O’Callaghan, M.; Bowatte, S.; et al. Effect of 7-year application of a nitrification inhibitor, dicyandiamide (DCD), on soil microbial biomass, protease and deaminase activities, and the abundance of bacteria and archaea in pasture soils. J. Soils Sediments 2013, 13, 753–759. [Google Scholar] [CrossRef]
- Wang, L.; George, T.S.; Feng, G. Concepts and consequences of the hyphosphere core microbiome for arbuscular mycorrhizal fungal fitness and function. New Phytol. 2024, 242, 1529–1533. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, T.; Chenu, C.; Genet, P.; Barot, S.; Perveen, N.; Mougin, C.; Fontaine, S. Contribution of exudates, arbuscular mycorrhizal fungi and litter depositions to the rhizosphere priming effect induced by grassland species. Soil Biol. Biochem. 2015, 80, 146–155. [Google Scholar] [CrossRef]
- Leifheit, E.F.; Verbruggen, E.; Rillig, M.C. Arbuscular mycorrhizal fungi reduce decomposition of woody plant litter while increasing soil aggregation. Soil Biol. Biochem. 2015, 81, 323–328. [Google Scholar] [CrossRef]
- Wu, C.; Kong, X.; He, X.; Song, F.; Lin, Y.; Jia, Y.; Kurakov, A.V.; He, Z. The Biotic and Abiotic Factors of Regulation of Arbuscular Mycorrhizal Fungi Activity in Litter Decomposition: Review. Eurasian Soil Sci. 2022, 55, 1446–1459. [Google Scholar] [CrossRef]
- Riggs, C.E.; Hobbie, S.E.; Cavender-Bares, J.; Savage, J.A.; Wei, X. Contrasting effects of plant species traits and moisture on the decomposition of multiple litter fractions. Oecologia 2015, 179, 573–584. [Google Scholar] [CrossRef]
- Stres, B.; Danevčič, T.; Pal, L.; Fuka, M.M.; Resman, L.; Leskovec, S.; Hacin, J.; Stopar, D.; Mahne, I.; Mandic-Mulec, I. Influence of temperature and soil water content on bacterial, archaeal and denitrifying microbial communities in drained fen grassland soil microcosms. FEMS Microbiol. Ecol. 2008, 66, 110–122. [Google Scholar] [CrossRef]
- Xiang, X.; De, K.; Lin, W.; Feng, T.; Li, F.; Wei, X.; Wang, W. Different fates and retention of deposited NH4+ and NO3− in the alpine grasslands of the Qinghai-Tibet plateau. Ecol. Indic. 2024, 158, 111415. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, J.; Zhang, J.-B.; Mary, B.; Cai, Z.-C. The mechanisms behind reduced NH4+ and NO3− accumulation due to litter decomposition in the acidic soil of subtropical forest. Plant Soil 2014, 378, 295–308. [Google Scholar] [CrossRef]
- Pauwels, R.; Graefe, J.; Bitterlich, M. An arbuscular mycorrhizal fungus alters soil water retention and hydraulic conductivity in a soil texture specific way. Mycorrhiza 2023, 33, 165–179. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.; He, Y.; Xu, X.; Umer, M.; Liu, X.; Xia, T.; Guo, Y.; Wu, B.; Xu, H.; Zang, L.; et al. Effects of AMF on plant nutrition and growth depend on substrate gravel content and patchiness in the karst species Bidens pilosa L. Front. Plant Sci. 2022, 13, 968719. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Yu, M.; Chen, H.; Zhao, H.; Huang, Y.; Su, W.; Xia, F.; Chang, S.X.; Brookes, P.C.; Dahlgren, R.A.; et al. Elevated temperature shifts soil N cycling from microbial immobilization to enhanced mineralization, nitrification and denitrification across global terrestrial ecosystems. Glob. Chang. Biol. 2020, 26, 5267–5276. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Zhou, J.; Tang, S.; Wu, L.; Ma, Q.; Marsden, K.A.; Chadwick, D.R.; Jones, D.L. Utilisation and transformation of organic and inorganic nitrogen by soil microorganisms and its regulation by excessive carbon and nitrogen availability. Biol. Fertil. Soils 2023, 59, 379–389. [Google Scholar] [CrossRef]
- Fang, C.; Xie, J.; Yang, X.; Yang, R.; Fransson, P.; Sheteiwy, M.S.; Weih, M.; Yang, H. Arbuscular mycorrhizal fungi drive soil nitrogen transformation under wheat varieties with different nitrogen utilization efficiencies. Rhizosphere 2023, 27, 100775. [Google Scholar] [CrossRef]
- Basiru, S.; Mhand, K.A.S.; Hijri, M. Deciphering the mechanisms through which arbuscular mycorrhizal symbiosis reduces nitrogen losses in agroecosystems. Appl. Soil Ecol. 2025, 206, 105799. [Google Scholar] [CrossRef]
- Veresoglou, S.D.; Sen, R.; Mamolos, A.P.; Veresoglou, D.S. Plant species identity and arbuscular mycorrhizal status modulate potential nitrification rates in nitrogen-limited grassland soils. J. Ecol. 2011, 99, 1339–1349. [Google Scholar] [CrossRef]
- Chen, Y.-L.; Chen, B.-D.; Hu, Y.-J.; Li, T.; Zhang, X.; Hao, Z.-P.; Wang, Y.-S. Direct and indirect influence of arbuscular mycorrhizal fungi on abundance and community structure of ammonia oxidizing bacteria and archaea in soil microcosms. Pedobiologia 2013, 56, 205–212. [Google Scholar] [CrossRef]
- Veresoglou, S.D.; Verbruggen, E.; Makarova, O.; Mansour, I.; Sen, R.; Rillig, M.C. Arbuscular Mycorrhizal Fungi Alter the Community Structure of Ammonia Oxidizers at High Fertility via Competition for Soil NH4+. Microb. Ecol. 2019, 78, 147–158. [Google Scholar] [CrossRef]
- He, C.; Ruan, Y.; Jia, Z. Effects of Nitrogen Addition on Soil Microbial Biomass: A Meta-Analysis. Agriculture 2024, 14, 1616. [Google Scholar] [CrossRef]
- Liu, B.; Fan, X.; Meng, D.; Liu, Z.; Gao, D.; Chang, Q.; Bai, E. Ectomycorrhizal trees rely on nitrogen resorption less than arbuscular mycorrhizal trees globally. Ecol. Lett. 2024, 27, e14346. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Zhang, X.; Du, J.; Gilliam, F.S.; Yang, S.; Tian, M.; Zhang, C.; Zhou, Y. Arbuscular Mycorrhizal Fungi Shift Soil Bacterial Community Composition and Reduce Soil Ammonia Volatilization and Nitrous Oxide Emissions. Microb. Ecol. 2023, 85, 951–964. [Google Scholar] [CrossRef] [PubMed]
- Mooshammer, M.; Hofhansl, F.; Frank, A.H.; Wanek, W.; Hammerle, I.; Leitner, S.; Schnecker, J.; Wild, B.; Watzka, M.; Keiblinger, K.M.; et al. Decoupling of microbial carbon, nitrogen, and phosphorus cycling in response to extreme temperature events. Sci. Adv. 2017, 3, e1602781. [Google Scholar] [CrossRef]
- Conceicao, T.A.; Andrade, G.; Brito, I. Influence of Intact Mycelium of Arbuscular Mycorrhizal Fungi on Soil Microbiome Functional Profile in Wheat under Mn Stress. Plants 2022, 11, 2598. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Xie, Y.; Yang, Y.; Jing, C.; You, X.; Yang, J.; Sun, C.; Qin, S.; Chen, J.; Cao, K.; et al. AMF colonization affects allelopathic effects of Zea mays L. root exudates and community structure of rhizosphere bacteria. Front. Plant Sci. 2022, 13, 1050104. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Wu, W.; Tian, K.; Jia, Y.; Siddiq, A.; Lin, H.; Tian, X. Promotive performance of shrimp Neocaridina denticulata on Typha angustifolia leaf litter decomposition. Hydrobiologia 2019, 827, 75–87. [Google Scholar] [CrossRef]
- L’Espérance, E.; Bouyoucef, L.S.; Dozois, J.A.; Yergeau, E. Tipping the plant-microbe competition for nitrogen in agricultural soils. iScience 2024, 27, 110973. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, W.; Nessa, A.; Yang, S.; Zhang, W.; Zhang, M. Metagenomic insight into antagonistic effects of a nitrification inhibitor and phosphate-solubilizing bacteria on soil protease activity. Land Degrad. Dev. 2023, 35, 455–466. [Google Scholar] [CrossRef]
- Wang, G.; Jin, Z.; George, T.S.; Feng, G.; Zhang, L. Arbuscular mycorrhizal fungi enhance plant phosphorus uptake through stimulating hyphosphere soil microbiome functional profiles for phosphorus turnover. New Phytol. 2023, 238, 2578–2593. [Google Scholar] [CrossRef]
- Jiang, F.; Zhang, L.; Zhou, J.; George, T.S.; Feng, G. Arbuscular mycorrhizal fungi enhance mineralisation of organic phosphorus by carrying bacteria along their extraradical hyphae. New Phytol. 2020, 230, 304–315. [Google Scholar] [CrossRef]
- Ji, Y.; Yang, X.; Ji, Z.; Zhu, L.; Ma, N.; Chen, D.; Jia, X.; Tang, J.; Cao, Y. DFT-Calculated IR Spectrum Amide I, II, and III Band Contributions of N-Methylacetamide Fine Components. ACS Omega 2020, 5, 8572–8578. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, E.Z. Microbial chitinases: Properties, enhancement and potential applications. Protoplasma 2021, 258, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, J.; George, T.S.; Limpens, E.; Feng, G. Arbuscular mycorrhizal fungi conducting the hyphosphere bacterial orchestra. Trends Plant Sci. 2022, 27, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Hall, S.J.; Huang, W.; Timokhin, V.I.; Hammel, K.E. Lignin lags, leads, or limits the decomposition of litter and soil organic carbon. Ecology 2020, 101, e03113. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Jørgensen, H.B. Carbon quality rather than stoichiometry controls litter decomposition in a tropical rain forest. J. Ecol. 2010, 98, 754–763. [Google Scholar] [CrossRef]
- Christensen, O.M.; Mather, J.G. Pesticide-induced surface migration by lumbricid earthworms in grassland: Life-stage and species differences. Ecotoxicol. Environ. Saf. 2004, 57, 89–99. [Google Scholar] [CrossRef]
- Javoreková, S.; Svrceková, I.; Maková, J. Influence of benomyl and prometryn on the soil microbial activities and community structures in pasture grasslands of Slovakia. J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes 2010, 45, 702–709. [Google Scholar] [CrossRef]
- Lee, L.S.; Sassman, S.A.; Bischoff, M.; Turco, R.F. Degradation of N,N′-Dibutylurea (DBU) in Soils Treated with only DBU and DBU-Fortified Benlate Fungicides. J. Environ. Qual. 2004, 33, 1771–1778. [Google Scholar] [CrossRef]
- Ullah, M.R.; Dijkstra, F.A. Fungicide and Bactericide Effects on Carbon and Nitrogen Cycling in Soils: A Meta-Analysis. Soil Syst. 2019, 3, 23. [Google Scholar] [CrossRef]
- Schwarzer, C.; Haselwandter, K. Enzymatic degradation of the nitrification inhibitor dicyandiamide by a soil bacterium. Soil Biol. Biochem. 1991, 23, 309–310. [Google Scholar] [CrossRef]
- Kelliher, F.M.; Clough, T.J.; Clark, H.; Rys, G.; Sedcole, J.R. The temperature dependence of dicyandiamide (DCD) degradation in soils: A data synthesis. Soil Biol. Biochem. 2008, 40, 1878–1882. [Google Scholar] [CrossRef]
- Ghose, T.K. Measurement of cellulase activities. Pure Appl. Chem. 1987, 59, 257–268. [Google Scholar] [CrossRef]
- Kandeler, E.; Tscherko, D.; Spiegel, H. Long-term monitoring of microbial biomass, N mineralisation and enzyme activities of a Chernozem under different tillage management. Biol. Fertil. Soils 1999, 28, 343–351. [Google Scholar] [CrossRef]
- Liu, L.; Jin, L.; Guo, Q. Effects of Soil Microbiomes and Enzymatic Activities on Glechoma longituba. HortScience 2020, 55, 515–521. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ceccanti, B.; Cervelli, S.; Matarese, E. Extraction of Phosphatase, Urease, Proteases, Organic Carbon, and Nitrogen from Soil. Soil Sci. Soc. Am. J. 1980, 44, 1011–1016. [Google Scholar] [CrossRef]
- Urbanová, M.; Šnajdr, J.; Brabcová, V.; Merhautová, V.; Dobiášová, P.; Cajthaml, T.; Vaněk, D.; Frouz, J.; Šantrůčková, H.; Baldrian, P. Litter decomposition along a primary post-mining chronosequence. Biol. Fertil. Soils 2014, 50, 827–837. [Google Scholar] [CrossRef]
- Vepsalainen, M.; Kukkonen, S.; Vestberg, M.; Sirvio, H.; Niemi, R.M. Application of soil enzyme activity test kit in a field experiment. Soil Biol. Biochem. 2001, 33, 1665–1672. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Between Subjects | Within Subjects | |||||
|---|---|---|---|---|---|---|---|
| B | DCD | B * DCD | T | DCD * T | B * T | B * DCD * T | |
| Swc | 1.313 | 0.082 | 0.049 | 33.005 *** | 0.327 | 0.572 | 0.130 |
| pH | 3.816 * | 10.966 | 1.995 | 23.685 *** | 22.234 | 0.395 | 0.054 |
| NO3− | 0.307 | 3.760 | 0.001 | 112.154 *** | 3.860 * | 3.907 * | 0.600 |
| NH4+ | 3.681 | 386.110 *** | 27.370 *** | 43.989 *** | 29.643 *** | 3.686 * | 3.400 * |
| Rnit | 31.070 *** | 29.491 *** | 1.563 | 115.186 *** | 32.921 *** | 4.951 ** | 0.616 |
| Ramm | 1.561 | 72.660 *** | 18.080 ** | 129.548 *** | 18.875 *** | 4.703 ** | 4.311 ** |
| Rmin | 0.997 | 10.315 * | 7.789 * | 53.186 *** | 5.076 ** | 11.271 *** | 2.793 |
| Ure | 0.056 | 0.4031 | 1.093 | 227.635 *** | 1.099 | 0.259 | 1.251 |
| Pro | 3.891 | 1.197 | 14.48 | 55.695 *** | 0.987 | 2.025 | 1.740 |
| Inv | 1.105 | 4.104 | 1.617 | 26.611 *** | 2.156 | 0.394 | 2.482 |
| NAG | 7.864 * | 0.042 | 0.747 | 4.161 * | 0.313 | 0.841 | 2.102 |
| BG | 3.347 | 5.807 * | 0.598 | 14.738 *** | 11.735 *** | 5.922 ** | 1.162 |
| Cx | 1.602 | 3.133 | 0.610 | 22.081 *** | 0.866 | 0.183 | 2.382 |
| ACP | 0.750 | 22.400 ** | 6.138 * | 372.874 *** | 13.051 *** | 1.619 | 4.178 * |
| ALP | 7.081 * | 7.221 * | 0.996 | 315.834 *** | 0.675 | 3.496 * | 1.637 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Long, X.; Liao, Y.; Lin, Y.; He, Z.; Kong, Q.; Kong, X.; He, X. Influence of Arbuscular Mycorrhizal Fungi on Nitrogen Dynamics During Cinnamomum camphora Litter Decomposition. Microorganisms 2025, 13, 151. https://doi.org/10.3390/microorganisms13010151
Gao Y, Long X, Liao Y, Lin Y, He Z, Kong Q, Kong X, He X. Influence of Arbuscular Mycorrhizal Fungi on Nitrogen Dynamics During Cinnamomum camphora Litter Decomposition. Microorganisms. 2025; 13(1):151. https://doi.org/10.3390/microorganisms13010151
Chicago/Turabian StyleGao, Yuehong, Xiaoyu Long, Yiqi Liao, Yonghui Lin, Zaihua He, Qin Kong, Xiangshi Kong, and Xingbing He. 2025. "Influence of Arbuscular Mycorrhizal Fungi on Nitrogen Dynamics During Cinnamomum camphora Litter Decomposition" Microorganisms 13, no. 1: 151. https://doi.org/10.3390/microorganisms13010151
APA StyleGao, Y., Long, X., Liao, Y., Lin, Y., He, Z., Kong, Q., Kong, X., & He, X. (2025). Influence of Arbuscular Mycorrhizal Fungi on Nitrogen Dynamics During Cinnamomum camphora Litter Decomposition. Microorganisms, 13(1), 151. https://doi.org/10.3390/microorganisms13010151