
Crown Gall Induced by a Natural Isolate of Brucella (Ochrobactrum) pseudogrignonense Containing a Tumor-Inducing Plasmid

Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Culture, DNA Isolation, and Virulence Test
2.2. Sequencing Methods
2.3. Data Processing Methods
2.4. Data Availability
3. Results
3.1. Crown Gall-Inducing Strain NBC51/LBA8980
3.2. Assembly of a High Quality, Complete NBC51/LBA8980 Genome
3.3. NBC51/LBA8980 Belongs to the Species Brucella (Ochrobactrum) pseudogrignonense
3.4. Functions Encoded by the Chromosomes and the Plasmids
3.5. Ti Plasmid pTi151
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dessaux, Y.; Petit, A.; Tempé, J. Chemistry and biochemistry of opines, chemical mediators of parasitism. Phytochemistry 1993, 34, 31–38. [Google Scholar] [CrossRef]
- Bevan, M.W.; Chilton, M.D. T-DNA of the Agrobacterium Ti and Ri plasmids. Ann. Rev. Genet. 1982, 16, 357–384. [Google Scholar] [CrossRef] [PubMed]
- Van Montagu, M.; Schell, J. The Ti plasmids of Agrobacterium. Curr. Top. Microbiol. Immunol. 1982, 96, 237–254. [Google Scholar] [PubMed]
- Zhu, J.; Oger, P.M.; Schrammeijer, B.; Hooykaas, P.J.; Farrand, S.K.; Winans, S.C. The bases of crown gall tumorigenesis. J. Bacteriol. 2000, 182, 3885–3895. [Google Scholar] [CrossRef]
- Gelvin, S.B. Agrobacterium-mediated plant transformation: The biology behind the “gene-jockeying” tool. Microbiol. Mol. Biol. Rev. 2003, 67, 16–37. [Google Scholar] [CrossRef]
- Lacroix, B.; Citovsky, V. Pathways of DNA transfer to plants from Agrobacterium tumefaciens and related bacterial species. Ann. Rev. Phytopathol. 2019, 57, 231–251. [Google Scholar] [CrossRef]
- Weisberg, A.J.; Wu, Y.; Chang, J.H.; Lai, E.M.; Kuo, C.H. Virulence and ecology of Agrobacteria in the context of evolutionary genomics. Ann. Rev. Phytopathol. 2023, 61, 1–23. [Google Scholar] [CrossRef]
- Brown, P.J.B.; Chang, J.H.; Fuqua, C. Agrobacterium tumefaciens: A transformative agent for fundamental insights into host-microbe interactions. J. Bacteriol. 2023, 205, e0000523. [Google Scholar] [CrossRef]
- Hooykaas, P.J.J. The Ti plasmid, driver of Agrobacterium pathogenesis. Phytopathology 2023, 113, 594–604. [Google Scholar] [CrossRef]
- Nabi, N.; Ben Hafsa, A.; Gaillard, V.; Nesme, X.; Chaouachi, M.; Vial, L. Evolutionary classification of tumor- and root-inducing plasmids based on T-DNAs and virulence regions. Mol. Phylogenet. Evol. 2022, 169, 107388. [Google Scholar] [CrossRef]
- Kerr, A.; Panagopoulos, C.G. Biotypes of Agrobacterium radiobacter var. tumefaciens and their biological control. Phytopath. Z. 1977, 90, 172–179. [Google Scholar]
- Mousavi, S.A.; Willems, A.; Nesme, X.; de Lajudie, P.; Lindstrom, K. Revised phylogeny of Rhizobiaceae: Proposal of the delineation of Pararhizobium gen. nov., and 13 new species combinations. Syst. Appl. Microbiol. 2015, 38, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Cantalapiedra, C.P.; Hernández-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. eggNOG-mapper v2, Functional annotation, orthology assignments, and domain prediction at the metagenomic scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R.; Glöckner, F.O.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Sardà Carbasse, J.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef]
- Harrison, P.W.; Lower, R.P.; Kim, N.K.; Young, J.P. Introducing the bacterial ‘chromid’: Not a chromosome, not a plasmid. Trends Microbiol. 2010, 18, 141–148. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef]
- Lang, A.S.; Zhaxybayeva, O.; Beatty, J.T. Gene transfer agents: Phage-like elements of genetic exchange. Nature reviews. Microbiology 2012, 10, 472–482. [Google Scholar] [PubMed]
- Poret-Peterson, A.T.; Bhatnagar, S.; McClean, A.E.; Kluepfel, D.A. Draft genome sequence of Agrobacterium tumefaciens biovar 1 strain 186, Isolated from Walnut. Genome Announc. 2017, 5, e01232-17. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.F.; Santos, M.N.M.; Cho, S.T.; Chang, H.H.; Tsai, Y.M.; Smith, D.A.; Kuo, C.H.; Chang, J.H.; Lai, E.M. Plant-pathogenic Agrobacterium tumefaciens strains have diverse Type VI effector-immunity pairs and vary in in-planta competitiveness. Mol. Plant-Microbe Interact. 2019, 32, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.; van Heusden, G.P.H.; Hooykaas, P.J.J. Complete sequence of succinamopine Ti-plasmid pTiEU6 reveals its evolutionary relatedness with nopaline-type Ti-plasmids. Genome Biol. Evol. 2019, 11, 2480–2491. [Google Scholar] [CrossRef]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef]
- Haryono, M.; Tsai, Y.M.; Lin, C.T.; Huang, F.C.; Ye, Y.C.; Deng, W.L.; Hwang, H.H.; Kuo, C.H. Presence of an Agrobacterium-Type Tumor-Inducing plasmid in Neorhizobium sp. NCHU2750 and the link to phytopathogenicity. Genome Biol. Evol. 2018, 10, 3188–3195. [Google Scholar] [CrossRef]
- diCenzo, G.C.; Yang, Y.; Young, J.P.W.; Kuzmanović, N. Refining the taxonomy of the order Hyphomicrobiales (Rhizobiales) based on whole genome comparisons of over 130 type strains. Int. J. System. Evol. Microbiol. 2024, 74, 006328. [Google Scholar] [CrossRef]
- Hooykaas, P.J.J.; Klapwijk, P.M.; Nuti, M.P.; Schilperoort, R.A.; Rorsch, A. Transfer of the Agrobacterium tumefaciens TI plasmid to avirulent Agrobacteria and to Rhizobium ex planta. J. Gen. Microbiol. 1977, 98, 477–484. [Google Scholar] [CrossRef]
- van Veen, R.J.M.; den Dulk-Ras, H.; Bisseling, T.; Schilperoort, R.A.; Hooykaas, P.J.J. Crown gall tumor and root nodule formation by the bacterium Phyllobacterium myrsinacearum after the introduction of an Agrobacterium Ti plasmid or a Rhizobium Sym plasmid. Mol. Plant Microbe Interact. 1988, 1, 231–234. [Google Scholar] [CrossRef]
- Teyssier-Cuvelle, S.; Oger, P.; Mougel, C.; Groud, K.; Farrand, S.K.; Nesme, X. A highly selectable and highly transferable Ti plasmid to study conjugal host range and Ti plasmid dissemination in complex ecosystems. Microb. Ecol. 2004, 48, 10–18. [Google Scholar] [CrossRef]
- Holsters, M.; Silva, B.; Van Vliet, F.; Hernalsteens, J.P.; Genetello, C.; Van Montagu, M.; Schell, J. In vivo transfer of the ti-plasmid of Agrobacterium tumefaciens to Escherichia coli. Mol. Gen. Genet. 1978, 163, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Weller, S.A.; Stead, D.E.; Young, J.P. Acquisition of an Agrobacterium Ri plasmid and pathogenicity by other alpha-Proteobacteria in cucumber and tomato crops affected by root mat. Appl. Environ. Microbiol. 2004, 70, 2779–2785. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Peng, W.; He, X.; Wang, B.; Gan, B.; Zhang, X. Mushroom tumor: A new disease on Flammulina velutipes caused by Ochrobactrum pseudogrignonense. FEMS Microbiol. Lett. 2016, 363, fnv226. [Google Scholar] [CrossRef] [PubMed]
- van Veen, R.J.M.; den Dulk-Ras, H.; Schilperoort, R.A.; Hooykaas, P.J.J. Ti plasmid containing Rhizobium meliloti are non-tumorigenic on plants, despite proper virulence gene induction and T-strand formation. Arch. Microbiol. 1989, 153, 85–89. [Google Scholar] [CrossRef]
- Hooykaas, M.J.G.; Hooykaas, P.J.J. Complete genomic sequence and phylogenomics analysis of Agrobacterium strain AB2/73, a new Rhizobium species with a unique mega-Ti plasmid. BMC Microbiol. 2021, 21, 295. [Google Scholar] [CrossRef]
- Kuzmanović, N.; diCenzo, G.C.; Bunk, B.; Spröer, C.; Frühling, A.; Neumann-Schaal, M.; Overmann, J.; Smalla, K. Genomics of the “tumorigenes” clade of the family Rhizobiaceae and description of Rhizobium rhododendri sp. nov. MicrobiologyOpen 2023, 12, e1352. [Google Scholar] [CrossRef]
- Otten, L.; Canaday, J.; Gérard, J.C.; Fournier, P.; Crouzet, P.; Paulus, F. Evolution of agrobacteria and their Ti plasmids—A review. Mol. Plant-Microbe Interact. 1992, 5, 279–287. [Google Scholar] [CrossRef]
- Broothaerts, W.; Mitchell, H.J.; Weir, B.; Kaines, S.; Smith, L.M.; Yang, W.; Mayer, J.E.; Roa-Rodríguez, C.; Jefferson, R.A. Gene transfer to plants by diverse species of bacteria. Nature 2005, 433, 629–633. [Google Scholar] [CrossRef]
- Rudder, S.; Doohan, F.; Creevey, C.J.; Wendt, T.; Mullins, E. Genome sequence of Ensifer adhaerens OV14 provides insights into its ability as a novel vector for the genetic transformation of plant genomes. BMC Genom. 2014, 15, 268. [Google Scholar] [CrossRef]
- Cho, H.J.; Moy, Y.; Rudnick, N.A.; Klein, T.M.; Yin, J.; Bolar, J.; Hendrick, C.; Beatty, M.; Castañeda, L.; Kinney, A.J.; et al. Development of an efficient marker-free soybean transformation method using the novel bacterium Ochrobactrum haywardense H1. Plant Biotechnol. J. 2022, 20, 977–990. [Google Scholar] [CrossRef]
- Yang, Z.; Chai, Z.; Wang, X.; Zhang, Z.; Zhang, F.; Kang, F.; Liu, W.; Ren, H.; Jin, Y.; Yue, J. Comparative genomic analysis provides insights into the genetic diversity and pathogenicity of the genus Brucella. Front. Microbiol. 2024, 15, 1389859. [Google Scholar] [CrossRef] [PubMed]
- Slater, S.C.; Goldman, B.S.; Goodner, B.; Setubal, J.C.; Farrand, S.K.; Nester, E.W.; Burr, T.J.; Banta, L.; Dickerman, A.W.; Paulsen, I.; et al. Genome sequences of three Agrobacterium biovars help elucidate the evolution of multichromosome genomes in bacteria. J. Bacteriol. 2009, 191, 2501–2511. [Google Scholar] [CrossRef] [PubMed]
- Machelart, A.; Willemart, K.; Zúñiga-Ripa, A.; Godard, T.; Plovier, H.; Wittmann, C.; Moriyón, I.; De Bolle, X.; Van Schaftingen, E.; Letesson, J.J.; et al. Convergent evolution of zoonotic Brucella species toward the selective use of the pentose phosphate pathway. Proc. Natl. Acad. Sci. USA 2020, 117, 26374–26381. [Google Scholar] [CrossRef] [PubMed]
- Moreno, E.; Blasco, J.M.; Letesson, J.J.; Gorvel, J.P.; Moriyón, I. Pathogenicity and its implications in taxonomy: The Brucella and Ochrobactrum case. Pathogens 2022, 11, 377. [Google Scholar] [CrossRef]
- Moreno, E.; Middlebrook, E.A.; Altamirano-Silva, P.; Al Dahouk, S.; Araj, G.F.; Arce-Gorvel, V.; Arenas-Gamboa, Á.; Ariza, J.; Barquero-Calvo, E.; Battelli, G.; et al. If you’re not confused, you’re not paying attention: Ochrobactrum is not Brucella. J. Clin. Microbiol. 2023, 61, e0043823. [Google Scholar] [CrossRef]
- Li, S.Y.; Huang, Y.E.; Chen, J.Y.; Lai, C.H.; Mao, Y.C.; Huang, Y.T.; Liu, P.Y. Genomics of Ochrobactrum pseudogrignonense (newly named Brucella pseudogrignonensis) reveals a new blaOXA subgroup. Microb. Genom. 2021, 7, 000626. [Google Scholar]
- Alnor, D.; Frimodt-Møller, N.; Espersen, F.; Frederiksen, W. Infections with the unusual human pathogens Agrobacterium species and Ochrobactrum anthropi. Clin. Infect. Dis. 1994, 18, 914–920. [Google Scholar] [CrossRef]
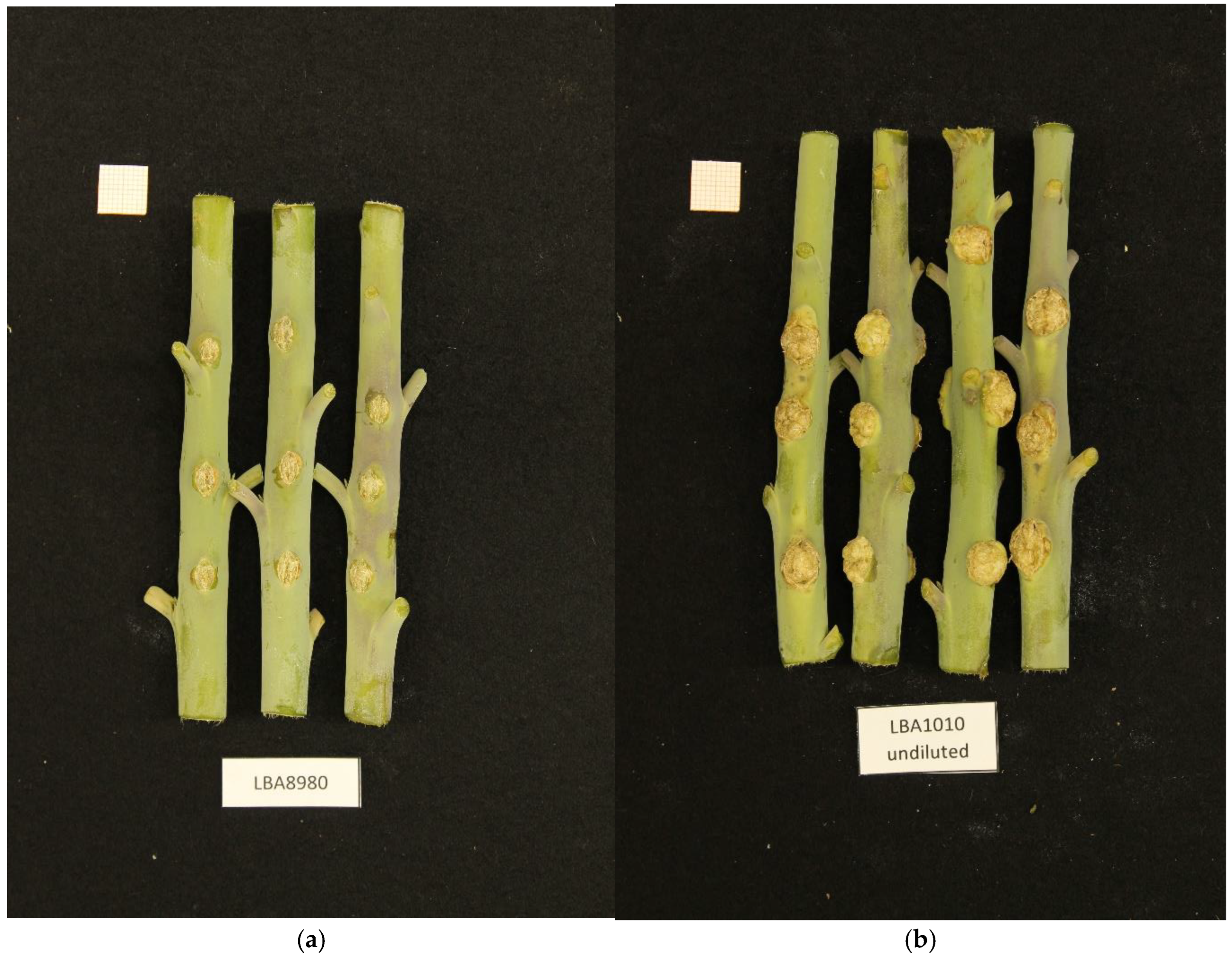
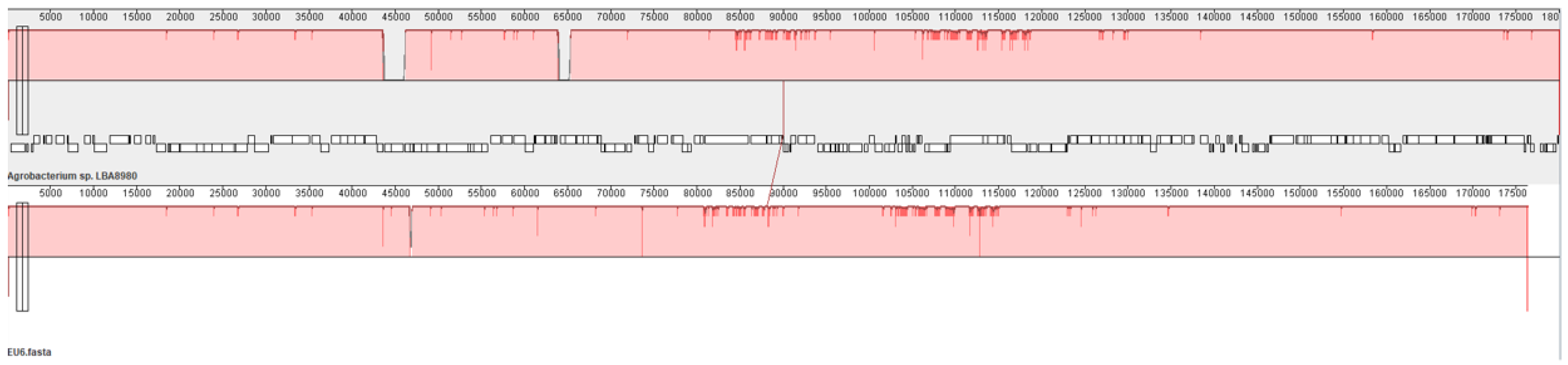

{kind=link}
{kind=link}
| ANIm [%] | Aligned [%] | Aligned [bp] | Total [bp] | |
|---|---|---|---|---|
| Ochrobactrum sp. CDB2 | 98.00 | 87.91 | 4,684,215 | 5,328,147 |
| Brucella pseudogrignonensis CCUG 30717 [T] | 97.87 | 88.24 | 4,701,446 | 5,328,147 |
| Brucella pseudogrignonensis K8 | 97.76 | 84.63 | 4,509,247 | 5,328,147 |
| Brucella pituitosa DSM 22207 [T] | 86.57 | 55.14 | 2,937,694 | 5,328,147 |
| Brucella anthropi ATCC 49188 [T] | 84.89 | 23.18 | 1,235,081 | 5,328,147 |
| Brucella intermedia LMG 3301 [T] | 84.68 | 22.27 | 1,186,669 | 5,328,147 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hooykaas, M.J.G.; Hooykaas, P.J.J. Crown Gall Induced by a Natural Isolate of Brucella (Ochrobactrum) pseudogrignonense Containing a Tumor-Inducing Plasmid. Microorganisms 2025, 13, 102. https://doi.org/10.3390/microorganisms13010102
Hooykaas MJG, Hooykaas PJJ. Crown Gall Induced by a Natural Isolate of Brucella (Ochrobactrum) pseudogrignonense Containing a Tumor-Inducing Plasmid. Microorganisms. 2025; 13(1):102. https://doi.org/10.3390/microorganisms13010102
Chicago/Turabian StyleHooykaas, Marjolein J. G., and Paul J. J. Hooykaas. 2025. "Crown Gall Induced by a Natural Isolate of Brucella (Ochrobactrum) pseudogrignonense Containing a Tumor-Inducing Plasmid" Microorganisms 13, no. 1: 102. https://doi.org/10.3390/microorganisms13010102
APA StyleHooykaas, M. J. G., & Hooykaas, P. J. J. (2025). Crown Gall Induced by a Natural Isolate of Brucella (Ochrobactrum) pseudogrignonense Containing a Tumor-Inducing Plasmid. Microorganisms, 13(1), 102. https://doi.org/10.3390/microorganisms13010102