

Veratryl Alcohol Attenuates the Virulence and Pathogenicity of Pseudomonas aeruginosa Mainly via Targeting las Quorum-Sensing System

Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Chemicals and Culture Conditions
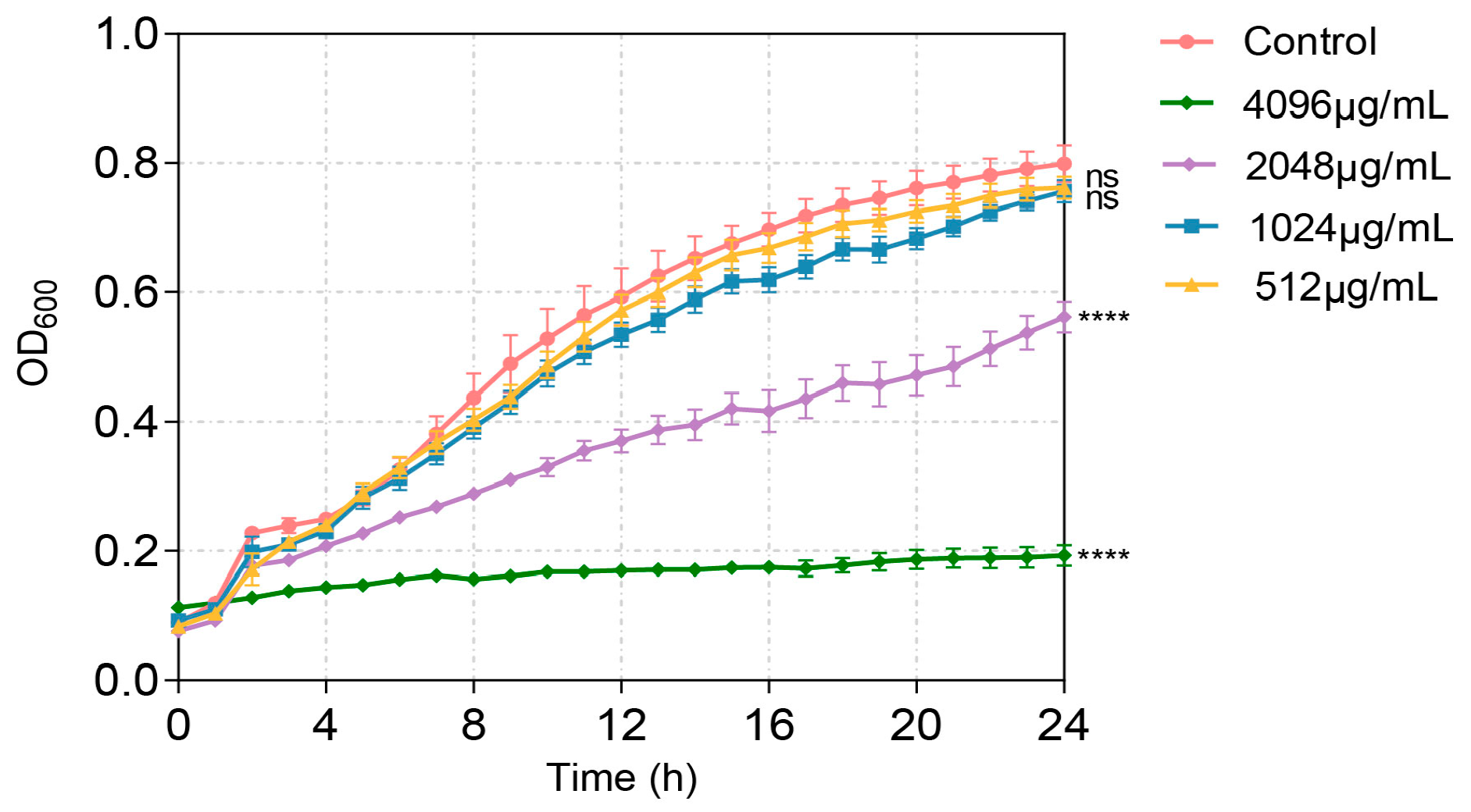
2.2. Minimum Inhibitory Concentration (MIC) Determination
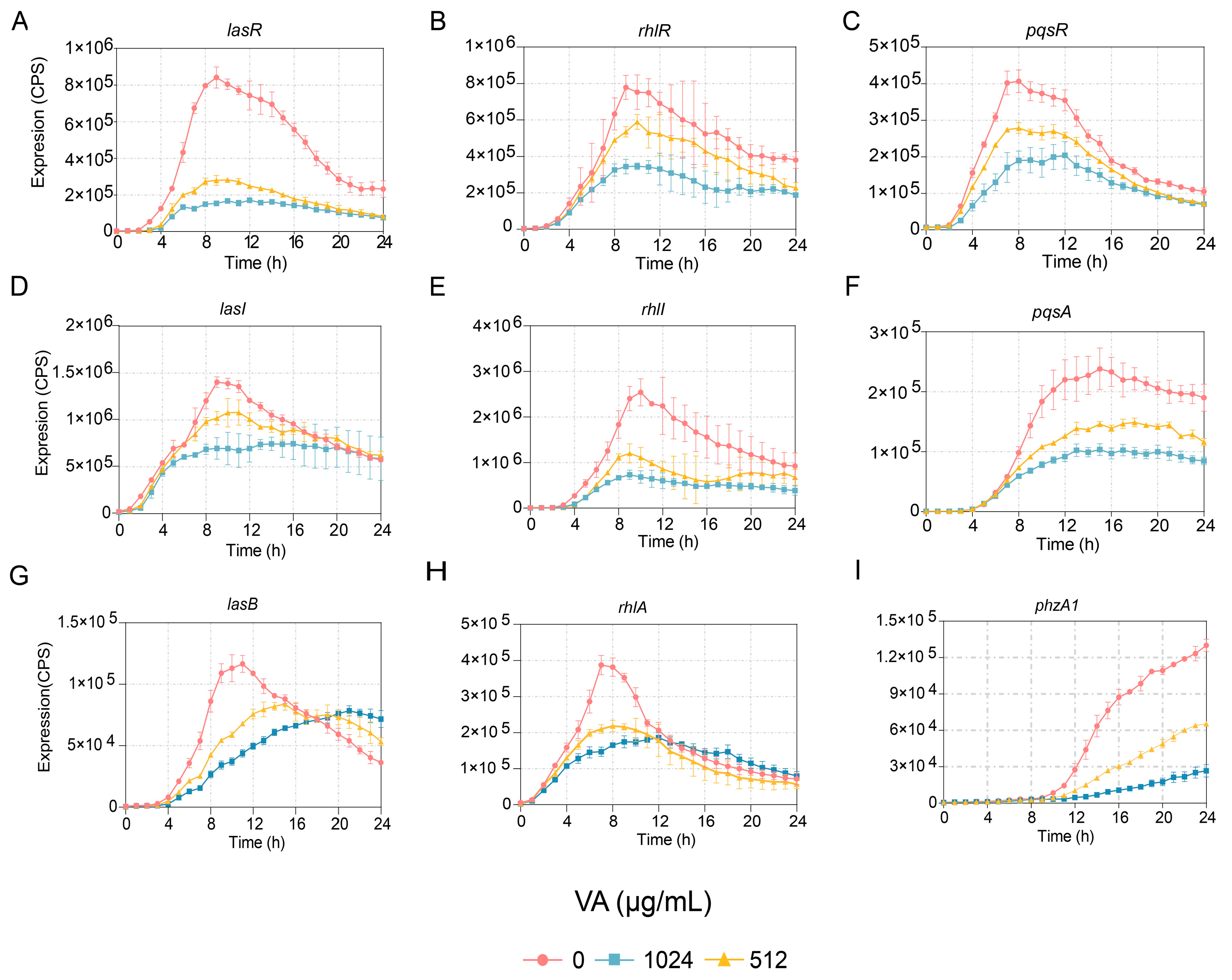
2.3. Evaluation of the Transcriptional Expression of Genes Associated with QS
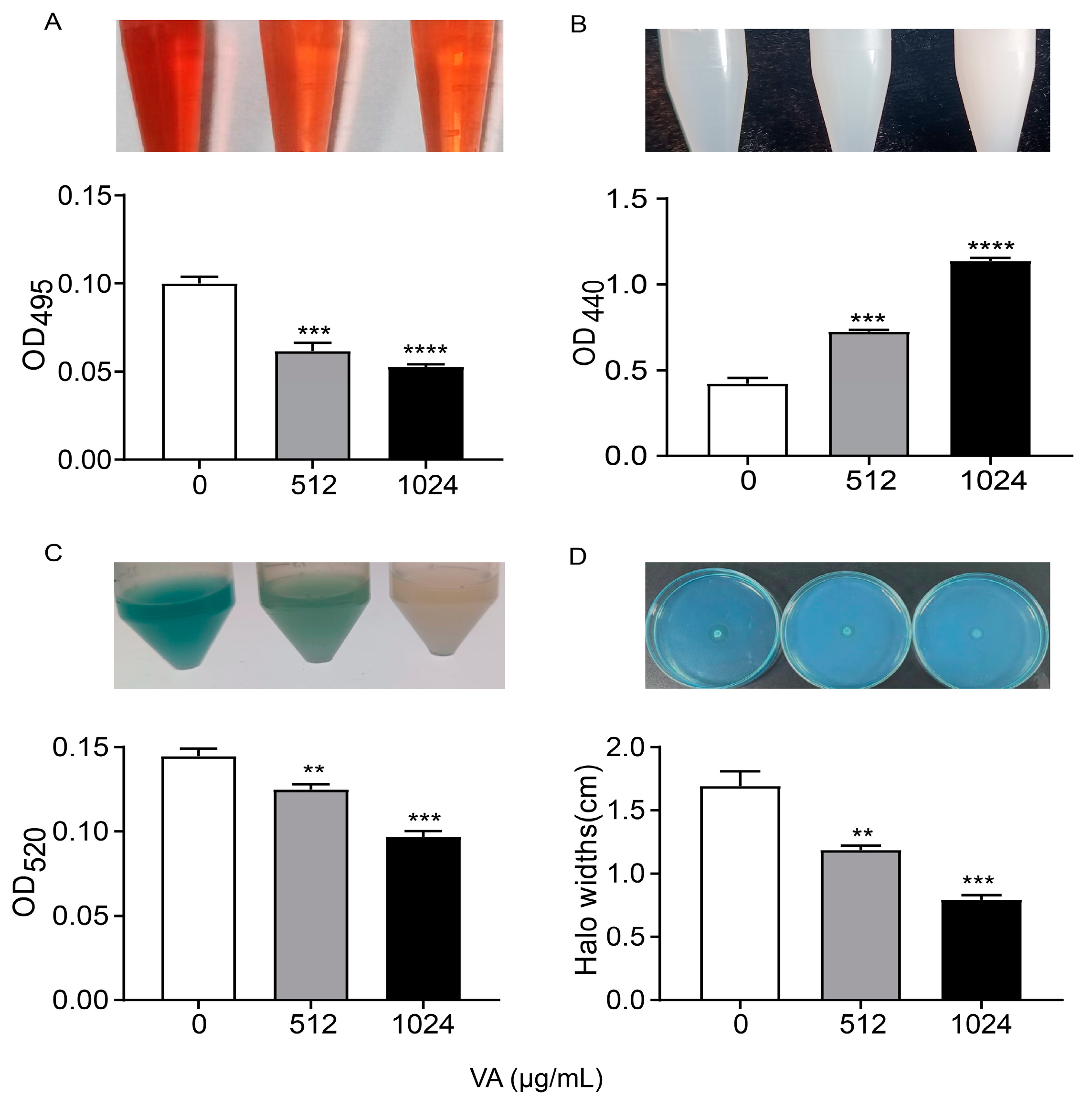
2.4. The Production of the QS-Associated Virulence Factors
2.4.1. Elastase
2.4.2. Protease
2.4.3. Pyocyanin Assay
2.4.4. Rhamnolipid
2.5. Determination of Motility
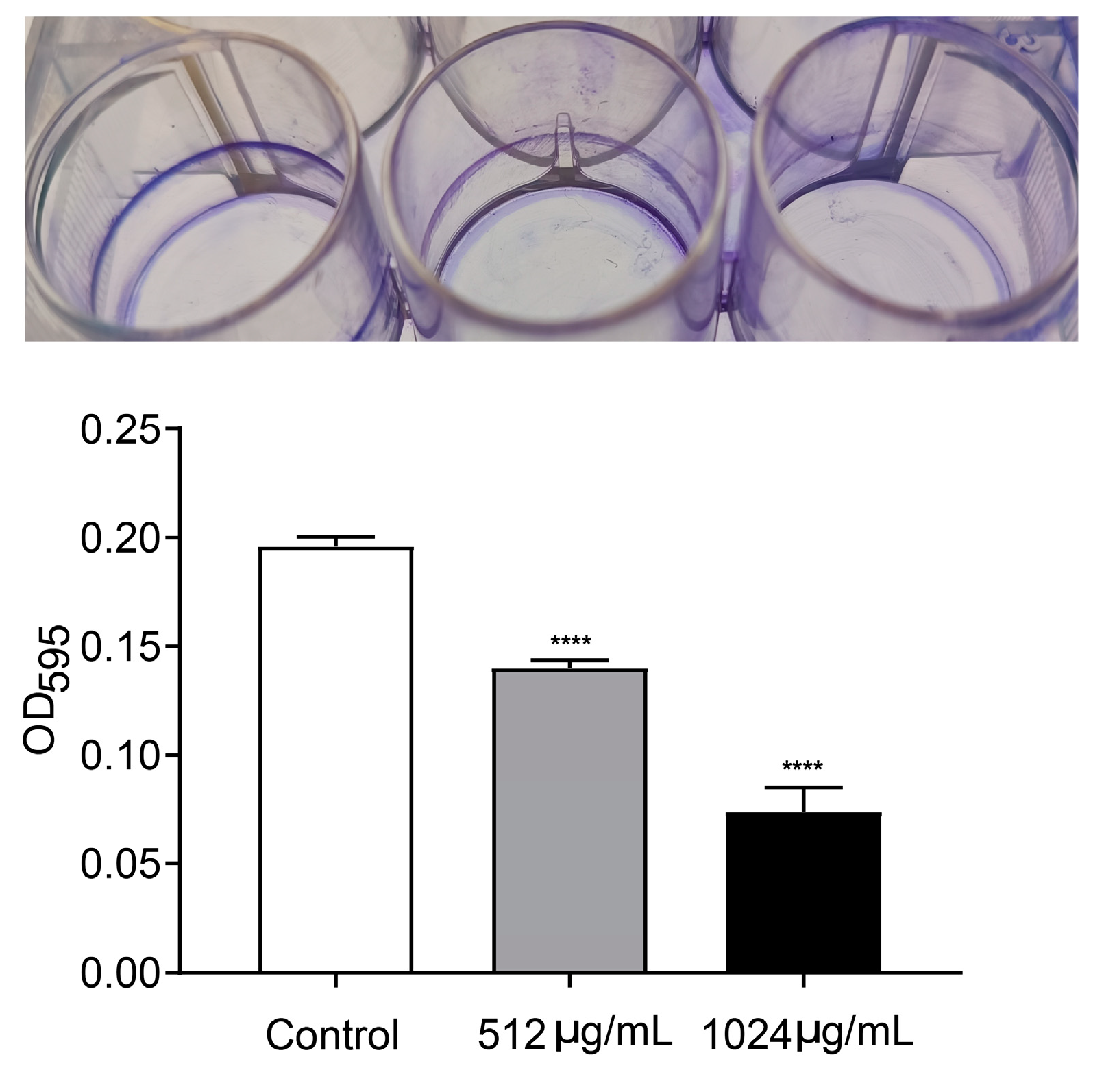
2.6. Qualification of Biofilm Formation
2.7. In Vivo Pathogenicity Assay
2.7.1. Chinese Cabbage Infection Assay
2.7.2. Drosophila melanogaster Infection Assessment
2.7.3. Caenorhabditis elegans Infection Analysis
2.8. Molecular Docking Analysis
2.9. Exploration of the Main Target of VA on QS System
2.10. Synergistic Antibacterial Effects of VA with Antibiotics
2.11. Statistical Analysis
3. Results
3.1. VA Inhibited QS-Associated Gene Expression in P. aeruginosa
3.2. VA Inhibited the Production of QS-Regulated Virulence Factors
3.3. VA Inhibited Motility and Biofilm Formation of P. aeruginosa
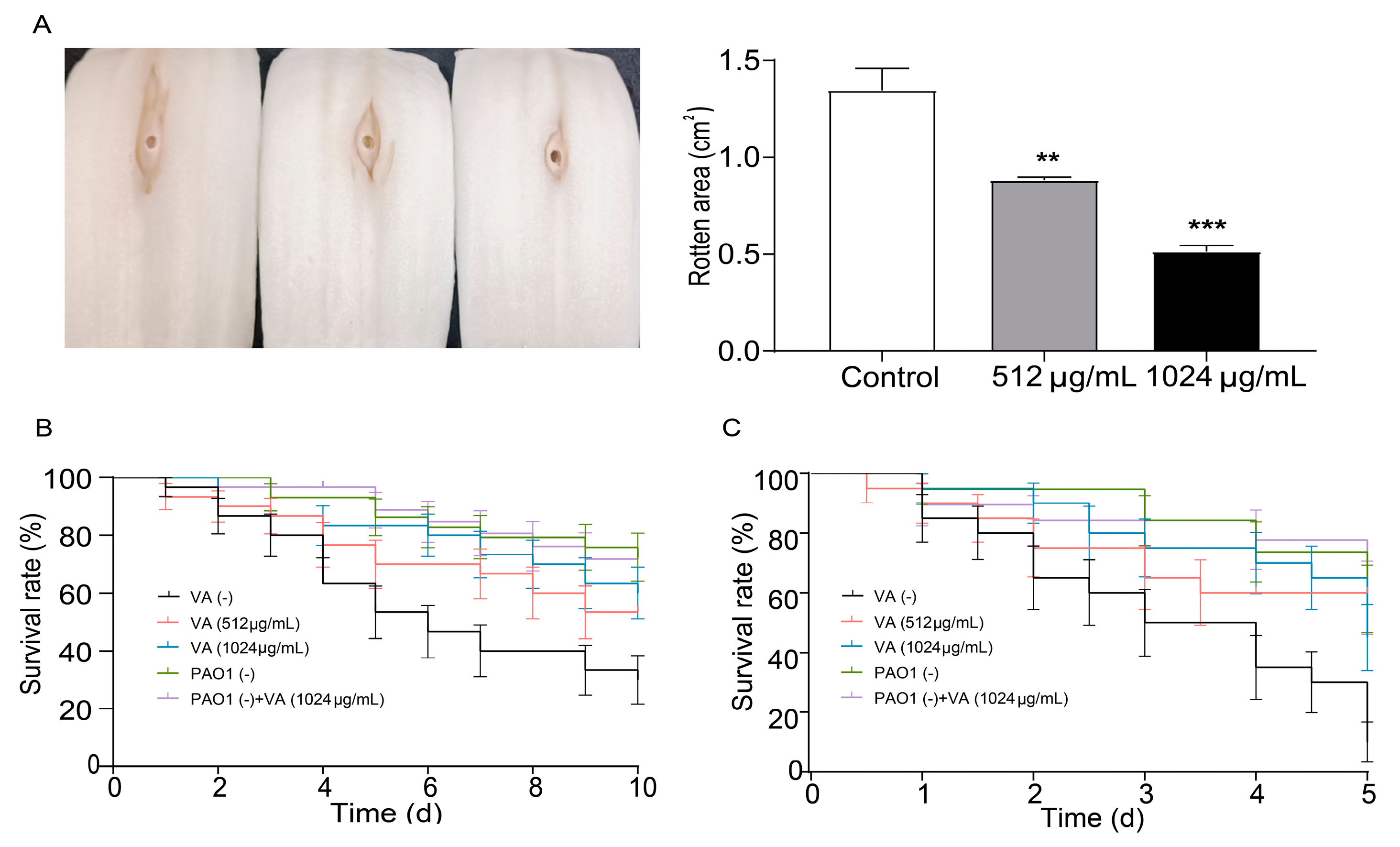
3.4. Attenuated Virulence and Pathogenicity of P. aeruginosa by Sub-Inhibitory VA
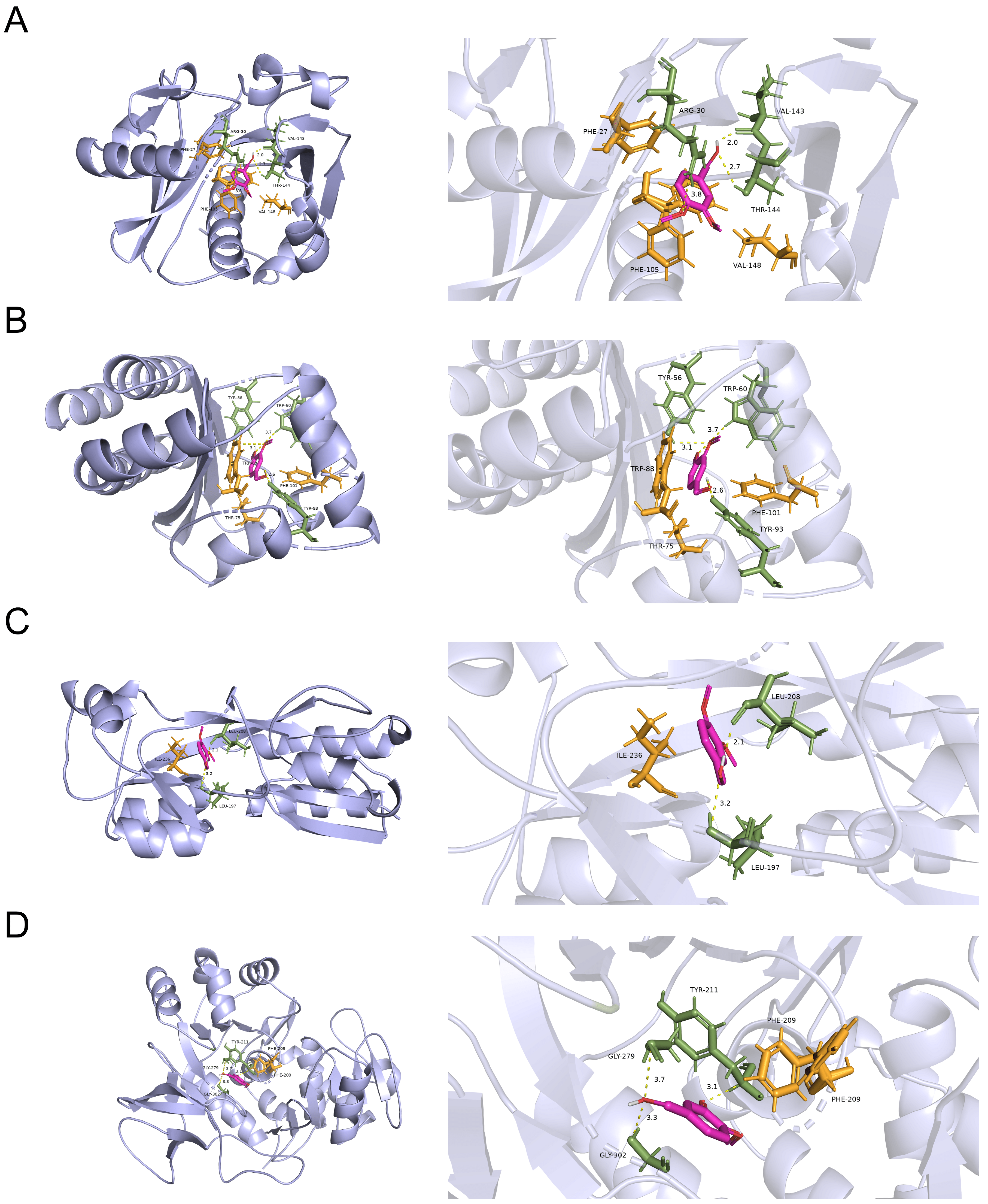
3.5. Docking Analysis for the Probable Targets of VA against the QS System of P. aeruginosa
3.6. VA Inhibits the QS System of P. aeruginosa Mainly by Targeting LasR
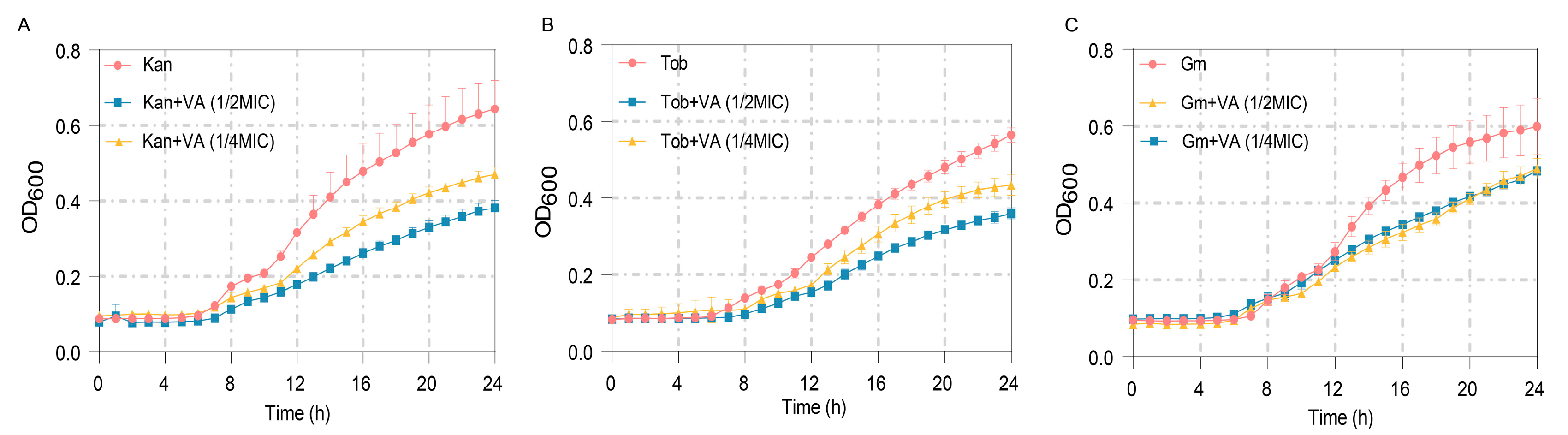
3.7. Synergistic Bacteriostatic Effects of VA and Antibiotics
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [CrossRef] [PubMed]
- de Kraker, M.E.; Stewardson, A.J.; Harbarth, S. Will 10 Million People Die a Year due to Antimicrobial Resistance by 2050? PLoS Med. 2016, 13, e1002184. [Google Scholar] [CrossRef]
- Diggle, S.P.; Whiteley, M. Microbe Profile: Pseudomonas aeruginosa: Opportunistic pathogen and lab rat. Microbiology 2020, 166, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Lamia, T.; Bousselmi, K.; Saida, B.R.; Allah, M.A. Epidemiological profile and antibiotic susceptibility of Pseudomonas aeruginosa isolates within the burned patient hospitalized in the intensive care burn unit. Tunis. Med. 2007, 85, 124–127. [Google Scholar] [PubMed]
- Pier, G.B. CFTR mutations and host susceptibility to Pseudomonas aeruginosa lung infection. Curr. Opin. Microbiol. 2002, 5, 81–86. [Google Scholar] [CrossRef]
- Nseir, S.; Ader, F.; Marquette, C.H. Nosocomial tracheobronchitis. Curr. Opin. Infect. Dis. 2009, 22, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Mendelson, M.H.; Gurtman, A.; Szabo, S.; Neibart, E.; Meyers, B.R.; Policar, M.; Cheung, T.W.; Lillienfeld, D.; Hammer, G.; Reddy, S.; et al. Pseudomonas aeruginosa bacteremia in patients with AIDS. Clin. Infect. Dis. 1994, 18, 886–895. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Hammoudi Halat, D.; Ayoub Moubareck, C. The Intriguing Carbapenemases of Pseudomonas aeruginosa: Current Status, Genetic Profile, and Global Epidemiology. Yale J. Biol. Med. 2022, 95, 507–515. [Google Scholar]
- Baraldi, E.; Lindahl, O.; Savic, M.; Findlay, D.; Årdal, C. Antibiotic Pipeline Coordinators. J. Law. Med. Ethics 2018, 46, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Liu, J.; Liu, C.; Yang, A.; Qiao, J. Quorum sensing for population-level control of bacteria and potential therapeutic applications. Cell Mol. Life Sci. 2020, 77, 1319–1343. [Google Scholar] [CrossRef] [PubMed]
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef] [PubMed]
- Chadha, J.; Harjai, K.; Chhibber, S. Revisiting the virulence hallmarks of Pseudomonas aeruginosa: A chronicle through the perspective of quorum sensing. Env. Microbiol. 2022, 24, 2630–2656. [Google Scholar] [CrossRef] [PubMed]
- Miranda, S.W.; Asfahl, K.L.; Dandekar, A.A.; Greenberg, E.P. Pseudomonas aeruginosa Quorum Sensing. Adv. Exp. Med. Biol. 2022, 1386, 95–115. [Google Scholar] [CrossRef]
- Pesci, E.C.; Milbank, J.B.; Pearson, J.P.; McKnight, S.; Kende, A.S.; Greenberg, E.P.; Iglewski, B.H. Quinolone signaling in the cell-to-cell communication system of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1999, 96, 11229–11234. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.P.; Pesci, E.C.; Iglewski, B.H. Roles of Pseudomonas aeruginosa las and rhl quorum-sensing systems in control of elastase and rhamnolipid biosynthesis genes. J. Bacteriol. 1997, 179, 5756–5767. [Google Scholar] [CrossRef]
- Gallagher, L.A.; McKnight, S.L.; Kuznetsova, M.S.; Pesci, E.C.; Manoil, C. Functions required for extracellular quinolone signaling by Pseudomonas aeruginosa. J. Bacteriol. 2002, 184, 6472–6480. [Google Scholar] [CrossRef]
- Lequette, Y.; Greenberg, E.P. Timing and localization of rhamnolipid synthesis gene expression in Pseudomonas aeruginosa biofilms. J. Bacteriol. 2005, 187, 37–44. [Google Scholar] [CrossRef]
- Zhang, Y.; Miller, R.M. Effect of a Pseudomonas rhamnolipid biosurfactant on cell hydrophobicity and biodegradation of octadecane. Appl. Environ. Microbiol. 1994, 60, 2101–2106. [Google Scholar] [CrossRef]
- Cowell, B.A.; Twining, S.S.; Hobden, J.A.; Kwong, M.S.F.; Fleiszig, S.M.J. Mutation of lasA and lasB reduces Pseudomonas aeruginosa invasion of epithelial cells. Microbiology 2003, 149, 2291–2299. [Google Scholar] [CrossRef]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef] [PubMed]
- Grandclément, C.; Tannières, M.; Moréra, S.; Dessaux, Y.; Faure, D. Quorum quenching: Role in nature and applied developments. FEMS Microbiol. Rev. 2016, 40, 86–116. [Google Scholar] [CrossRef] [PubMed]
- Vashistha, A.; Sharma, N.; Nanaji, Y.; Kumar, D.; Singh, G.; Barnwal, R.P.; Yadav, A.K. Quorum sensing inhibitors as Therapeutics: Bacterial biofilm inhibition. Bioorg. Chem. 2023, 136, 106551. [Google Scholar] [CrossRef]
- Pinar, O.; Rodríguez-Couto, S. Biologically active secondary metabolites from white-rot fungi. Front. Chem. 2024, 12, 1363354. [Google Scholar] [CrossRef]
- Khindaria, A.; Yamazaki, I.; Aust, S.D. Veratryl alcohol oxidation by lignin peroxidase. Biochemistry 1995, 34, 16860–16869. [Google Scholar] [CrossRef]
- Han, X.; Nan, M.; Cai, X.; Qiao, B.; Chen, L.; Shen, L. Sennoside A inhibits quorum sensing system to attenuate its regulated virulence and pathogenicity via targeting LasR in Pseudomonas aeruginosa. Front. Microbiol. 2022, 13, 1042214. [Google Scholar] [CrossRef] [PubMed]
- Duan, K.; Dammel, C.; Stein, J.; Rabin, H.; Surette, M.G. Modulation of Pseudomonas aeruginosa gene expression by host microflora through interspecies communication. Mol. Microbiol. 2003, 50, 1477–1491. [Google Scholar] [CrossRef]
- Duan, K.; Surette, M.G. Environmental regulation of Pseudomonas aeruginosa PAO1 Las and Rhl quorum-sensing systems. J. Bacteriol. 2007, 189, 4827–4836. [Google Scholar] [CrossRef]
- Liang, H.; Li, L.; Dong, Z.; Surette, M.G.; Duan, K. The YebC family protein PA0964 negatively regulates the Pseudomonas aeruginosa quinolone signal system and pyocyanin production. J. Bacteriol. 2008, 190, 6217–6227. [Google Scholar] [CrossRef]
- Becher, A.; Schweizer, H.P. Integration-proficient Pseudomonas aeruginosa vectors for isolation of single-copy chromosomal lacZ and lux gene fusions. Biotechniques 2000, 29, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Kessler, E.; Safrin, M. Elastinolytic and proteolytic enzymes. Methods Mol. Biol. 2014, 1149, 135–169. [Google Scholar] [CrossRef] [PubMed]
- Essar, D.W.; Eberly, L.; Hadero, A.; Crawford, I.P. Identification and characterization of genes for a second anthranilate synthase in Pseudomonas aeruginosa: Interchangeability of the two anthranilate synthases and evolutionary implications. J. Bacteriol. 1990, 172, 884–900. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, M.; Kalme, S.; Tamboli, D.; Govindwar, S. Rhamnolipid from Pseudomonas desmolyticum NCIM-2112 and its role in the degradation of Brown 3REL. J. Basic. Microbiol. 2011, 51, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.; Kornberg, A. Inorganic polyphosphate is needed for swimming, swarming, and twitching motilities of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2000, 97, 4885–4890. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Feng, Y.; Han, X.; Cai, X.; Yang, L.; Liu, C.; Shen, L. Inhibition of Virulence Factors and Biofilm Formation by Wogonin Attenuates Pathogenicity of Pseudomonas aeruginosa PAO1 via Targeting pqs Quorum-Sensing System. Int. J. Mol. Sci. 2021, 22, 12699. [Google Scholar] [CrossRef]
- Starkey, M.; Rahme, L.G. Modeling Pseudomonas aeruginosa pathogenesis in plant hosts. Nat. Protoc. 2009, 4, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Chugani, S.A.; Whiteley, M.; Lee, K.M.; D’Argenio, D.; Manoil, C.; Greenberg, E.P. QscR, a modulator of quorum-sensing signal synthesis and virulence in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2001, 98, 2752–2757. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.W.; Mahajan-Miklos, S.; Ausubel, F.M. Killing of Caenorhabditis elegans by Pseudomonas aeruginosa used to model mammalian bacterial pathogenesis. Proc. Natl. Acad. Sci. USA 1999, 96, 715–720. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Wen, F.; Wu, Y.; Yuan, Y.; Yang, X.; Ran, Q.; Gan, X.; Guo, Y.; Wang, X.; Chu, Y.; Zhao, K. Discovery of psoralen as a quorum sensing inhibitor suppresses Pseudomonas aeruginosa virulence. Appl. Microbiol. Biotechnol. 2024, 108, 222. [Google Scholar] [CrossRef] [PubMed]
- Barnes, V.L.; Heithoff, D.M.; Mahan, S.P.; House, J.K.; Mahan, M.J. Antimicrobial susceptibility testing to evaluate minimum inhibitory concentration values of clinically relevant antibiotics. STAR Protoc. 2023, 4, 102512. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, M.; Anju, C.P.; Biswas, L.; Anil Kumar, V.; Gopi Mohan, C.; Biswas, R. Antibiotic resistance in Pseudomonas aeruginosa and alternative therapeutic options. Int. J. Med. Microbiol. 2016, 306, 48–58. [Google Scholar] [CrossRef]
- Zeng, Y.X.; Liu, J.S.; Wang, Y.J.; Tang, S.; Wang, D.Y.; Deng, S.M.; Jia, A.Q. Actinomycin D: A novel Pseudomonas aeruginosa quorum sensing inhibitor from the endophyte Streptomyces cyaneochromogenes RC1. World J. Microbiol. Biotechnol. 2022, 38, 170. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Park, J.S.; Choi, H.Y.; Yoon, S.S.; Kim, W.G. Terrein is an inhibitor of quorum sensing and c-di-GMP in Pseudomonas aeruginosa: A connection between quorum sensing and c-di-GMP. Sci. Rep. 2018, 8, 8617. [Google Scholar] [CrossRef]
- Yang, W.; Wei, Q.; Tong, Q.; Cui, K.; He, G.; Lin, L.; Ma, L.Z.; Cornelis, P.; Wang, Y. Traditional Chinese Medicine Tanreqing Inhibits Quorum Sensing Systems in Pseudomonas aeruginosa. Front. Microbiol. 2020, 11, 517462. [Google Scholar] [CrossRef]
- Zhao, Z.G.; Yan, S.S.; Yu, Y.M.; Mi, N.; Zhang, L.X.; Liu, J.; Li, X.L.; Liu, F.; Xu, J.F.; Yang, W.Q.; et al. An aqueous extract of Yunnan Baiyao inhibits the quorum-sensing-related virulence of Pseudomonas aeruginosa. J. Microbiol. 2013, 51, 207–212. [Google Scholar] [CrossRef]
- Lou, Z.; Wang, H.; Tang, Y.; Chen, X. The effect of burdock leaf fraction on adhesion, biofilm formation, quorum sensing and virulence factors of Pseudomonas aeruginosa. J. Appl. Microbiol. 2017, 122, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Hao, S.; Zhao, L.; Shi, F.; Ye, G.; Zou, Y.; Song, X.; Li, L.; Yin, Z.; He, X.; et al. Paeonol Attenuates Quorum-Sensing Regulated Virulence and Biofilm Formation in Pseudomonas aeruginosa. Front. Microbiol. 2021, 12, 692474. [Google Scholar] [CrossRef]
- Benny, A.T.; Rathinam, P.; Dev, S.; Mathew, B.; Radhakrishnan, E.K. Perillaldehyde mitigates virulence factors and biofilm formation of Pseudomonas aeruginosa clinical isolates, by acting on the quorum sensing mechanism in vitro. J. Appl. Microbiol. 2022, 133, 385–399. [Google Scholar] [CrossRef]
- Zhao, C.; Zheng, H.; Zhou, L.; Ji, H.; Zhao, L.; Yu, W.; Gong, Q. Falcarindiol Isolated from Notopterygium incisum Inhibits the Quorum Sensing of Pseudomonas aeruginosa. Molecules 2021, 26, 5896. [Google Scholar] [CrossRef] [PubMed]
- Kalia, V.C.; Purohit, H.J. Quenching the quorum sensing system: Potential antibacterial drug targets. Crit. Rev. Microbiol. 2011, 37, 121–140. [Google Scholar] [CrossRef] [PubMed]
- Markus, V.; Golberg, K.; Teralı, K.; Ozer, N.; Kramarsky-Winter, E.; Marks, R.S.; Kushmaro, A. Assessing the Molecular Targets and Mode of Action of Furanone C-30 on Pseudomonas aeruginosa Quorum Sensing. Molecules 2021, 26, 1620. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Chen, J.; Wang, B.; Peng, A.Y.; Mao, Z.W.; Xia, W. Inhibition of Quorum-Sensing Regulator from Pseudomonas aeruginosa Using a Flavone Derivative. Molecules 2022, 27, 2439. [Google Scholar] [CrossRef] [PubMed]
- Boopathi, S.; Vashisth, R.; Mohanty, A.K.; Jia, A.Q.; Sivakumar, N.; Arockiaraj, J. Bacillus subtilis BR4 derived stigmatellin Y interferes Pqs-PqsR mediated quorum sensing system of Pseudomonas aeruginosa. J. Basic. Microbiol. 2022, 62, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Mok, N.; Chan, S.Y.; Liu, S.Y.; Chua, S.L. Vanillin inhibits PqsR-mediated virulence in Pseudomonas aeruginosa. Food Funct. 2020, 11, 6496–6508. [Google Scholar] [CrossRef] [PubMed]
- Kai, T.; Tateda, K.; Kimura, S.; Ishii, Y.; Ito, H.; Yoshida, H.; Kimura, T.; Yamaguchi, K. A low concentration of azithromycin inhibits the mRNA expression of N-acyl homoserine lactone synthesis enzymes, upstream of lasI or rhlI, in Pseudomonas aeruginosa. Pulm. Pharmacol. Ther. 2009, 22, 483–486. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Xu, J.L.; Hu, J.; Wang, L.H.; Ong, S.L.; Leadbetter, J.R.; Zhang, L.H. Acyl-homoserine lactone acylase from Ralstonia strain XJ12B represents a novel and potent class of quorum-quenching enzymes. Mol. Microbiol. 2003, 47, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Sunder, A.V.; Utari, P.D.; Ramasamy, S.; van Merkerk, R.; Quax, W.; Pundle, A. Penicillin V acylases from gram-negative bacteria degrade N-acylhomoserine lactones and attenuate virulence in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2017, 101, 2383–2395. [Google Scholar] [CrossRef]
- Liu, N.; Yu, M.; Zhao, Y.; Cheng, J.; An, K.; Zhang, X.H. PfmA, a novel quorum-quenching N-acylhomoserine lactone acylase from Pseudoalteromonas flavipulchra. Microbiology 2017, 163, 1389–1398. [Google Scholar] [CrossRef]
- Ren, Y.; Zhu, R.; You, X.; Li, D.; Guo, M.; Fei, B.; Liu, Y.; Yang, X.; Liu, X.; Li, Y. Quercetin: A promising virulence inhibitor of Pseudomonas aeruginosa LasB in vitro. Appl. Microbiol. Biotechnol. 2024, 108, 57. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains or Plasmids | Description | Reference |
|---|---|---|
| Strains | ||
| P. aeruginosa (PAO1) | Wild-type strain | This study |
| E. coli OP50 | E. coli uracil-auxotrophic strain | This study |
| PAO1 (ΔlasR) | LasR knocked mutant of PAO1 | [27] |
| PAO1 (ΔlasIΔrhlIΔpqsAΔrhlRΔpqsR) | PAO1 quintuple mutant with lasI, rhlI, pqsA, rhlR and pqsR knocked out | [27] |
| PAO1 (ΔlasIΔrhlIΔpqsA ΔlasRΔpqsR) | PAO1 quintuple mutant with lasI, rhlI, pqsA, lasR and pqsR knocked out | [27] |
| PAO1 (ΔlasIΔrhlIΔpqsA ΔlasRΔrhlR) | PAO1 quintuple mutant with lasI, rhlI, pqsA, lasR and rhlR knocked out | [27] |
| Plasmids | ||
| PMS402 | Expression reporter vector carrying the promoterless luxCDABE; Kanr, Tmpr | [28] |
| pKD-lasI | pMS402 containing lasI promoter region; Kanr, Tmpr | [29] |
| pKD-lasR | pMS402 containing lasR promoter region; Kanr, Tmpr | [29] |
| pKD-rhlI | pMS402 containing rhlI promoter region; Kanr, Tmpr | [29] |
| pKD-rhlR | pMS402 containing rhlR promoter region; Kanr, Tmpr | [29] |
| pKD-pqsA | pMS402 containing pqsA promoter region; Kanr, Tmpr | [30] |
| pKD-pqsR | pMS402 containing pqsR promoter region; Kanr, Tmpr | [30] |
| pKD-lasB | pMS402 containing lasB promoter region; Kanr, Tmpr | [30] |
| pKD-rhlA | pMS402 containing rhlA promoter region; Kanr, Tmpr | [30] |
| pKD-phzA1 | pMS402 containing phzA1promoter region; Kanr, Tmpr | [30] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, S.; Song, W.; Han, X.; Chen, L.; Shen, L. Veratryl Alcohol Attenuates the Virulence and Pathogenicity of Pseudomonas aeruginosa Mainly via Targeting las Quorum-Sensing System. Microorganisms 2024, 12, 985. https://doi.org/10.3390/microorganisms12050985
Fu S, Song W, Han X, Chen L, Shen L. Veratryl Alcohol Attenuates the Virulence and Pathogenicity of Pseudomonas aeruginosa Mainly via Targeting las Quorum-Sensing System. Microorganisms. 2024; 12(5):985. https://doi.org/10.3390/microorganisms12050985
Chicago/Turabian StyleFu, Songzhe, Wenxu Song, Xiaofeng Han, Lin Chen, and Lixin Shen. 2024. "Veratryl Alcohol Attenuates the Virulence and Pathogenicity of Pseudomonas aeruginosa Mainly via Targeting las Quorum-Sensing System" Microorganisms 12, no. 5: 985. https://doi.org/10.3390/microorganisms12050985
APA StyleFu, S., Song, W., Han, X., Chen, L., & Shen, L. (2024). Veratryl Alcohol Attenuates the Virulence and Pathogenicity of Pseudomonas aeruginosa Mainly via Targeting las Quorum-Sensing System. Microorganisms, 12(5), 985. https://doi.org/10.3390/microorganisms12050985