


An Exploratory Review of the Potential of Lytic Proteins and Bacteriophages for the Treatment of Tuberculosis
 and
and
Abstract
1. Introduction
2. Tuberculosis (TB), Mycobacterium tuberculosis (Mtb), and the Rise of Multidrug-Resistant TB
3. Available Treatment Options for Tuberculosis
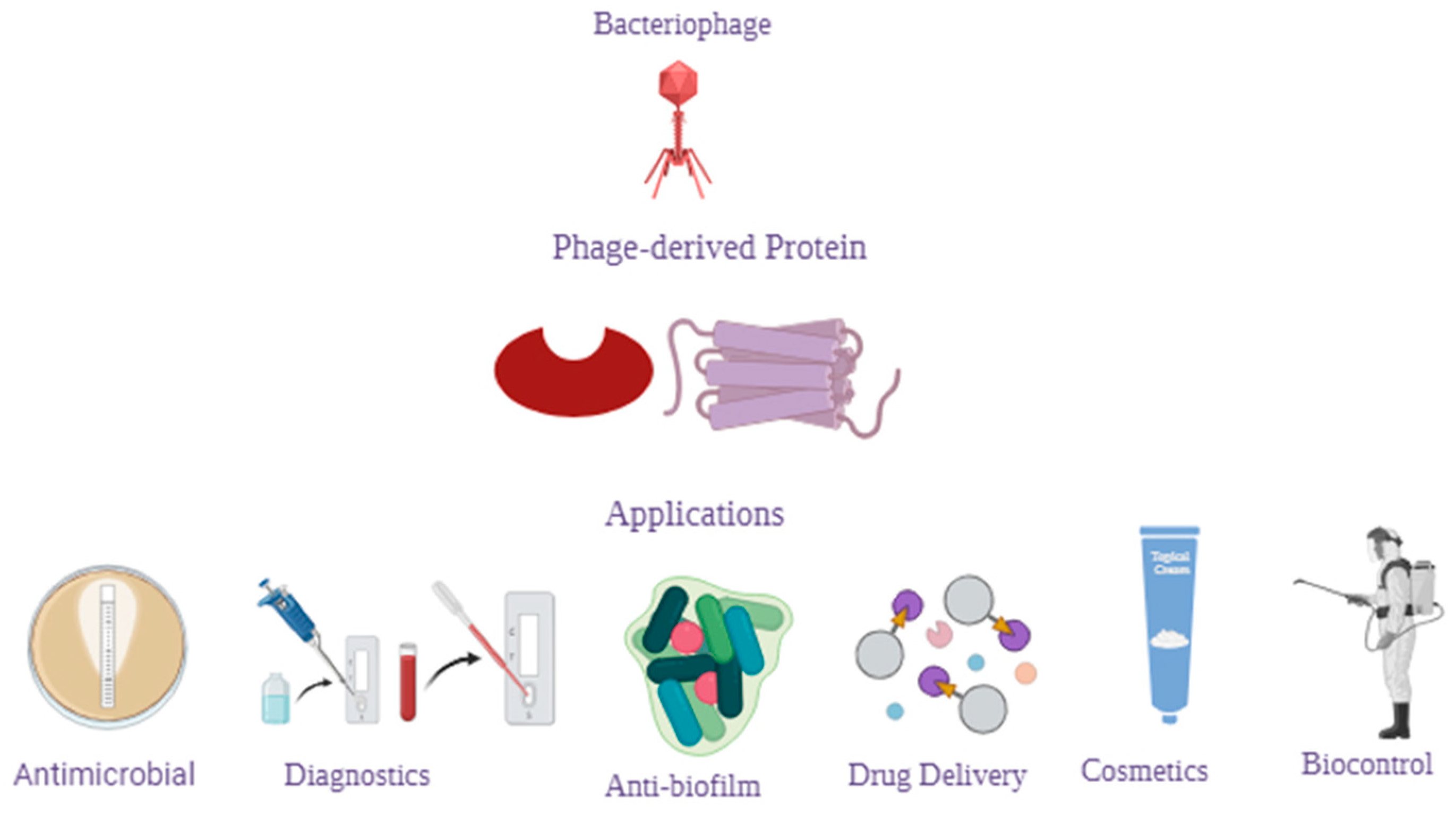
4. Bacteriophages, Their Derivatives, and Their Therapeutic Potential
4.1. Phage-Derived Proteins
4.1.1. Phage-Lytic Proteins
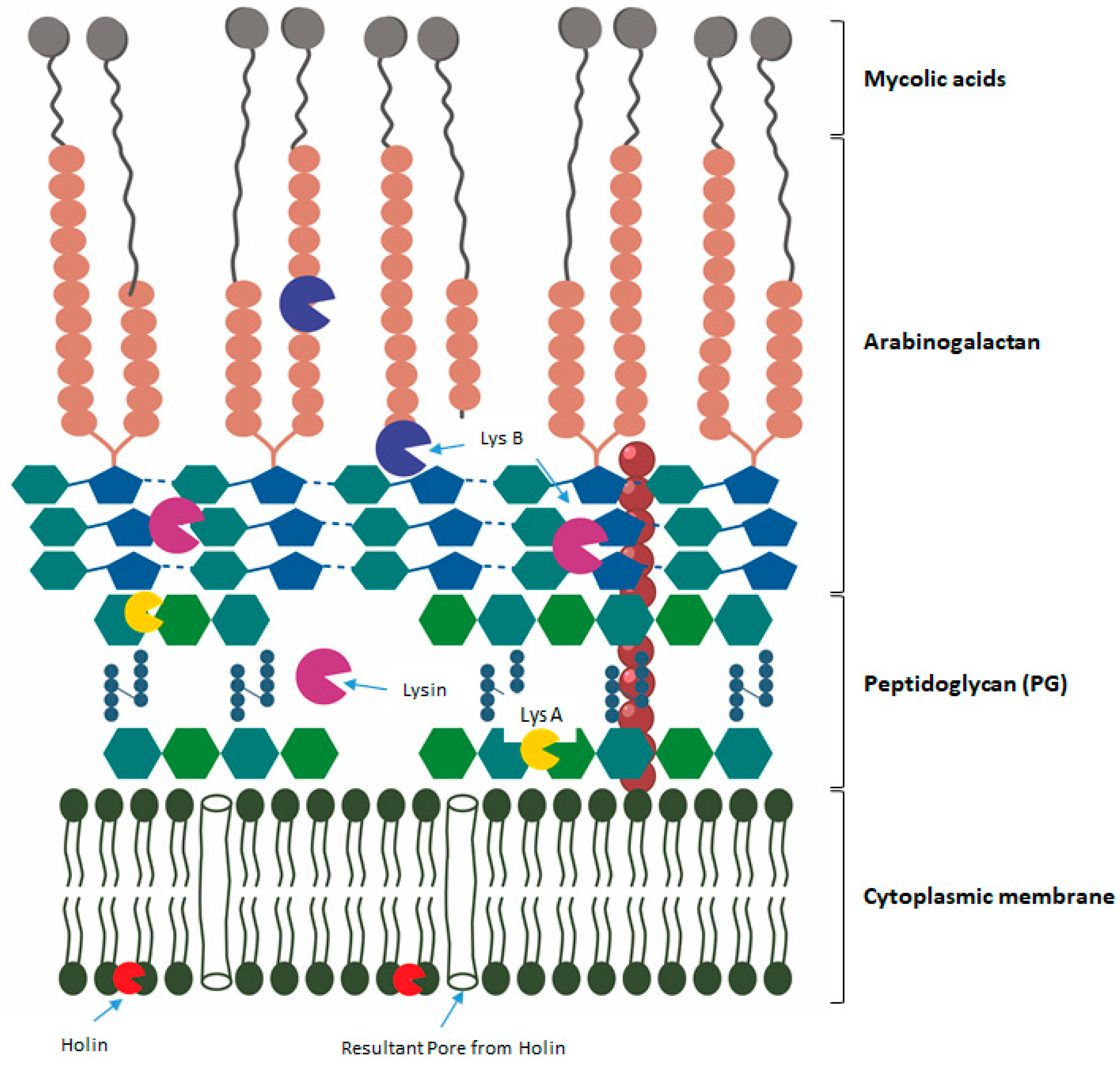
Endolysin-Peptidoglycan Degrading Enzymes
Holins-Cytoplasmic Membrane Degrading Enzymes
VAPGH—Virion-Associated Peptidoglycan Hydrolases
4.1.2. Therapeutic Potential of Phage-Lytic Proteins and Combination Therapy to Treat TB
5. Conclusions and Future Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance. In Virulence Mechanisms of Bacterial Pathogens; Wiley: Hoboken, NJ, USA, 2016; pp. 481–511. [Google Scholar]
- Reygaert, W.C. An overview of the antimicrobial resistance mechanisms of bacteria. AIMS Microbiol. 2018, 4, 482. [Google Scholar] [CrossRef] [PubMed]
- Briers, Y. Phage lytic enzymes. Viruses 2019, 11, 113. [Google Scholar] [CrossRef]
- Yang, H.; Wang, M.; Yu, J.; Wei, H. Antibacterial activity of a novel peptide-modified lysin against Acinetobacter baumannii and Pseudomonas aeruginosa. Front. Microbiol. 2015, 6, 1471. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.-J.; Liu, C.-C.; Jiang, S.-J.; Soo, P.-C.; Tu, M.-H.; Lee, J.-J.; Chen, Y.-H.; Chang, K.-C. Antimycobacterial activities of endolysins derived from a mycobacteriophage, BTCU-1. Molecules 2015, 20, 19277–19290. [Google Scholar] [CrossRef] [PubMed]
- Vasava, M.S.; Bhoi, M.N.; Rathwa, S.K.; Borad, M.A.; Nair, S.G.; Patel, H.D. Drug development against tuberculosis: Past, present and future. Indian J. Tuberc. 2017, 64, 252–275. [Google Scholar] [CrossRef]
- Catalão, M.J.; Pimentel, M. Mycobacteriophage lysis enzymes: Targeting the mycobacterial cell envelope. Viruses 2018, 10, 428. [Google Scholar] [CrossRef]
- Abouhmad, A.; Korany, A.H.; Grey, C.; Dishisha, T.; Hatti-Kaul, R. Exploring the enzymatic and antibacterial activities of novel mycobacteriophage lysin B enzymes. Int. J. Mol. Sci. 2020, 21, 3176. [Google Scholar] [CrossRef]
- Horsburgh, C.R., Jr.; Barry, C.E., III; Lange, C. Treatment of tuberculosis. N. Engl. J. Med. 2015, 373, 2149–2160. [Google Scholar] [CrossRef]
- Davies, C.G.; Reilly, K.; Altermann, E.; Hendrickson, H.L. PLAN-M.; Mycobacteriophage endolysins fused to biodegradable nanobeads mitigate mycobacterial growth in liquid and on surfaces. Front. Microbiol. 2021, 12, 562748. [Google Scholar] [CrossRef]
- World Health Organization. The End TB Strategy. Global Strategy and Targets for Tuberculosis Prevention, Care and Control after 2015; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Lange, C.; Aarnoutse, R.; Alffenaar, J.; Bothamley, G.; Brinkmann, F.; Costa, J.; Chesov, D.; Van Crevel, R.; Dedicoat, M.; Dominguez, J. Management of patients with multidrug-resistant tuberculosis. Int. J. Tuberc. Lung Dis. 2019, 23, 645–662. [Google Scholar] [CrossRef]
- Kendall, E.A.; Azman, A.S.; Cobelens, F.G.; Dowdy, D.W. MDR-TB treatment as prevention: The projected population-level impact of expanded treatment for multidrug-resistant tuberculosis. PLoS ONE 2017, 12, e0172748. [Google Scholar] [CrossRef]
- Nyang’wa, B.-T.; Berry, C.; Kazounis, E.; Motta, I.; Parpieva, N.; Tigay, Z.; Solodovnikova, V.; Liverko, I.; Moodliar, R.; Dodd, M. A 24-week, all-oral regimen for rifampin-resistant tuberculosis. N. Engl. J. Med. 2022, 387, 2331–2343. [Google Scholar] [CrossRef]
- Berry, C.; du Cros, P.; Fielding, K.; Gajewski, S.; Kazounis, E.; McHugh, T.D.; Merle, C.; Motta, I.; Moore, D.A.; Nyang’wa, B.-T. TB-PRACTECAL: Study protocol for a randomised, controlled, open-label, phase II–III trial to evaluate the safety and efficacy of regimens containing bedaquiline and pretomanid for the treatment of adult patients with pulmonary multidrug-resistant tuberculosis. Trials 2022, 23, 484. [Google Scholar]
- Wallis, R.S. Sputum culture conversion in new TB regimens. Lancet Respir. Med. 2015, 3, e18–e19. [Google Scholar] [CrossRef]
- Wallis, R.S.; Maeurer, M.; Mwaba, P.; Chakaya, J.; Rustomjee, R.; Migliori, G.B.; Marais, B.; Schito, M.; Churchyard, G.; Swaminathan, S. Tuberculosis—Advances in development of new drugs, treatment regimens, host-directed therapies, and biomarkers. Lancet Infect. Dis. 2016, 16, e34–e46. [Google Scholar] [CrossRef]
- Uplekar, M.; Weil, D.; Lonnroth, K.; Jaramillo, E.; Lienhardt, C.; Dias, H.M.; Falzon, D.; Floyd, K.; Gargioni, G.; Getahun, H. WHO’s new end TB strategy. Lancet 2015, 385, 1799–1801. [Google Scholar] [CrossRef]
- Bastos, M.L.; Lan, Z.; Menzies, D. An updated systematic review and meta-analysis for treatment of multidrug-resistant tuberculosis. Eur. Respir. J. 2017, 49, 1600803. [Google Scholar] [CrossRef] [PubMed]
- Paterson, D.L. “Collateral damage” from cephalosporin or quinolone antibiotic therapy. Clin. Infect. Dis. 2004, 38, S341–S345. [Google Scholar] [CrossRef] [PubMed]
- Weerasuriya, C.K.; Harris, R.C.; McQuaid, C.F.; Bozzani, F.; Ruan, Y.; Li, R.; Li, T.; Rade, K.; Rao, R.; Ginsberg, A.M. The epidemiologic impact and cost-effectiveness of new tuberculosis vaccines on multidrug-resistant tuberculosis in India and China. BMC Med. 2021, 19, 60. [Google Scholar] [CrossRef] [PubMed]
- Laurenzi, M.; Ginsberg, A.; Spigelman, M. Challenges associated with current and future TB treatment. Infect. Disord.-Drug Targets (Former. Curr. Drug Targets-Infect. Disord.) 2007, 7, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Dean, G.L.; Edwards, S.G.; Ives, N.J.; Matthews, G.; Fox, E.F.; Navaratne, L.; Fisher, M.; Taylor, G.P.; Miller, R.; Taylor, C.B. Treatment of tuberculosis in HIV-infected persons in the era of highly active antiretroviral therapy. Aids 2002, 16, 75–83. [Google Scholar] [CrossRef]
- Ramappa, V.; Aithal, G.P. Hepatotoxicity related to anti-tuberculosis drugs: Mechanisms and management. J. Clin. Exp. Hepatol. 2013, 3, 37–49. [Google Scholar] [CrossRef]
- Farazi, A.; Sofian, M.; Jabbariasl, M.; Keshavarz, S. Adverse reactions to antituberculosis drugs in Iranian tuberculosis patients. Tuberc. Res. Treat. 2014, 201, 412893. [Google Scholar] [CrossRef]
- Ginsberg, A.M. Drugs in development for tuberculosis. Drugs 2010, 70, 2201–2214. [Google Scholar] [CrossRef]
- Tiberi, S.; du Plessis, N.; Walzl, G.; Vjecha, M.J.; Rao, M.; Ntoumi, F.; Mfinanga, S.; Kapata, N.; Mwaba, P.; McHugh, T.D. Tuberculosis: Progress and advances in development of new drugs, treatment regimens, and host-directed therapies. Lancet Infect. Dis. 2018, 18, e183–e198. [Google Scholar] [CrossRef] [PubMed]
- Horsburgh, C.R., Jr.; O’Donnell, M.; Chamblee, S.; Moreland, J.L.; Johnson, J.; Marsh, B.J.; Narita, M.; Johnson, L.S.; von Reyn, C.F. Revisiting rates of reactivation tuberculosis: A population-based approach. Am. J. Respir. Crit. Care Med. 2010, 182, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Dooley, K.E.; Kaplan, R.; Mwelase, N.; Grinsztejn, B.; Ticona, E.; Lacerda, M.; Sued, O.; Belonosova, E.; Ait-Khaled, M.; Angelis, K. Dolutegravir-based antiretroviral therapy for patients coinfected with tuberculosis and human immunodeficiency virus: A multicenter, noncomparative, open-label, randomized trial. Clin. Infect. Dis. 2020, 70, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rubio, L.; Martínez, B.; Donovan, D.M.; Rodríguez, A.; García, P. Bacteriophage virion-associated peptidoglycan hydrolases: Potential new enzybiotics. Crit. Rev. Microbiol. 2013, 39, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Yu, J.; Wei, H. Engineered bacteriophage lysins as novel anti-infectives. Front. Microbiol. 2014, 5, 542. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, S.; Ross, R.P.; Coffey, A. Bacteriophage and their lysins for elimination of infectious bacteria. FEMS Microbiol. Rev. 2009, 33, 801–819. [Google Scholar] [CrossRef] [PubMed]
- Fenton, M.; McAuliffe, O.; O’Mahony, J.; Coffey, A. Recombinant bacteriophage lysins as antibacterials. Bioeng. Bugs 2010, 1, 9–16. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, M.; Zhang, H.; Dai, J.; Guo, Z.; Li, X.; Ji, Y.; Cai, R.; Xi, H.; Wang, X. Antibacterial effects of phage lysin LysGH15 on planktonic cells and biofilms of diverse staphylococci. Appl. Environ. Microbiol. 2018, 84, e00886-18. [Google Scholar] [CrossRef]
- Domingo-Calap, P.; Georgel, P.; Bahram, S. Back to the future: Bacteriophages as promising therapeutic tools. HLA 2016, 87, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Keary, R.; Mcauliffe, O.; O’Mahony, J.; Coffey, A. Bacteriophage-derived peptidase CHAPK eliminates and prevents staphylococcal biofilms. Int. J. Microbiol. 2013, 2013, 625341. [Google Scholar]
- Kutter, E.; De Vos, D.; Gvasalia, G.; Alavidze, Z.; Gogokhia, L.; Kuhl, S.; Abedon, S.T. Phage therapy in clinical practice: Treatment of human infections. Curr. Pharm. Biotechnol. 2010, 11, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T.; García, P.; Mullany, P.; Aminov, R. Phage therapy: Past, present and future. Front. Microbiol. 2017, 8, 981. [Google Scholar] [CrossRef] [PubMed]
- Roach, D.R.; Leung, C.Y.; Henry, M.; Morello, E.; Singh, D.; Di Santo, J.P.; Weitz, J.S.; Debarbieux, L. Synergy between the host immune system and bacteriophage is essential for successful phage therapy against an acute respiratory pathogen. Cell Host Microbe 2017, 22, 38–47.e34. [Google Scholar] [CrossRef] [PubMed]
- Luria, S.E.; Delbrück, M. Mutations of bacteria from virus sensitivity to virus resistance. Genetics 1943, 28, 491. [Google Scholar] [CrossRef] [PubMed]
- Luria, S.E.; Human, M.L. A nonhereditary, host-induced variation of bacterial viruses. J. Bacteriol. 1952, 64, 557–569. [Google Scholar] [CrossRef]
- Pontali, E.; Raviglione, M.C.; Migliori, G.B. Regimens to treat multidrug-resistant tuberculosis: Past, present and future perspectives. Eur. Respir. Rev. 2019, 28, 190035. [Google Scholar] [CrossRef]
- Gordillo Altamirano, F.L.; Barr, J.J. Phage therapy in the postantibiotic era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar] [CrossRef] [PubMed]
- Fischetti, V.A. Development of phage lysins as novel therapeutics: A historical perspective. Viruses 2018, 10, 310. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, L.; Martínez, B.; Zhou, Y.; Rodríguez, A.; Donovan, D.M.; García, P. Lytic activity of the virion-associated peptidoglycan hydrolase HydH5 of Staphylococcus aureus bacteriophage vB_SauS-phiIPLA88. BMC Microbiol. 2011, 11, 138. [Google Scholar] [CrossRef] [PubMed]
- Gerstmans, H.; Criel, B.; Briers, Y. Synthetic biology of modular endolysins. Biotechnol. Adv. 2018, 36, 624–640. [Google Scholar] [CrossRef] [PubMed]
- Heselpoth, R.D.; Swift, S.M.; Linden, S.B.; Mitchell, M.S.; Nelson, D.C. Enzybiotics: Endolysins and bacteriocins. In Bacteriophages Biology, Technology, Therapy; Springer: Cham, Switzerland, 2021; pp. 989–1030. [Google Scholar]
- Rahman, M.U.; Wang, W.; Sun, Q.; Shah, J.A.; Li, C.; Sun, Y.; Li, Y.; Zhang, B.; Chen, W.; Wang, S. Endolysin, a promising solution against antimicrobial resistance. Antibiotics 2021, 10, 1277. [Google Scholar] [CrossRef]
- Belete, T.M. Recent progress in the development of novel mycobacterium cell wall inhibitor to combat drug-resistant tuberculosis. Microbiol. Insights 2022, 15, 11786361221099878. [Google Scholar] [CrossRef]
- van Schie, L.; Borgers, K.; Michielsen, G.; Plets, E.; Vuylsteke, M.; Tiels, P.; Festjens, N.; Callewaert, N. Exploration of synergistic action of cell wall-degrading enzymes against Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2021, 65, AAC0065921. [Google Scholar] [CrossRef]
- Silva, S.; Arinaminpathy, N.; Atun, R.; Goosby, E.; Reid, M. Economic impact of tuberculosis mortality in 120 countries and the cost of not achieving the Sustainable Development Goals tuberculosis targets: A full-income analysis. Lancet Glob. Health 2021, 9, e1372–e1379. [Google Scholar] [CrossRef]
- Gondil, V.S.; Harjai, K.; Chhibber, S. Endolysins as emerging alternative therapeutic agents to counter drug-resistant infections. Int. J. Antimicrob. Agents 2020, 55, 105844. [Google Scholar] [CrossRef]
- Grabowski, Ł.; Łepek, K.; Stasiłojć, M.; Kosznik-Kwaśnicka, K.; Zdrojewska, K.; Maciąg-Dorszyńska, M.; Węgrzyn, G.; Węgrzyn, A. Bacteriophage-encoded enzymes destroying bacterial cell membranes and walls, and their potential use as antimicrobial agents. Microbiol. Res. 2021, 248, 126746. [Google Scholar] [CrossRef]
- Pastagia, M.; Schuch, R.; Fischetti, V.A.; Huang, D.B. Lysins: The arrival of pathogen-directed anti-infectives. J. Med. Microbiol. 2013, 62, 1506–1516. [Google Scholar] [CrossRef]
- Pastagia, M.; Euler, C.; Chahales, P.; Fuentes-Duculan, J.; Krueger, J.G.; Fischetti, V.A. A novel chimeric lysin shows superiority to mupirocin for skin decolonization of methicillin-resistant and-sensitive Staphylococcus aureus strains. Antimicrob. Agents Chemother. 2011, 55, 738–744. [Google Scholar] [CrossRef]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef]
- Puiu, M.; Julius, C. Bacteriophage gene products as potential antimicrobials against tuberculosis. Biochem. Soc. Trans. 2019, 47, 847–860. [Google Scholar] [CrossRef]
- Nau, R.; Eiffert, H. Modulation of release of proinflammatory bacterial compounds by antibacterials: Potential impact on course of inflammation and outcome in sepsis and meningitis. Clin. Microbiol. Rev. 2002, 15, 95–110. [Google Scholar] [CrossRef]
- Nelson, D.C.; Schmelcher, M.; Rodriguez-Rubio, L.; Klumpp, J.; Pritchard, D.G.; Dong, S.; Donovan, D.M. Endolysins as antimicrobials. Adv. Virus Res. 2012, 83, 299–365. [Google Scholar] [PubMed]
- Gutiérrez, D.; Fernández, L.; Rodríguez, A.; García, P. Are phage lytic proteins the secret weapon to kill Staphylococcus aureus? MBio 2018, 9, e01923-17. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, R.; García, E.; García, P. Phage lysins for fighting bacterial respiratory infections: A new generation of antimicrobials. Front. Immunol. 2018, 9, 2252. [Google Scholar] [CrossRef]
- Loeffler, J.M.; Djurkovic, S.; Fischetti, V.A. Phage lytic enzyme Cpl-1 as a novel antimicrobial for pneumococcal bacteremia. Infect. Immun. 2003, 71, 6199–6204. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.-N.; Smith, D.L.; Young, R. Holins: The protein clocks of bacteriophage infections. Annu. Rev. Microbiol. 2000, 54, 799–825. [Google Scholar] [CrossRef]
- Cahill, J.; Young, R. Phage lysis: Multiple genes for multiple barriers. Adv. Virus Res. 2019, 103, 33–70. [Google Scholar]
- Kim, M.; Jo, Y.; Hwang, Y.J.; Hong, H.W.; Hong, S.S.; Park, K.; Myung, H. Phage-antibiotic synergy via delayed lysis. Appl. Environ. Microbiol. 2018, 84, e02085-18. [Google Scholar] [CrossRef]
- Catalao, M.J.; Gil, F.; Moniz-Pereira, J.; Sao-Jose, C.; Pimentel, M. Diversity in bacterial lysis systems: Bacteriophages show the way. FEMS Microbiol. Rev. 2013, 37, 554–571. [Google Scholar] [CrossRef]
- Drulis-Kawa, Z.; Majkowska-Skrobek, G.; Maciejewska, B. Bacteriophages and phage-derived proteins–application approaches. Curr. Med. Chem. 2015, 22, 1757–1773. [Google Scholar] [CrossRef] [PubMed]
- Aslam, B.; Arshad, M.I.; Aslam, M.A.; Muzammil, S.; Siddique, A.B.; Yasmeen, N.; Khurshid, M.; Rasool, M.; Ahmad, M.; Rasool, M.H. Bacteriophage proteome: Insights and potentials of an alternate to antibiotics. Infect. Dis. Ther. 2021, 10, 1171–1193. [Google Scholar] [CrossRef] [PubMed]
- CHATAIN-LY, M.H. The factors affecting effectiveness of treatment in phages therapy. Front. Microbiol. 2014, 5, 51. [Google Scholar] [CrossRef]
- Viertel, T.M.; Ritter, K.; Horz, H.-P. Viruses versus bacteria—Novel approaches to phage therapy as a tool against multidrug-resistant pathogens. J. Antimicrob. Chemother. 2014, 69, 2326–2336. [Google Scholar] [CrossRef] [PubMed]
- Krut, O.; Bekeredjian-Ding, I. Contribution of the immune response to phage therapy. J. Immunol. 2018, 200, 3037–3044. [Google Scholar] [CrossRef] [PubMed]
- Luong, T.; Salabarria, A.-C.; Roach, D.R. Phage therapy in the resistance era: Where do we stand and where are we going? Clin. Ther. 2020, 42, 1659–1680. [Google Scholar] [CrossRef] [PubMed]
- Kamilla, S.; Jain, V. Mycobacteriophage D29 holin C-terminal region functionally assists in holin aggregation and bacterial cell death. FEBS J. 2016, 283, 173–190. [Google Scholar] [CrossRef]
- Gigante, A.M.; Hampton, C.M.; Dillard, R.S.; Gil, F.; Catalão, M.J.; Moniz-Pereira, J.; Wright, E.R.; Pimentel, M. The Ms6 mycolyl-arabinogalactan esterase LysB is essential for an efficient mycobacteriophage-induced lysis. Viruses 2017, 9, 343. [Google Scholar] [CrossRef]
- Gil, F.; Grzegorzewicz, A.E.; Catalao, M.J.; Vital, J.; McNeil, M.R.; Pimentel, M. Mycobacteriophage Ms6 LysB specifically targets the outer membrane of Mycobacterium smegmatis. Microbiology 2010, 156, 1497. [Google Scholar] [CrossRef]
- Payne, K.; Sun, Q.; Sacchettini, J.; Hatfull, G.F. Mycobacteriophage Lysin B is a novel mycolylarabinogalactan esterase. Mol. Microbiol. 2009, 73, 367–381. [Google Scholar] [CrossRef]
- Lin, D.M.; Koskella, B.; Lin, H.C. Phage therapy: An alternative to antibiotics in the age of multi-drug resistance. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 162. [Google Scholar] [CrossRef] [PubMed]
- Lin, K. Exploiting Bacterial Lysis for Tuberculosis Drug Target Validation and Vaccine Development. Ph.D. Thesis, Weill Medical College of Cornell University, New York, NY, USA, 2018. [Google Scholar]
- Roach, D.R.; Donovan, D.M. Antimicrobial bacteriophage-derived proteins and therapeutic applications. Bacteriophage 2015, 5, e1062590. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Ao, Z. Phage in the diagnosis and treatment of tuberculosis. Front. Biosci. 2012, 17, 2691–2697. [Google Scholar] [CrossRef]
- Rybniker, J.; Kramme, S.; Small, P.L. Host range of 14 mycobacteriophages in Mycobacterium ulcerans and seven other mycobacteria including Mycobacterium tuberculosis–application for identification and susceptibility testing. J. Med. Microbiol. 2006, 55, 37–42. [Google Scholar] [CrossRef]
- Azimi, T.; Mosadegh, M.; Nasiri, M.J.; Sabour, S.; Karimaei, S.; Nasser, A. Phage therapy as a renewed therapeutic approach to mycobacterial infections: A comprehensive review. Infect. Drug Resist. 2019, 12, 2943–2959. [Google Scholar] [CrossRef] [PubMed]
- Albert, H.; Heydenrych, A.; Brookes, R.; Mole, R.J.; Harley, B.; Subotsky, E.; Henry, R.; Azevedo, V. Performance of a rapid phage-based test, FASTPlaqueTB™, to diagnose pulmonary tuberculosis from sputum specimens in South Africa. Int. J. Tuberc. Lung Dis. 2002, 6, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Swift, B.M.; Meade, N.; Barron, E.S.; Bennett, M.; Perehenic, T.; Hughes, V.; Stevenson, K.; Rees, C.E. The development and use of Actiphage® to detect viable mycobacteria from bovine tuberculosis and Johne’s disease-infected animals. Microb. Biotechnol. 2020, 13, 738–746. [Google Scholar] [CrossRef]
- Shield, C.G.; Swift, B.M.; McHugh, T.D.; Dedrick, R.M.; Hatfull, G.F.; Satta, G. Application of bacteriophages for mycobacterial infections, from diagnosis to treatment. Microorganisms 2021, 9, 2366. [Google Scholar] [CrossRef]
- Wei, L.; Wu, J.; Liu, H.; Yang, H.; Rong, M.; Li, D.; Zhang, P.; Han, J.; Lai, R. A mycobacteriophage-derived trehalose-6,6′-dimycolate-binding peptide containing both antimycobacterial and anti-inflammatory abilities. FASEB J. 2013, 27, 3067–3077. [Google Scholar] [CrossRef]
- Sharma, U.; Vipra, A.; Channabasappa, S. Phage-derived lysins as potential agents for eradicating biofilms and persisters. Drug Discov. Today 2018, 23, 848–856. [Google Scholar] [CrossRef]
- Śliwka, P.; Ochocka, M.; Skaradzińska, A. Applications of bacteriophages against intracellular bacteria. Crit. Rev. Microbiol. 2022, 48, 222–239. [Google Scholar] [CrossRef] [PubMed]
- Düzgüneş, N.; Sessevmez, M.; Yildirim, M. Bacteriophage therapy of bacterial infections: The rediscovered frontier. Pharmaceuticals 2021, 14, 34. [Google Scholar] [CrossRef] [PubMed]
- Allué-Guardia, A.; Saranathan, R.; Chan, J.; Torrelles, J.B. Mycobacteriophages as Potential Therapeutic Agents against Drug-Resistant Tuberculosis. Int. J. Mol. Sci. 2021, 22, 735. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, M.; Bajpai, K. The chamber of secretome in Mycobacterium tuberculosis as a potential therapeutic target. Biotechnol. Genet. Eng. Rev. 2022, 39, 1–44. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Name of Lytic Enzyme (Synonym) | Model | Target Pathogen | Result Summary | Reference |
|---|---|---|---|---|
| BTCU-1_ORF7 and BTCU-1_ORF8 | In vitro | M. smegmatis | Endolysin has been shown to effectively reduce TB infection, endolysins derived from the mycobacteriophage BTCU-1 have antimycobacterial activity | [5] |
| PK34 | Mouse | M. tuberculosis | PK34 binds to the glycolipid and cleared TB in the mouse model and reduced level of proinflammatory cytokines | [86] |
| TM4 | Mouse | M. avium, M. tuberculosis | Reduction of infection when lytic bacteriophage TM4 was delivered transiently kills both M. tuberculosis and M. avium residing within macrophages | [81] |
| LysB/Ms6 | In vitro | Mycobacterium spp. | Growth inhibition with surfactants | [61] |
| LysB/Bxz2 | In vitro | Mycobacterium spp. | Growth inhibition with surfactants | |
| LysA/BTCU-1 | In vitro | Mycobacterium spp. | Intracellular killing of M. smegatis | |
| LysB/BTCU-1 | In vitro | Mycobacterium spp. | Intracellular killing of M. smegatis | |
| Ms6 | M. smegmatis | The Ms6 lysis cassette has five genes involved in disruption of mycobacterium outer membrane | [7] | |
| Phage-D29-derived LysB | In vitro | M. smegmatis | Showed synergy with various anti-TB drugs against M. smegmatis cells | [87] |
| D29 phage | Mouse | M. tuberculosis | Significantly reduced the burden of M. tuberculosis at 24 h and 3 weeks post-infection compared to untreated mice. | [88] |
| D29 phage | Animal models | M. tuberculosis | Shows preventive measures against M. tuberculosis infection when administered in an inhalation formulation. | [89] |
| DS6A, TM4, D29, BTCU-1, SWU1 and Ms6 | In vivo (guinea pigs) | M. tuberculosis, M. ulcerans, M. avium | Elimination of M. tuberculosis. Reduction of infection in a number of organs and lesions | [82,90] |
| T7, P4, PDRPv, Bo4, Bxz2 | In vitro and in vivo | M. smegmatisi, M. tuberculosis, M. bovis | Reduction of infection in a number of organs and reduced lesions on some organs | [82,91] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mtimka, S.; Pillay, P.; Kwezi, L.; Pooe, O.J.; Tsekoa, T.L. An Exploratory Review of the Potential of Lytic Proteins and Bacteriophages for the Treatment of Tuberculosis. Microorganisms 2024, 12, 570. https://doi.org/10.3390/microorganisms12030570
Mtimka S, Pillay P, Kwezi L, Pooe OJ, Tsekoa TL. An Exploratory Review of the Potential of Lytic Proteins and Bacteriophages for the Treatment of Tuberculosis. Microorganisms. 2024; 12(3):570. https://doi.org/10.3390/microorganisms12030570
Chicago/Turabian StyleMtimka, Sibongile, Priyen Pillay, Lusisizwe Kwezi, Ofentse Jacob Pooe, and Tsepo Lebiletsa Tsekoa. 2024. "An Exploratory Review of the Potential of Lytic Proteins and Bacteriophages for the Treatment of Tuberculosis" Microorganisms 12, no. 3: 570. https://doi.org/10.3390/microorganisms12030570
APA StyleMtimka, S., Pillay, P., Kwezi, L., Pooe, O. J., & Tsekoa, T. L. (2024). An Exploratory Review of the Potential of Lytic Proteins and Bacteriophages for the Treatment of Tuberculosis. Microorganisms, 12(3), 570. https://doi.org/10.3390/microorganisms12030570