
Wolbachia and Asaia Distribution among Different Mosquito Vectors Is Affected by Tissue Localization and Host Species

 , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Collection and Identification
2.1.1. European Collections
2.1.2. African Collections
2.1.3. USA Collections
2.2. DNA Extraction
2.3. Wolbachia and Asaia Detection
2.4. Metagenomics Analysis
2.5. Asaia and Wolbachia Quantification via qPCR
3. Results
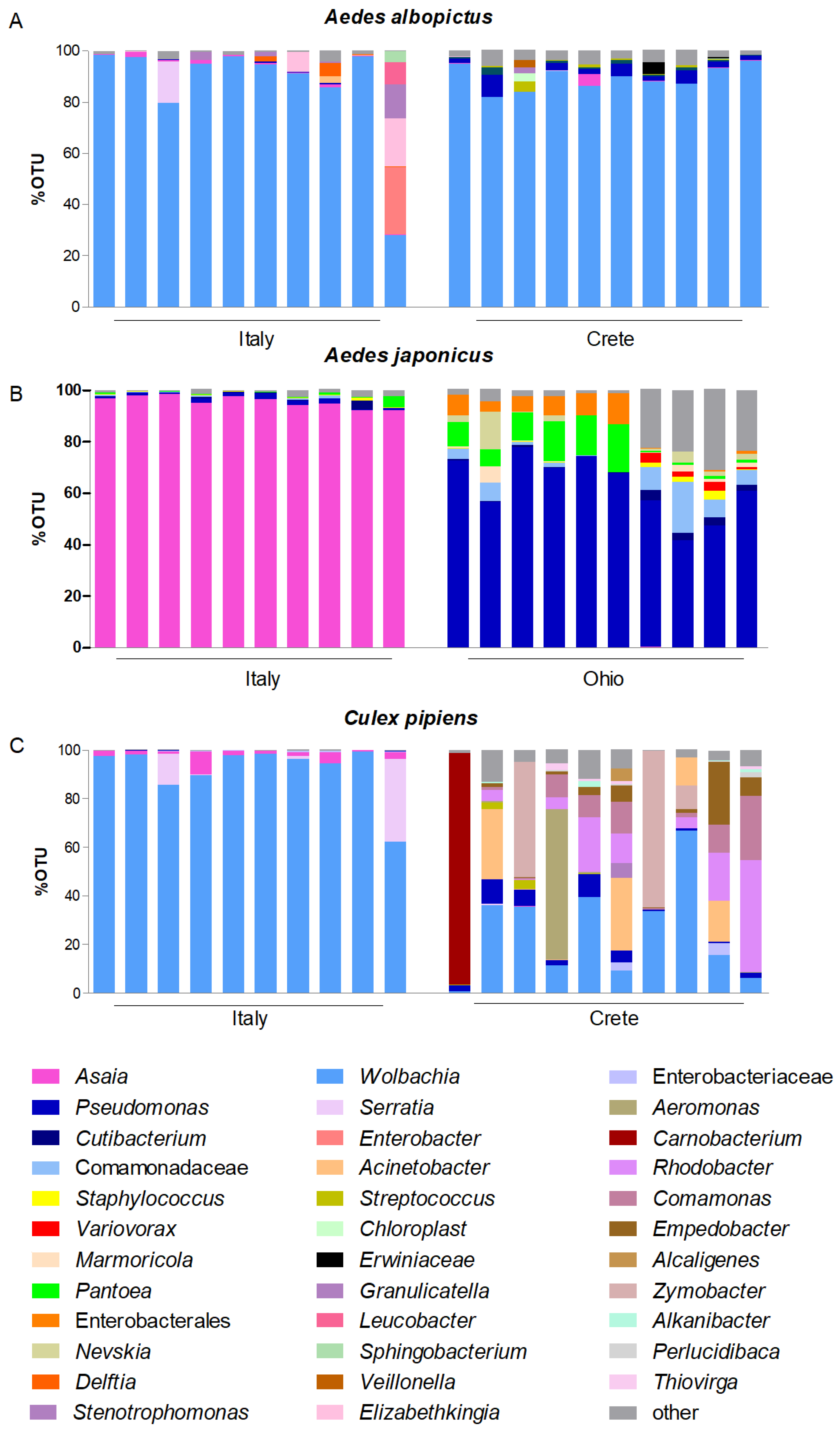
3.1. Asaia–Wolbachia Distribution in Different Mosquito Populations
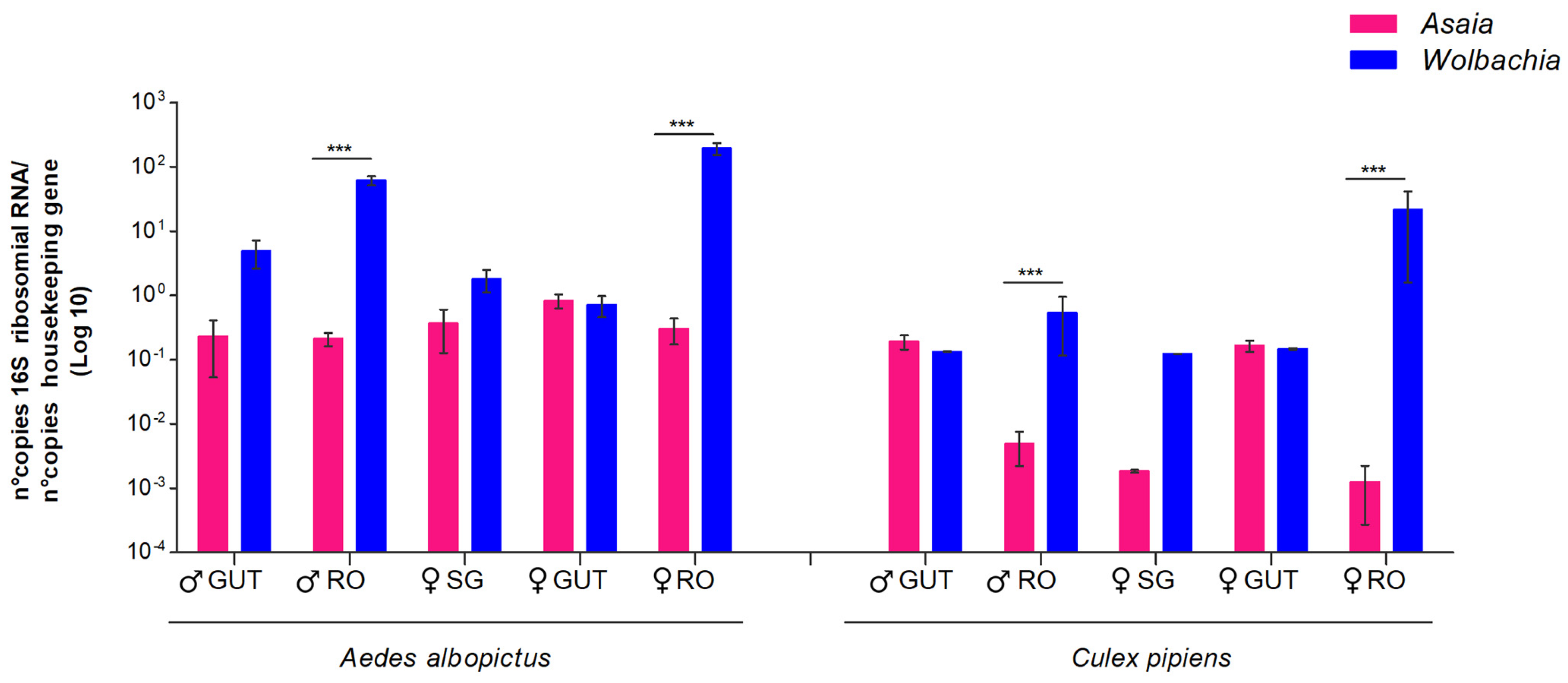
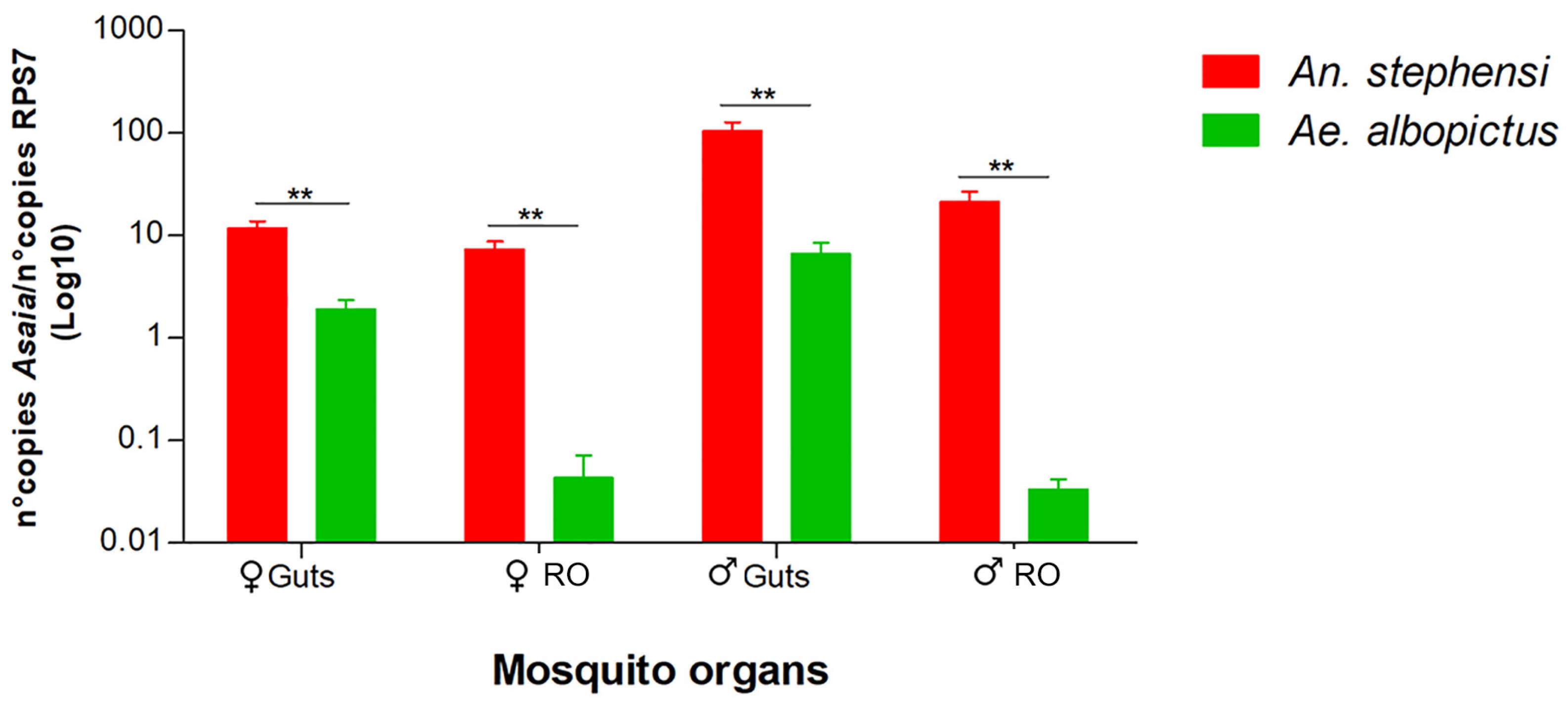
3.2. Competition in Different Mosquito Organs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Gao, L.; Aksoy, S. Microbiota in disease-transmitting vectors. Nat. Rev. Microbiol. 2023, 21, 604–618. [Google Scholar] [CrossRef] [PubMed]
- Riehle, M.A.; Jacobs-Lorena, M. Using bacteria to express and display anti-parasite molecules in mosquitoes: Current and future strategies. Insect Biochem. Mol. Biol. 2005, 35, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Didion, E.M.; Doyle, M.; Benoit, J.B. Bacterial Communities of Lab and Field Northern House Mosquitoes (Diptera: Culicidae) Throughout Diapause. J. Med. Entomol. 2022, 59, 648–658. [Google Scholar] [CrossRef]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef]
- Favia, G.; Ricci, I.; Damiani, C.; Raddadi, N.; Crotti, E.; Marzorati, M.; Rizzi, A.; Urso, R.; Brusetti, L.; Borin, S.; et al. Bacteria of the genus Asaia stably associate with Anopheles stephensi, an Asian malarial mosquito vector. Proc. Natl. Acad. Sci. USA 2007, 104, 9047–9051. [Google Scholar] [CrossRef] [PubMed]
- Mercant Osuna, A.; Gidley, A.; Mayi, M.P.A.; Bamou, R.; Dhokiya, V.; Antonio-Nkondjio, C.; Jeffries, C.L.; Walker, T. Diverse novel Wolbachia bacteria strains and genera-specific co-infections with Asaia bacteria in Culicine mosquitoes from ecologically diverse regions of Cameroon. Wellcome Open Res. 2023, 8, 267. [Google Scholar] [CrossRef]
- Frentiu, F.D.; Zakir, T.; Walker, T.; Popovici, J.; Pyke, A.T.; van den Hurk, A.; McGraw, E.A.; O’Neill, S.L. Limited dengue virus replication in field-collected Aedes aegypti mosquitoes infected with Wolbachia. PLoS Negl. Trop. Dis. 2014, 8, e2688. [Google Scholar] [CrossRef]
- Pinto, S.B.; Riback, T.I.S.; Sylvestre, G.; Costa, G.; Peixoto, J.; Dias, F.B.S.; Tanamas, S.K.; Simmons, C.P.; Dufault, S.M.; Ryan, P.A.; et al. Effectiveness of Wolbachia-infected mosquito deployments in reducing the incidence of dengue and other Aedes-borne diseases in Niterói, Brazil: A quasi-experimental study. PLoS Negl. Trop. Dis. 2021, 15, e0009556. [Google Scholar] [CrossRef]
- Utarini, A.; Indriani, C.; Ahmad, R.A.; Tantowijoyo, W.; Arguni, E.; Ansari, M.R.; Supriyati, E.; Wardana, D.S.; Meitika, Y.; Ernesia, I.; et al. Efficacy of Wolbachia-Infected Mosquito Deployments for the Control of Dengue. N. Engl. J. Med. 2021, 384, 2177–2186. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Ahmad, N.W.; Keong, W.M.; Ling, C.Y.; Ahmad, N.A.; Golding, N.; Tierney, N.; Jelip, J.; Putit, P.W.; Mokhtar, N.; et al. Introduction of Aedes aegypti mosquitoes carrying wAlbB Wolbachia sharply decreases dengue incidence in disease hotspots. iScience 2024, 27, 108942. [Google Scholar] [CrossRef]
- Ahmad, N.A.; Mancini, M.V.; Ant, T.H.; Martinez, J.; Kamarul, G.M.R.; Nazni, W.A.; Hoffmann, A.A.; Sinkins, S.P. Wolbachia strain w AlbB maintains high density and dengue inhibition following introduction into a field population of Aedes aegypti. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2021, 376, 20190809. [Google Scholar] [CrossRef] [PubMed]
- Nazni, W.A.; Hoffmann, A.A.; NoorAfizah, A.; Cheong, Y.L.; Mancini, M.V.; Golding, N.; Kamarul, G.M.R.; Arif, M.A.K.; Thohir, H.; NurSyamimi, H.; et al. Establishment of Wolbachia Strain wAlbB in Malaysian Populations of Aedes aegypti for Dengue Control. Curr. Biol. 2019, 29, 4241–4248.e4245. [Google Scholar] [CrossRef] [PubMed]
- Bian, G.; Joshi, D.; Dong, Y.; Lu, P.; Zhou, G.; Pan, X.; Xu, Y.; Dimopoulos, G.; Xi, Z. Wolbachia invades Anopheles stephensi populations and induces refractoriness to Plasmodium infection. Science 2013, 340, 748–751. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xi, Z. Wolbachia Transinfection Via Embryonic Microinjection. Methods Mol. Biol. 2024, 2739, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Shane, J.L.; Grogan, C.L.; Cwalina, C.; Lampe, D.J. Blood meal-induced inhibition of vector-borne disease by transgenic microbiota. Nat. Commun. 2018, 9, 4127. [Google Scholar] [CrossRef] [PubMed]
- Grogan, C.; Bennett, M.; Lampe, D.J. An evaluation of fusion partner proteins for paratransgenesis in Asaia bogorensis. PLoS ONE 2022, 17, e0273568. [Google Scholar] [CrossRef] [PubMed]
- Cappelli, A.; Damiani, C.; Mancini, M.V.; Valzano, M.; Rossi, P.; Serrao, A.; Ricci, I.; Favia, G. Asaia Activates Immune Genes in Mosquito Eliciting an Anti-Plasmodium Response: Implications in Malaria Control. Front. Genet. 2019, 10, 836. [Google Scholar] [CrossRef] [PubMed]
- Gonella, E.; Mandrioli, M.; Tedeschi, R.; Crotti, E.; Pontini, M.; Alma, A. Activation of Immune Genes in Leafhoppers by Phytoplasmas and Symbiotic Bacteria. Front. Physiol. 2019, 10, 795. [Google Scholar] [CrossRef]
- Pascar, J.; Middleton, H.; Dorus, S. Aedes aegypti microbiome composition covaries with the density of Wolbachia infection. Microbiome 2023, 11, 255. [Google Scholar] [CrossRef]
- Hughes, G.L.; Dodson, B.L.; Johnson, R.M.; Murdock, C.C.; Tsujimoto, H.; Suzuki, Y.; Patt, A.A.; Cui, L.; Nossa, C.W.; Barry, R.M.; et al. Native microbiome impedes vertical transmission of Wolbachia in Anopheles mosquitoes. Proc. Natl. Acad. Sci. USA 2014, 111, 12498–12503. [Google Scholar] [CrossRef]
- Rossi, P.; Ricci, I.; Cappelli, A.; Damiani, C.; Ulissi, U.; Mancini, M.V.; Valzano, M.; Capone, A.; Epis, S.; Crotti, E.; et al. Mutual exclusion of Asaia and Wolbachia in the reproductive organs of mosquito vectors. Parasit. Vectors 2015, 8, 278. [Google Scholar] [CrossRef] [PubMed]
- Andreychuk, S.; Yakob, L. Mathematical modelling to assess the feasibility of Wolbachia in malaria vector biocontrol. J. Theor. Biol. 2022, 542, 111110. [Google Scholar] [CrossRef] [PubMed]
- Montarsi, F.; Martini, S.; Dal Pont, M.; Delai, N.; Ferro Milone, N.; Mazzucato, M.; Soppelsa, F.; Cazzola, L.; Cazzin, S.; Ravagnan, S.; et al. Distribution and habitat characterization of the recently introduced invasive mosquito Aedes koreicus [Hulecoeteomyia koreica], a new potential vector and pest in north-eastern Italy. Parasit. Vectors 2013, 6, 292. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Valentini, A.; Dejean, T.; Montarsi, F.; Taberlet, P.; Glaizot, O.; Fumagalli, L. Detection of Invasive Mosquito Vectors Using Environmental DNA (eDNA) from Water Samples. PLoS ONE 2016, 11, e0162493. [Google Scholar] [CrossRef] [PubMed]
- Becker, N.; Petrić, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes: Identification, Ecology and Control, 3rd ed.; Springer Nature: Cham, Switzerland, 2020. [Google Scholar]
- Fotakis, E.A.; Mavridis, K.; Kampouraki, A.; Balaska, S.; Tanti, F.; Vlachos, G.; Gewehr, S.; Mourelatos, S.; Papadakis, A.; Kavalou, M.; et al. Mosquito population structure, pathogen surveillance and insecticide resistance monitoring in urban regions of Crete, Greece. PLoS Negl. Trop. Dis. 2022, 16, e0010186. [Google Scholar] [CrossRef] [PubMed]
- Gillies, M.; De Meillon, B. The Anophelinae of Africa South of the Sahara; South African Institute for Medical Research: Johannesburg, South Africa, 1968; p. 343. [Google Scholar]
- Gillies, M.; Coetzee, M. Supplement to the anophelinae of Africa south of the Sahara (afrotropical region). S. Afr. Inst. Med. Res. 1987, 55, 143. [Google Scholar]
- Chabi, J.; Van’t Hof, A.; N’dri, L.K.; Datsomor, A.; Okyere, D.; Njoroge, H.; Pipini, D.; Hadi, M.P.; de Souza, D.K.; Suzuki, T.; et al. Rapid high throughput SYBR green assay for identifying the malaria vectors Anopheles arabiensis, Anopheles coluzzii and Anopheles gambiae s.s. Giles. PLoS ONE 2019, 14, e0215669. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Nanfack-Minkeu, F.; Delong, A.; Luri, M.; Poelstra, J.W. Invasive Aedes japonicus Mosquitoes Dominate the Aedes Fauna Collected with Gravid Traps in Wooster, Northeastern Ohio, USA. Insects 2023, 14, 56. [Google Scholar] [CrossRef]
- Capone, A.; Ricci, I.; Damiani, C.; Mosca, M.; Rossi, P.; Scuppa, P.; Crotti, E.; Epis, S.; Angeletti, M.; Valzano, M.; et al. Interactions between Asaia, Plasmodium and Anopheles: New insights into mosquito symbiosis and implications in malaria symbiotic control. Parasit. Vectors 2013, 6, 182. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Author Correction: Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 1091. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Baldini, F.; Rougé, J.; Kreppel, K.; Mkandawile, G.; Mapua, S.A.; Sikulu-Lord, M.; Ferguson, H.M.; Govella, N.; Okumu, F.O. First report of natural Wolbachia infection in the malaria mosquito Anopheles arabiensis in Tanzania. Parasit. Vectors 2018, 11, 635. [Google Scholar] [CrossRef] [PubMed]
- Sawadogo, S.P.; Kabore, D.A.; Tibiri, E.B.; Hughes, A.; Gnankine, O.; Quek, S.; Diabaté, A.; Ranson, H.; Hughes, G.L.; Dabiré, R.K. Lack of robust evidence for a Wolbachia infection in Anopheles gambiae from Burkina Faso. Med. Vet. Entomol. 2022, 36, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.L.; Liew, J.W.K.; Wong, W.K.; Pramasivan, S.; Mohamed Hassan, N.; Wan Sulaiman, W.Y.; Jeyaprakasam, N.K.; Leong, C.S.; Low, V.L.; Vythilingam, I. Natural Wolbachia infection in field-collected Anopheles and other mosquito species from Malaysia. Parasit. Vectors 2020, 13, 414. [Google Scholar] [CrossRef] [PubMed]
- Waymire, E.; Duddu, S.; Yared, S.; Getachew, D.; Dengela, D.; Bordenstein, S.R.; Balkew, M.; Zohdy, S.; Irish, S.R.; Carter, T.E. Wolbachia 16S rRNA haplotypes detected in wild Anopheles stephensi in eastern Ethiopia. Parasit. Vectors 2022, 15, 178. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.A.; Robinson, K.L.; Yang, Q.; Callahan, A.G.; Schmidt, T.L.; Axford, J.K.; Coquilleau, M.P.; Staunton, K.M.; Townsend, M.; Ritchie, S.A.; et al. A decade of stability for wMel Wolbachia in natural Aedes aegypti populations. PLoS Pathog. 2022, 18, e1010256. [Google Scholar] [CrossRef]
- Damiani, C.; Cappelli, A.; Comandatore, F.; Montarsi, F.; Serrao, A.; Michelutti, A.; Bertola, M.; Mancini, M.V.; Ricci, I.; Bandi, C.; et al. Wolbachia in Aedes koreicus: Rare Detections and Possible Implications. Insects 2022, 13, 216. [Google Scholar] [CrossRef]
- Muturi, E.J.; Ramirez, J.L.; Rooney, A.P.; Kim, C.H. Comparative analysis of gut microbiota of mosquito communities in central Illinois. PLoS Negl. Trop. Dis. 2017, 11, e0005377. [Google Scholar] [CrossRef]
- Rosso, F.; Tagliapietra, V.; Albanese, D.; Pindo, M.; Baldacchino, F.; Arnoldi, D.; Donati, C.; Rizzoli, A. Reduced diversity of gut microbiota in two Aedes mosquitoes species in areas of recent invasion. Sci. Rep. 2018, 8, 16091. [Google Scholar] [CrossRef]
- Alfano, N.; Tagliapietra, V.; Rosso, F.; Manica, M.; Arnoldi, D.; Pindo, M.; Rizzoli, A. Changes in Microbiota Across Developmental Stages of. Front. Microbiol. 2019, 10, 2832. [Google Scholar] [CrossRef]
- Crotti, E.; Damiani, C.; Pajoro, M.; Gonella, E.; Rizzi, A.; Ricci, I.; Negri, I.; Scuppa, P.; Rossi, P.; Ballarini, P.; et al. Asaia, a versatile acetic acid bacterial symbiont, capable of cross-colonizing insects of phylogenetically distant genera and orders. Environ. Microbiol. 2009, 11, 3252–3264. [Google Scholar] [CrossRef] [PubMed]
- Möhlmann, T.W.R.; Vogels, C.B.F.; Göertz, G.P.; Pijlman, G.P.; Ter Braak, C.J.F.; Te Beest, D.E.; Hendriks, M.; Nijhuis, E.H.; Warris, S.; Drolet, B.S.; et al. Impact of Gut Bacteria on the Infection and Transmission of Pathogenic Arboviruses by Biting Midges and Mosquitoes. Microb. Ecol. 2020, 80, 703–717. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.; Camargo, C.; Avila, F.W. Characterization of the reproductive tract bacterial microbiota of virgin, mated, and blood-fed Aedes aegypti and Aedes albopictus females. Parasit. Vectors 2021, 14, 592. [Google Scholar] [CrossRef] [PubMed]
- Zouache, K.; Raharimalala, F.N.; Raquin, V.; Tran-Van, V.; Raveloson, L.H.; Ravelonandro, P.; Mavingui, P. Bacterial diversity of field-caught mosquitoes, Aedes albopictus and Aedes aegypti, from different geographic regions of Madagascar. FEMS Microbiol. Ecol. 2011, 75, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Raharimalala, F.N.; Boukraa, S.; Bawin, T.; Boyer, S.; Francis, F. Molecular detection of six (endo-) symbiotic bacteria in Belgian mosquitoes: First step towards the selection of appropriate paratransgenesis candidates. Parasitol. Res. 2016, 115, 1391–1399. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Name | Target | Forward Oligonucleotides (5′→3′) | Reverse Oligonucleotides (5′→3′) | Reference |
|---|---|---|---|---|
| Wolbachia | 16S rRNA | GAAGATAATGACGGTACTCAC | R2: GTCAGATTTGAACCAGATAGA R3: GTCACTGATCCCACTTTAAATAAC | [21] |
| AsaiaNew | 16S rRNA | GCGCGTAGGCGGTTTACAC | AGCGTCAGTAATGAGCCAGGTT | [5] |
| Asaia qPCR | 16S rRNA | TAGCGTTGCTCGGAATGACTGG | CGTATCAAATGCAGCCCCAAGG | [32] |
| Wolbachia qPCR | 16S rRNA | GAAGATAATGACGGTACTCAC | CCTACGCGCTCTTTACGCCCA | This work |
| Ae-rps7 | RPS7 | CGCGCTCGTGAGATCGA | GCACCGGGACGTAGATCA | [21] |
| Cx-rps3 | RPS3 | AGCGTGCCAAGTCGATGAG | ACGTACTCGTTGCACGGATCTC | [21] |
| As-rps7 | RPS7 | AGCAGCAGCAGCACTTGATTTG | TAAACGGCTTTCTGCGTCACCC | [32] |
| Mosquito Species | Collection Site | Country | Asaia % (Positive/Total) | Wolbachia % (Positive/Total) |
|---|---|---|---|---|
| An. arabiensis | Yangah | Cameroon | 9/37 (24.3) | 0/37 (0) |
| An. coluzzii | Yangah | Cameroon | 1/10 (10) | 0/10 (0) |
| Yaoundé | Cameroon | 33/39 (84.6) | 0/39 (0) | |
| Douala | Cameroon | 20/38 (52.6) | 0/38 (0) | |
| Mbalmayo | Cameroon | 34/34 (100) | 0/34 (0) | |
| An. funestus | Ouagadougou | Burkina Faso | 32/69 (46.4) | 0/69 (0) |
| An. gambiae | Santchou | Cameroon | 15/45 (33.3) | 0/45(0) |
| Yaoundé | Cameroon | 16/25 (64) | 0/25 (0) | |
| Adamaoua | Cameroon | 1/48 (2.1) | 0/48 (0) | |
| Douala | Cameroon | 13/25 (52) | 0/25 (0) | |
| Mbandjock | Cameroon | 41/41 (100) | 0/41(0) | |
| Mbalmayo | Cameroon | 5/5 (100) | 0/5 (0) | |
| An. pharoensis | Yangah | Cameroon | 14/48 (29.2) | 0/48 (0) |
| An. ziemanni | Yangah | Cameroon | 9/48 (18.8) | 0/48 (0) |
| Ae. aegypti | Ouagadougou | Burkina Faso | 3/7 (42.9) | 0/7 (0) |
| Ae. albopictus | Petriolo | Italy | 72/72 (100) | 72/72 (100) |
| Sospirolo | Italy | 11/12 (91.6) | 12/12 (100) | |
| Pedavena | Italy | 11/11 (100) | 11/11 (100) | |
| Feltre | Italy | 1/1 (100) | 1/1 (100) | |
| Dschang | Cameroon | 44/44 (100) | 44/44 (100) | |
| Gazi | Crete | 0/5 (0) | 5/5 (100) | |
| Heraklion | Crete | 1/8 (12.5) | 8/8 (100) | |
| Rethymnon | Crete | 9/21 (42.9) | 21/21 (100) | |
| Hersonissos | Crete | 4/26 (15.4) | 26/26 (100) | |
| Ae. koreicus | Sospirolo | Italy | 32/33 (97) | 0/33 (0) |
| Pedavena | Italy | 52/52 (100) | 0/52 (0) | |
| Feltre | Italy | 20/20 (100) | 0/20 (0) | |
| Alano di Piave | Italy | 24/24 (100) | 0/24 (0) | |
| Ae. japonicus | Sospirolo | Italy | 20/20 (100) | 0/20 (0) |
| Pedavena | Italy | 29/29 (100) | 0/29 (0) | |
| Feltre | Italy | 37/38 (97.4) | 0/38 (0) | |
| Wooster | Ohio (USA) | 19/39 (48.7) | 0/39 (0) | |
| Cx. pipiens | Camerino | Italy | 38/38 (100) | 38/38 (100) |
| Pedavena | Italy | 2/2 (100) | 2/2 (100) | |
| Pisa | Italy | 42/42 (100) | 42/42 (100) | |
| Rethymnon | Crete | 6/11 (54.5) | 11/11 (100) | |
| Heraklion | Crete | 4/9 (44.4) | 9/9 (100) | |
| Hersonissos | Crete | 8/22 (36.4) | 22/22 (100) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ilbeigi Khamseh Nejad, M.; Cappelli, A.; Damiani, C.; Falcinelli, M.; Catapano, P.L.; Nanfack-Minkeu, F.; Mayi, M.P.A.; Currà, C.; Ricci, I.; Favia, G. Wolbachia and Asaia Distribution among Different Mosquito Vectors Is Affected by Tissue Localization and Host Species. Microorganisms 2024, 12, 545. https://doi.org/10.3390/microorganisms12030545
Ilbeigi Khamseh Nejad M, Cappelli A, Damiani C, Falcinelli M, Catapano PL, Nanfack-Minkeu F, Mayi MPA, Currà C, Ricci I, Favia G. Wolbachia and Asaia Distribution among Different Mosquito Vectors Is Affected by Tissue Localization and Host Species. Microorganisms. 2024; 12(3):545. https://doi.org/10.3390/microorganisms12030545
Chicago/Turabian StyleIlbeigi Khamseh Nejad, Mahdokht, Alessia Cappelli, Claudia Damiani, Monica Falcinelli, Paolo Luigi Catapano, Ferdinand Nanfack-Minkeu, Marie Paul Audrey Mayi, Chiara Currà, Irene Ricci, and Guido Favia. 2024. "Wolbachia and Asaia Distribution among Different Mosquito Vectors Is Affected by Tissue Localization and Host Species" Microorganisms 12, no. 3: 545. https://doi.org/10.3390/microorganisms12030545
APA StyleIlbeigi Khamseh Nejad, M., Cappelli, A., Damiani, C., Falcinelli, M., Catapano, P. L., Nanfack-Minkeu, F., Mayi, M. P. A., Currà, C., Ricci, I., & Favia, G. (2024). Wolbachia and Asaia Distribution among Different Mosquito Vectors Is Affected by Tissue Localization and Host Species. Microorganisms, 12(3), 545. https://doi.org/10.3390/microorganisms12030545