
The Efficacy of Selected Probiotic Strains and Their Combination to Inhibit the Interaction of Adherent-Invasive Escherichia coli (AIEC) with a Co-Culture of Caco-2:HT29-MTX Cells
 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Probiotic Strains
2.2. E. coli Strains
2.3. Co-Culture of Cells
2.4. Adhesion Assay
2.5. Invasion Assay
2.6. Translocation Assay
2.7. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guzman, J.R.; Conlin, V.S.; Jobin, C. Diet, microbiome, and the intestinal epithelium: An essential triumvirate? BioMed Res. Int. 2013, 2013, 425146. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic mechanisms of action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Chervy, M.; Barnich, N.; Denizot, J. Adherent-invasive E. coli: Update on the lifestyle of a troublemaker in Crohn’s disease. Int. J. Mol. Sci. 2020, 21, 3734. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.; Blaser, M.J. The human microbiome: At the interface of health and disease. Nat. Rev. Gen. 2012, 13, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Manichanh, C.; Borruel, N.; Casellas, F.; Guarner, F. The gut microbiota in IBD. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; Darfeuillemichaud, A. The commensal microbiota and enteropathogens in the pathogenesis of inflammatory bowel diseases. Gastroenterology 2011, 140, 1720–1728.e3. [Google Scholar] [CrossRef] [PubMed]
- Palmela, C.; Chevarin, C.; Xu, Z.; Torres, J.; Sevrin, G.; Hirten, R.; Barnich, N.; Ng, S.C.; Colombel, J.F. Adherent-invasive Escherichia coli in inflammatory bowel disease. Gut 2018, 67, 574–587. [Google Scholar] [CrossRef]
- Conte, M.P.; Longhi, C.; Marazzato, M.; Conte, A.L.; Aleandri, M.; Lepanto, M.S.; Zagaglia, C.; Nicoletti, M.; Aloi, M.; Totino, V.; et al. Adherent-invasive Escherichia coli (AIEC) in pediatric Crohn’s disease patients: Phenotypic and genetic pathogenic features. BMC Res. Notes 2014, 7, 748. [Google Scholar] [CrossRef]
- Martinez-Medina, M.; Denizot, J.; Dreux, N.; Robin, F.; Billard, E.; Bonnet, R.; Darfeuille-Michaud, A.; Barnich, N. Western diet induces dysbiosis with increased E coli in CEABAC10 mice, alters host barrier function favouring AIEC colonisation. Gut 2014, 63, 116–124. [Google Scholar] [CrossRef]
- Yang, Y.; Liao, Y.; Ma, Y.; Gong, W.; Zhu, G. The role of major virulence factors of AIEC involved in inflammatory bowl disease—A mini-review. Appl. Microbiol. Biotechnol. 2017, 101, 7781–7787. [Google Scholar] [CrossRef] [PubMed]
- Danese, S.; Fiocchi, C. Etiopathogenesis of inflammatory bowel diseases. World J. Gastroenterol. 2006, 12, 4807–4812. [Google Scholar] [CrossRef] [PubMed]
- Darfeuille-Michaud, A.; Boudeau, J.; Bulois, P.; Neut, C.; Glasser, A.L.; Barnich, N.; Bringer, M.A.; Swidsinski, A.; Beaugerie, L.; Colombel, J.F. High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohn’s disease. Gastroenterology 2004, 127, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Kostic, A.D.; Xavier, R.J.; Gevers, D. The microbiome in inflammatory bowel disease: Current status and the future ahead. Gastroenterology 2014, 146, 1489–1499. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Medina, M.; Aldeguer, X.; Lopez-Siles, M.; González-Huix, F.; López-Oliu, C.; Dahbi, G.; Bianco, J.E.; Blanco, J.; Garcia-Gil, L.J.; Darfeuille-Michaud, A. Molecular diversity of Escherichia coli in the human gut: New ecological evidence supporting the role of adherent-invasive E. coli (AIEC) in Crohn’s disease. Inflamm. Bowel Dis. 2009, 15, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.T.; Sharma, V.; Elmén, L.; Peterson, S.N. Immune homeostasis, dysbiosis and therapeutic modulation of the gut microbiota. Clin. Exp. Immunol. 2015, 179, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Hertzberger, R.Y.; Knaus, U.G. Hydrogen peroxide production by lactobacilli promotes epithelial restitution during colitis. Redox Biol. 2018, 16, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.L.; Farias, A.Q.; Rezaie, A. Gastrointestinal motility and absorptive disorders in patients with inflammatory bowel diseases: Prevalence, diagnosis and treatment. World J. Gastroenterol. 2019, 25, 4414–4426. [Google Scholar] [CrossRef]
- McLean, L.P.; Cross, R.K. Integrin antagonists as potential therapeutic options for the treatment of Crohns disease. Expert Opin. Investig. Drugs 2016, 25, 263–273. [Google Scholar] [CrossRef]
- Varankovich, N.V.; Nickerson, M.T.; Korber, D.R. Probiotic-based strategies for therapeutic and prophylactic use against multiple gastrointestinal diseases. Front. Microbiol. 2015, 6, 685. [Google Scholar] [CrossRef]
- Nami, Y.; Haghshenas, B.; Abdullah, N.; Barzegari, A.; Radiah, D.; Rosli, R.; Khosroushahi, A.Y. Probiotics or antibiotics: Future challenges in medicine. J. Med. Microbiol. 2015, 64, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Jakobsson, H.E.; Jernberg, C.; Andersson, A.F.; Sjölund-Karlsson, M.; Jansson, J.K.; Engstrand, L. Short-term antibiotic treatment has differing long- term impacts on the human throat and gut microbiome. PLoS ONE 2010, 5, e9836. [Google Scholar] [CrossRef] [PubMed]
- Hotel, A. Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria. Prevention 2001, 5, 1–10. [Google Scholar]
- McFarland, L.V. From Yaks to Yogurt: The History, Development, and Current Use of Probiotics. Clin. Infect. Dis. 2015, 60 (Suppl. S2), S85–S90. [Google Scholar] [CrossRef] [PubMed]
- Adriana, N.; Ilona, M.; Katarzyna, Ś.; Zdzisława, L.; Elżbieta, K. Adherence of probiotic bacteria to human colon epithelial cells and inhibitory effect against enteric pathogens—In vitro study. Int. J. Dairy. Technol. 2016, 69, 532–539. [Google Scholar] [CrossRef]
- Collado, M.C.; Meriluoto, J.; Salminen, S. In vitro analysis of probiotic strain combinations to inhibit pathogen adhesion to human intestinal mucus. Food Res. Int. 2007, 40, 629–636. [Google Scholar] [CrossRef]
- Collado, M.C.; Meriluoto, J.; Salminen, S. Development of new probiotics by strain combinations: Is it possible to improve the adhesion to intestinal mucus? J. Dairy. Sci. 2007, 90, 2710–2716. [Google Scholar] [CrossRef]
- Kechagia, M.; Basoulis, D.; Konstantopoulou, S.; Dimitriadi, D.; Gyftopoulou, K.; Skarmoutsou, N.; Fakiri, E.M. Health benefits of probiotics: A review. ISRN Nutr. 2013, 2013, 481651. [Google Scholar] [CrossRef]
- Santacroce, L.; Charitos, I.A.; Bottalico, L. A successful history: Probiotics and their potential as antimicrobials. Expert. Rev. Anti-Infect. Ther. 2019, 17, 635–645. [Google Scholar] [CrossRef]
- Fontana, L.; Bermudez-Brito, M.; Plaza-Diaz, J.; Muñoz-Quezada, S.; Gil, A. Sources, isolation, characterisation and evaluation of probiotics. Br. J. Nutr. 2013, 109 (Suppl. S2), S35–S50. [Google Scholar] [CrossRef]
- Parvez, S.; Malik, K.A.; Ah Kang, S.; Kim, H.Y. Probiotics and their fermented food products are beneficial for health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- Laksemi, D.A.A.S.; Ketut Tunas, I.; Diarthini, N.L.P.E. A brief review of probiotic: Action mechanism, benefits, and clinical application on human health. J. Glob. Pharma Technol. 2018, 10, 19–29. [Google Scholar]
- Sanders, M.E.; Merenstein, D.; Merrifield, C.A.; Hutkins, R. Probiotics for human use. Nutr. Bull. 2018, 43, 212–225. [Google Scholar] [CrossRef]
- Tuomola, E.; Crittenden, R.; Playne, M.; Isolauri, E.; Salminen, S. Quality assurance criteria for probiotic bacteria. Am. J. Clin. Nutr. 2001, 73 (Suppl. S2), 393S–398S. [Google Scholar] [CrossRef] [PubMed]
- Piatek, J.; Gibas-Dorna, M.; Olejnik, A.; Krauss, H.; Wierzbicki, K.; Zukiewicz-Sobczak, W.; Głowacki, M. The viability and intestinal epithelial cell adhesion of probiotic strain combination—In vitro study. Ann. Agric. Environ. Med. 2012, 19, 99–102. [Google Scholar]
- Shewale, R.N.; Sawale, P.D.; Khedkar, C.D.; Singh, A. Selection criteria for probiotics: A review. Int. J. Probiotics Prebiotics 2014, 9, 17–22. [Google Scholar]
- Collado, M.C.; Meriluoto, J.; Salminen, S. Role of commercial probiotic strains against human pathogen adhesion to intestinal mucus. Lett. Appl. Microbiol. 2007, 45, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Salminen, S.; Nybom, S.; Meriluoto, J.; Collado, M.C.; Vesterlund, S.; El-Nezami, H. Interaction of probiotics and pathogens-benefits to human health? Curr. Opin. Biotechnol. 2010, 21, 157–167. [Google Scholar] [CrossRef]
- Nagpal, R.; Kumar, A.; Kumar, M.; Behare, P.V.; Jain, S.; Yadav, H. Probiotics, their health benefits and applications for developing healthier foods: A review. FEMS Microbiol. Lett. 2012, 334, 1–15. [Google Scholar] [CrossRef]
- Roy, D. Probiotics. In Comprehensive Biotechnology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 649–661. [Google Scholar]
- Stavropoulou, E.; Bezirtzoglou, E. Probiotics in Medicine: A Long Debate. Front. Immunol. 2020, 11, 2192. [Google Scholar] [CrossRef]
- Arena, M.P.; Capozzi, V.; Russo, P.; Drider, D.; Spano, G.; Fiocco, D. Immunobiosis and probiosis: Antimicrobial activity of lactic acid bacteria with a focus on their antiviral and antifungal properties. Appl. Microbiol. Biotechnol. 2018, 102, 9949–9958. [Google Scholar] [CrossRef] [PubMed]
- McFarland, L.V. Efficacy of Single-Strain Probiotics Versus Multi-Strain Mixtures: Systematic Review of Strain and Disease Specificity. Dig. Dis. Sci. 2021, 66, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Kajander, K.; Korpela, R. Clinical studies on alleviating the symptoms of irritable bowel syndrome with a probiotic combination. Asia Pac. J. Clin. Nutr. 2006, 15, 576–580. [Google Scholar] [PubMed]
- Wang, Y.; Xie, Q.; Zhang, Y.; Ma, W.; Ning, K.; Xiang, J.Y.; Cui, J.; Xiang, H. Combination of probiotics with different functions alleviate DSS-induced colitis by regulating intestinal microbiota, IL-10, and barrier function. Appl. Microbiol. Biotechnol. 2020, 104, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Gerritsen, J.; Smidt, H.; Rijkers, G.T.; De Vos, W.M. Intestinal microbiota in human health and disease: The impact of probiotics. Genes Nutr. 2011, 6, 209–240. [Google Scholar] [CrossRef]
- Soni, R.; Jain, N.K.; Shah, V.; Soni, J.; Suthar, D.; Gohel, P. Development of probiotic yogurt: Effect of strain combination on nutritional, rheological, organoleptic and probiotic properties. J. Food Sci. Technol. 2020, 57, 2038–2050. [Google Scholar] [CrossRef]
- Chapman, C.M.; Gibson, G.R.; Rowland, I. Health benefits of probiotics: Are mixtures more effective than single strains? Eur. J. Nutr. 2011, 50, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Verdenelli, M.C.; Silvi, S.; Cecchini, C.; Orpianesi, C.; Cresci, A. Influence of a combination of two potential probiotic strains, Lactobacillus rhamnosus IMC 501® and Lactobacillus paracasei IMC 502® on bowel habits of healthy adults. Lett. Appl. Microbiol. 2011, 52, 596–602. [Google Scholar] [CrossRef]
- Collado, M.C.; Grześkowiak, Ł.; Salminen, S. Probiotic strains and their combination inhibit in vitro adhesion of pathogens to pig intestinal mucosa. Curr. Microbiol. 2007, 55, 260–265. [Google Scholar] [CrossRef]
- Rohani, M.; Noohi, N.; Talebi, M.; Katouli, M.; Pourshafie, M.R. Highly heterogeneous probiotic Lactobacillus species in healthy iranians with low functional activities. PLoS ONE 2015, 10, e0144467. [Google Scholar] [CrossRef]
- Astley, D.J.; Masters, N.; Kuballa, A.; Katouli, M. Commonality of adherent-invasive Escherichia coli isolated from patients with extraintestinal infections, healthy individuals and the environment. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Bibalan, M.H.; Eshaghi, M.; Rohani, M.; Esghaei, M.; Darban-Sarokhalil, D.; Pourshafie, M.R.; Talebi, M. Isolates of Lactobacillus plantarum and L. Reuteri display greater antiproliferative and antipathogenic activity than other Lactobacillus isolates. J. Med. Microbiol. 2017, 66, 1416–1420. [Google Scholar] [CrossRef] [PubMed]
- Smit, B.; Chinaka, C.C.; Scott, A.A.; Gaiduschek, K.; Hatje, E.; Kuballa, A.; Coulson, S.; Finlayson, W.; Katouli, M. Efficacy of Selected Live Biotherapeutic Candidates to Inhibit the Interaction of an Adhesive-Invasive Escherichia coli Strain with Caco-2, HT29-MTX Cells and Their Co-Culture. Biomedicines 2022, 10, 2245. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, N.L.; Katouli, M.; Polkinghorne, A. Genomic comparison of translocating and non-translocating Escherichia coli. PLoS ONE 2015, 10, e0137131. [Google Scholar] [CrossRef] [PubMed]
- Nettelbladt, C.G.; Katouli, M.; Bark, T.; Svenberg, T.; Möllby, R.; Ljungqvist, O. Evidence of bacterial translocation in fatal hemorrhagic pancreatitis. J. Trauma. 2000, 48, 314–315. [Google Scholar] [CrossRef] [PubMed]
- Ramos, N.L.; Lamprokostopoulou, A.; Chapman, T.A.; Chin, J.C.; Römling, U.; Brauner, A.; Katouli, M. Virulence characteristics of translocating Escherichia coli and the interleukin-8 response to infection. Microb. Pathog. 2011, 50, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Mahler, G.J.; Shuler, M.L.; Glahn, R.P. Characterization of Caco-2 and HT29-MTX cocultures in an in vitro digestion/cell culture model used to predict iron bioavailability. J. Nutr. Biochem. 2009, 20, 494–502. [Google Scholar] [CrossRef]
- Wan, L.Y.M.; Allen, K.J.; Turner, P.C.; El-nezami, H. Modulation of mucin mRNA (MUC5AC AND MUC5B) expression and protein production and secretion in Caco-2/HT29-MTX Co-cultures following exposure to individual and combined Fusarium mycotoxins. Toxicol. Sci. 2014, 139, 83–98. [Google Scholar] [CrossRef]
- Duncan, M.J.; Li, G.; Shin, J.S.; Carson, J.L.; Abraham, S.N. Bacterial Penetration of Bladder Epithelium through Lipid Rafts. J. Biol. Chem. 2004, 279, 18944–18951. [Google Scholar] [CrossRef]
- Alizadeh Behbahani, B.; Noshad, M.; Falah, F. Inhibition of Escherichia coli adhesion to human intestinal Caco-2 cells by probiotic candidate Lactobacillus plantarum strain L15. Microb. Pathog. 2019, 136, 103677. [Google Scholar] [CrossRef]
- Gagnon, M.; Zihler Berner, A.; Chervet, N.; Chassard, C.; Lacroix, C. Comparison of the Caco-2, HT-29 and the mucus-secreting HT29-MTX intestinal cell models to investigate Salmonella adhesion and invasion. J. Microbiol. Methods 2013, 94, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Celebioglu, H.U.; Olesen, S.V.; Prehn, K.; Lahtinen, S.J.; Brix, S.; Abou Hachem, M.; Svensson, B. Mucin- and carbohydrate-stimulated adhesion and subproteome changes of the probiotic bacterium Lactobacillus acidophilus NCFM. J. Proteom. 2017, 163, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Morrin, S.T.; Hickey, R.M. New insights on the colonization of the human gut by health-promoting bacteria. Appl. Microbiol. Biotechnol. 2020, 104, 1511–1515. [Google Scholar] [CrossRef] [PubMed]
- Reale, O.; Huguet, A.; Fessard, V. Co-culture model of Caco-2/HT29-MTX cells: A promising tool for investigation of phycotoxins toxicity on the intestinal barrier. Chemosphere 2020, 273, 128497. [Google Scholar] [CrossRef] [PubMed]
- Sekirov, I.; Russell, S.L.; Caetano M Antunes, L.; Finlay, B.B. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [PubMed]
- Afolabi, O.; Bankole, O.; Olaitan, O. Production and characterization of antimicrobial agents by Lactic Acid Bacteria Isolated from Fermented Foods. Internet J. Microbiol. 2008, 4, 1–7. [Google Scholar]
- Van den Abbeele, P.; Marzorati, M.; Derde, M.; De Weirdt, R.; Joan, V.; Possemiers, S.; Van de Wiele, T. Arabinoxylans, inulin and Lactobacillus reuteri 1063 repress the adherent-invasive Escherichia coli from mucus in a mucosa-comprising gut model. NPJ Biofilms Microbiomes 2016, 2, 16016. [Google Scholar] [CrossRef]
- Bernet, M.F.; Brassart, D.; Neeser, J.R.; Servin, A.L. Lactobacillus acidophilus LA 1 binds to cultured human intestinal cell lines and inhibits cell attachment and cell invasion by enterovirulent bacteria. Gut 1994, 35, 483–489. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, N.; Prete, R.; Battista, N.; Corsetti, A. Adhesion Properties of Food-Associated Lactobacillus plantarum Strains on Human Intestinal Epithelial Cells and Modulation of IL-8 Release. Front. Microbiol. 2018, 9, 2392. [Google Scholar] [CrossRef]
- Katouli, M.; Ramos, N.L.; Nettelbladt, C.G.; Ljungdahl, M.; Robinson, W.; Ison, H.M.; Brauner, A.; Möllby, R. Host species-specific translocation of Escherichia coli. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 1095–1103. [Google Scholar] [CrossRef]
- Wine, E.; Ossa, J.C.; Gray-Owen, S.D.; Sherman, P.M. Adherent-invasive Escherichia coli target the epithelial barrier. Gut Microbes 2010, 1, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Agus, A.; Massier, S.; Darfeuille-Michaud, A.; Billard, E.; Barnich, N. Understanding Host-Adherent-Invasive Escherichia coli Interaction in Crohn’s Disease: Opening Up New Therapeutic Strategies. BioMed Res. Int. 2014, 2014, 567929. [Google Scholar] [CrossRef] [PubMed]
- Ingrassia, I.; Leplingard, A.; Darfeuille-Michaud, A. Lactobacillus casei DN-114 001 Inhibits the Ability of Adherent-Invasive Escherichia coli Isolated from Crohn’s Disease Patients To Adhere to and To Invade Intestinal Epithelial Cells. Appl. Environ. Microbiol. 2005, 71, 2880–2887. [Google Scholar] [CrossRef]
- Ghanavati, R.; Mohammadi, F.; Javadi, A.; Shahosseini, Z.; Talebi, M.; Rohani, M. The effects of a mixture of Lactobacillus species on colorectal tumor cells activity through modulation of Hes1 pathway. PharmaNutrition 2020, 13, 100207. [Google Scholar] [CrossRef]
- Ghanavati, R.; Akbari, A.; Mohammadi, F.; Asadollahi, P.; Javadi, A.; Talebi, M.; Rohani, M. Lactobacillus species inhibitory effect on colorectal cancer progression through modulating the Wnt/β-catenin signaling pathway. Mol. Cell Biochem. 2020, 470, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Aghamohammad, S.; Sepehr, A.; Miri, S.T.; Najafi, S.; Pourshafie, M.R.; Rohani, M. Anti-inflammatory and immunomodulatory effects of Lactobacillus spp. as a preservative and therapeutic agent for IBD control. Immun. Inflamm. Dis. 2022, 10, e635. [Google Scholar] [CrossRef] [PubMed]
- Fijan, S.; Šulc, D.; Steyer, A. Study of the In Vitro Antagonistic Activity of Various Single-Strain and Multi-Strain Probiotics against Escherichia coli. Int. J. Environ. Res. Public Health 2018, 15, 1539. [Google Scholar] [CrossRef] [PubMed]
- Campana, R.; van Hemert, S.; Baffone, W. Strain-specific probiotic properties of lactic acid bacteria and their interference with human intestinal pathogens invasion. Gut Pathog. 2017, 9, 12. [Google Scholar] [CrossRef]
- Asadollahi, P.; Ghanavati, R.; Rohani, M.; Razavi, S.; Esghaei, M.; Talebi, M. Anti-cancer effects of Bifidobacterium species in colon cancer cells and a mouse model of carcinogenesis. PLoS ONE 2020, 15, e0232930. [Google Scholar]
- Béduneau, A.; Tempesta, C.; Fimbel, S.; Pellequer, Y.; Jannin, V.; Demarne, F.; Lamprecht, A. A tunable Caco-2/HT29-MTX co-culture model mimicking variable permeabilities of the human intestine obtained by an original seeding procedure. Eur. J. Pharm. Biopharm. 2014, 87, 290–298. [Google Scholar] [CrossRef]
- Smit, B.; Kuballa, A.; Coulson, S.; Katouli, M. Interaction of Candida albicans with human gut epithelium in the presence of Live Biotherapeutic Products (LBPs). Microbiol. Aust. 2021, 42, 120–124. [Google Scholar] [CrossRef]
- Timmerman, H.M.; Niers, L.E.M.; Ridwan, B.U.; Koning, C.J.M.; Mulder, L.; Akkermans, L.M.A.; Rombouts, F.M.; Rijkers, G.T. Design of a multispecies probiotic mixture to prevent infectious complications in critically ill patients. Clin. Nutr. 2007, 26, 450–459. [Google Scholar] [CrossRef]
 and in the presence of the Lactobacillus strains when co-inoculated
and in the presence of the Lactobacillus strains when co-inoculated  and pre-inoculated
and pre-inoculated  . (b) The number (mean ± SEM) of adhering AIEC alone and in the presence of the Lactobacillus strains when co-inoculated and pre-inoculated . E. coli strain 46.4 used as negative control
. (b) The number (mean ± SEM) of adhering AIEC alone and in the presence of the Lactobacillus strains when co-inoculated and pre-inoculated . E. coli strain 46.4 used as negative control  . * p < 0.0001 co-inoculation versus pre-inoculation ** p < 0.0001 combination versus individual strain.
and in the presence of the Lactobacillus strains when co-inoculated and pre-inoculated . (b) The number (mean ± SEM) of adhering AIEC alone and in the presence of the Lactobacillus strains when co-inoculated and pre-inoculated . E. coli strain 46.4 used as negative control . * p < 0.0001 co-inoculation versus pre-inoculation ** p < 0.0001 combination versus individual strain.
. * p < 0.0001 co-inoculation versus pre-inoculation ** p < 0.0001 combination versus individual strain.
and in the presence of the Lactobacillus strains when co-inoculated and pre-inoculated . (b) The number (mean ± SEM) of adhering AIEC alone and in the presence of the Lactobacillus strains when co-inoculated and pre-inoculated . E. coli strain 46.4 used as negative control . * p < 0.0001 co-inoculation versus pre-inoculation ** p < 0.0001 combination versus individual strain.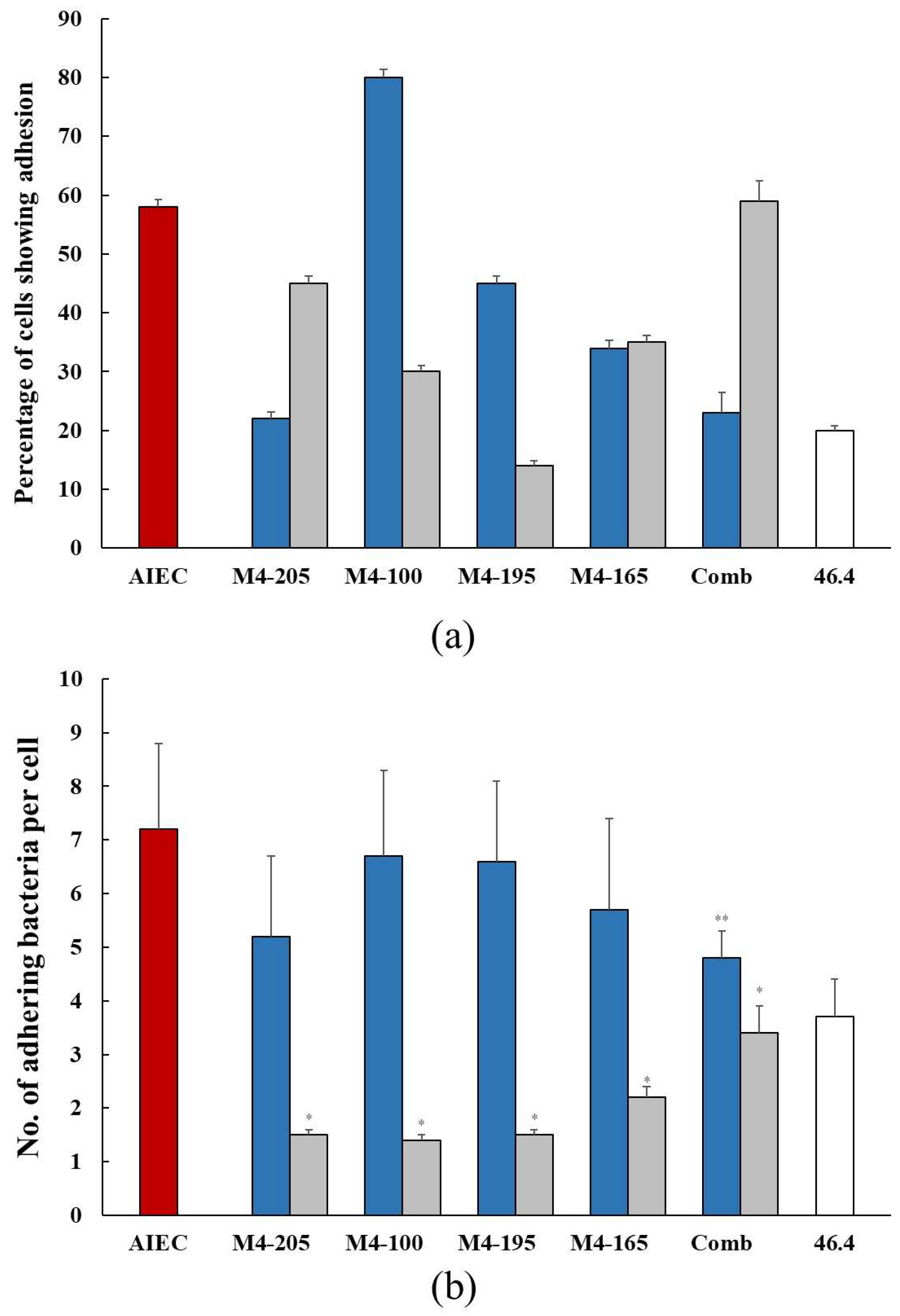 and in the presence of Lactobacillus strains when co-inoculated and pre-inoculated . E. coli strain 46.4 used as negative control . * p < 0.0001 co-inoculation versus pre-inoculation.
and in the presence of Lactobacillus strains when co-inoculated and pre-inoculated . E. coli strain 46.4 used as negative control . * p < 0.0001 co-inoculation versus pre-inoculation.
and in the presence of Lactobacillus strains when co-inoculated and pre-inoculated . E. coli strain 46.4 used as negative control . * p < 0.0001 co-inoculation versus pre-inoculation.
and in the presence of Lactobacillus strains when co-inoculated and pre-inoculated . E. coli strain 46.4 used as negative control . * p < 0.0001 co-inoculation versus pre-inoculation.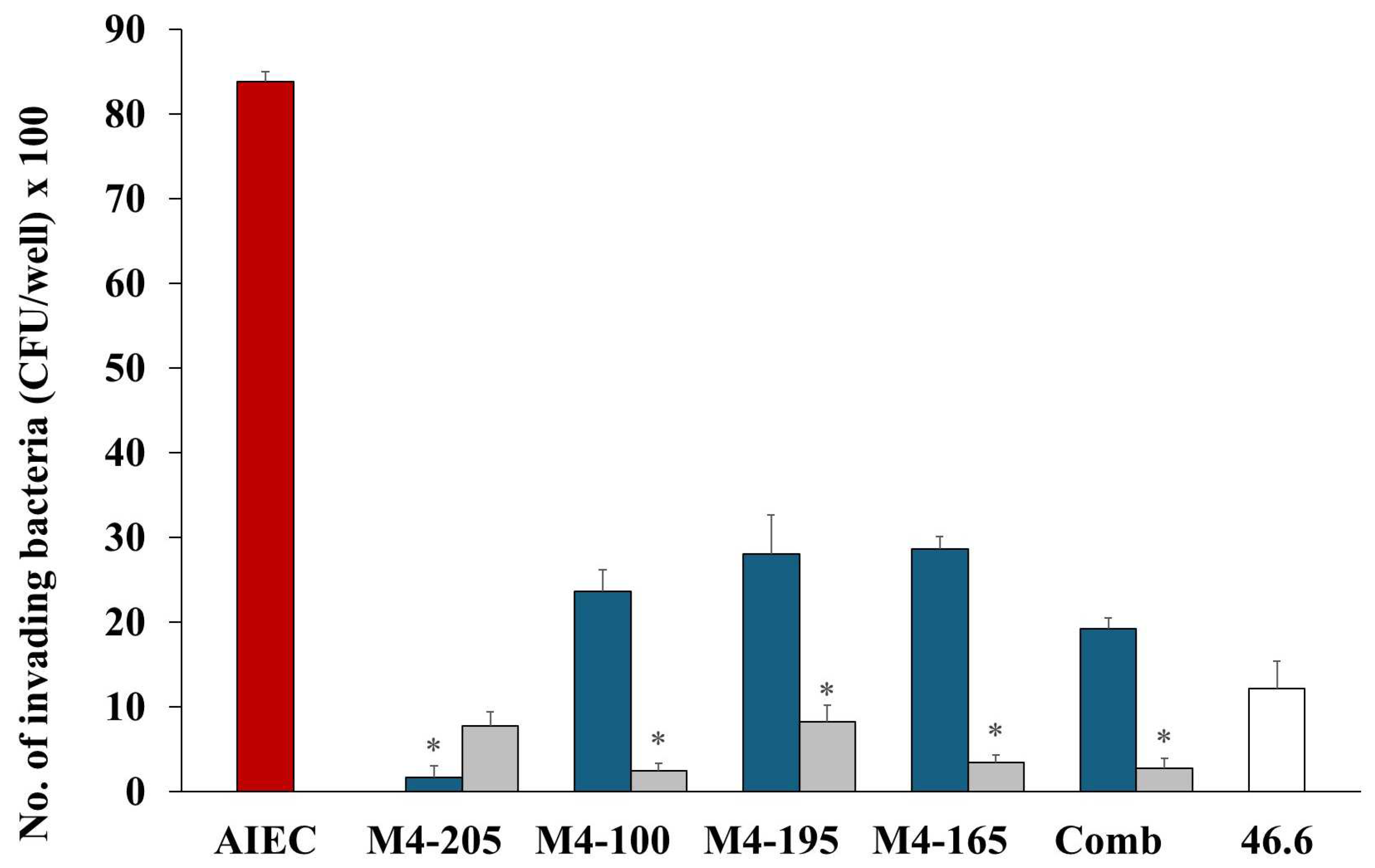 , when in the presence of Lactobacillus strains M4-205 , M4-100 , M4-195
, when in the presence of Lactobacillus strains M4-205 , M4-100 , M4-195  , M4-165
, M4-165  , and their combination
, and their combination  . E. coli strain HMLN1 was used a positive control
. E. coli strain HMLN1 was used a positive control  and E. coli strain JM109 as a negative control . (a) Co-inoculation of AIEC and probiotic candidates after 30 min interaction; (b) co-inoculation of AIEC and probiotic candidates after 120 min interaction; (c) pre-inoculation of AIEC and probiotic candidates after 30 min interaction; (d) pre-inoculation of AIEC and probiotic candidates after 120 min interaction.
, when in the presence of Lactobacillus strains M4-205 , M4-100 , M4-195 , M4-165 , and their combination . E. coli strain HMLN1 was used a positive control and E. coli strain JM109 as a negative control . (a) Co-inoculation of AIEC and probiotic candidates after 30 min interaction; (b) co-inoculation of AIEC and probiotic candidates after 120 min interaction; (c) pre-inoculation of AIEC and probiotic candidates after 30 min interaction; (d) pre-inoculation of AIEC and probiotic candidates after 120 min interaction.
and E. coli strain JM109 as a negative control . (a) Co-inoculation of AIEC and probiotic candidates after 30 min interaction; (b) co-inoculation of AIEC and probiotic candidates after 120 min interaction; (c) pre-inoculation of AIEC and probiotic candidates after 30 min interaction; (d) pre-inoculation of AIEC and probiotic candidates after 120 min interaction.
, when in the presence of Lactobacillus strains M4-205 , M4-100 , M4-195 , M4-165 , and their combination . E. coli strain HMLN1 was used a positive control and E. coli strain JM109 as a negative control . (a) Co-inoculation of AIEC and probiotic candidates after 30 min interaction; (b) co-inoculation of AIEC and probiotic candidates after 120 min interaction; (c) pre-inoculation of AIEC and probiotic candidates after 30 min interaction; (d) pre-inoculation of AIEC and probiotic candidates after 120 min interaction.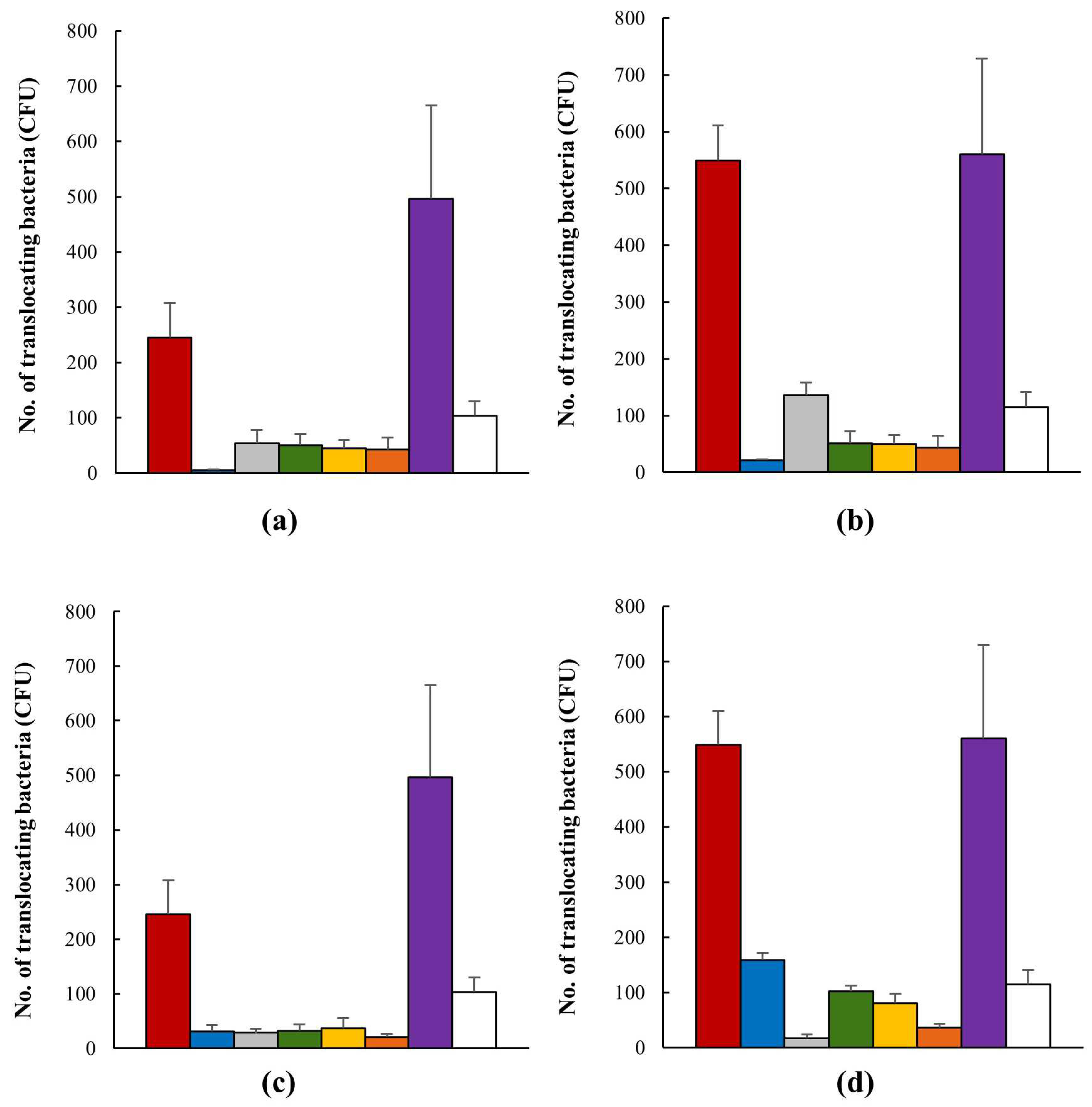

{kind=link}
{kind=link}
{kind=link}
| Probiotic Candidates | Percentage of Reduction | |||||
|---|---|---|---|---|---|---|
| Colonisation | Adhesion | Invasion | ||||
| Co- Inoculation | Pre- Inoculation | Co- Inoculation | Pre- Inoculation | Co- Inoculation | Pre- Inoculation | |
| Lactobacillus M4-205 | 36 | 13 | 28 | 79 | 98 | 91 |
| Lactobacillus M4-100 | -22 | 28 | 8 | 80 | 72 | 97 |
| Lactobacillus M4-195 | 13 | 44 | 9 | 80 | 67 | 90 |
| Lactobacillus M4-165 | 24 | 23 | 21 | 70 | 66 | 96 |
| Combination of four probiotics | 35 | −1 | 34 | 53 | 77 | 97 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bradford, G.; Asgari, B.; Smit, B.; Hatje, E.; Kuballa, A.; Katouli, M. The Efficacy of Selected Probiotic Strains and Their Combination to Inhibit the Interaction of Adherent-Invasive Escherichia coli (AIEC) with a Co-Culture of Caco-2:HT29-MTX Cells. Microorganisms 2024, 12, 502. https://doi.org/10.3390/microorganisms12030502
Bradford G, Asgari B, Smit B, Hatje E, Kuballa A, Katouli M. The Efficacy of Selected Probiotic Strains and Their Combination to Inhibit the Interaction of Adherent-Invasive Escherichia coli (AIEC) with a Co-Culture of Caco-2:HT29-MTX Cells. Microorganisms. 2024; 12(3):502. https://doi.org/10.3390/microorganisms12030502
Chicago/Turabian StyleBradford, Georgia, Behnoush Asgari, Bronwyn Smit, Eva Hatje, Anna Kuballa, and Mohammad Katouli. 2024. "The Efficacy of Selected Probiotic Strains and Their Combination to Inhibit the Interaction of Adherent-Invasive Escherichia coli (AIEC) with a Co-Culture of Caco-2:HT29-MTX Cells" Microorganisms 12, no. 3: 502. https://doi.org/10.3390/microorganisms12030502
APA StyleBradford, G., Asgari, B., Smit, B., Hatje, E., Kuballa, A., & Katouli, M. (2024). The Efficacy of Selected Probiotic Strains and Their Combination to Inhibit the Interaction of Adherent-Invasive Escherichia coli (AIEC) with a Co-Culture of Caco-2:HT29-MTX Cells. Microorganisms, 12(3), 502. https://doi.org/10.3390/microorganisms12030502