
Genetic and Phenotypic Characterization of Bacillus velezensis Strain BV379 for Human Probiotic Applications



Abstract
1. Introduction
2. Materials and Methods
2.1. Strain Isolation and Spore Preparation
2.2. Genome Sequencing and Annotation
2.3. Taxonomic Classification
2.4. In Silico Bacillus Toxin Screening
2.5. In Silico Biogenic Amine Screening
2.6. In Silico Secondary Metabolite Screening
2.7. In Silico Virulence Factor Screening
2.8. Antimicrobial Resistance Gene and Mobile Genetic Element Screening
2.9. Antibiotic Minimum Inhibitory Concentration (MIC) Assay
2.10. Blood Hemolysis Assay
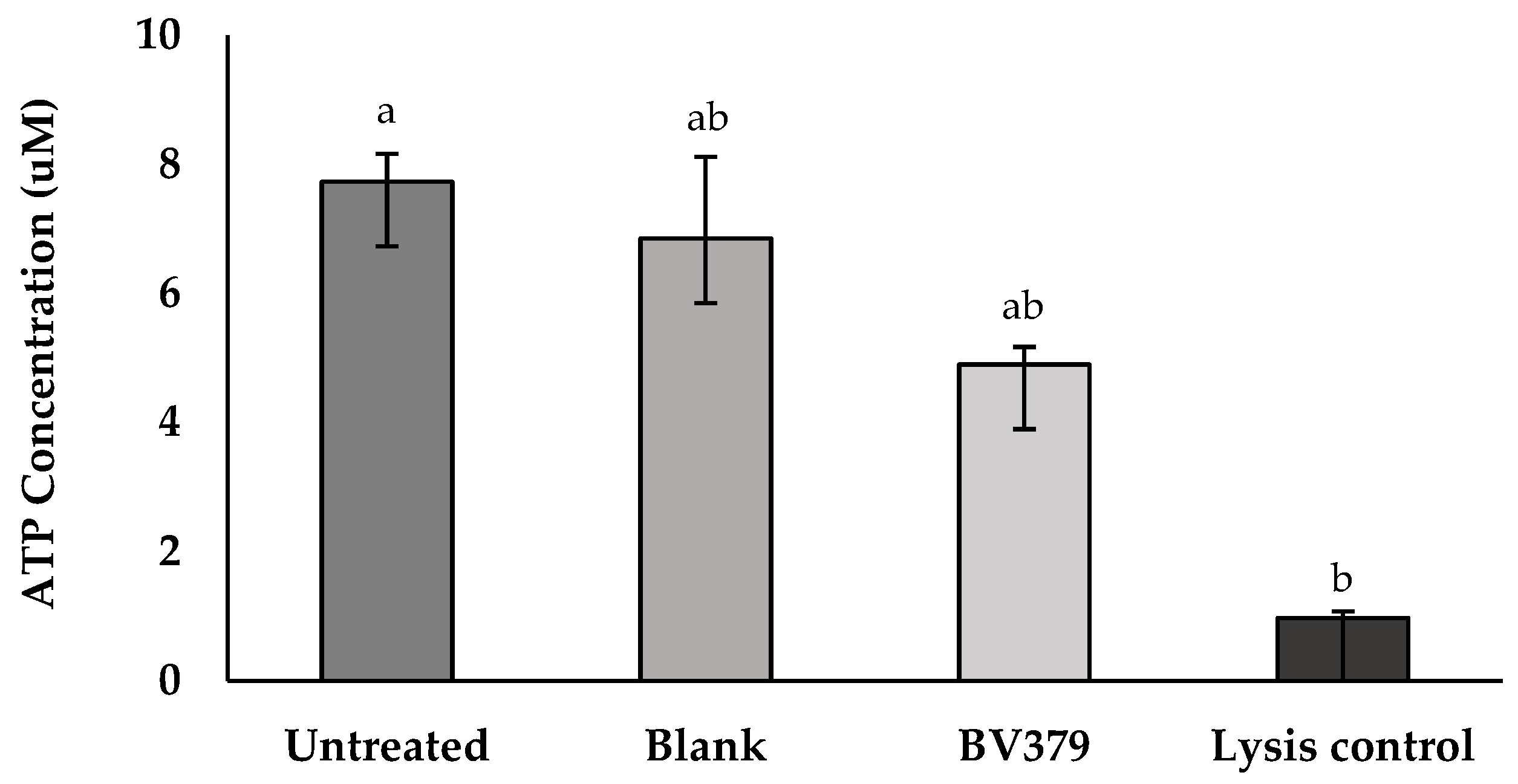
2.11. Caco-2 Cell Viability Assay
2.12. Caco-2 Cell Transepithelial Electrical Resistance (TEER) Assay
2.13. Antimicrobial Activity Assay
2.14. Carbohydrate Utilization Screening
2.15. Enzymatic Activity Screening
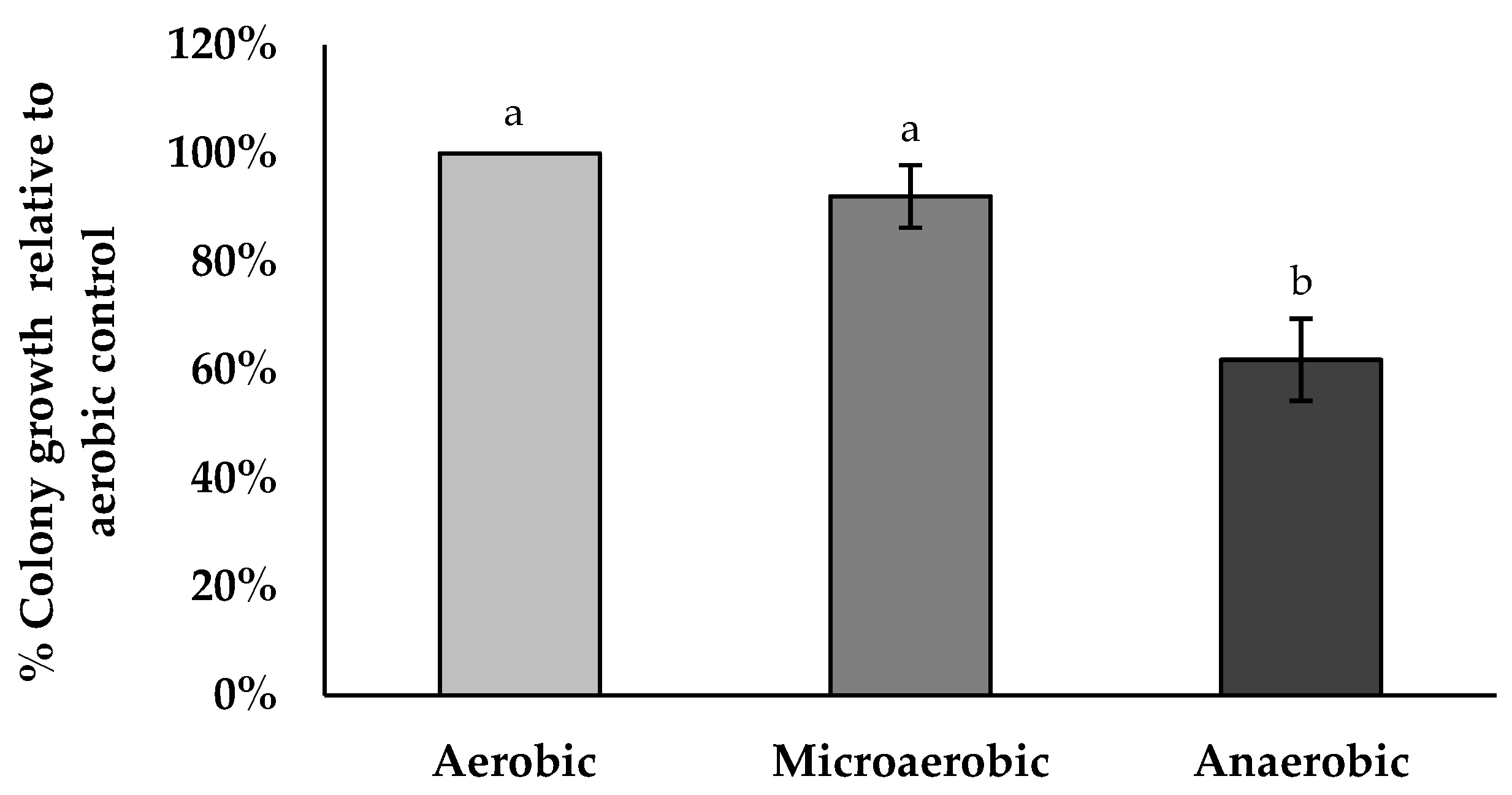
2.16. Anaerobic Tolerance Assay
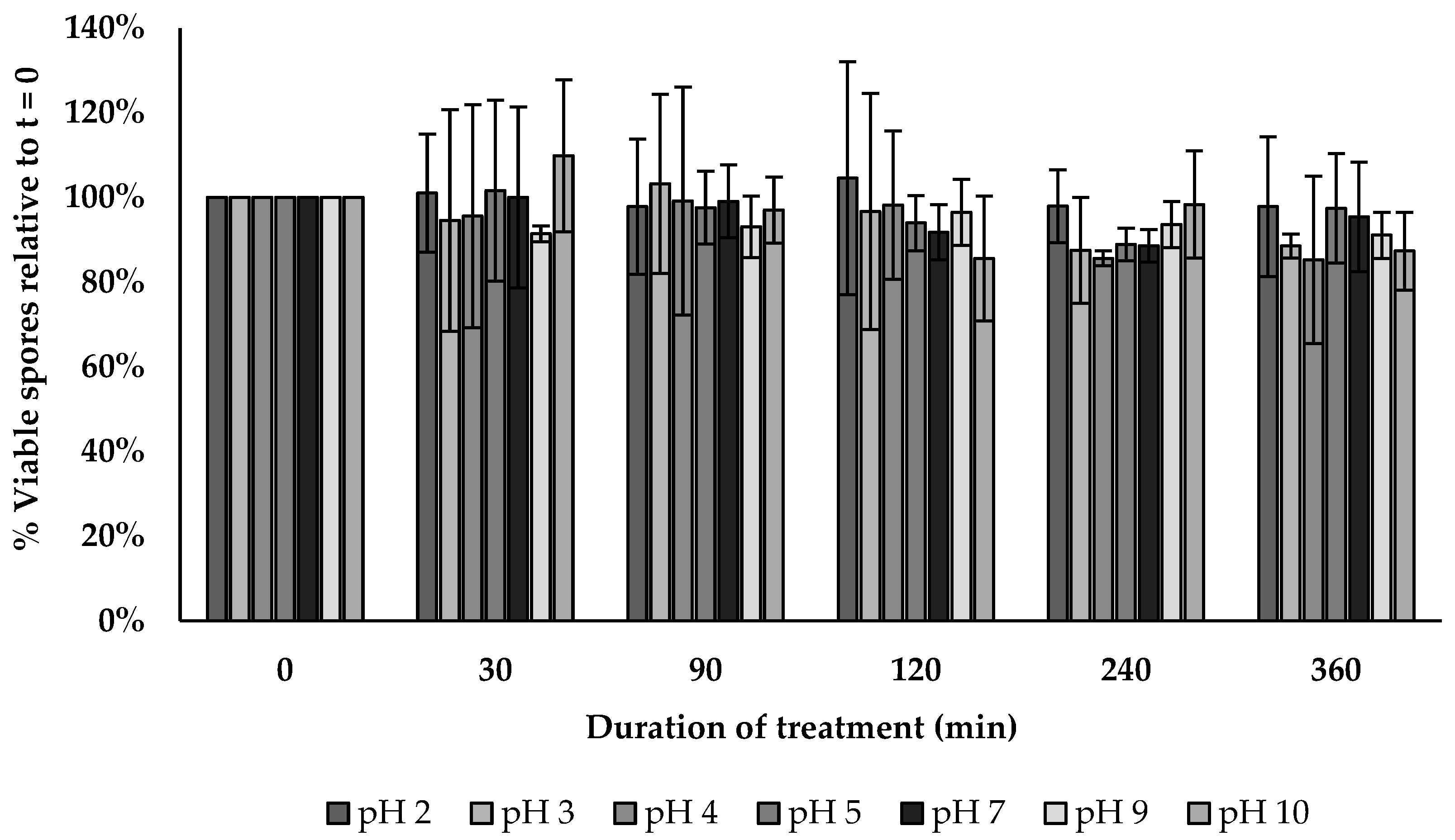
2.17. pH Tolerance Assay
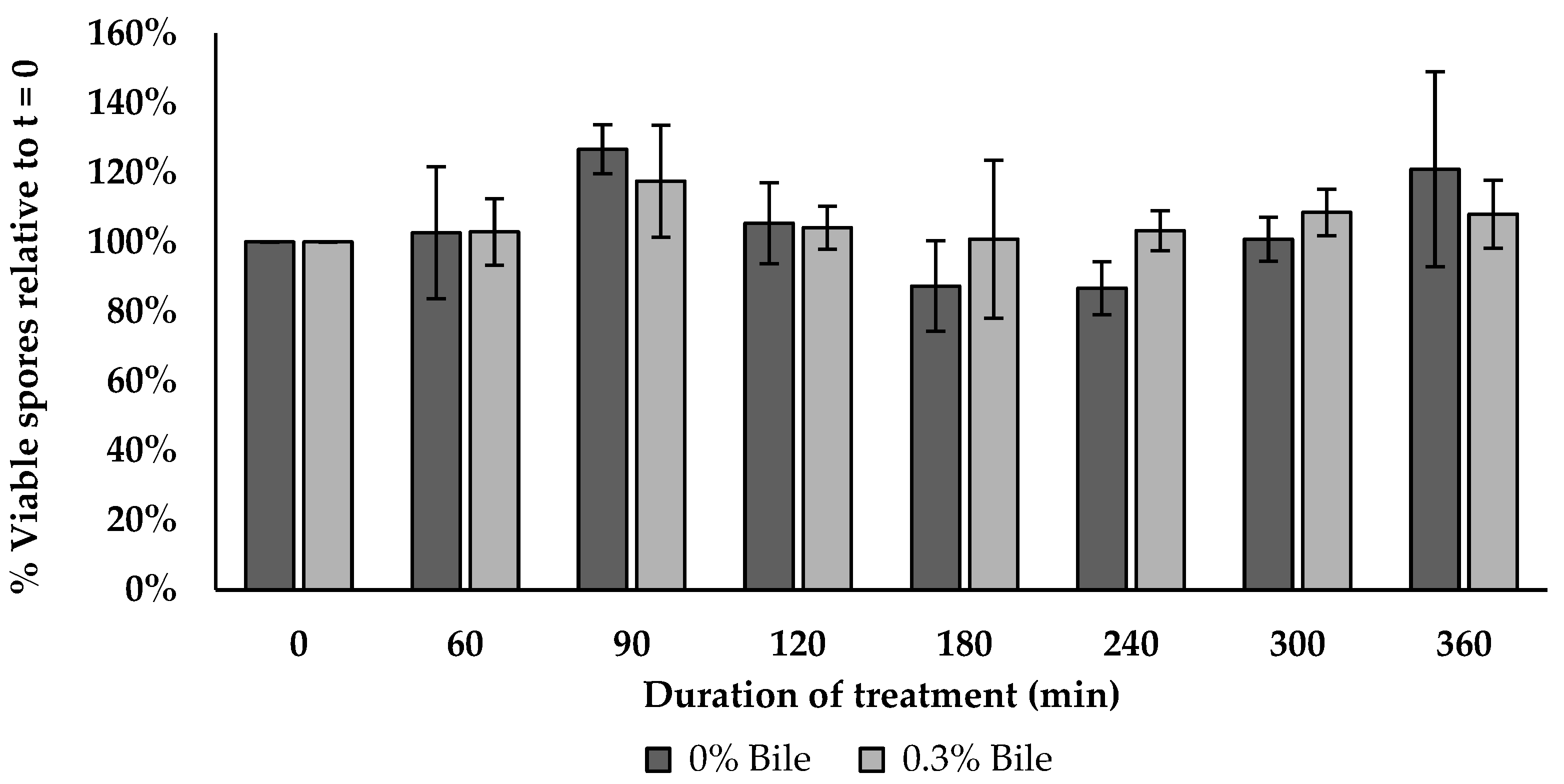
2.18. Bile Salt Tolerance Assay
2.19. Thermostability Assay
2.20. Statistical Analyses
3. Results
3.1. Taxonomic Classification
3.2. Bacillus Toxin Screening
3.3. Biogenic Amine Screening
3.4. Secondary Metabolite Screening
3.5. Virulence Factor Screening
3.6. Antimicrobial Resistance Gene and Mobile Genetic Element Assay
3.7. Antibiotic Minimum Inhibitory Concentration (MIC) Assay
3.8. Blood Hemolysis Assay
3.9. Caco-2 Cell Viability and Transepithelial Electrical Resistance (TEER) Assays
3.10. Antimicrobial Activity Assay
3.11. Carbohydrate Utilization Screening
3.12. Enzymatic Activity Screening
3.13. Anaerobic Tolerance Assay
3.14. pH Tolerance Assay
3.15. Bile Salt Tolerance Assay
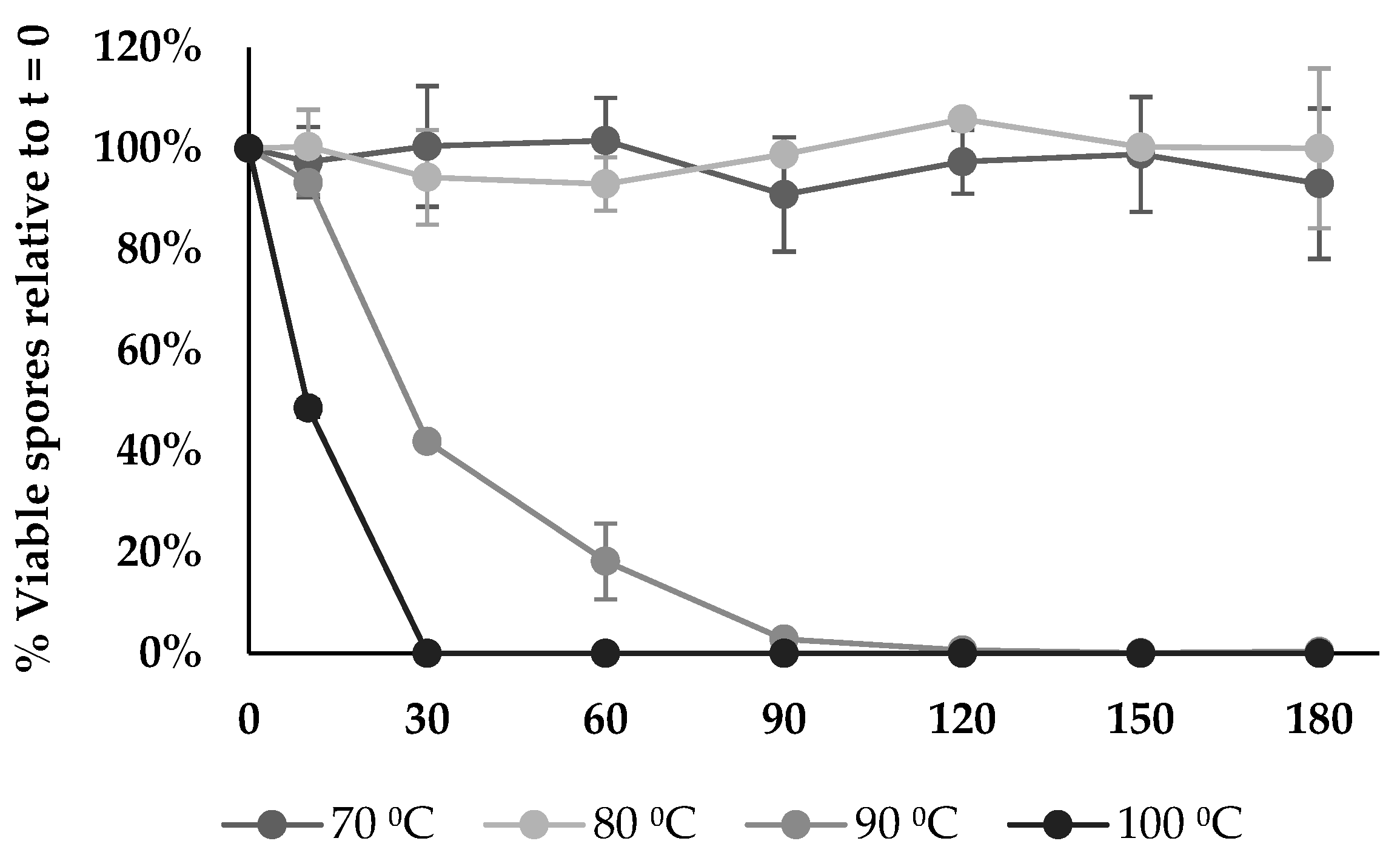
3.16. Thermostability Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harwood, C.R. Introduction to the Biotechnology of Bacillus. In Bacillus; Springer: Boston, MA, USA, 1989; pp. 1–4. [Google Scholar]
- Zhao, X.; Kuipers, O.P. Identification and Classification of Known and Putative Antimicrobial Compounds Produced by a Wide Variety of Bacillales Species. BMC Genom. 2016, 17, 882. [Google Scholar] [CrossRef]
- Grubbs, K.J.; Bleich, R.M.; Santa Maria, K.C.; Allen, S.E.; Farag, S.; Shank, E.A.; Bowers, A.A. Large-Scale Bioinformatics Analysis of Bacillus Genomes Uncovers Conserved Roles of Natural Products in Bacterial Physiology. mSystems 2017, 2, e00040-17. [Google Scholar] [CrossRef]
- Mandic-Mulec, I.; Stefanic, P.; van Elsas, J.D. Ecology of Bacillaceae. Microbiol Spectr. 2015, 3, TBS-0017-2013. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Liu, C.; Fang, H.; Zhang, D. Bacillus subtilis: A Universal Cell Factory for Industry, Agriculture, Biomaterials and Medicine. Microb. Cell Factories 2020, 19, 173. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Aguilar, J.G.; Sato, H.H. Microbial Proteases: Production and Application in Obtaining Protein Hydrolysates. Food Res. Int. 2018, 103, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Al-Dossary, A.; Hussain, M.; Setlow, P.; Li, J. Applications of Bacillus subtilis Spores in Biotechnology and Advanced Materials. Appl. Environ. Microbiol. 2020, 86, e01096-20. [Google Scholar] [CrossRef] [PubMed]
- Lajis, A.F.B. Biomanufacturing Process for the Production of Bacteriocins from Bacillaceae Family. Bioresour. Bioprocess. 2020, 7, 8. [Google Scholar] [CrossRef]
- Setlow, P. Spores of Bacillus subtilis: Their Resistance to and Killing by Radiation, Heat and Chemicals. J. Appl. Microbiol. 2006, 101, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Elshaghabee, F.M.F.; Rokana, N.; Gulhane, R.D.; Sharma, C.; Panwar, H. Bacillus as Potential Probiotics: Status, Concerns, and Future Perspectives. Front. Microbiol. 2017, 8, 1490. [Google Scholar] [CrossRef] [PubMed]
- Mingmongkolchai, S.; Panbangred, W. Bacillus Probiotics: An Alternative to Antibiotics for Livestock Production. J. Appl. Microbiol. 2018, 124, 1334–1346. [Google Scholar] [CrossRef]
- Cutting, S.M. Bacillus Probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of Action of Probiotics. Adv. Nutr. 2019, 10, S49–S66. [Google Scholar] [CrossRef] [PubMed]
- Del Toro-Barbosa, M.; Hurtado-Romero, A.; Garcia-Amezquita, L.E.; García-Cayuela, T. Psychobiotics: Mechanisms of Action, Evaluation Methods and Effectiveness in Applications with Food Products. Nutrients 2020, 12, 3896. [Google Scholar] [CrossRef] [PubMed]
- González-Rodríguez, I.; Ruiz, L.; Gueimonde, M.; Margolles, A.; Sánchez, B. Factors Involved in the Colonization and Survival of Bifidobacteria in the Gastrointestinal Tract. FEMS Microbiol. Lett. 2013, 340, 1–10. [Google Scholar] [CrossRef]
- Caillard, R.; Lapointe, N. In Vitro Gastric Survival of Commercially Available Probiotic Strains and Oral Dosage Forms. Int. J. Pharm. 2017, 519, 125–127. [Google Scholar] [CrossRef] [PubMed]
- Arslan, S.; Erbas, M.; Tontul, I.; Topuz, A. Microencapsulation of Probiotic Saccharomyces Cerevisiae Var. Boulardii with Different Wall Materials by Spray Drying. LWT-Food Sci. Technol. 2015, 63, 685–690. [Google Scholar] [CrossRef]
- Tam, N.K.M.; Uyen, N.Q.; Hong, H.A.; Duc, L.H.; Hoa, T.T.; Serra, C.R.; Henriques, A.O.; Cutting, S.M. The Intestinal Life Cycle of Bacillus subtilis and Close Relatives. J. Bacteriol. 2006, 188, 2692–2700. [Google Scholar] [CrossRef]
- Penet, C.; Kramer, R.; Little, R.; Spears, J.L.; Parker, J.; Iyer, J.K.; Guthrie, N.; Evans, M. A Randomized, Double-Blind, Placebo-Controlled, Parallel Study Evaluating the Efficacy of Bacillus subtilis MB40 to Reduce Abdominal Discomfort, Gas, and Bloating. Altern. Ther. Health Med. 2021, 27, 146–157. [Google Scholar]
- Garvey, S.M.; Mah, E.; Blonquist, T.M.; Kaden, V.N.; Spears, J.L. The Probiotic Bacillus subtilis BS50 Decreases Gastrointestinal Symptoms in Healthy Adults: A Randomized, Double-Blind, Placebo-Controlled Trial. Gut Microbes 2022, 14, 1. [Google Scholar] [CrossRef]
- Stecker, R.A.; Moon, J.M.; Russo, T.J.; Ratliff, K.M.; Mumford, P.W.; Jäger, R.; Purpura, M.; Kerksick, C.M. Bacillus Coagulans GBI-30, 6086 Improves Amino Acid Absorption from Milk Protein. Nutr. Metab. 2020, 17, 93. [Google Scholar] [CrossRef]
- Tarik, M.; Ramakrishnan, L.; Bhatia, N.; Goswami, R.; Kandasamy, D.; Roy, A.; Chandran, D.S.; Singh, A.; Upadhyay, A.D.; Kalaivani, M.; et al. The Effect of Bacillus Coagulans Unique IS-2 Supplementation on Plasma Amino Acid Levels and Muscle Strength in Resistance Trained Males Consuming Whey Protein: A Double-Blind, Placebo-Controlled Study. Eur. J. Nutr. 2022, 61, 2673–2685. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, T.; Xu, X.; Ahmarani, J. A Comprehensive Review of Post-Market Clinical Studies Performed in Adults with an Asian Probiotic Formulation. Benef. Microbes 2010, 1, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Hanifi, A.; Culpepper, T.; Mai, V.; Anand, A.; Ford, A.L.; Ukhanova, M.; Christman, M.; Tompkins, T.A.; Dahl, W.J. Evaluation of Bacillus subtilis R0179 on Gastrointestinal Viability and General Wellness: A Randomised, Double-Blind, Placebo-Controlled Trial in Healthy Adults. Benef. Microbes 2015, 6, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Horosheva, T.V.; Vodyanoy, V.; Sorokulova, I. Efficacy of Bacillus Probiotics in Prevention of Antibiotic-associated Diarrhoea: A Randomized, Double-blind, Placebo-controlled Clinical Trial. JMM Case Rep. 2014, 1, 1–6. [Google Scholar] [CrossRef]
- Kalman, D.S.; Schwartz, H.I.; Alvarez, P.; Feldman, S.; Pezzullo, J.C.; Krieger, D.R. A Prospective, Randomized, Double-Blind, Placebo-Controlled Parallel-Group Dual Site Trial to Evaluate the Effects of a Bacillus Coagulans-Based Product on Functional Intestinal Gas Symptoms. BMC Gastroenterol. 2009, 9, 85. [Google Scholar] [CrossRef] [PubMed]
- Maity, C.; Gupta, A.K. A Prospective, Interventional, Randomized, Double-Blind, Placebo-Controlled Clinical Study to Evaluate the Efficacy and Safety of Bacillus Coagulans LBSC in the Treatment of Acute Diarrhea with Abdominal Discomfort. Eur. J. Clin. Pharmacol. 2019, 75, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Spears, J.L.; Kramer, R.; Nikiforov, A.I.; Rihner, M.O.; Lambert, E.A. Safety Assessment of Bacillus subtilis MB40 for Use in Foods and Dietary Supplements. Nutrients 2021, 13, 733. [Google Scholar] [CrossRef]
- Majeed, M.; Nagabhushanam, K.; Paulose, S.; Arumugam, S.; Mundkur, L. The Effects of Bacillus Coagulans MTCC 5856 on Functional Gas and Bloating in Adults: A Randomized, Double-Blind, Placebo-Controlled Study. Medicine 2023, 102, e33109. [Google Scholar] [CrossRef]
- Kumar, V.; Sudha, K.; Bennur, S.; Dhanasekar, K. A Prospective, Randomized, Open-Label, Placebo-Controlled Comparative Study of Bacillus Coagulans GBI-30,6086 with Digestive Enzymes in Improving Indigestion in Geriatric Population. J. Fam. Med. Prim. Care 2020, 9, 1108. [Google Scholar] [CrossRef]
- Wauters, L.; Slaets, H.; De Paepe, K.; Ceulemans, M.; Wetzels, S.; Geboers, K.; Toth, J.; Thys, W.; Dybajlo, R.; Walgraeve, D.; et al. Efficacy and Safety of Spore-Forming Probiotics in the Treatment of Functional Dyspepsia: A Pilot Randomised, Double-Blind, Placebo-Controlled Trial. Lancet Gastroenterol. Hepatol. 2021, 6, 784–792. [Google Scholar] [CrossRef]
- Dolin, B.J. Effects of a Propietary Bacillus Coagulans Preparation on Symptoms of Diarrhea-Predominant Irritable Bowel Syndrome. Methods Find. Exp. Clin. Pharmacol. 2009, 31, 655. [Google Scholar] [CrossRef]
- Hun, L. Original Research: Bacillus Coagulans Significantly Improved Abdominal Pain and Bloating in Patients with IBS. Postgrad. Med. 2009, 121, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Majeed, M.; Nagabhushanam, K.; Natarajan, S.; Sivakumar, A.; Ali, F.; Pande, A.; Majeed, S.; Karri, S.K. Bacillus Coagulans MTCC 5856 Supplementation in the Management of Diarrhea Predominant Irritable Bowel Syndrome: A Double Blind Randomized Placebo Controlled Pilot Clinical Study. Nutr. J. 2016, 15, 21. [Google Scholar] [CrossRef] [PubMed]
- Madempudi, R.S.; Ahire, J.J.; Neelamraju, J.; Tripathi, A.; Nanal, S. Randomized Clinical Trial: The Effect of Probiotic Bacillus Coagulans Unique IS2 vs. Placebo on the Symptoms Management of Irritable Bowel Syndrome in Adults. Sci. Rep. 2019, 9, 12210. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Maity, C. Efficacy and Safety of Bacillus Coagulans LBSC in Irritable Bowel Syndrome: A Prospective, Interventional, Randomized, Double-Blind, Placebo-Controlled Clinical Study [CONSORT Compliant]. Medicine 2021, 100, e23641. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Kim, W.S.; Paik, H.D. Bacillus Strains as Human Probiotics: Characterization, Safety, Microbiome, and Probiotic Carrier. Food Sci. Biotechnol. 2019, 28, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Khatri, I.; Sharma, G.; Subramanian, S. Composite Genome Sequence of Bacillus clausii, a Probiotic Commercially Available as Enterogermina®, and Insights into Its Probiotic Properties. BMC Microbiol. 2019, 19, 307. [Google Scholar] [CrossRef]
- Ghelardi, E.; Abreu Y Abreu, A.T.; Marzet, C.B.; Calatayud, G.Á.; Perez, M.I.I.I.; Castro, A.P.M. Current Progress and Future Perspectives on the Use of Bacillus clausii. Microorganisms 2022, 10, 1246. [Google Scholar] [CrossRef]
- Ruiz-García, C.; Béjar, V.; Martínez-Checa, F.; Llamas, I.; Quesada, E. Bacillus velezensis Sp. Nov., a Surfactant-Producing Bacterium Isolated from the River Vélez in Málaga, Southern Spain. Int. J. Syst. Evol. Microbiol. 2005, 55, 191–195. [Google Scholar] [CrossRef]
- Ye, M.; Tang, X.; Yang, R.; Zhang, H.; Li, F.; Tao, F.; Li, F.; Wang, Z. Characteristics and Application of a Novel Species of Bacillus: Bacillus velezensis. ACS Chem. Biol. 2018, 13, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Rabbee, M.; Ali, M.; Choi, J.; Hwang, B.; Jeong, S.; Baek, K. Bacillus velezensis: A Valuable Member of Bioactive Molecules within Plant Microbiomes. Molecules 2019, 24, 1046. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Guan, X.; Zhong, X.; Zhou, X.; Yang, F. Bacillus velezensis DP -2 Isolated from Douchi and Its Application in Soybean Meal Fermentation. J. Sci. Food Agric. 2021, 101, 1861–1868. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Zhang, X.; Gao, Z.; Yang, Y.; Tian, X.; Lu, M.; He, L.; Li, C.; Zeng, X. Isolation of Bacillus Spp. with High Fibrinolytic Activity and Performance Evaluation in Fermented Douchi. J. Food Prot. 2021, 84, 717–727. [Google Scholar] [CrossRef]
- Na, H.-E.; Heo, S.; Kim, Y.-S.; Kim, T.; Lee, G.; Lee, J.-H.; Jeong, D.-W. The Safety and Technological Properties of Bacillus velezensis DMB06 Used as a Starter Candidate Were Evaluated by Genome Analysis. LWT 2022, 161, 113398. [Google Scholar] [CrossRef]
- Moon, S.; Park, Y.; Kim, I.; Chang, H. Isolation and Characterization of Bacillus velezensis SS360-1 from Seed Soy Sauce. Korean J. Community Living Sci. 2018, 29, 49–58. [Google Scholar] [CrossRef]
- Yang, H.; Liu, Y.; Ning, Y.; Wang, C.; Zhang, X.; Weng, P.; Wu, Z. Characterization of an Intracellular Alkaline Serine Protease from Bacillus velezensis SW5 with Fibrinolytic Activity. Curr. Microbiol. 2020, 77, 1610–1621. [Google Scholar] [CrossRef]
- Gil, N.-Y.; Jang, Y.-J.; Gwon, H.-M.; Jeong, W.-S.; Yeo, S.-H.; Kim, S.-Y. Comparative Evaluation of Quality and Metabolite Profiles in Meju Using Starter Cultures of Bacillus velezensis and Aspergillus oryzae. Foods 2021, 11, 68. [Google Scholar] [CrossRef]
- Yao, Z.; Kim, J.A.; Kim, J.H. Characterization of a Fibrinolytic Enzyme Secreted by Bacillus velezensis BS2 Isolated from Sea Squirt Jeotgal. J. Microbiol. Biotechnol. 2019, 29, 347–356. [Google Scholar] [CrossRef]
- Mi, X.J.; Tran, T.H.M.; Park, H.R.; Xu, X.Y.; Subramaniyam, S.; Choi, H.S.; Kim, J.; Koh, S.C.; Kim, Y.J. Immune-Enhancing Effects of Postbiotic Produced by Bacillus velezensis Kh2-2 Isolated from Korea Foods. Food Res. Int. 2022, 152, 110911. [Google Scholar] [CrossRef]
- Heo, S.; Kim, J.H.; Kwak, M.S.; Sung, M.H.; Jeong, D.W. Functional Annotation Genome Unravels Potential Probiotic Bacillus velezensis Strain KMU01 from Traditional Korean Fermented Kimchi. Foods 2021, 10, 563. [Google Scholar] [CrossRef]
- Heo, S.; Kim, J.H.; Kwak, M.S.; Jeong, D.W.; Sung, M.H. Draft Genome Sequence of Bacillus velezensis Strain KMUS1 from Kimchi. Korean J. Microbiol. 2021, 57, 122–124. [Google Scholar] [CrossRef]
- Liu, H.; Luo, S.; Liu, J.; Yan, Q.; Yang, S.; Jiang, Z. Novel Green Soybean Shuidouchi Fermented by Bacillus velezensis with Multibioactivities. Food Sci. Nutr. 2021, 9, 6538–6547. [Google Scholar] [CrossRef]
- Kathiresan, N.; Renuka, A.N.; Gopal, L.; Abraham, D.R. Preparation of Probiotic Chocolate and Jelly with Bacillus Sp. and Lactobacillus Sp. Isolated from Fermented Gooseberry (Phyllanthus emblica) and Indian Scramberry (Flacourtia jangomas). Asian J. Res. Biosci. 2021, 3, 191–202. [Google Scholar]
- Luo, Y.Y.; Guo, Y.; Hu, X.Y.; Liu, W.H.; Liu, B.Q.; Yang, J.; Tu, Z.C.; Huang, Y.H. Flavor Improvement of Fermented Soybean Foods by Co-Fermentation with Bacillus velezensis and Lactiplantibacillus plantarum. LWT 2023, 186, 115257. [Google Scholar] [CrossRef]
- Ge, Z.; Kuang, Z.; Chen, J.; Chen, J.; Liu, T.; She, Z.; Lu, Y. Comparative Genomics Analysis of Bacillus velezensis LOH112 Isolated from a Nonagenarian Provides Insights into Its Biocontrol and Probiotic Traits. Gene 2022, 835, 146644. [Google Scholar] [CrossRef]
- O’Donnell, M.M.; Hegarty, J.W.; Healy, B.; Schulz, S.; Walsh, C.J.; Hill, C.; Ross, R.P.; Rea, M.C.; Farquhar, R.; Chesnel, L. Identification of ADS024, a Newly Characterized Strain of Bacillus velezensis with Direct Clostridiodes Difficile Killing and Toxin Degradation Bio-Activities. Sci. Rep. 2022, 12, 9283. [Google Scholar] [CrossRef] [PubMed]
- Konaté, S.; Zgheib, R.; Camara, A.; Doumbo, O.; Djimdé, A.; Koné, A.K.; Théra, M.A.; Fournier, P.-E.; Tidjani Alou, M.; Raoult, D.; et al. Draft Genome Sequence of Bacillus velezensis Strain Marseille-Q1230, Isolated from a Stool Sample from a Severely Malnourished Child. Microbiol. Resour. Announc. 2021, 10, 16–18. [Google Scholar] [CrossRef] [PubMed]
- Vittoria, M.; Saggese, A.; Isticato, R.; Baccigalupi, L.; Ricca, E. Probiotics as an Alternative to Antibiotics: Genomic and Physiological Characterization of Aerobic Spore Formers from the Human Intestine. Microorganisms 2023, 11, 1978. [Google Scholar] [CrossRef] [PubMed]
- Adeniji, A.A.; Loots, D.T.; Babalola, O.O. Bacillus velezensis: Phylogeny, Useful Applications, and Avenues for Exploitation. Appl. Microbiol. Biotechnol. 2019, 103, 3669–3682. [Google Scholar] [CrossRef] [PubMed]
- Khalid, F.; Khalid, A.; Fu, Y.; Hu, Q.; Zheng, Y.; Khan, S.; Wang, Z. Potential of Bacillus velezensis as a Probiotic in Animal Feed: A Review. J. Microbiol. 2021, 59, 627–633. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel; Bampidis, V.; Azimonti, G.; Bastos, M.d.L.; Christensen, H.; Dusemund, B.; Kouba, M.; Fašmon Durjava, M.; López-Alonso, M.; López Puente, S.; et al. Safety and Efficacy of CorrelinkTM ABS1781 Bacillus subtilis (Bacillus velezensis NRRL B-67259) as a Feed Additive for All Growing Poultry Species. EFSA J. 2020, 18, e06279. [Google Scholar] [CrossRef]
- EFSA Panel; Bampidis, V.; Azimonti, G.; Bastos, M.d.L.; Christensen, H.; Dusemund, B.; Kouba, M.; Fašmon Durjava, M.; López-Alonso, M.; López Puente, S.; et al. Safety and Efficacy of Bacillus subtilis PB6 (Bacillus velezensis ATCC PTA-6737) as a Feed Additive for Chickens for Fattening, Chickens Reared for Laying, Minor Poultry Species (except for Laying Purposes), Ornamental, Sporting and Game Birds. EFSA J. 2020, 18, e06280. [Google Scholar] [CrossRef]
- EFSA Panel; Bampidis, V.; Azimonti, G.; Bastos, M.d.L.; Christensen, H.; Dusemund, B.; Fašmon Durjava, M.; Kouba, M.; López-Alonso, M.; López Puente, S.; et al. Safety and Efficacy of a Feed Additive Consisting of Bacillus velezensis DSM 15544 (Calsporin®) for Piglets (Suckling and Weaned), Pigs for Fattening, Sows in Order to Have Benefit in Piglets, Ornamental Fish, Dogs and All Avian Species (Asahi Biocycle Co.). EFSA J. 2021, 19, e06903. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel; Bampidis, V.; Azimonti, G.; Bastos, M.d.L.; Christensen, H.; Dusemund, B.; Fašmon Durjava, M.; Kouba, M.; López-Alonso, M.; López Puente, S.; et al. Safety and Efficacy of the Feed Additive Consisting of Bacillus velezensis CECT 5940 (Ecobiol®) for Turkeys for Fattening, Turkeys Reared for Breeding, Minor Poultry Species for Fattening and Reared for Laying and Ornamental Birds (Evonik Operations GmbH). EFSA J. 2021, 19, e06620. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel; Bampidis, V.; Azimonti, G.; Bastos, M.d.L.; Christensen, H.; Dusemund, B.; Fašmon Durjava, M.; Kouba, M.; López-Alonso, M.; López Puente, S.; et al. Safety and Efficacy of a Feed Additive Consisting of Bacillus velezensis PTA-6507, B. velezensis NRRL B-50013 and B. velezensis NRRL B-50104 (Enviva® PRO 202 GT) for Turkeys for Fattening (Danisco Animal Nutrition). EFSA J. 2021, 19, e06535. [Google Scholar] [CrossRef]
- EFSA Panel; Bampidis, V.; Azimonti, G.; Bastos, M.d.L.; Christensen, H.; Dusemund, B.; Kouba, M.; Fašmon Durjava, M.; López-Alonso, M.; López Puente, S.; et al. Safety and Efficacy of a Feed Additive Consisting of Bacillus velezensis NITE BP-01844 (BA-KING®) for Chickens for Fattening, Chickens Reared for Laying, Turkeys for Fattening, Turkeys Reared for Breeding and All Avian Species for Fattening, or Rearing To. EFSA J. 2022, 20, e07152. [Google Scholar] [CrossRef]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F.; Lindqvist, R.; et al. Update of the List of QPS-Recommended Biological Agents Intentionally Added to Food or Feed as Notified to EFSA 14: Suitability of Taxonomic Units Notified to EFSA until March 2021. EFSA J. 2021, 19, e06689. [Google Scholar] [CrossRef]
- Koutsoumanis, K.; Allende, A.; Álvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; de Cesare, A.; Hilbert, F.; Lindqvist, R.; Nauta, M.; et al. Update of the List of Qualified Presumption of Safety (QPS) Recommended Microorganisms Intentionally Added to Food or Feed as Notified to EFSA. EFSA J. 2023, 21, e07747. [Google Scholar] [CrossRef]
- EFSA Panel; Bampidis, V.; Azimonti, G.; Bastos, M.d.L.; Christensen, H.; Dusemund, B.; Fašmon Durjava, M.; Kouba, M.; López-Alonso, M.; López Puente, S.; et al. Assessment of the Application for Renewal of the Authorisation of Calsporin® (Bacillus velezensis DSM 15544) as a Feed Additive for Weaned Piglets. EFSA J. 2020, 18, 6283. [Google Scholar] [CrossRef]
- Jeong, H.; Kim, J.; Choi, S.-K.; Pan, J.-G. Genome Sequence of the Probiotic Strain Bacillus velezensis Variant Polyfermenticus GF423. Microbiol. Resour. Announc. 2018, 7, e01000-18. [Google Scholar] [CrossRef]
- Hatanaka, M.; Yamamoto, K.; Suzuki, N.; Iio, S.; Takara, T.; Morita, H.; Takimoto, T.; Nakamura, T. Effect of Bacillus subtilis C-3102 on Loose Stools in Healthy Volunteers. Benef. Microbes 2018, 9, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Hatanaka, M.; Kanzato, H.; Tsuda, R.; Nadaoka, I.; Yasue, M.; Hoshino, T.; Iio, S.-I.; Takara, T. Safety Evaluation of the Excessive Intake of Bacillus subtilis C-3102 in Healthy Japanese Adults: A Randomized, Placebo-Controlled, Double-Blind, Parallel-Group, Comparison Trial. Toxicol. Rep. 2020, 7, 46–58. [Google Scholar] [CrossRef]
- Takimoto, T.; Hatanaka, M.; Hoshino, T.; Takara, T.; Tanaka, K.; Shimizu, A.; Morita, H.; Nakamura, T. Effect of Bacillus subtilis C-3102 on Bone Mineral Density in Healthy Postmenopausal Japanese Women: A Randomized, Placebo-Controlled, Double-Blind Clinical Trial. Biosci. Microbiota Food Health 2018, 37, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-S.; Park, H.; Cho, I.-Y.; Paik, H.-D.; Park, E. Dietary Supplementation of Probiotic Bacillus Polyfermenticus, Bispan Strain, Modulates Natural Killer Cell and T Cell Subset Populations and Immunoglobulin G Levels in Human Subjects. J. Med. Food 2006, 9, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Brutscher, L.M.; Borgmeier, C.; Garvey, S.M.; Spears, J.L. Preclinical Safety Assessment of Bacillus subtilis BS50 for Probiotic and Food Applications. Microorganisms 2022, 10, 1038. [Google Scholar] [CrossRef] [PubMed]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of Long, Error-Prone Reads Using Repeat Graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef]
- Olson, R.D.; Assaf, R.; Brettin, T.; Conrad, N.; Cucinell, C.; Davis, J.J.; Dempsey, D.M.; Dickerman, A.; Dietrich, E.M.; Kenyon, R.W.; et al. Introducing the Bacterial and Viral Bioinformatics Resource Center (BV-BRC): A Resource Combining PATRIC, IRD and ViPR. Nucleic Acids Res. 2023, 51, D678–D689. [Google Scholar] [CrossRef]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A Modular and Extensible Implementation of the RAST Algorithm for Building Custom Annotation Pipelines and Annotating Batches of Genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef]
- Altschul, S.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief. Bioinform. 2018, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.; Peplies, J. JSpeciesWS: A Web Server for Prokaryotic Species Circumscription Based on Pairwise Genome Comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Mei, X.; Xu, K.; Yang, L.; Yuan, Z.; Mahillon, J.; Hu, X. The Genetic Diversity of Cereulide Biosynthesis Gene Cluster Indicates a Composite Transposon Tnces in Emetic Bacillus Weihenstephanensis. BMC Microbiol. 2014, 14, 149. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Fricker, M.; Grallert, H.; Rieck, P.; Wagner, M.; Scherer, S. Cereulide Synthetase Gene Cluster from Emetic Bacillus Cereus: Structure and Location on a Mega Virulence Plasmid Related to Bacillus anthracis Toxin Plasmid pXO1. BMC Microbiol. 2006, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving Cluster Detection and Comparison Capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: A General Classification Scheme for Bacterial Virulence Factors. Nucleic Acids Res. 2022, 50, D912–D917. [Google Scholar] [CrossRef]
- Pearson, W.R. An Introduction to Sequence Similarity (“Homology”) Searching. Curr. Protoc. Bioinform. 2013, 42, 1286–1292. [Google Scholar] [CrossRef]
- McArthur, A.G.; Waglechner, N.; Nizam, F.; Yan, A.; Azad, M.A.; Baylay, A.J.; Bhullar, K.; Canova, M.J.; De Pascale, G.; Ejim, L.; et al. The Comprehensive Antibiotic Resistance Database. Antimicrob. Agents Chemother. 2013, 57, 3348–3357. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic Resistome Surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Leplae, R.; Lima-Mendez, G.; Toussaint, A. ACLAME: A CLAssification of Mobile Genetic Elements, Update 2010. Nucleic Acids Res. 2009, 38, 57–61. [Google Scholar] [CrossRef]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The Reference Centre for Bacterial Insertion Sequences. Nucleic Acids Res. 2006, 34, 32–36. [Google Scholar] [CrossRef]
- Maamar, S.B.; Glawe, A.J.; Brown, T.K.; Hellgeth, N.; Hu, J.; Wang, J.P.; Huttenhower, C.; Hartmann, E.M. Mobilizable Antibiotic Resistance Genes Are Present in Dust Microbial Communities. PLoS Pathog. 2020, 16, e1008211. [Google Scholar] [CrossRef]
- Weinstein, M.P.; Patel, J.B.; Burnhman, C.-A.; ZImmer, B.L. M07 Methods for Dilution Antiests for Bacteria That Grow Aerobically; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.d.L.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; Gropp, J.; et al. Guidance on the Characterisation of Microorganisms Used as Feed Additives or as Production Organisms. EFSA J. 2018, 16, e05206. [Google Scholar] [CrossRef] [PubMed]
- EFSA. EFSA Panel on Additives and Products or Substances Used in Animal Feed (FEEDAP). Scientific Opinion Guidance on the Assessment of Bacterial Susceptibility to Antimicrobials of Human and Veterinary Importance. EFSA J. 2012, 10, 2740. [Google Scholar] [CrossRef]
- Zheng, J.; Ge, Q.; Yan, Y.; Zhang, X.; Huang, L.; Yin, Y. dbCAN3: Automated Carbohydrate-Active Enzyme and Substrate Annotation. Nucleic Acids Res. 2023, 51, W115–W121. [Google Scholar] [CrossRef] [PubMed]
- Dawson, P.A. Bile Secretion and the Enterohepatic Circulation of Bile Acids. In Sleisenger and Fordtran’s Gastrointestinal and Liver disease: Pathophysiology/Diagnosis/Management; W. B. Saunders, Co.: Philadelphia, PA, USA, 2014; pp. 1052–1063. [Google Scholar]
- Hempel, P.P.; Yao, M.; Yannarell, S.; Shevchenko, O.; Vogt, F.; Donofrio, N.; Maresca, J.A. Complete Genome Sequence of Bacillus velezensis Strain S4, Isolated from Biochar-Treated Soil. Microbiol. Resour. Announc. 2020, 9, 46–47. [Google Scholar] [CrossRef] [PubMed]
- Piewngam, P.; Khongthong, S.; Roekngam, N.; Theapparat, Y.; Sunpaweravong, S.; Faroongsarng, D.; Otto, M. Probiotic for Pathogen-Specific Staphylococcus aureus Decolonisation in Thailand: A Phase 2, Double-Blind, Randomised, Placebo-Controlled Trial. Lancet Microbe 2023, 4, e75–e83. [Google Scholar] [CrossRef] [PubMed]
- Konstantinidis, K.T.; Tiedje, J.M. Genomic Insights That Advance the Species Definition for Prokaryotes. Proc. Natl. Acad. Sci. USA 2005, 102, 2567–2572. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Rosselló-Móra, R. Shifting the Genomic Gold Standard for the Prokaryotic Species Definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [PubMed]
- Lücking, G.; Frenzel, E.; Rütschle, A.; Marxen, S.; Stark, T.D.; Hofmann, T.; Scherer, S.; Ehling-Schulz, M. Ces Locus Embedded Proteins Control the Non-Ribosomal Synthesis of the Cereulide Toxin in Emetic Bacillus Cereus on Multiple Levels. Front. Microbiol. 2015, 6, 1101. [Google Scholar] [CrossRef]
- Comas-Basté, O.; Sánchez-Pérez, S.; Veciana-Nogués, M.T.; Latorre-Moratalla, M.; Vidal-Carou, M.D.C. Histamine Intolerance: The Current State of the Art. Biomolecules 2020, 10, 1181. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, F.; Montanari, C.; Gardini, F.; Tabanelli, G. Biogenic Amine Production by Lactic Acid Bacteria: A Review. Foods 2019, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Bardócz, S. Polyamines in Food and Their Consequences for Food Quality and Human Health. Trends Food Sci. Technol. 1995, 6, 341–346. [Google Scholar] [CrossRef]
- Kalač, P.; Krausová, P. A Review of Dietary Polyamines: Formation, Implications for Growth and Health and Occurrence in Foods. Food Chem. 2005, 90, 219–230. [Google Scholar] [CrossRef]
- Soni, R.; Keharia, H.; Bose, A.; Pandit, N.; Doshi, J.; Rao, S.V.R.; Paul, S.S.; Raju, M.V.L.N. Genome Assisted Probiotic Characterization and Application of Bacillus velezensis ZBG17 as an Alternative to Antibiotic Growth Promoters in Broiler Chickens. Genomics 2021, 113, 4061–4074. [Google Scholar] [CrossRef]
- Devi, S.; Kiesewalter, H.T.; Kovács, R.; Frisvad, J.C.; Weber, T.; Larsen, T.O.; Kovács, Á.T.; Ding, L. Depiction of Secondary Metabolites and Antifungal Activity of Bacillus velezensis DTU001. Synth. Syst. Biotechnol. 2019, 4, 142–149. [Google Scholar] [CrossRef]
- Sharma, S.; Bhatnagar, R.; Gaur, D. Bacillus anthracis Poly-γ-d-Glutamate Capsule Inhibits Opsonic Phagocytosis by Impeding Complement Activation. Front. Immunol. 2020, 11, 462. [Google Scholar] [CrossRef]
- Kocianova, S.; Vuong, C.; Yao, Y.; Voyich, J.M.; Fischer, E.R.; DeLeo, F.R.; Otto, M. Key Role of Poly-γ-dl-Glutamic Acid in Immune Evasion and Virulence of Staphylococcus Epidermidis. J. Clin. Investig. 2005, 115, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Ijadi Bajestani, M.; Mousavi, S.M.; Mousavi, S.B.; Jafari, A.; Shojaosadati, S.A. Purification of Extra Cellular Poly-γ-Glutamic Acid as an Antibacterial Agent Using Anion Exchange Chromatography. Int. J. Biol. Macromol. 2018, 113, 142–149. [Google Scholar] [CrossRef]
- Bhat, A.R.; Irorere, V.U.; Bartlett, T.; Hill, D.; Kedia, G.; Morris, M.R.; Charalampopoulos, D.; Radecka, I. Bacillus subtilis Natto: A Non-Toxic Source of Poly-γ-Glutamic Acid That Could Be Used as a Cryoprotectant for Probiotic Bacteria. AMB Express 2013, 3, 36. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-E.; Rhee, J.-H.; Park, C.; Sung, M.-H.; Lee, I.-H. Distribution of Poly-Gamma-Glutamate Producers in Korean Fermented Foods, Cheongkukjang, Doenjang, and Kochujang. Food Sci. Biotechnol. 2005, 14, 704–708. [Google Scholar]
- Deng, F.; Chen, Y.; Sun, T.; Wu, Y.; Su, Y.; Liu, C.; Zhou, J.; Deng, Y.; Wen, J. Antimicrobial Resistance, Virulence Characteristics and Genotypes of Bacillus Spp. from Probiotic Products of Diverse Origins. Food Res. Int. 2021, 139, 109949. [Google Scholar] [CrossRef] [PubMed]
- Yelin, I.; Flett, K.B.; Merakou, C.; Mehrotra, P.; Stam, J.; Snesrud, E.; Hinkle, M.; Lesho, E.; McGann, P.; McAdam, A.J.; et al. Genomic and Epidemiological Evidence of Bacterial Transmission from Probiotic Capsule to Blood in ICU Patients. Nat. Med. 2019, 25, 1728–1732. [Google Scholar] [CrossRef]
- Kulkarni, T.; Majarikar, S.; Deshmukh, M.; Ananthan, A.; Balasubramanian, H.; Keil, A.; Patole, S. Probiotic Sepsis in Preterm Neonates—A Systematic Review. Eur. J. Pediatr. 2022, 181, 2249–2262. [Google Scholar] [CrossRef]
- Simonen, M.; Palva, I. Protein Secretion in Bacillus Species. Microbiol. Rev. 1993, 57, 109–137. [Google Scholar] [CrossRef]
- Harwood, C.R.; Cranenburgh, R. Bacillus Protein Secretion: An Unfolding Story. Trends Microbiol. 2008, 16, 73–79. [Google Scholar] [CrossRef]
- Clincial and Laboratory Standards Institute (CLSI). M02 Performance Standards for Antimicrobial Disk Susceptibility Tests, 13th ed.; Weinstein, M.P., Ed.; CLSI: Wayne, PA, USA, 2018; ISBN 1-56238-835-5. [Google Scholar]
- Yamamura, R.; Inoue, K.Y.; Nishino, K.; Yamasaki, S. Intestinal and Fecal pH in Human Health. Front. Microbiomes 2023, 2, 1192316. [Google Scholar] [CrossRef]
- Ruiz, L.; Margolles, A.; Sánchez, B. Bile Resistance Mechanisms in Lactobacillus and Bifidobacterium. Front. Microbiol. 2013, 4, 396. [Google Scholar] [CrossRef] [PubMed]
- Koopman, N.; Remijas, L.; Seppen, J.; Setlow, P.; Brul, S. Mechanisms and Applications of Bacterial Sporulation and Germination in the Intestine. Int. J. Mol. Sci. 2022, 23, 3405. [Google Scholar] [CrossRef] [PubMed]
- Colom, J.; Freitas, D.; Simon, A.; Brodkorb, A.; Buckley, M.; Deaton, J.; Deaton, J. Presence and Germination of the Probiotic Bacillus subtilis DE111® in the Human Small Intestinal Tract: A Randomized, Crossover, Double- Blind, and Placebo-Controlled Study. Front. Microbiol. 2021, 12, 715863. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, W.G.; Lowndes, R.H.; Young, H.L. Intraoperative Tissue Oximetry in the Human Gastrointestinal Tract. Am. J. Surg. 1990, 159, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Albenberg, L.; Esipova, T.V.; Judge, C.P.; Bittinger, K.; Chen, J.; Laughlin, A.; Grunberg, S.; Baldassano, R.N.; Lewis, J.D.; Li, H.; et al. Correlation between Intraluminal Oxygen Gradient and Radial Partitioning of Intestinal Microbiota. Gastroenterology 2014, 147, 1055–1063.e8. [Google Scholar] [CrossRef]
- Hartel, R.W.; Von Elbe, J.H.; Hofberger, R. Confectionery Science and Technology; Springer International Publishing: Cham, Switzerland, 2018; ISBN 978-3-319-61740-4. [Google Scholar]
- Tripathi, M.K.; Giri, S.K. Probiotic Functional Foods: Survival of Probiotics during Processing and Storage. J. Funct. Foods 2014, 9, 225–241. [Google Scholar] [CrossRef]
- International Dairy Foods Association. Available online: https://www.idfa.org/pasteurization (accessed on 1 February 2024).
- U.S. Department of Health and Human Services; National Institutes of Health; National Library of Medicine. The Effects of a Novel Probiotic on Gastrointestinal Health in Generally Healthy Adults. Available online: https://clinicaltrials.gov/study/NCT05595980 (accessed on 1 February 2024).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Organism | Accession | Max Score | e Value | % Identity |
|---|---|---|---|---|---|
| GatA | B. subtilis | NP_388550.1 | 6600 | 0 | 96.5 |
| MetG | B. cereus | WP_079994147.1 | 1019 | 0 | 74.3 |
| EntFM | B. cereus | AAX14641.1 | 112 | 3 × 10−26 | 54.6 |
| CesA | B. cereus | WP_002081542.1 | 968 | 0 | 35.5 |
| CesB | B. cereus | WP_000953496.1 | 653 | 0 | 34.8 |
| CesC | B. cereus | WP_000590108.1 | 139 | 3 × 10−36 | 32.6 |
| CesH | B. cereus | WP_000291846.1 | 32.7 | 8 × 10−1 | 20.9 |
| CesP | B. cereus | WP_000680399.1 | 128 | 6 × 10−33 | 34.4 |
| CesT | B. cereus | WP_000764755.1 | 60.8 | 1 × 10−17 | 30.7 |
| CesD | B. cereus | WP_001008264.1 | No significant similarity found | ||
| cytK | B. mycoides | AAW56196.1 | |||
| cytK | B. cereus | AAY84864.1 | |||
| NheA | B. mycoides | AAZ82480.1 | |||
| NheB | B. mycoides | AAZ82481.1 | |||
| NheC | B. mycoides | AAZ82482.1 | |||
| NheA | B. cereus | ABI52601.1 | |||
| NheB | B. cereus | ABI52602.1 | |||
| NheC | B. cereus | ABI52603.1 | |||
| NheA | B. cereus | CBL95107.1 | |||
| NheA, partial | B. thuringiensis | ACM18211.1 | |||
| NheB | B. thuringiensis | ACM18212.1 | |||
| NheC, partial | B. thuringiensis | ACM18213.1 | |||
| HblD | B. cereus | AFN08801.1 | |||
| HblC | B. cereus | AFN08807.1 | |||
| HblA | B. cereus | AII31101.1 | |||
| HblD | B. licheniformis | AIR07774.1 | |||
| HblA | B. licheniformis | AIR07775.1 | |||
| cytK | B. licheniformis | AIS75096.1 | |||
| Protein | Products | Species | Accession | Max Score | e Value | % Identity |
|---|---|---|---|---|---|---|
| Ornithine decarboxylase | Putrescine | P. wasatchensis | UHY45096.1 | 83.6 | 4 × 10−17 | 23.4% |
| Ornithine decarboxylase | Putrescine/ Cadaverine | B. amyloliquefaciens | MCG1032488.1 | 74.3 | 3 × 10−14 | 33.8% |
| Ornithine/ Lysine decarboxylase | Putrescine | P. wasatchensis | WP_044011228.1 | 69.7 | 6 × 10−13 | 27.2% |
| Lysine decarboxylase 1 | Putrescine | E. coli str. K-12 | NP_418555 | 82.8 | 6 × 10−17 | 25.3% |
| Lysine decarboxylase 2 | Cadaverine | E. coli str. K-12 | NP_414728 | 79.7 | 5 × 10−16 | 25.4% |
| Lysine decarboxylase | Cadaverine | B. amyloliquefaciens | MCG1033704.1 | 75.1 | 1 × 10−14 | 25.8% |
| Histidine decarboxylase | Histamine | L. parabuchneri | CUA78690.1 | No significant similarity found | ||
| Histidine decarboxylase | Histamine | B. licheniformis | BAJ05382.1 | |||
| Tyrosine decarboxylase | Tyramine | L. brevis | WP_021742084.1 | |||
| Tyrosine decarboxylase | Tyramine | B. cereus | WP_098989318.1 | |||
| Agmatine deiminase | Putrescine | L. brevis | ABS19477.1 | |||
| Agmatine deiminase | Putrescine | B. massiliogorillae | WP_042353033.1 | |||
| Cluster Type | Most Similar Cluster | % Identity |
|---|---|---|
| NRPS, Betalactone, Transat-PKS | Fengycin | 100% |
| Transat-PKS, NRPS-Like, NRPS, T3PKS | Bacillaene | 100% |
| Transat-PKS | Macrolactin H | 100% |
| Other | Bacilysin | 100% |
| Ripp-Like, NRPS | Bacillibactin | 100% |
| Transat-PKS | Difficidin | 93% |
| LAP, RRE-Containing | Plantazolicin | 91% |
| NRPS-Like, NRPS | Surfactin | 91% |
| PKS-Like | Butirosin A/Butirosin B | 7% |
| Terpene | - | - |
| Terpene | - | - |
| T3pks | - | - |
| ARO Term (Gene) | AMR Gene Family | Drug Class | % Identity | % Length | RGI Criteria |
|---|---|---|---|---|---|
| qacG | SMR antibiotic efflux pump | Disinfecting agents and antiseptics | 42.5 | 113.1 | Strict |
| qacJ | SMR antibiotic efflux pump | Disinfecting agents and antiseptics | 44.9 | 112.2 | Strict |
| qacJ | SMR antibiotic efflux pump | Disinfecting agents and antiseptics | 37.4 | 97.2 | Strict |
| qacJ | SMR antibiotic efflux pump | Disinfecting agents and antiseptics | 56.3 | 76.6 | Strict |
| clbA | Cfr 23S ribosomal RNAmethyltransferase | Lincosamide; Streptogramin; Streptogramin A; Oxazolidinone; Phenicol; Pleuromutilin | 99.7 | 100 | Strict |
| Antibiotics | Type | MIC (µg/mL) | EFSA MIC (µg/mL) Resistance Threshold a |
|---|---|---|---|
| Chloramphenicol | Phenicol | 1.00 | 8 |
| Clindamycin | Macrolides, lincosamides | 0.50 | 4 |
| Erythromycin | Macrolides, lincosamides | <0.0625 | 4 |
| Gentamicin | Aminoglycosides | <0.0625 | 4 |
| Kanamycin | Aminoglycoside | 0.500 | 8 |
| Streptomycin | Aminoglycoside | 3.906 | 8 |
| Oxytetracycline | Tetracycline | 0.125 | 8 |
| Vancomycin | Glycopeptide | 0.25 | 4 |
| Gram Reaction | Strains | Activity |
|---|---|---|
| Gram-positive | Bordetella bronchiseptica | ++ |
| Escherichia coli O157:H7 | + | |
| Pseudomonas aeruginosa | + | |
| Salmonella typhimurim | ++ | |
| Salmonella heidelberg | + | |
| Salmonella enterica subsp. enterica serovar Abaetetuba | + | |
| Gram-negative | Listeria monocytogenes | ++ |
| Staphylococcus aureus subsp. aureus | + | |
| Streptococcus agalactiae | + |
| Carbohydrate | Putative Genetic Mediator(s) |
|---|---|
| Amygdalin | N/A |
| Arbutin | arbF |
| D-Cellobiose | arbF, bglP, chbB |
| D-Fructose | SORD, xylA, ptf1, scrK |
| D-Glucose | pgm, pgi |
| D-Lactose (bovine origin) | araQ, araN, araP |
| D-Maltose | glva, MGAM |
| D-Mannitol | N/A |
| D-Raffinose | GLA, sacA, msmE, msmF, msmG |
| D-Ribose | rbsA, rbsB, rbsC |
| D-Saccharose (sucrose) | sacA, malL, MGAM |
| D-Sorbitol | SORD |
| D-Trehalose | treC |
| D-Xylose | xylA, lyxA, xylB |
| Esculin + ferric citrate | N/A |
| Glycerol | glpk, GCY1 |
| Glycogen | amyA |
| Inositol | iolG |
| L-Arabinose | AraA, AraB |
| Methyl-α-D-glucopyranoside | N/A |
| N-Acetylglucosamine | murA |
| Salicin | arbF |
| Starch (amidon) | amyA |
| Enzyme | Result | Gene |
|---|---|---|
| Alkaline phosphatase | + | Present |
| Esterase (C 4) | + | Present |
| Esterase lipase (C 8) | + | Present |
| Lipase (C 14) | − | N/A |
| Leucine arylamidase | + | Present |
| Valine arylamidase | − | Present |
| Cystine arylamidase | − | N/A |
| Trypsin | − | N/A |
| D-chymotrypsin | − | N/A |
| Acid phosphatase | + | Present |
| Naphthol-AS-BI-phosphohydrolase | + | N/A |
| α-Galactosidase | − | Present |
| β-Galactosidase (lactase) | − | Present |
| β-Glucuronidase | − | N/A |
| α-Glucosidase | + | Present |
| β-Glucosidase | − | Present |
| N-acetyl-β-glucosaminidase | − | N/A |
| α-Mannosidase | − | N/A |
| α-Fucosidase | − | N/A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brutscher, L.M.; Gebrechristos, S.; Garvey, S.M.; Spears, J.L. Genetic and Phenotypic Characterization of Bacillus velezensis Strain BV379 for Human Probiotic Applications. Microorganisms 2024, 12, 436. https://doi.org/10.3390/microorganisms12030436
Brutscher LM, Gebrechristos S, Garvey SM, Spears JL. Genetic and Phenotypic Characterization of Bacillus velezensis Strain BV379 for Human Probiotic Applications. Microorganisms. 2024; 12(3):436. https://doi.org/10.3390/microorganisms12030436
Chicago/Turabian StyleBrutscher, Laura M., Sebhat Gebrechristos, Sean M. Garvey, and Jessica L. Spears. 2024. "Genetic and Phenotypic Characterization of Bacillus velezensis Strain BV379 for Human Probiotic Applications" Microorganisms 12, no. 3: 436. https://doi.org/10.3390/microorganisms12030436
APA StyleBrutscher, L. M., Gebrechristos, S., Garvey, S. M., & Spears, J. L. (2024). Genetic and Phenotypic Characterization of Bacillus velezensis Strain BV379 for Human Probiotic Applications. Microorganisms, 12(3), 436. https://doi.org/10.3390/microorganisms12030436