
Molecular Characteristics of Colistin Resistance in Acinetobacter baumannii and the Activity of Antimicrobial Combination Therapy in a Tertiary Care Medical Center in Lebanon

 , ,
, ,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Source of Isolates
2.2. Identification of Isolates
2.3. Antimicrobial Susceptibility Testing
2.4. Antimicrobial Combination Therapy
2.5. Growth Rate Assay
2.6. Serial Passaging of Colistin-Resistant Isolates
2.7. Whole-Genome Sequencing (WGS)
2.8. Bioinformatics Analysis of the Isolates
3. Results
3.1. Antimicrobial Susceptibility Testing
3.2. Antimicrobial Combination Therapy
3.3. Serial Passaging of Colistin-Resistant Isolates
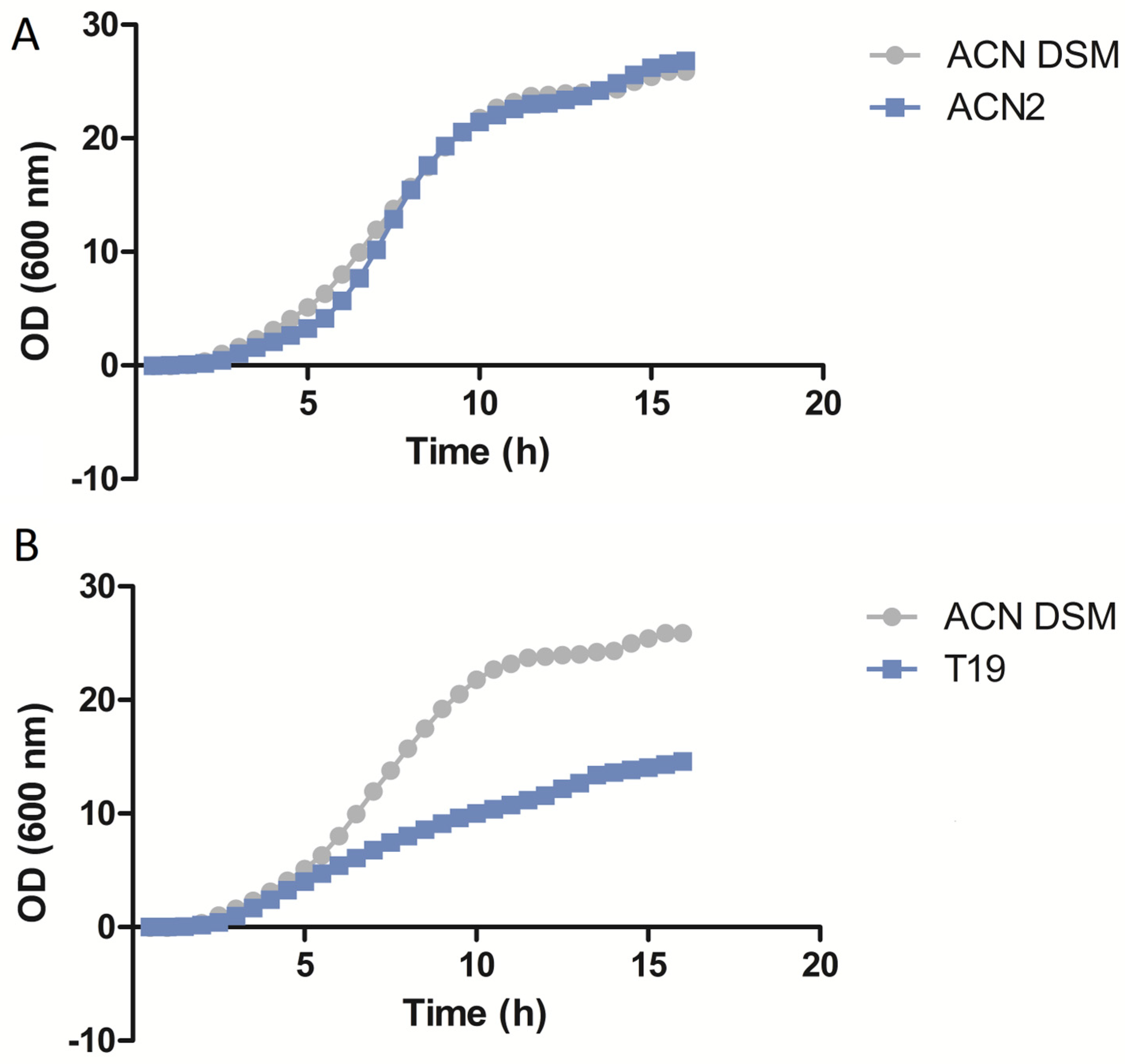
3.4. Growth Rate
3.5. Whole-Genome Sequencing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Logan, L.K.; Weinstein, R.A. The Epidemiology of Carbapenem-Resistant Enterobacteriaceae: The Impact and Evolution of a Global Menace. J. Infect. Dis. 2017, 215, S28–S36. [Google Scholar] [CrossRef]
- Sleiman, A.; Abou Fayad, A.G.; Banna, H.; Matar, G.M. Prevalence and Molecular Epidemiology of Carbapenem-Resistant Gram-Negative Bacilli and Their Resistance Determinants in the Eastern Mediterranean Region over the Last Decade. J. Glob. Antimicrob. Resist. 2021, 25, 209–221. [Google Scholar] [CrossRef]
- Fan, B.; Guan, J.; Wang, X.; Cong, Y. Activity of Colistin in Combination with Meropenem, Tigecycline, Fosfomycin, Fusidic Acid, Rifampin or Sulbactam against Extensively Drug-Resistant Acinetobacter Baumannii in a Murine Thigh-Infection Model. PLoS ONE 2016, 11, e0157757. [Google Scholar] [CrossRef]
- Nhu, N.T.K.; Riordan, D.W.; Nhu, T.D.H.; Thanh, D.P.; Thwaites, G.; Lan, N.P.H.; Wren, B.W.; Baker, S.; Stabler, R.A. The Induction and Identification of Novel Colistin Resistance Mutations in Acinetobacter Baumannii and Their Implications. Sci. Rep. 2016, 6, 28291. [Google Scholar] [CrossRef]
- Talat, A.; Miranda, C.; Poeta, P.; Khan, A.U. Farm to table: Colistin resistance hitchhiking through food. Arch. Microbiol. 2023, 205, 167. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.D.; Nickel, G.C.; Bajaksouzian, S.; Lavender, H.; Murthy, A.R.; Jacobs, M.R.; Bonomo, R.A. Resistance to Colistin in Acinetobacter Baumannii Associated with Mutations in the PmrAB Two-Component System. Antimicrob. Agents Chemother. 2009, 53, 3628–3634. [Google Scholar] [CrossRef]
- Moffatt, J.H.; Harper, M.; Harrison, P.; Hale, J.D.F.; Vinogradov, E.; Seemann, T.; Henry, R.; Crane, B.; St. Michael, F.; Cox, A.D.; et al. Colistin Resistance in Acinetobacter Baumannii Is Mediated by Complete Loss of Lipopolysaccharide Production. Antimicrob. Agents Chemother. 2010, 54, 4971–4977. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Petrosillo, N.; Ioannidou, E.; Falagas, M.E. Colistin Monotherapy vs. Combination Therapy: Evidence from Microbiological, Animal and Clinical Studies. Clin. Microbiol. Infect. 2008, 14, 816–827. [Google Scholar] [CrossRef]
- Viehman, J.A.; Nguyen, M.H.; Doi, Y. Treatment Options for Carbapenem-Resistant and Extensively Drug-Resistant Acinetobacter Baumannii Infections. Drugs 2014, 74, 1315–1333. [Google Scholar] [CrossRef]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF Mass Spectrometry: An Emerging Technology for Microbial Identification and Diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef]
- EM100 Connect—CLSI M100 ED29:2019. Available online: http://em100.edaptivedocs.net/GetDoc.aspx?doc=CLSIM100ED29:2019&xormat=SPDF&src=BB (accessed on 2 April 2020).
- Kanafani, Z.A.; Sleiman, A.; Frem, J.A.; Doumat, G.; Gharamti, A.; El Hafi, B.; Doumith, M.; AlGhoribi, M.F.; Kanj, S.S.; Araj, G.F.; et al. Molecular characterization and differential effects of levofloxacin and ciprofloxacin on the potential for developing quinolone resistance among clinical Pseudomonas aeruginosa isolates. Front. Microbiol. 2023, 14, 1209224. [Google Scholar] [CrossRef]
- Jain, S.N.; Vishwanatha, T.; Reena, V.; Divyashree, B.C.; Sampath, A.; Siddhalingeshwara, K.G.; Venugopal, N.; Ramesh, I. Antibiotic Synergy Test: Checkerboard Method on Multidrug Resistant Pseudomonas Aeruginosa. Int. Res. J. Pharm. 2011, 2, 196–198. [Google Scholar]
- Mu, X.; Wang, N.; Li, X.; Shi, K.; Zhou, Z.; Yu, Y.; Hua, X. The Effect of Colistin Resistance-Associated Mutations on the Fitness of Acinetobacter Baumannii. Front. Microbiol. 2016, 7, 1715. [Google Scholar] [CrossRef]
- Liao, W.; Lin, J.; Jia, H.; Zhou, C.; Zhang, Y.; Lin, Y.; Ye, J.; Cao, J.; Zhou, T. Resistance and Heteroresistance to Colistin in Escherichia Coli Isolates from Wenzhou, China. Infect. Drug Resist. 2020, 13, 3551–3561. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving Bacterial Genome Assemblies from Short and Long Sequencing Reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Wood, D.E.; Lu, J.; Langmead, B. Improved Metagenomic Analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of Acquired Antimicrobial Resistance Genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Naas, T.; Oueslati, S.; Bonnin, R.A.; Dabos, M.L.; Zavala, A.; Dortet, L.; Retailleau, P.; Iorga, B.I. Beta-Lactamase Database (BLDB)—Structure and Function. J. Enzym. Inhib. Med. Chem. 2017, 32, 917–919. [Google Scholar] [CrossRef]
- Gaiarsa, S.; Batisti Biffignandi, G.; Esposito, E.P.; Castelli, M.; Jolley, K.A.; Brisse, S.; Sassera, D.; Zarrilli, R. Comparative Analysis of the Two Acinetobacter Baumannii Multilocus Sequence Typing (MLST) Schemes. Front. Microbiol. 2019, 10, 930. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-Access Bacterial Population Genomics: BIGSdb Software, the PubMLST.Org Website and Their Applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Pormohammad, A.; Mehdinejadiani, K.; Gholizadeh, P.; Mohtavinejad, N.; Dadashi, M.; Karimaei, S.; Safari, H.; Azimi, T. Global Prevalence of Colistin Resistance in Clinical Isolates of Acinetobacter Baumannii: A Systematic Review and Meta-Analysis. Microb. Pathog. 2020, 139, 103887. [Google Scholar] [CrossRef]
- Chamoun, K.; Farah, M.; Araj, G.; Daoud, Z.; Moghnieh, R.; Salameh, P.; Saade, D.; Mokhbat, J.; Abboud, E.; Hamze, M.; et al. Surveillance of Antimicrobial Resistance in Lebanese Hospitals: Retrospective Nationwide Compiled Data. Int. J. Infect. Dis. 2016, 46, 64–70. [Google Scholar] [CrossRef]
- Hraiech, S.; Roch, A.; Lepidi, H.; Atieh, T.; Audoly, G.; Rolain, J.M.; Raoult, D.; Brunel, J.M.; Papazian, L.; Brégeon, F. Impaired Virulence and Fitness of a Colistin-Resistant Clinical Isolate of Acinetobacter Baumannii in a Rat Model of Pneumonia. Antimicrob. Agents Chemother. 2013, 57, 5120. [Google Scholar] [CrossRef]
- Gerson, S.; Betts, J.W.; Lucaßen, K.; Nodari, C.S.; Wille, J.; Josten, M.; Göttig, S.; Nowak, J.; Stefanik, D.; Roca, I.; et al. Investigation of Novel PmrB and EptA Mutations in Isogenic Acinetobacter Baumannii Isolates Associated with Colistin Resistance and Increased Virulence In Vivo. Antimicrob. Agents Chemother. 2019, 63, e01586-18. [Google Scholar] [CrossRef]
- Osman, M.; B Halimeh, F.; Rafei, R.; Mallat, H.; Tom, J.E.; Raad, E.B.; M Diene, S.; Jamal, S.; Al Atrouni, A.; Dabboussi, F.; et al. Investigation of an XDR-Acinetobacter Baumannii ST2 Outbreak in an Intensive Care Unit of a Lebanese Tertiary Care Hospital. Future Microbiol. 2020, 15, 1535–1542. [Google Scholar] [CrossRef]
- Nawfal Dagher, T.; Al-Bayssari, C.; Chabou, S.; Antar, N.; Diene, S.M.; Azar, E.; Rolain, J.M. Investigation of Multidrug-Resistant ST2 Acinetobacter Baumannii Isolated from Saint George Hospital in Lebanon. BMC Microbiol. 2019, 19, 29. [Google Scholar] [CrossRef]
- Higgins, P.G.; Hagen, R.M.; Kreikemeyer, B.; Warnke, P.; Podbielski, A.; Frickmann, H.; Loderstädt, U. Molecular Epidemiology of Carbapenem-Resistant Acinetobacter Baumannii Isolates from Northern Africa and the Middle East. Antibiotics 2021, 10, 291. [Google Scholar] [CrossRef]

{kind=link}
| Isolates | MIC (µg/mL) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Colistin (R ≥ 4 µg/mL) | Zerbaxa | Amikacin (R ≥ 64 µg/mL) | Bactrim (R ≥ 4/76 µg/mL) | Levofloxacin (R ≥ 8 µg/mL) | Azithromycin | Teicoplanin | Tigecyclin (R ≥ 0.5 µg/mL) | Meropenem (R ≥ 8 µg/mL) | Imipenem | Gentamycin | Cefipeme | Tazocin | Ceftazidime | Ciprofloxacin | Tetracyclin | |
| T11 | 4 | 64 | >4096 | 64 | 16 | 256 | 256 | 64 | 128 | 256/128 | >1024 | 256 | 512 | 128 | 64 | 1024 |
| T19 | 512 | 64 | 128 | 32 | 16 | 64/32 | 1024 | 64 | 32 | 128 | >1024 | 256 | 256 | 256 | 256/128 | 1024 |
| T20 | 4 | 128 | >4096 | 128 | 64 | >256 | 8 | 64 | 32 | 64/128 | >1024 | 512 | 512 | 512 | 128 | 1024 |
| T31 | 512 | 8 | 128 | 16 | 8 | 128 | 32 | 16 | 8 | 16 | 8 | 64 | 128 | 128 | 64 | 512 |
| T34 | 8 | 128 | >4096 | 64 | 32 | 256 | 256 | 64 | 128 | 128 | >1024 | 256 | 512 | 512/256 | 64 | 1024 |
| T36 | 512 | 128 | >4096 | 8 | 32 | 256 | 128 | 16 | 16 | 16 | >1024 | 128 | 256 | 512 | 128 | 1024 |
| T38 | 4 | 128 | >4096 | 128 | 32 | 256 | 128 | 64 | 128 | 128 | 1024 | 256 | 1024 | 512 | 64 | 1024 |
| T39 | 4 | 128 | >4096 | 64 | 32 | >256 | 256 | 32 | 256 | 128 | >1024 | 512 | 1024 | 512 | 64 | 1024 |
| T40 | 8 | 128 | >4096 | 64 | 32 | >256 | 256 | 32 | 128 | 64 | >1024 | 265 | 512 | 512 | 128 | 1024 |
| T47 | 4 | 512 | >4096 | 64 | 32 | >128 | 512 | 64 | 128 | 128 | >2048 | 512 | 256 | 512 | 64 | 1024 |
| T48 | 4 | 64 | >4096 | 64 | 32 | >128 | 512 | 64 | 128 | 64 | >2048 | 128 | 512 | 256 | 64 | 1024 |
| ACN1 | 512 | 128 | >4096 | 32 | 16 | 256 | 64 | 32 | 64 | 256 | >10,204 | 256 | 512 | 512 | 64 | 1024 |
| ACN2 | 64 | 16 | >4096 | 64 | 64 | >256 | 128 | 32 | 128 | 128 | >1024 | 128 | 512 | 256 | 128 | 1024 |
| ACN3 | 512 | 16 | >4096 | 32 | 32 | 8 | 128 | 64 | 8 | 16 | 2 | 128 | 256 | 256 | 128 | 1024 |
| ACN4 | 1 | 128 | >4096 | 1 | 64 | >256 | 256 | 64 | 128 | 64 | >1024 | 128 | 256 | 512 | 128 | |
| ACN5 | 512 | 16 | >4096 | 64 | 16 | >256 | 32 | 64 | 64 | 128 | 1024 | 256 | 512 | 512 | 64 | 1024 |
| Isolate | Aminoglycosides | Sulphon-Amides | Tetra-Cyclines | Macrolides | Beta-Lactams |
|---|---|---|---|---|---|
| T11 | aph(3″)-Ib; aph(6)-Id; armA; aph(3′)-Ia | sul2 | tet(B) | mph(E); msr(E) | OXA-66; OXA-23; ADC-25; TEM-1D |
| T19 | aph(3″)-Ib; aph(6)-Id; armA; aph(3′)-Ia | N/A | tet(B) | mph(E); msr(E) | OXA-66; OXA-23; ADC-25; TEM-1D |
| T20 | aph(3″)-Ib; aph(6)-Id; armA; aph(3′)-Ia | N/A | tet(B) | mph(E); msr(E) | OXA-66; OXA-23; ADC-25; TEM-1D |
| T31 | armA; aph(3′)-Ia | N/A | N/A | mph(E); msr(E) | OXA-66; blOXA-23; ADC-25; TEM-1D |
| T34 | aph(3″)-Ib; aph(6)-Id; armA; aph(3′)-Ia | N/A | tet(B) | mph(E); msr(E) | OXA-66; OXA-23; ADC-25; TEM-1D |
| T36 | aph(3″)-Ib; aph(6)-Id; armA; aph(3′)-Ia | N/A | tet(B) | mph(E); msr(E) | OXA-66; OXA-23 ADC-25; TEM-1D |
| T38 | aph(3″)-Ib; aph(6)-Id; armA; aph(3′)-Ia | N/A | tet(B) | mph(E); msr(E) | OXA-66; OXA-23; ADC-25; TEM-1D |
| T39 | aph(3″)-Ib; aph(6)-Id; aph(3′)-VIa | sul2 | tet(B) | N/A | OXA-66; OXA-23; ADC-25 |
| T40 | aph(3″)-Ib; aph(6)-Id; armA; aph(3′)-Ia | sul2 | tet(B) | mph(E); msr(E) | OXA-66; OXA-23; ADC-25; TEM-1D |
| T47 | armA; aph(3′)-Ia; aph(3″)-Ib; aph(6)-Id; aph(3′)-VIa | sul2 | tet(B) | mph(E); msr(E) | OXA-66; OXA-225; ADC-25; TEM-1D |
| T48 | aph(3″)-Ib; aph(6)-Id; armA; aph(3′)-Ia | sul2 | tet(B) | mph(E); msr(E) | OXA-66; OXA-23; ADC-25; TEM-1D |
| ACN1 | armA; aph(3′)-Ia; aph(3″)-Ib; aph(6)-Id; aph(3′)-VIa | sul2 | tet(B) | mph(E); msr(E) | OXA-66; OXA-23; ADC-25; TEM-1D |
| ACN2 | armA; aph(3′)-Ia; aph(3″)-Ib; aph(6)-Id; aph(3′)-VIa | sul2 | tet(B) | mph(E); msr(E) | OXA-66; OXA-23; ADC-25; TEM-1D |
| ACN3 | armA; aph(3′)-Ia; aph(3″)-Ib; aph(6)-Id; aph(3′)-VIa | sul2 | tet(B) | mph(E); msr(E) | OXA-66; OXA-23; ADC-25; TEM-1D |
| ACN5 | armA; aph(3′)-Ia; aph(3″)-Ib; aph(6)-Id; aph(3′)-VIa | sul2 | tet(B) | mph(E); msr(E) | OXA-66; OXA-23; ADC-25; TEM-1D |
| Isolate Code | Colistin MIC (µg/mL) | Resistance Determinant | Mutations |
|---|---|---|---|
| T11 | 4 | pmrC variant | G326A and R109H |
| T19 | 512 | - | - |
| T20 | 4 | pmrC variant | G326A and R109H |
| T31 | 512 | pmrC variant | C950T and A317V |
| T34 | 8 | pmrC variant | G326A and R109H |
| T36 | 512 | pmrC variant | G326A and R109H |
| T38 | 4 | pmrC variant | G326A and R109H |
| T39 | 4 | pmrC variant | G326A and R109H |
| T40 | 8 | pmrC variant | G326A and R109H |
| T47 | 4 | pmrC variant | G326A and R109H |
| T48 | 4 | pmrC variant | G326A and R109H |
| ACN1 | 512 | pmrC variant | G326A and R109H |
| ACN2 | 64 | pmrC variant | G326A and R109H |
| ACN3 | 512 | pmrC variant | G326A and R109H |
| ACN5 | 512 | pmrC variant | G326A and R109H |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abou Fayad, A.; Haraoui, L.-P.; Sleiman, A.; Hussein, H.; Grenier, F.; Derbaj, G.; Itani, D.; Iweir, S.; Sherri, N.; Bazzi, W.; et al. Molecular Characteristics of Colistin Resistance in Acinetobacter baumannii and the Activity of Antimicrobial Combination Therapy in a Tertiary Care Medical Center in Lebanon. Microorganisms 2024, 12, 349. https://doi.org/10.3390/microorganisms12020349
Abou Fayad A, Haraoui L-P, Sleiman A, Hussein H, Grenier F, Derbaj G, Itani D, Iweir S, Sherri N, Bazzi W, et al. Molecular Characteristics of Colistin Resistance in Acinetobacter baumannii and the Activity of Antimicrobial Combination Therapy in a Tertiary Care Medical Center in Lebanon. Microorganisms. 2024; 12(2):349. https://doi.org/10.3390/microorganisms12020349
Chicago/Turabian StyleAbou Fayad, Antoine, Louis-Patrick Haraoui, Ahmad Sleiman, Hadi Hussein, Frédéric Grenier, Ghada Derbaj, Dana Itani, Sereen Iweir, Nour Sherri, Wael Bazzi, and et al. 2024. "Molecular Characteristics of Colistin Resistance in Acinetobacter baumannii and the Activity of Antimicrobial Combination Therapy in a Tertiary Care Medical Center in Lebanon" Microorganisms 12, no. 2: 349. https://doi.org/10.3390/microorganisms12020349
APA StyleAbou Fayad, A., Haraoui, L.-P., Sleiman, A., Hussein, H., Grenier, F., Derbaj, G., Itani, D., Iweir, S., Sherri, N., Bazzi, W., Rasheed, S., Tanelian, A., Miari, M., el Hafi, B., Kanj, S. S., Kanafani, Z. A., Daoud, Z., Araj, G. F., & Matar, G. M. (2024). Molecular Characteristics of Colistin Resistance in Acinetobacter baumannii and the Activity of Antimicrobial Combination Therapy in a Tertiary Care Medical Center in Lebanon. Microorganisms, 12(2), 349. https://doi.org/10.3390/microorganisms12020349