
Microbial Community Establishment, Succession, and Temporal Dynamics in an Industrial Semi-Synthetic Metalworking Fluid Operation: A 50-Week Real-Time Tracking
Abstract
1. Introduction
2. Materials and Methods
2.1. Fluid Sampling
2.2. Quantification of Total Microbial Population
2.2.1. Microscopy
2.2.2. Culturing
2.2.3. Viable but Nonculturable (VBNC) Count
2.3. Targeted Analysis of Microbial Community
2.3.1. Selective Culturing and Identification Using PCR-Amplicon Sequencing (Targeted Community Composition Analysis)
2.3.2. Quantitative Real-Time PCR for Specific Genera/Groups (Targeted Community Dynamics)
2.4. Unbiased (PCR-DGGE) Analysis of Whole Community Composition and Dynamics
2.5. Endotoxin Analysis in MWF
3. Results and Discussion
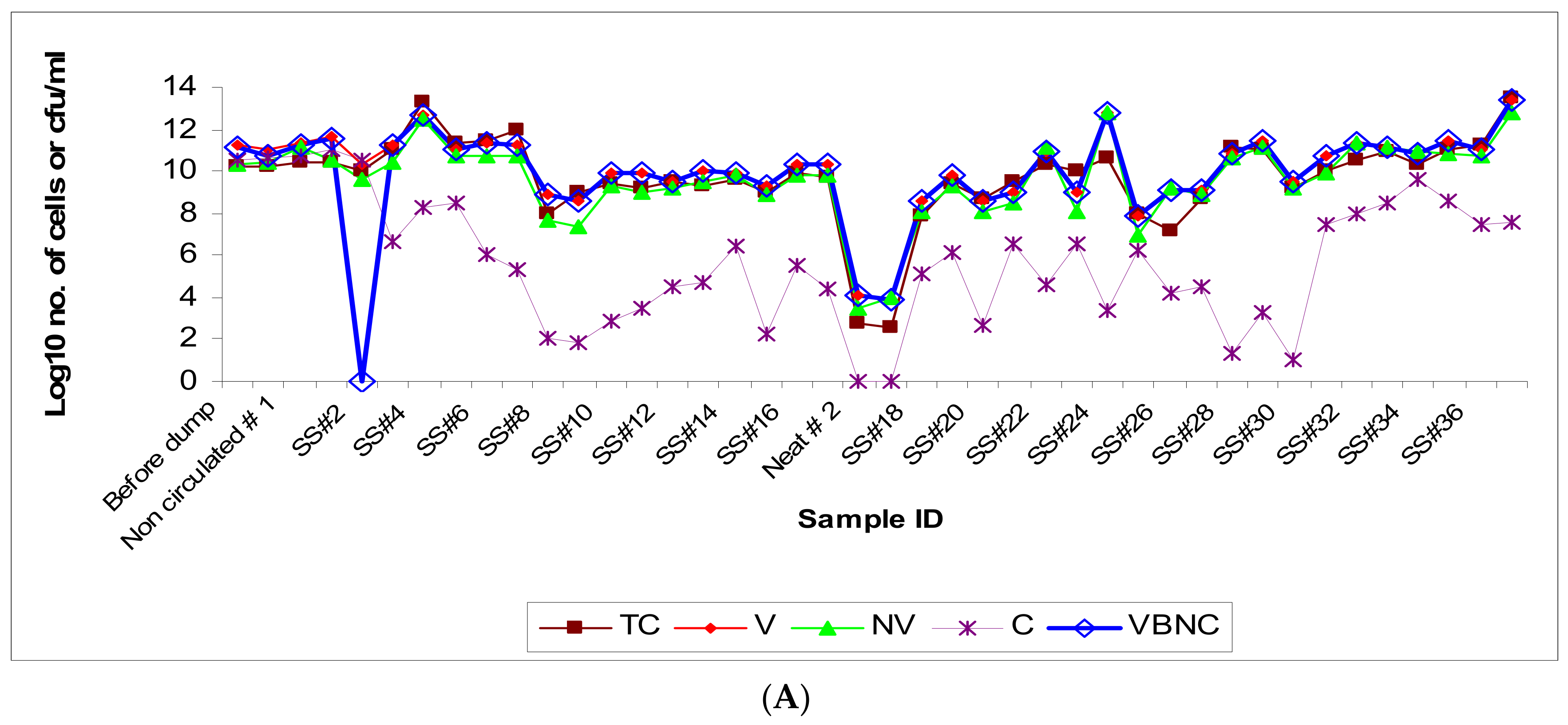
3.1. Microbial Population Dynamics (Viable versus Non-Viable)
3.2. Microbial Community Diversity (Composition and Dynamics)
3.2.1. Targeted Community Profile
Selective Culturing and DNA Sequencing
Real-Time qPCR and Amplicon Sequencing
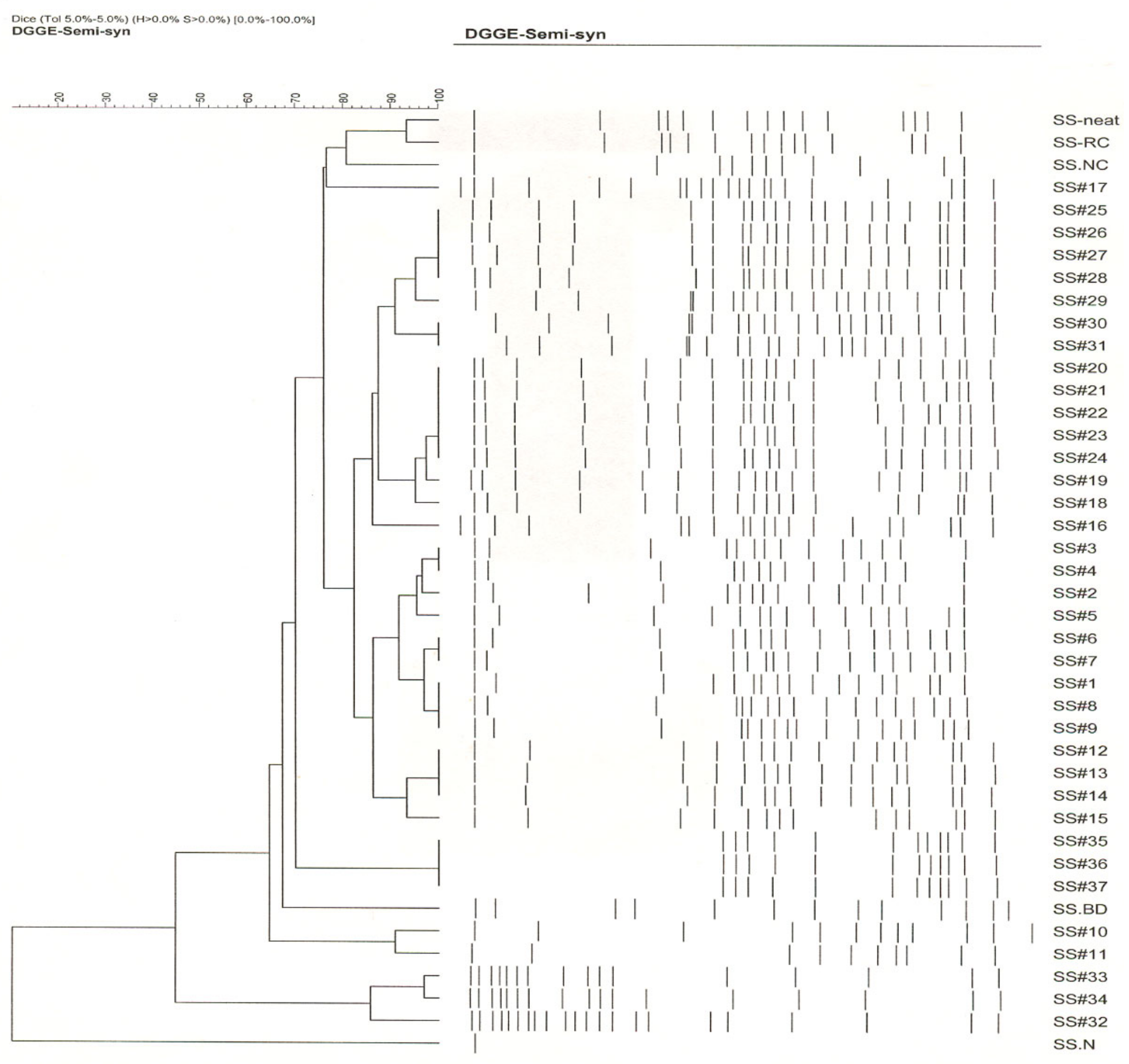
3.3. Whole Community Composition and Succession (Based on DGGE Analysis)
3.4. Endotoxin Build-Up and Dynamics
4. Study Limitations and Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robertson, A.S.; Weir, D.C.; Sherwood Burge, P. Occpational asthma due to oil mists. Thorax 1988, 43, 200–205. [Google Scholar] [CrossRef]
- National Institute of Occupational Safety and Health. Metalworking Fluids: Recommendation for Chronic Inhalation Studies; NIOSH: Cincinnati, OH, USA, 2001.
- Gauthier, S.L. Metalworking fluids: Oil mist and beyond. Appl. Occup. Environ. Hyg. 2003, 18, 818–824. [Google Scholar] [CrossRef]
- Saha, R.; Donofrio, R.S. The microbiology of metalworking fluids. Appl. Microbiol. Biotechnol. 2012, 94, 1119–1130. [Google Scholar] [CrossRef]
- Yan, L.; Yao, X.; Zhang, T.; Jiang, F.; Shui, Y.; Xie, H.; Xiang, Z.; Li, Y.; Lin, L. Passivation effect of the chlorinated paraffin added in the cutting fluid on the surface corrosion resistance of the stainless steel. Molecules 2023, 28, 3648. [Google Scholar] [CrossRef]
- Trafny, E.A. Microorganisms in metalworking fluids: Current issues in research and management. Int. J. Occup. Med. Environ. Health 2013, 26, 4–15. [Google Scholar] [CrossRef]
- Elansky, S.N.; Chudinova, E.M.; Elansky, A.S.; Kah, M.O.; Sandzhieva, D.A.; Mukabenova, B.A.; Dedov, A.G. Microorganisms in spent water-miscible metalworking fluids as a resource of strains for their disposal. J. Clean. Prod. 2022, 350, 131438. [Google Scholar] [CrossRef]
- Koch, T. Microbiology of metalworking fluids: What we know and lessons to be learnt. Acta Mech. Autom. 2023, 17, 166–172. [Google Scholar] [CrossRef]
- Józwik, J.; Michałowska, M.; Budzik, G.; Legutko, S.; Kupczyk, M. Microbiological analysis of coolant used in machining. Adv. Sci. Technol. Res. J. 2023, 17, 206–214. [Google Scholar] [CrossRef]
- Khan, I.U.; Selvaraju, S.B.; Yadav, J.S. Occurrence and characterization of multiple novel genotypes of Mycobacterium immunogenum and Mycobacterium chelonae in metalworking fluids. FEMS Microbiol. Ecol. 2005, 54, 329–338. [Google Scholar] [CrossRef]
- Perkins, S.D.; Angenent, L.T. Potential pathogenic bacteria in metalworking fluids and aerosols from a machining facility. FEMS Microbiol. Ecol. 2010, 74, 643–654. [Google Scholar] [CrossRef]
- Omura, H.; Kajiki, A.; Ikegame, S.; Aono, A.; Mitarai, S.; Kitahara, Y. Mycobacterium immunogenum isolated from a metal worker in Japan. Kekkaku 2012, 87, 341–344. (In Japanese) [Google Scholar]
- Martinez, V.; Matabang, M.A.; Miller, D.; Aggarwal, R.; LaFortune, A. First case report on Empedobacter falsenii bacteremia. IDCases 2023, 33, e01814. [Google Scholar] [CrossRef]
- Shen, Y.; Dong, Y.; Zhu, H.; Dong, L. Pseudomonas xiamenensis in the cutting fluids on corrosion behavior of aluminum alloy 2219. Bioelectrochemistry 2023, 150, 108350. [Google Scholar] [CrossRef]
- Sandin, M.; Allenmark, S.; Edebo, L. Selective toxicity of alkanolamines. Antimicrob. Agents Chemother. 1990, 34, 491–493. [Google Scholar] [CrossRef]
- Bruze, M.; Hradil, E.; Eriksohn, I.L.; Gruvberger, B.; Widstorm, L. Occupational allergic contact dermatitis from alkanolamineborates in metalworking fluids. Contact Dermat. 1995, 32, 24–27. [Google Scholar] [CrossRef]
- Kreiss, K.; Cox-Ganser, J. Metalworking fluid associated hypersensitivity pneumonitis: A workshop summary. Am. J. Ind. Med. 1997, 32, 423–432. [Google Scholar] [CrossRef]
- Rosenman, K.D. Asthma, hypersensitivity pneumonitis and other respiratory diseases caused by metalworking fluids. Curr. Opin. Allergy Clin. Immunol. 2009, 9, 97–102. [Google Scholar] [CrossRef]
- Ferraz de Campos, F.P.; Felipe-Silva, A.; Lopes, A.C.F.M.M.; Passadore, L.F.; Guida, S.M.; Balabakis, A.J.; dos Santos Martines, J.A. Community-acquired Pseudomonas aeruginosa-pneumonia in a previously healthy man occupationally exposed to metalworking fluids. Autops. Case Rep. 2014, 4, 31–37. [Google Scholar] [CrossRef][Green Version]
- Cummings, K.J.; Stanton, M.L.; Nett, R.J.; Segal, L.N.; Kreiss, K.; Abraham, J.L.; Colby, T.V.; Franko, A.D.; Green, F.H.Y.; Sanyal, S.; et al. Severe lung disease characterized by lymphocytic bronchiolitis, alveolar ductitis, and emphysema (BADE) in industrial machine-manufacturing workers. Am. J. Ind. Med. 2019, 62, 927–937. [Google Scholar] [CrossRef]
- Bellanger, A.P.; Morisse-Pradier, H.; Reboux, G.; Scherer, E.; Pramil, S.; Dominique, S.; Millon, L. Hypersensitivity pneumonitis in a cystic fibrosis patient. Occup. Med. 2019, 69, 632–634. [Google Scholar] [CrossRef]
- Kespohl, S.; Warfolomeow, I.; Schneider, G.; Maryska, S.; Meurer, U.; Raulf, M. Microbial contamination in water-based metalworking fluid as trigger for occupational hypersensitivity pneumonitis—Development of specific IgG tools for a suspected clinical case. Allergol. Sel. 2020, 4, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.G.; Kapoor, B.; Cummings, K.J.; Stanton, M.L.; Nett, R.J.; Kreiss, K.; Abraham, J.L.; Colby, T.V.; Franko, A.D.; Green, F.H.Y.; et al. Evidence for environmental-human microbiota transfer at a manufacturing facility with novel work-related respiratory disease. Am. J. Respir. Crit. Care Med. 2020, 202, 1678–1688. [Google Scholar] [CrossRef] [PubMed]
- Tustin, A.W.; Cooney, R.; Lamson, G.E.; Hodgson, M.J. A cluster of hypersensitivity pneumonitis associated with exposure to metalworking fluids. Am. J. Ind. Med. 2021, 64, 915–923. [Google Scholar] [CrossRef]
- Nett, R.J.; Stanton, M.; Grimes, G.R. Occupational respiratory and skin diseases among workers exposed to metalworking fluids. Curr. Opin. Allergy Clin. Immunol. 2021, 21, 121–127. [Google Scholar] [CrossRef]
- Walters, G.I.; Huntley, C.C. Novel occupational causes of hypersensitivity pneumonitis. Curr. Opin. Allergy Clin. Immunol. 2023, 23, 85–91. [Google Scholar] [CrossRef]
- Falkinham, J.O., III. Mycobacterial aerosols and respiratory diseases. Emerg. Infect. Dis. 2003, 9, 763–767. [Google Scholar] [CrossRef]
- Castellan, R.M.; Olenchock, S.A.; Kinsley, K.B.; Hankinson, J.L. Inhaled endotoxin and decreased spirometric values. An exposure-response relation for cotton dust. N. Engl. J. Med. 1987, 317, 605–610. [Google Scholar] [CrossRef]
- Gordon, T. Dose-dependent pulmonary effects of inhaled endotoxin in guinea pigs. Environ. Res. 1992, 59, 416–426. [Google Scholar] [CrossRef]
- Thorne, P.S.; DeKoster, J.A. Pulmonary effects of machining fluids in guinea pigs and mice. Am. Ind. Hyg. Assoc. J. 1996, 57, 1168–1172. [Google Scholar] [CrossRef]
- Thorne, P.S. Inhalation toxicology models of endotoxin- and bioaerosol-induced inflammation. Toxicology 2000, 152, 13–23. [Google Scholar] [CrossRef]
- Cummings, K.J.; Stanton, M.L.; Kreiss, K.; Boylstein, R.J.; Park, J.H.; Cox-Ganser, J.M.; Virji, M.A.; Edwards, N.T.; Segal, L.N.; Blaser, M.J.; et al. Work-related adverse respiratory health outcomes at a machine manufacturing facility with a cluster of bronchiolitis, alveolar ductitis and emphysema (BADE). Occup. Environ. Med. 2020, 77, 386–392. [Google Scholar] [CrossRef]
- Brassard, J.; Roy, J.; Lemay, A.-M.; Beaulieu, M.-J.; Bernatchez, E.; Veillette, M.; Duchaine, C.; Blanchet, M.-R. Exposure to the gram-negative bacteria Pseudomonas aeruginosa influences the lung dendritic cell population signature by interfering with cd103 expression. Front. Cell. Infect. Microbiol. 2021, 11, 617481. [Google Scholar] [CrossRef]
- Gordon, T.; Nadziejko, C.; Galdanes, K.; Lewis, D.; Donnelly, K. Mycobacterium immunogenum causes hypersensitivity pneumonitis-like pathology in mice. Inhal. Toxicol. 2006, 18, 449–456. [Google Scholar] [CrossRef]
- Thorne, P.S.; Adamcakova-Dodd, A.; Kelly, K.M.; O’neill, M.E.; Duchaine, C. Metalworking fluid with mycobacteria and endotoxin induces hypersensitivity pneumonitis in mice. Am. J. Respir. Crit. Care Med. 2006, 173, 759–768. [Google Scholar] [CrossRef]
- Johansson, E.; Boivin, G.P.; Yadav, J.S. Early immunopathological events in acute model of mycobacterial hypersensitivity pneumonitis in mice. J. Immunotoxicol. 2017, 14, 77–88. [Google Scholar] [CrossRef]
- Mattsby-Baltzer, I.; Sandin, M.; Ahlström, B.; Allenmark, S.; Edebo, M.; Falsen, E.; Pedersen, K.; Rodin, N.; Thompson, R.A.; Edebo, L. Thompson, and L. Edebo. Microbial growth and accumulation in industrial metal-working fluids. Appl. Environ. Microbiol. 1989, 55, 2681–2689. [Google Scholar] [CrossRef]
- Lonon, M.K.; Abanto, M.; Findlay, R.H. A pilot study for monitoring changes in the microbiological component of metalworking fluids as a function of time and use in the system. Am. Ind. Hyg. Assoc. J. 1999, 60, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, R.; Selvaraju, S.B.; Yadav, J.S. Extended tracking of the microbial community structure and dynamics in an industrial synthetic metalworking fluid system. FEMS Microbiol. Ecol. 2014, 87, 664–677. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lange, J.L.; Thorne, P.S.; Lynch, N. Application of flow cytometry and fluorescent in situ hybridization for assessment of exposures to airborne bacteria. Appl. Environ. Microbiol. 1997, 63, 1557–1563. [Google Scholar] [CrossRef] [PubMed]
- Edwards, C. Problems posed by natural environments for monitoring microorganisms. Mol. Biotechnol. 2000, 15, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Pace, N.R. A molecular view of microbial diversity and the biosphere. Science 1997, 276, 734–740. [Google Scholar] [CrossRef]
- Thorne, P.S.; DeKoster, J.A.; Subramanian, P. Environmental assessment of aerosols, bioaerosols, and airborne endotoxin in machining plant. Am. Ind. Hyg. Assoc. J. 1996, 57, 1163–1167. [Google Scholar] [CrossRef]
- Selvaraju, S.B.; Khan, I.U.H.; Yadav, J.S. Specific detection and quantification of culturable and non-culturable mycobacteria in metalworking fluids by fluorescence-based methods. Lett. Appl. Microbiol. 2008, 47, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Vanhauteghem, D.; Audenaert, K.; Demeyere, K.; Hoogendoorn, F.; Janssens, G.P.J.; Meyer, E. Flow cytometry, a powerful novel tool to rapidly assess bacterial viability in metal working fluids: Proof-of-principle. PLoS ONE 2019, 14, e0211583. [Google Scholar] [CrossRef] [PubMed]
- Yadav, J.S.; Khan, I.U.; Fakhari, F.; Soellner, M.B. DNA-based methodologies for rapid detection, quantification, and species- or strain-level identification of respiratory pathogens (Mycobacteria and Pseudomonads) in metalworking fluids. Appl. Occup. Environ. Hyg. 2003, 18, 966–975. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.U.; Yadav, J.S. Real-time PCR assays for genus-specific detection and quantification of culturable and non-culturable mycobacteria and pseudomonads in metalworking fluids. Mol. Cell. Probes 2004, 18, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Selvaraju, S.B.; Khan, I.U.H.; Yadav, J.S. An improved method for species differentiation in Mycobacterium chelonae complex based on amplified hsp65 restriction analysis (AHSPRA). Mol. Cell. Probes 2005, 19, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.U.H.; Selvaraju, S.B.; Yadav, J.S. Species differentiation for Mycobacterium chelonae complex based on 16S-23S Ribosomal DNA internal transcribed spacer (ITS) sequencing and ITS PCR-restriction analysis (ITSPRA). J. Clin. Microbiol. 2005, 43, 4466–4472. [Google Scholar] [CrossRef] [PubMed]
- Selvaraju, S.B.; Kapoor, R.; Yadav, J.S. Peptide Nucleic Acid-Fluorescence in-situ hybridization (PNA-FISH) assay for specific detection of Mycobacterium immunogenum and DNA-FISH assay for analysis of pseudomonads in metalworking fluids and sputum. Mol. Cell. Probes 2008, 22, 273–280. [Google Scholar] [CrossRef]
- Kapoor, R.; Yadav, J.S. Development of a species-specific colorimetric-PCR assay for detection and species differentiation of Mycobacterium immunogenum and M. chelonae and its comparison with quantitative real-time PCR for metalworking fluids. Mol. Cell. Probes 2009, 23, 75–82. [Google Scholar] [CrossRef]
- Di Maiuta, N.; Rüfenacht, A.; Küenzi, P. Assessment of bacteria and archaea in metalworking fluids using massive parallel 16S rRNA gene tag sequencing. Lett. Appl. Microbiol. 2017, 65, 266–273. [Google Scholar] [CrossRef]
- Das, M.; Royer, T.V.; Leff, L.G. Diversity of fungi, bacteria, and actinomycetes on leaves decomposing in a stream. Appl. Environ. Microbiol. 2007, 73, 756–767. [Google Scholar] [CrossRef]
- Ibekwe, A.M.; Lyon, S.R.; Leddy, M.; Jacobson-Meyers, M. Impact of plant density and microbial composition on water quality from a free water surface constructed wetland. J. Appl. Microbiol. 2007, 102, 921–936. [Google Scholar] [CrossRef] [PubMed]
- Labbe, D.; Margesin, R.; Schinner, F.; Whyte, L.G.; Greer, C.W. Comparative phylogenetic analysis of microbial communities in pristine and hydrocarbon-contaminated Alpine soils. FEMS Microbiol. Ecol. 2007, 59, 466–475. [Google Scholar] [CrossRef]
- Gilbert, Y.; Veillette, M.; Duchaine, C. Metalworking fluids biodiversity characterization. J. Appl. Microbiol. 2010, 108, 437–449. [Google Scholar] [CrossRef]
- Veillette, M.; Thorne, P.S.; Gordon, T.; Duchaine, C. Six month tracking of microbial growth in a metalworking fluid after system cleaning and recharging. Ann. Occup. Hyg. 2004, 48, 541–546. [Google Scholar] [PubMed]
- Murat, J.-B.; Grenouillet, F.; Reboux, G.; Penven, E.; Batchili, A.; Dalphin, J.-C.; Thaon, I.; Millon, L. Factors influencing the microbial composition of metalworking fluids and potential implications for machine operator’s lung. Appl. Environ. Microbiol. 2012, 78, 34–38. [Google Scholar] [CrossRef]
- Vincent, C. The direct or Breed method for counting bacteria in tomato catsup, pulp or paste. J. Bacteriol. 1918, 3, 183–185. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.U.H.; Yadav, J.S. Developement of a single tube, cell lysis-based, Genus specific PCR method for rapid identification of Mycobacteria: Optimization of cell lysis, PCR primers and conditions and restriction pattern analysis. J. Clin. Microbiol. 2004, 42, 453–457. [Google Scholar] [CrossRef]
- Johnsen, K.; Enger, O.; Jacobsen, C.S.; Thirup, L.; Torsvik, V. Quantitative selective PCR of 16S ribosomal DNA correlates well with selective agar plating in describing population dynamics of indigenous Pseudomonas spp. in soil hot spots. Appl. Environ. Microbiol. 1999, 65, 1786–1788. [Google Scholar] [CrossRef]
- Bayardelle, P.; Zafarullah, M. Development of oligonucleotide primers for the specific PCR-based detection of the most frequent Enterobacteriaceae species DNA using wec gene templates. Can. J. Microbiol. 2002, 48, 113–122. [Google Scholar] [CrossRef]
- Wellinghausen, N.; Frost, C.; Marre, R. Detection of legionellae in hospital water samples by quantitative real-time LightCycler PCR. Appl. Environ. Microbiol. 2001, 67, 3985–3993. [Google Scholar] [CrossRef]
- Heuer, H.; Krsek, M.; Baker, P.; Smalla, K.; Wellington, E.M. Analysis of actinomycete communities by specific amplification of genes encoding 16S rRNA and gel-electrophoretic separation in denaturing gradients. Appl. Environ. Microbiol. 1997, 63, 3233–3241. [Google Scholar] [CrossRef]
- White, T.J.; Brum, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribososmal RNA genes for phylogenetics. In PCR Protocols: Guide to Methods and Application; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–321. [Google Scholar]
- McBain, A.J.; Bartolo, R.G.; Catrenich, C.E.; Charbonneau, D.; Ledder, R.G.; Price, B.B.; Gilbert, P. Exposure of sink drain microcosms to triclosan: Population dynamics and antimicrobial susceptibility. Appl. Environ. Microbiol. 2003, 69, 5433–5442. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, D.I.; Lummus, Z.L.; Santilli, G.; Siskosky, J.; Bernstein, I.L. Machine operator’s lung. A hypersensitivity pneumonitis disorder associated with exposure to metalworking fluid aerosols. Chest 1995, 108, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Bukowski, J.A. Review of respiratory morbidity from occupational exposure to oil mists. Appl. Occup. Environ. Hyg. 2003, 18, 828–837. [Google Scholar] [CrossRef]
- Simpson, A.T.; Stear, M.; Groves, J.A.; Piney, M.; Bradley, S.D.; Stagg, S.; Crook, B. Occupational Exposure to Metalworking Fluid Mist and Sump Fluid Contaminants. Ann. Occup. Hyg. 2003, 47, 17–30. [Google Scholar]
- Gordon, T. Metalworking Fluid—The Toxicity of a Complex Mixture. J. Toxicol. Environ. Health A 2004, 67, 209–219. [Google Scholar] [CrossRef]
- Gupta, A.; Rosenman, K.D. Hypersensitivity pneumonitis due to metal working fluids: Sporadic or under reported? Am. J. Ind. Med. 2006, 49, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Cook, P.E.; Gaylarde, C.C. Biofilm formation in aqueous metal working fluids. Int. Biodeterior. 1988, 24, 265–270. [Google Scholar] [CrossRef]
- Özcan, S.S.; Dieser, M.; Parker, A.E.; Balasubramanian, N.; Foreman, C.M. Quorum sensing inhibition as a promising method to control biofilm growth in metalworking fluids. J. Ind. Microbiol. Biotechnol. 2019, 46, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Foxall-VanAken, S.; Brown, J.A., Jr.; Young, W.; Salmeen, I.; McClure, T.; Napier, S., Jr.; Olsen, R.H. Common components of industrial metal-working fluids as sources of carbon for bacterial growth. Appl. Environ. Microbiol. 1986, 51, 1165–1169. [Google Scholar] [CrossRef] [PubMed]
- Sloyer, J.L.; Novitsky, T.J.; Nugent, S. Rapid bacterial counts in metalworking fluids. J. Ind. Microbiol. Biotechnol. 2002, 29, 323–324. [Google Scholar] [CrossRef] [PubMed]
- van Der Gast, C.J.; Whiteley, A.S.; Lilley, A.K.; Knowles, C.J.; Thompson, I.P. Bacterial community structure and function in a metal-working fluid. Environ. Microbiol. 2003, 5, 453–461. [Google Scholar] [CrossRef]
- Selvaraju, S.B.; Khan, I.U.H.; Yadav, J.S. Susceptibility of Mycobacterium immunogenum and Pseudomonas fluorescens to formaldehyde and non-formaldehyde biocides in semi-synthetic metalworking fluids. Int. J. Mol. Sci. 2011, 12, 725–741. [Google Scholar] [CrossRef]
- Virji, M.A.; Woskie, S.R.; Sama, S.R.; Kriebel, D.; Eberiel, D. Identifying the determinants of viable microorganisms in the air and bulk metalworking fluids. Am. Ind. Hyg. Assoc. J. 2000, 61, 788–797. [Google Scholar] [CrossRef]
- Bracker, A.; Storey, E.; Yang, C.; Hodgson, M.J. An outbreak of hypersensitivity pneumonitis at a metalworking plant: A longitudinal assessment of intervention effectiveness. Appl. Occup. Environ. Hyg. 2003, 18, 95–108. [Google Scholar] [CrossRef]
- Marchand, G.; Lavioe, J.; Racine, L.; Lacombe, N.; Cloutier, Y.; Belanger, E.; Lemelin, C.; Desroches, J. Evaluation of bacterial contamination and control methods in soluble metalworking fluids. J. Occup. Environ. Hyg. 2010, 7, 358–366. [Google Scholar] [CrossRef]
- Jabeen, I.; Islam, S.; Hassan, A.K.M.I.; Tasnim, Z.; Shuvo, S.R. A brief insight into Citrobacter species—A growing threat to public health. Front. Antibiot. 2023, 2, 127. [Google Scholar] [CrossRef]
- Rossmore, H.W. Microbial degradation of water-based metalworking fluids. In Comprehensive Biotechnology, Vol. 2, The Principles of Biotechnology: Engineering Considerations; Moo-Young, M., Cooney, C.L., Humphrey, A., Eds.; Pergamon Press: New York, NY, USA, 1985; pp. 249–268. [Google Scholar]
- Wang, W.; Luo, B.; Yuan, X.; Xu, L.; Gao, J.; Liu, S.; Den, C.; Tan, Y.; Su, G.; Chen, Z.; et al. Diagnosis of fungal keratitis caused by Nectria haematococca through next-generation sequencing: Review of literature and report of three cases. Int. J. Infect. Dis. 2023, 131, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.J., Jr.; Zhang, Y.; Wilson, R.W.; Mann, L.; Rossmore, H. Presence of a Single Genotype of the Newly Described Species Mycobacterium immunogenum in Industrial Metalworking Fluids Associated with Hypersensitivity Pneumonitis. Appl. Environ. Microbiol. 2002, 68, 5580–5584. [Google Scholar] [CrossRef]
- Trout, D.; Weissman, D.N.; Lewis, D.; Brundage, R.A.; Franzblau, A.; Remick, D. Evaluation of Hypersensitivity Pneumonitis Among Workers Exposed to Metal Removal Fluids. Appl. Environ. Microbiol. 2003, 18, 953–960. [Google Scholar] [CrossRef]
- Beckett, W.; Kallay, M.; Sood, A.; Zuo, Z.; Milton, D. Hypersensitivity Pneumonitis associated with environmental Mycobacteria. Environ. Health Perspect. 2005, 113, 767–770. [Google Scholar] [CrossRef] [PubMed]
- Marx, J.J., Jr.; Emanuel, D.A.; Dovenbarger, W.V.; Reinecke, V.; Roberts, R.C.; Treuhaft, M.W. Farmer’s lung disease among farmers with precipitating antibodies to the thermophilic actinomycetes: A clinical and immunologic study. J. Allergy Clin. Immunol. 1978, 62, 185–189. [Google Scholar] [PubMed]
- van der Gast, C.J.; Knowles, C.J.; Starkey, M.; Thompson, I.P. Selection of microbial consortia for treating metal-working fluids. J. Ind. Microbiol. Biotech. 2002, 29, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Burton, C.M.; BCrook HSaife GHEvans, C.M. Barber. Systematic review of respiratory outbreak associated with exposure to water-based metalworking fluids. Ann. Occup. Hyg. 2012, 56, 374–388. [Google Scholar] [PubMed]
- Mattsby-Baltzer, I.; Edebo, L.; Järvholm, B.; Lavenius, B. Serum antibodies to Pseudomonas pseudoalcali genes in metal workers exposed to infected metal-working fluids. Int. Arch. Allergy Immunol. 1989, 88, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Milton, D.K.; Gere, R.J.; Feldman, H.A.; Greaves, I.A. Endotoxin measurement: Aerosol sampling and application of a new Limulus method. Am. Ind. Hyg. Assoc. 1990, 51, 331–337. [Google Scholar] [CrossRef]
- Cyprowski, M.; Piotrowska, M.; Zakowska, Z.; Szadkowska-Stańczyk, I. Microbial and endotoxin contamination of water-soluble metalworking fluids. Int. J. Occup. Med. Environ. Health 2007, 20, 365–371. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targeted Group/Genus | Primer Sequence | Source Reference |
|---|---|---|
| Pseudomonads | Forward-5′ GAG TTT GAT CCT GGC TCA G 3′ Reverse-5′ CCT TCC TCC CAA CTT 3′ | [61] |
| Enteric Bacteria | Forward-5′ TCC GTG AAA AGG GCA CTA AC 3′ Reverse-5′ CAG ATC GGA CAT CAA ATA GC 3′ | [62] |
| Mycobacteria | Forward-5′ CTG GTC AAG GAA GGT CTG GC 3′ Reverse-5′ GAT GAC ACC CYTC GTT GCC AAC 3′ | [47] |
| Legionellae | Forward-5′ AGG GTT GAT AGG TTA AGA GC 3′ Reverse-5′ CCA ACA GCT AGT TGA CAT CG 3′ | [63] |
| Actinomycetes | Forward-5′ GGA TGA GCC CGC GGC CTA 3′ Reverse-5′ CGG CCG CGG CTG CTG GCA CGT A 3′ | [64] |
| Fungi | Forward-5′ GGC TCT CGC ATC GAT GAA GAA C 3′ Reverse-5′ CTT TTC CTC CGC TTA TTG ATA TGC 3′ | [65] |
| Culture Medium ¥ | Isolate(s) |
|---|---|
| TSA | Alcaligenes sp., Caulobacter sp., Citrobacter sp., Methylobacterium sp., Pseudomonas sp., Staphylococcus sp. * |
| EMB | Acidovorax sp., Alcaligenes sp., Bacillus cereus *, Citrobacter sp., Morganella morganii, Pseudomonas sp., Sinorhizobium sp. |
| PIA | Achromobacter sp., Aeromonas sp., Citrobacter sp., Morganella morganii, Serratia sp., Stenotrophomonas sp. |
| MBA | Alcaligenes sp., Bacillus cereus * |
| BCYE | Sinorhozobium sp., Sphignobacterium sp. |
| AIA | Bacillus cereus *, Citrobacter sp., Nectria haematococca # |
| SDA | Enterobacter sp., Nectria haematococca # |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapoor, R.; Selvaraju, S.B.; Subramanian, V.; Yadav, J.S. Microbial Community Establishment, Succession, and Temporal Dynamics in an Industrial Semi-Synthetic Metalworking Fluid Operation: A 50-Week Real-Time Tracking. Microorganisms 2024, 12, 267. https://doi.org/10.3390/microorganisms12020267
Kapoor R, Selvaraju SB, Subramanian V, Yadav JS. Microbial Community Establishment, Succession, and Temporal Dynamics in an Industrial Semi-Synthetic Metalworking Fluid Operation: A 50-Week Real-Time Tracking. Microorganisms. 2024; 12(2):267. https://doi.org/10.3390/microorganisms12020267
Chicago/Turabian StyleKapoor, Renuka, Suresh Babu Selvaraju, Venkataramanan Subramanian, and Jagjit S. Yadav. 2024. "Microbial Community Establishment, Succession, and Temporal Dynamics in an Industrial Semi-Synthetic Metalworking Fluid Operation: A 50-Week Real-Time Tracking" Microorganisms 12, no. 2: 267. https://doi.org/10.3390/microorganisms12020267
APA StyleKapoor, R., Selvaraju, S. B., Subramanian, V., & Yadav, J. S. (2024). Microbial Community Establishment, Succession, and Temporal Dynamics in an Industrial Semi-Synthetic Metalworking Fluid Operation: A 50-Week Real-Time Tracking. Microorganisms, 12(2), 267. https://doi.org/10.3390/microorganisms12020267