
Antibacterial, Resistance Modulation, Anti-Biofilm Formation, and Efflux Pump Inhibition Properties of Loeseneriella africana (Willd.) N. Halle (Celastraceae) Stem Extract and Its Constituents
 ,
,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Drugs and Chemicals
2.2. Bacterial Strains and Inoculum Standardization
2.3. General Experimental Procedures
2.4. Plant Material Collection
2.5. Preliminary Phytochemical Screening
2.6. Preparation of Extracts and Fractions
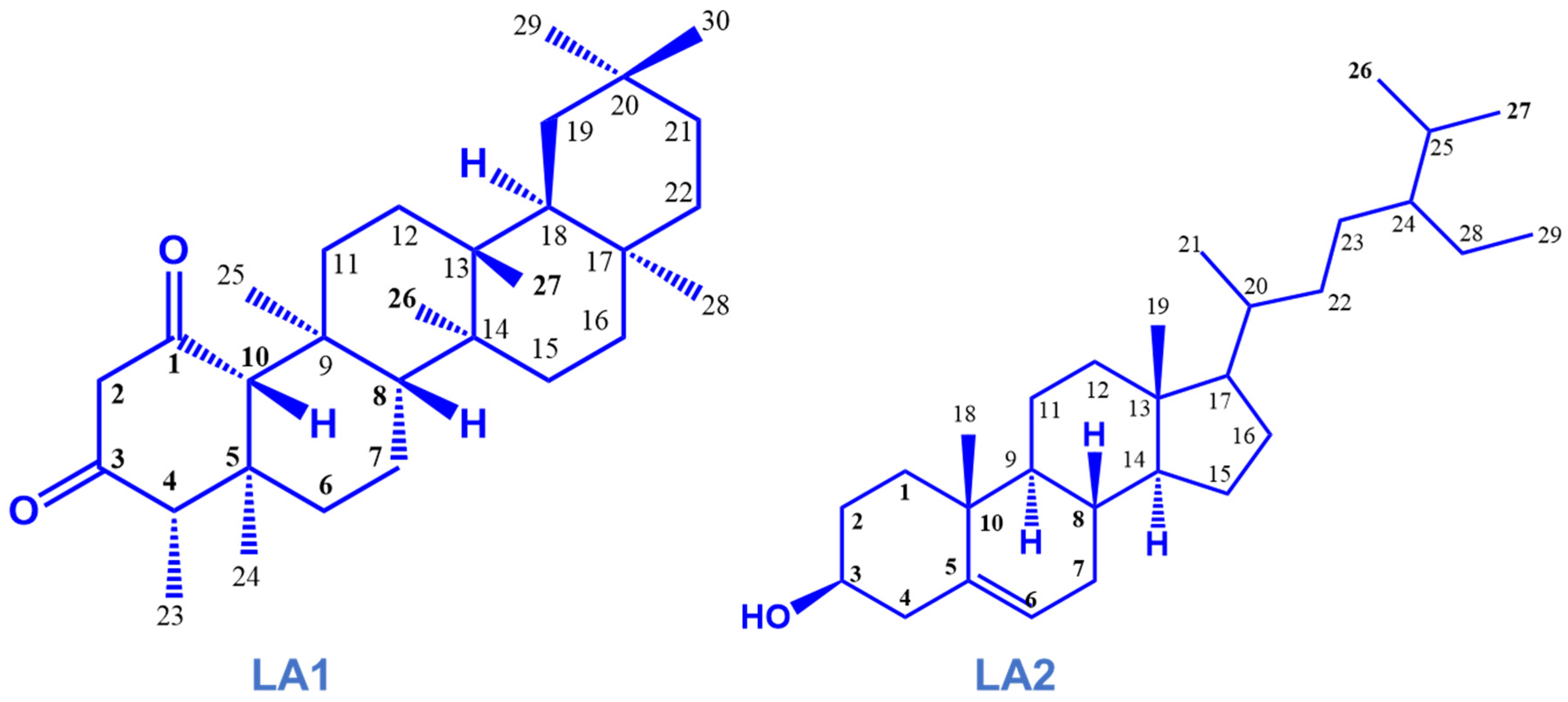
2.7. Isolation and Characterization of Phytoconstituents
2.8. Antimicrobial Testing
2.8.1. Evaluation of Antibacterial Activity of Crude Extracts and Fractions
2.8.2. Determination of the Antibacterial Activity and Antibiotic Modulation Effect of Isolated Compounds
2.8.3. Biofilm Inhibition Assay
2.8.4. Efflux Pump Inhibition Assay
3. Results
3.1. Preliminary Phytochemical Investigation of the Stem of L. africana
3.2. Isolated Compounds from the Stem of L. africana
3.3. Antibacterial Activity of L. africana Stem Extract, Major Fractions and Isolated Compounds
3.4. Antibiotic Modulation Effect of Isolated Compounds from L. africana Stem
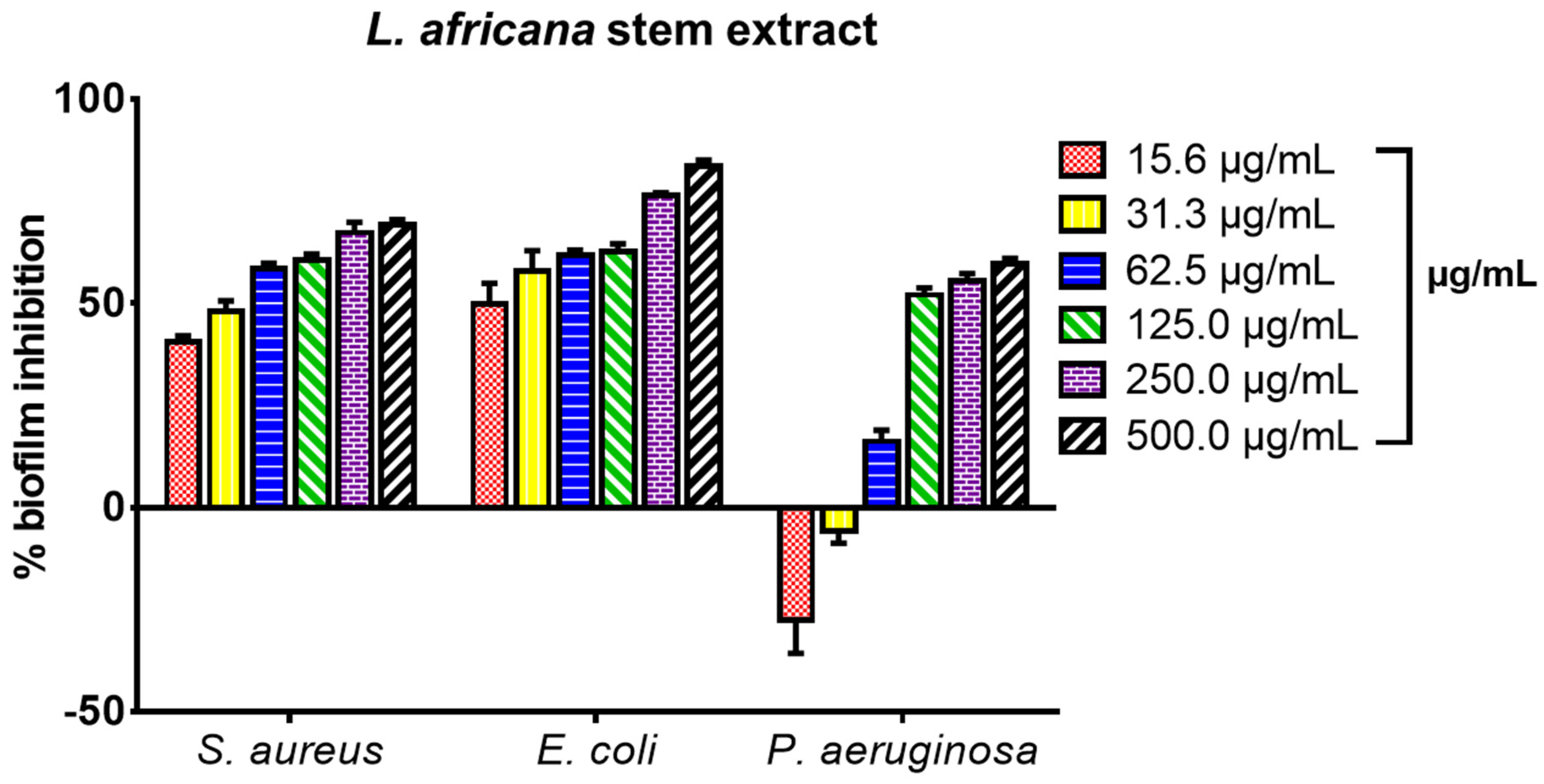
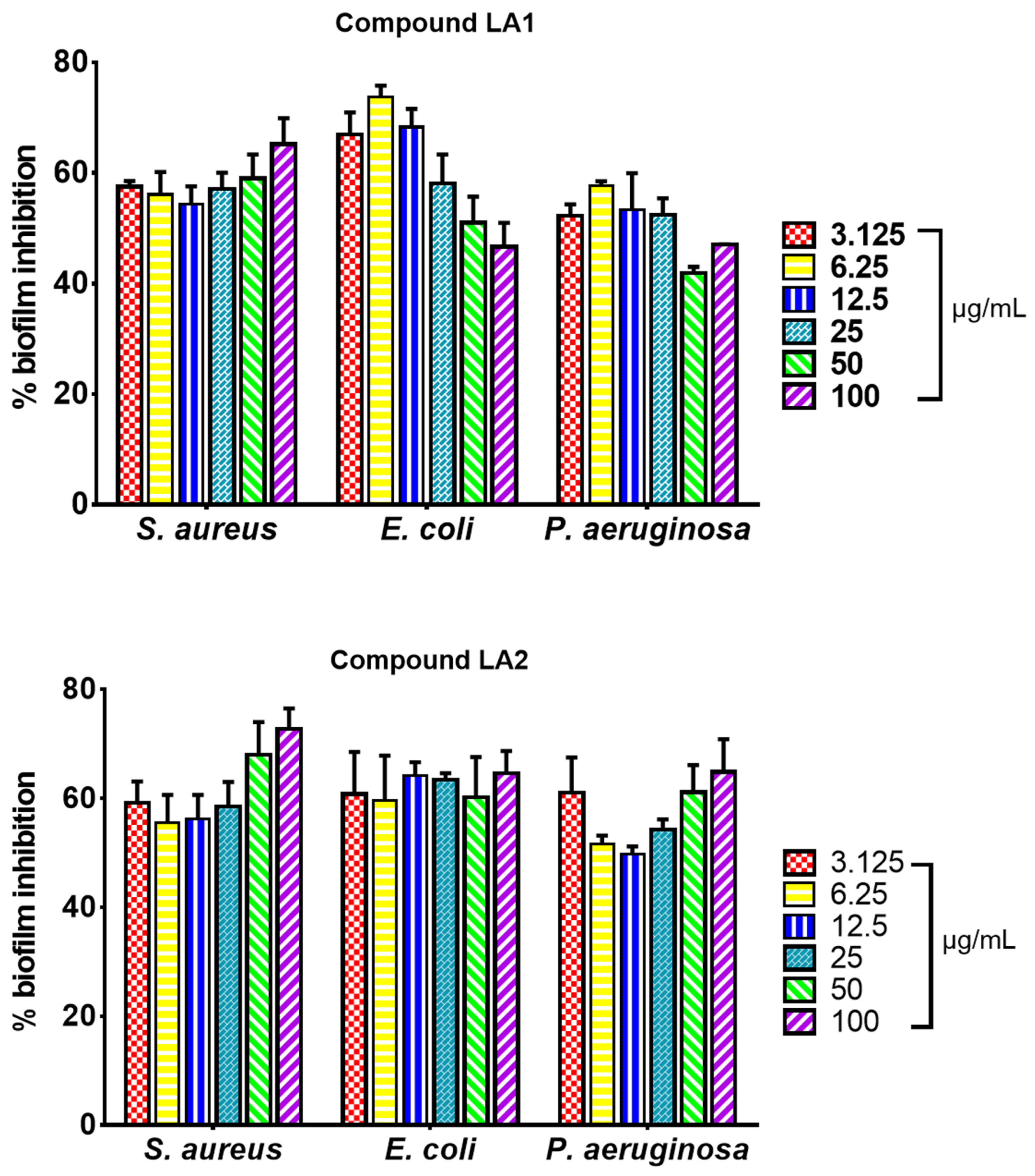
3.5. Biofilm Inhibitory Effect of L. africana Stem Extract and Isolated Compounds
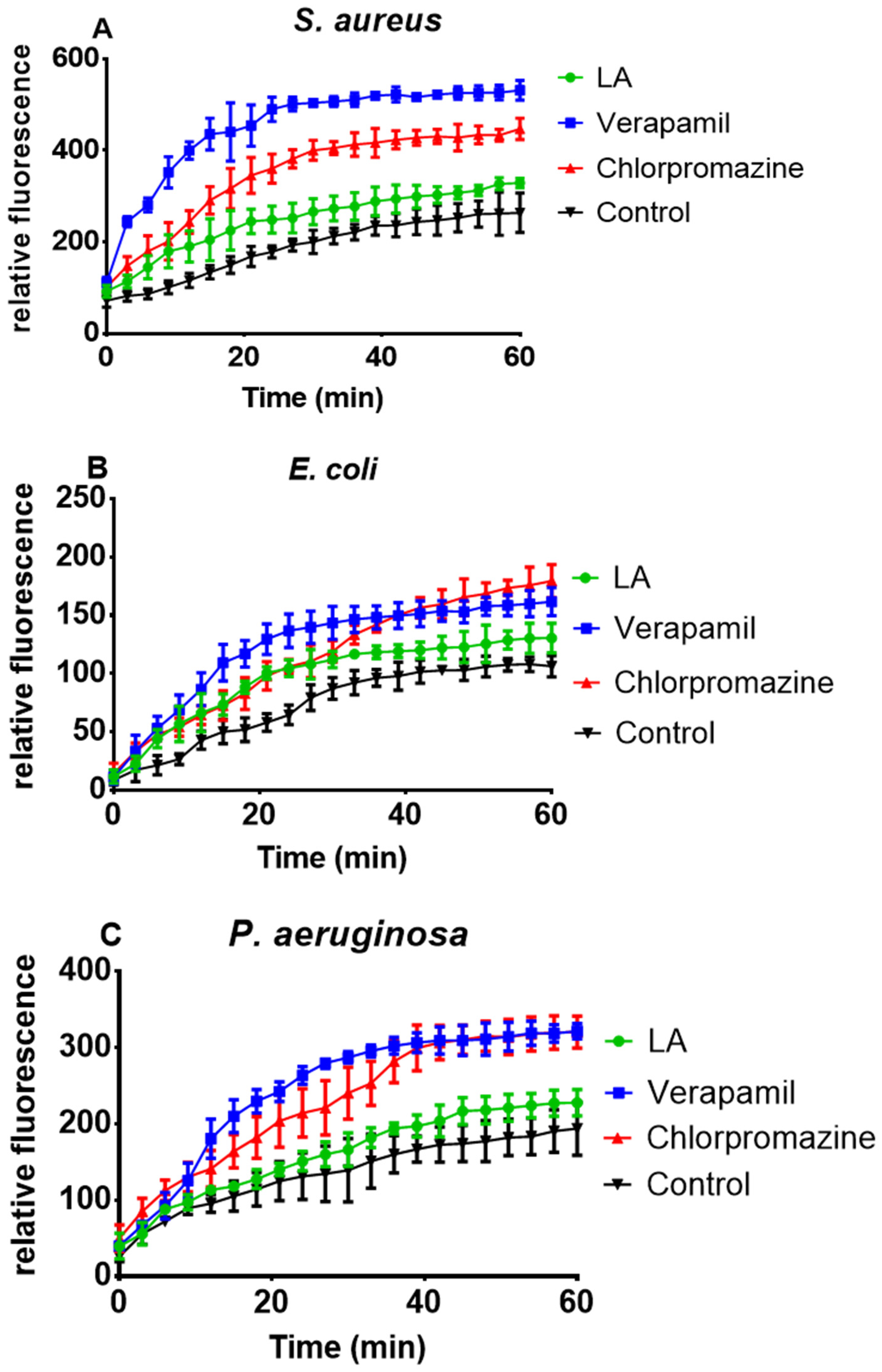
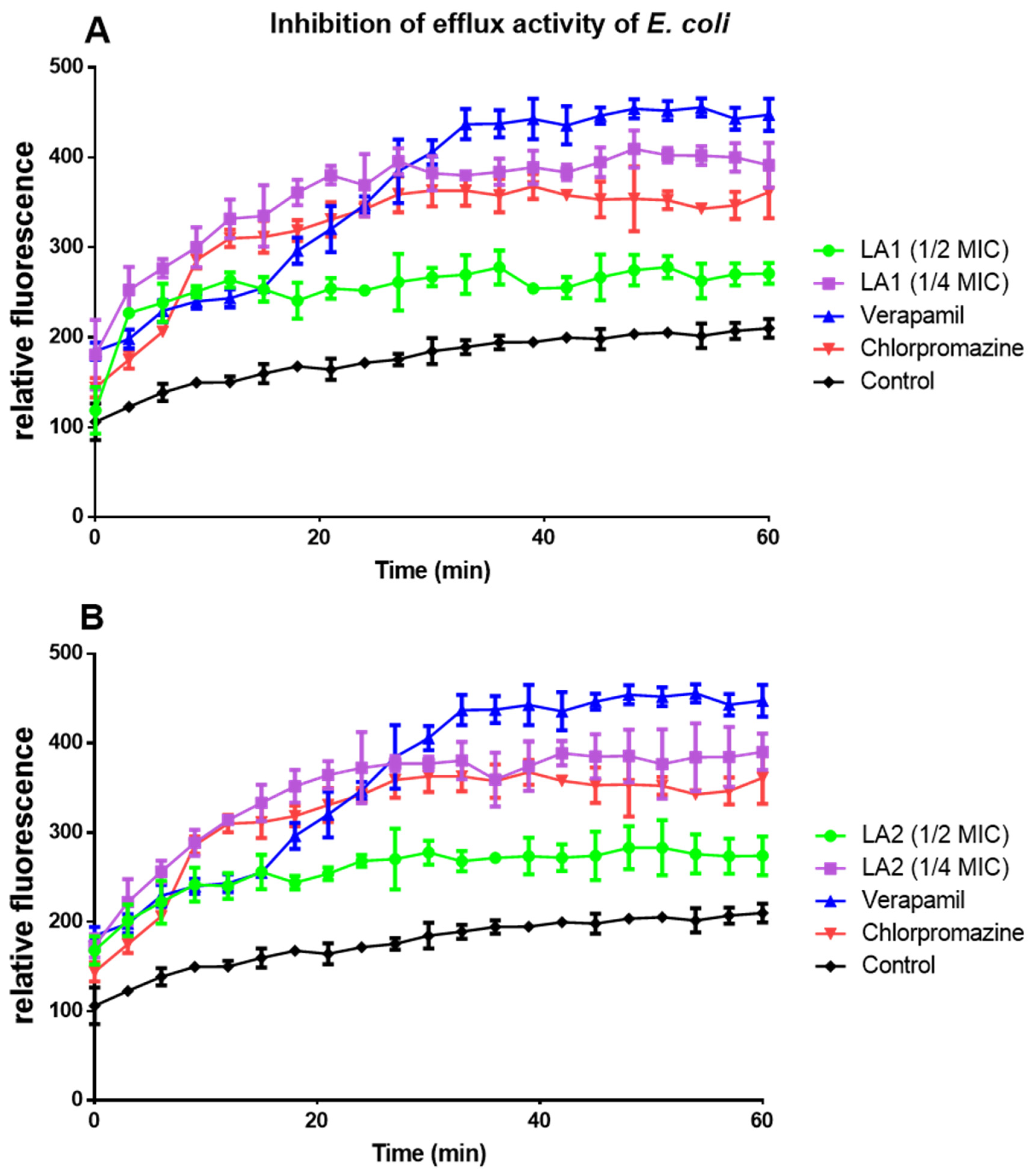
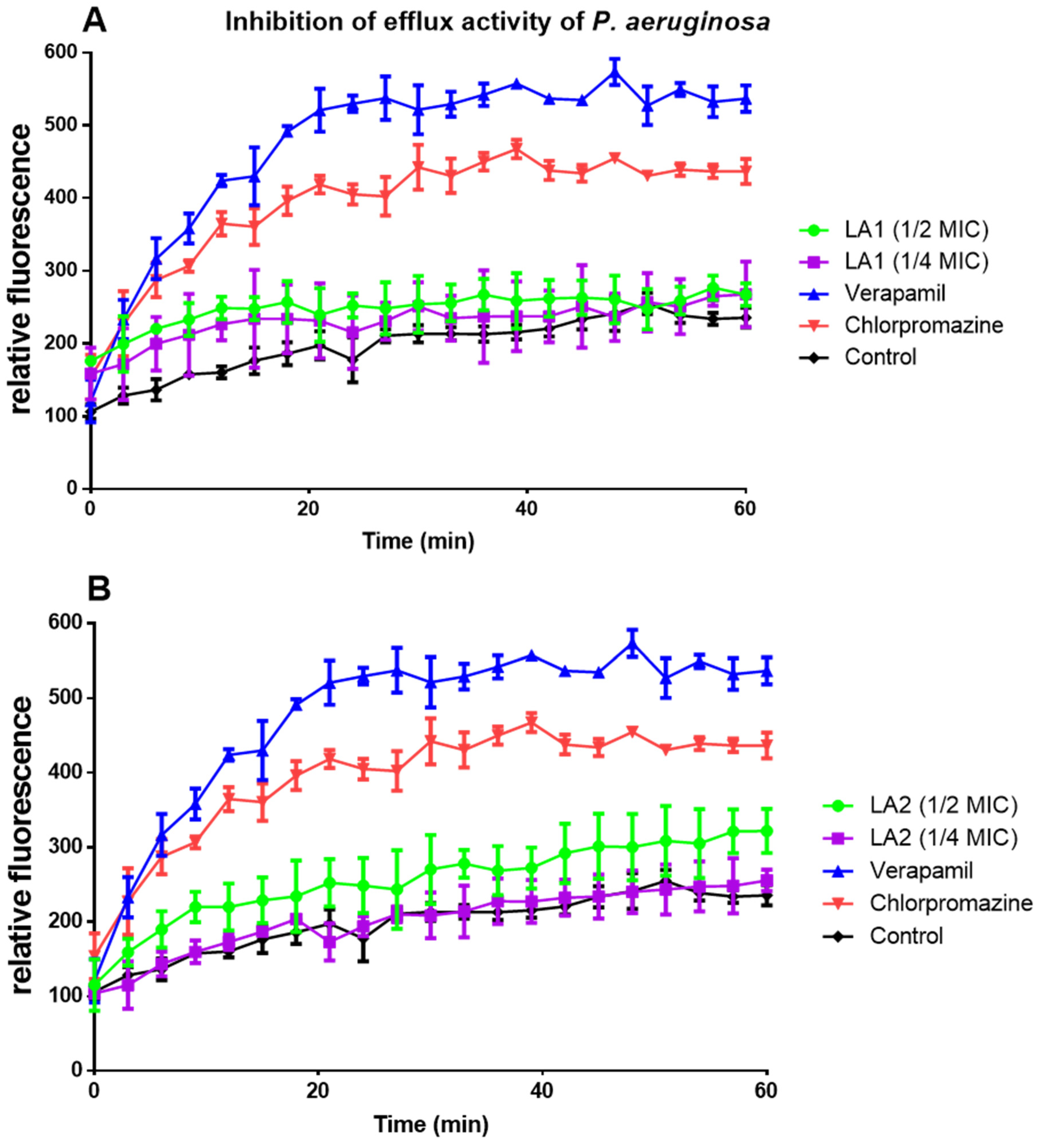
3.6. Effect of L. africana Stem Extract and Isolated Compounds on Ethidium Bacterial Efflux Pump
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seukep, A.J.; Kuete, V.; Nahar, L.; Sarker, S.D.; Guo, M. Plant-derived secondary metabolites as the main source of efflux pump inhibitors and methods for identification. J. Pharm. Anal. 2020, 10, 277–290. [Google Scholar] [CrossRef]
- Laukkanen, M.O. Return of Communicable Diseases as a Result of Antibiotic Resistance? Antioxid. Redox Signal. 2021, 34, 439–441. [Google Scholar] [CrossRef]
- Igarashi, M. New natural products to meet the antibiotic crisis: A personal journey. J. Antibiot. 2019, 72, 890–898. [Google Scholar] [CrossRef]
- Kvist, M.; Hancock, V.; Klemm, P. Inactivation of efflux pumps abolishes bacterial biofilm formation. Appl. Environ. Microbiol. 2008, 74, 7376–7382. [Google Scholar] [CrossRef]
- Tamfu, A.N.; Ceylan, O.; Cârâc, G.; Talla, E.; Dinica, R.M. Antibiofilm and anti-quorum sensing potential of cycloartane-type triterpene acids from cameroonian grassland propolis: Phenolic profile and antioxidant activity of crude extract. Molecules 2022, 27, 4872. [Google Scholar] [CrossRef]
- Sánchez, E.; Morales, C.R.; Castillo, S.; Leos-Rivas, C.; García-Becerra, L.; Martínez, D.M.O. Antibacterial and antibiofilm activity of methanolic plant extracts against nosocomial microorganisms. Evid. Based Complement. Altern. Med. eCAM 2016, 2016, 1572697. [Google Scholar] [CrossRef]
- Touani, F.K.; Seukep, A.J.; Djeussi, D.E.; Fankam, A.G.; Noumedem, J.A.; Kuete, V. Antibiotic-potentiation activities of four Cameroonian dietary plants against multidrug-resistant Gram-negative bacteria expressing efflux pumps. BMC Complement. Altern. Med. 2014, 14, 258. [Google Scholar] [CrossRef]
- Wolcott, R.; Costerton, J.; Raoult, D.; Cutler, S. The polymicrobial nature of biofilm infection. Clin. Microbiol. Infect. 2013, 19, 107–112. [Google Scholar] [CrossRef]
- Tamfu, A.N.; Ceylan, O.; Fru, G.C.; Ozturk, M.; Duru, M.E.; Shaheen, F. Antibiofilm, antiquorum sensing and antioxidant activity of secondary metabolites from seeds of Annona senegalensis, Persoon. Microb. Pathog. 2020, 144, 104191. [Google Scholar] [CrossRef] [PubMed]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef] [PubMed]
- Van Wyk, A.S.; Prinsloo, G. Health, safety and quality concerns of plant-based traditional medicines and herbal remedies. S. Afr. J. Bot. 2020, 133, 54–62. [Google Scholar] [CrossRef]
- Burkill, H. The useful plants of West Africa, families MR. R. Bot. Gard. Kew 1985, 4, 359. [Google Scholar]
- Compaoré, M.; Meda, R.N.-T.; Bakasso, S.; Vlase, L.; Kiendrebeogo, M. Antioxidative, anti-inflammatory potentials and phytochemical profile of Commiphora africana (A. Rich.) Engl. (Burseraceae) and Loeseneriella africana (Willd.) (Celastraceae) stem leaves extracts. Asian Pac. J. Trop. Biomed. 2016, 6, 665–670. [Google Scholar] [CrossRef]
- Anokwah, D.; Asante-Kwatia, E.; Jibira, Y.; Amponsah, I.K.; Kyei, S.; Cobbold, G.P.; Adzekey, B.; Obese, E.; Ameyaw, E.O.; Biney, R.P. Pharmacognostic study, anti-inflammatory and antioxidant activities of Loeseneriella africana. S. Afr. J. Bot. 2023, 157, 174–188. [Google Scholar] [CrossRef]
- Evans, W.C. Trease and Evans’ Pharmacognosy; Elsevier Health Sciences: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Anokwah, D.; Asante-Kwatia, E.; Mensah, A.Y.; Danquah, C.A.; Harley, B.K.; Amponsah, I.K.; Oberer, L. Bioactive constituents with antibacterial, resistance modulation, anti-biofilm formation and efflux pump inhibition properties from Aidia genipiflora stem bark. Clin. Phytosci. 2021, 7, 28. [Google Scholar] [CrossRef]
- Danquah, C.A.; Maitra, A.; Gibbons, S.; Faull, J.; Bhakta, S. HT-SPOTi: A rapid drug susceptibility test (DST) to evaluate antibiotic resistance profiles and novel chemicals for anti-infective drug discovery. Curr. Protoc. Microbiol. 2016, 40, 17.8.1–17.8.12. [Google Scholar] [CrossRef]
- Cos, P.; Vlietinck, A.J.; Berghe, D.V.; Maes, L. Anti-infective potential of natural products: How to develop a stronger in vitro ‘proof-of-concept’. J. Ethnopharmacol. 2006, 106, 290–302. [Google Scholar] [CrossRef]
- Prasch, S.; Duran, A.G.; Chinchilla, N.; Molinillo, J.M.; Macías, F.A.; Bucar, F. Resistance modulatory and efflux-inhibitory activities of capsaicinoids and capsinoids. Bioorg. Chem. 2019, 82, 378–384. [Google Scholar] [CrossRef]
- Abidi, S.H.; Ahmed, K.; Sherwani, S.K.; Bibi, N.; Kazmi, S.U. Detection of Mycobacterium smegmatis biofilm and its control by natural agents. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 801–812. [Google Scholar]
- Stavri, M.; Piddock, L.J.; Gibbons, S. Bacterial efflux pump inhibitors from natural sources. J. Antimicrob. Chemother. 2007, 59, 1247–1260. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V. Potential of Cameroonian plants and derived products against microbial infections: A review. Planta Medica 2010, 76, 1479–1491. [Google Scholar] [CrossRef] [PubMed]
- Simoes, M.; Bennett, R.N.; Rosa, E.A. Understanding antimicrobial activities of phytochemicals against multidrug resistant bacteria and biofilms. Nat. Prod. Rep. 2009, 26, 746–757. [Google Scholar] [CrossRef] [PubMed]
- Aligiannis, N.; Kalpoutzakis, E.; Mitaku, S.; Chinou, I.B. Composition and antimicrobial activity of the essential oils of two Origanum species. J. Agric. Food Chem. 2001, 49, 4168–4170. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, M.S.; Hazell, S.L. Isolation, analysis and antimicrobial activity of the acidic fractions of Mastic, Kurdica, Mutica and Cabolica gums from genus Pistacia. Glob. J. Health Sci. 2012, 4, 217. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, M.S. Antimicrobial activities of triterpenoids against Methicillin-resistant Staphylococcus aureus (MRSA). Worldw. Res. Efforts Fight. Against Microb. Pathog. 2013, 9, 51. [Google Scholar]
- Ododo, M.M.; Choudhury, M.K.; Dekebo, A.H. Structure elucidation of β-sitosterol with antibacterial activity from the root bark of Malva parviflora. SpringerPlus 2016, 5, 1210. [Google Scholar] [CrossRef] [PubMed]
- Doğan, A.; Otlu, S.; Çelebi, Ö.; Aksu, P.; Sağlam, A.G.; Doğan, A.N.C.; Mutlu, N. An investigation of antibacterial effects of steroids. Turk. J. Vet. Anim. Sci. 2017, 41, 302–305. [Google Scholar] [CrossRef]
- Sen, A.; Dhavan, P.; Shukla, K.K.; Singh, S.; Tejovathi, G. Analysis of IR, NMR and antimicrobial activity of β-sitosterol isolated from Momordica charantia. Sci. Secur. J. Biotechnol. 2012, 1, 9–13. [Google Scholar]
- Mailafiya, M.M.; Yusuf, A.J.; Abdullahi, M.I.; Aleku, G.A.; Ibrahim, I.A.; Yahaya, M.; Abubakar, H.; Sanusi, A.; Adamu, H.W.; Alebiosu, C.O. Antimicrobial activity of stigmasterol from the stem bark of Neocarya macrophylla. J. Med. Plants Econ. Dev. 2018, 2, a38. [Google Scholar]
- Anand, U.; Jacobo-Herrera, N.; Altemimi, A.; Lakhssassi, N. A comprehensive review on medicinal plants as antimicrobial therapeutics: Potential avenues of biocompatible drug discovery. Metabolites 2019, 9, 258. [Google Scholar] [CrossRef]
- Catteau, L.; Zhu, L.; Van Bambeke, F.; Quetin-Leclercq, J. Natural and hemi-synthetic pentacyclic triterpenes as antimicrobials and resistance modifying agents against Staphylococcus aureus: A review. Phytochem. Rev. 2018, 17, 1129–1163. [Google Scholar] [CrossRef]
- da Silva, G.N.S.; Primon-Barros, M.; Macedo, A.J.; Gnoatto, S.C.B. Triterpene derivatives as relevant scaffold for new antibiofilm drugs. Biomolecules 2019, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Sibanda, T.; Okoh, A. The challenges of overcoming antibiotic resistance: Plant extracts as potential sources of antimicrobial and resistance modifying agents. Afr. J. Biotechnol. 2007, 6, 2886–2896. [Google Scholar]
- Wrońska, N.; Szlaur, M.; Zawadzka, K.; Lisowska, K. The synergistic effect of triterpenoids and flavonoids—New approaches for treating bacterial infections? Molecules 2022, 27, 847. [Google Scholar] [CrossRef] [PubMed]
- Romero, C.M.; Vivacqua, C.G.; Abdulhamid, M.B.; Baigori, M.D.; Slanis, A.C.; Allori, M.C.G.d.; Tereschuk, M.L. Biofilm inhibition activity of traditional medicinal plants from Northwestern Argentina against native pathogen and environmental microorganisms. Rev. Soc. Bras. Med. Trop. 2016, 49, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Sasirekha, B.; Megha, D.; Chandra, M.; Soujanya, R. Study on effect of different plant extracts on microbial biofilms. Asian J. Biotechnol. 2015, 7, 1–12. [Google Scholar] [CrossRef]
- Blair, J.M.; Piddock, L.J. How to measure export via bacterial multidrug resistance efflux pumps. mBio 2016, 7, e00840-16. [Google Scholar] [CrossRef]
- Kumar, R.; Jandaik, S.; Patial, P. Screening of medicinal plants of Himachal Pradesh for efflux pump inhibitory activity against Escherichia coli. J. Pharmacogn. Phytochem. 2016, 5, 96–100. [Google Scholar]
- World Health Organization. 2019 Antibacterial Agents in Clinical Development: An Analysis of the Antibacterial Clinical Development Pipeline; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Shriram, V.; Khare, T.; Bhagwat, R.; Shukla, R.; Kumar, V. Inhibiting bacterial drug efflux pumps via phyto-therapeutics to combat threatening antimicrobial resistance. Front. Microbiol. 2018, 9, 2990. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Secondary Metabolite | Result |
|---|---|
| Reducing sugars | + |
| Tannins | + |
| Flavonoids | + |
| Coumarins | + |
| Triterpenoids | + |
| Phytosterols | + |
| Saponins | − |
| Alkaloids | + |
| Position | LA1 | LA2 | ||
|---|---|---|---|---|
| δ C | δ H (J in Hz) | δ C | δ H (J in Hz) | |
| 1 | 202.8 (C) | - | 37.2 (CH2) | 1.81 |
| 2 | 60.6 (CH2) | 3.21, d (15.9 Hz) 3.43, d (15.9 Hz) | 31.6 (CH2) | 1.81 |
| 3 | 204.2 (C) | - | 71.8 (CH) | 3.50 |
| 4 | 59.0 (CH) | 2.55, q (6.7 Hz) | 42.2 (CH2) | 2.21, 2.27 |
| 5 | 37.8 (C) | - | 140.7 (C) | - |
| 6 | 40.5 (CH2) | 1.35, m 1.86 m (12. 5, 6.8, 3.2 Hz) | 121.7 (CH) | 5.33, d (4.7 Hz) |
| 7 | 18.0 (CH2) | 1.42, 1.51 | 31.9 (CH2) | 1.55, 1.56 |
| 8 | 52.1 (CH) | 1.22 | 31.9 (CH) | 1.42 |
| 9 | 37.13 (C) | - | 50.1 (CH) | 0.89 |
| 10 | 71.79 (CH) | 2.35, s | 36.5 (C) | - |
| 11 | 34.5 (CH2) | 1.12, m 2.12, m (13.6, 6.8, 3.5 Hz) | 21.0 (CH2) | 1.44, 1.47 |
| 12 | 30.1 (CH2) | 1.26, 1.38 | 39.7 (CH2) | 1.13, 1.99 |
| 13 | 39.4 (C) | - | 42.2 (C) | - |
| 14 | 38.2 (C) | - | 56.7 (CH) | 0.96 |
| 15 | 32.3 (CH2) | 1.50, m | 24.3 (CH2) | 1.03, 1.56 |
| 16 | 35.8 (CH2) | 1.33, 1.52 | 28.2 (CH2) | 1.26, 1.81 |
| 17 | 30.0 (C) | - | 56.0 (CH) | 1.08 |
| 18 | 42.6 (CH) | 1.54, m | 11.8 (CH3) | 0.65 s |
| 19 | 35.2 (CH2) | 1.32, m | 19.4 (CH3) | 0.98, s |
| 20 | 28.1 (C) | - | 36.1 (CH) | 1.32, m |
| 21 | 32.7 (CH2) | 1.24, m | 18.7 (CH3) | 0.90, d (6.5 Hz) |
| 22 | 39.2 (CH2) | 0.90, 1.47 | 33.9 (CH2) | 0.97, 1.30, dd (8.4, 15.0 Hz) |
| 23 | 7.3 (CH3) | 1.02, d (6.7 Hz) | 26.0 (CH2) | 1.13, dd (8.4, 15.0 Hz) |
| 24 | 15.9 (CH3) | 0.66, s | 45.8 (CH) | 0.90 |
| 25 | 18.0 (CH3) | 1.17, s | 29.1 (CH) | 1.65 |
| 26 | 20.3 (CH3) | 1.00, s | 19.0 (CH3) | 0.81, d (2.5 Hz) |
| 27 | 18.7 (CH3) | 0.99, s | 19.8 (CH3) | 0.79, d (6.8 Hz) |
| 28 | 32.0 (CH3) | 1.15, s | 23.0 (CH2) | 1.21, 1.25 |
| 29 | 31.7 (CH3) | 0.97, s | 12.0 (CH3) | 0.82, t (6.0 Hz) |
| 30 | 35.0 (CH3) | 0.91, s | 22.3 (CH2) | 0.84, m |
| Microorganism | Minimum Inhibitory Concentration (µg/mL) | ||||
|---|---|---|---|---|---|
| LA | LAPE | LAEt | LAM | Amox | |
| S. aureus | 125 | 500 | >500 | 250 | 3.91 |
| S. pyogenes | 500 | >500 | >500 | >500 | 1.95 |
| E. faecalis | 125 | >500 | 250 | >500 | 0.49 |
| P. aeruginosa | 500 | >500 | >500 | >500 | 500 |
| P. mirabilis | >500 | >500 | 250 | >500 | 31.25 |
| K. pneumoniae | 125 | >500 | 250 | >500 | 31.25 |
| S. typhi | 500 | >500 | 500 | >500 | 62.50 |
| E. coli | 500 | >500 | >500 | >500 | 125 |
| V. cholerae | 125 | 500 | 250 | 125 | 125 |
| Microorganism | Minimum Inhibitory Concentration (µg/mL) | ||
|---|---|---|---|
| LA1 | LA2 | Amoxicillin | |
| S. aureus | 31.25 | 31.25 | 10 |
| E. faecalis | 31.25 | 31.25 | 10 |
| E. coli | 62.5 | 125 | 20 |
| P. aeruginosa | 62.5 | 125 | >320 |
| Microorganism | MIC (µg/mL) | MIC Combined (µg/mL) | Modulation Factor | ||
|---|---|---|---|---|---|
| Amoxicillin Only | LA1 | LA2 | LA1 | LA2 | |
| P. aeruginosa | >320 | <31.25 | <31.25 | >10 | >10 |
| E. coli | 20 | <0.625 | <0.625 | >32 | >32 |
| MIC | Compound | % Inhibition of Efflux Pump Activity | |||
|---|---|---|---|---|---|
| 15 min | 30 min | 45 min | 60 min | ||
| 1/4 | LA1 | 111.0 ± 19.03 c | 108.4 ± 15.43 c | 100.0 ± 11.05 c | 87.1 ± 12.47 b |
| LA2LA2 | 109.6 ± 12.89 c | 105.4 ± 12.17 c | 95.4 ± 12.82 c | 86.3 ± 9.34 b | |
| 1/2 | LA1 | 59.5 ± 11.35 b | 45.3 ± 8.01 a | 34.6 ± 5.00 a | 29.1 ± 1.78 a |
| LA2 | 61.3 ± 12.55 b | 50.8 ± 6.35 b | 39.2 ± 12.55 a | 30.7 ± 7.10 a | |
| Verapamil | 60.3 ± 5.41 b | 120.3 ± 6.82 c | 125.8 ± 7.23 c | 113.4 ±8.29 c | |
| Chlorpromazine | 95.6 ± 7.91 b | 97.9 ± 15.04 b | 78.64 ± 5.78 b | 71.9 ± 4.49 b | |
| MIC | Compound | % Inhibition of Efflux Pump Activity | |||
|---|---|---|---|---|---|
| 15 min | 30 min | 45 min | 60 min | ||
| 1/4 | LA1 | 35.3 ± 26.88 a | 18.1 ± 10.87 a | 6.7 ± 10.69 a | 14.5 ± 14.21 a |
| LA2 | 6.7 ± 5.34 a | 2.2 ± 7.66 a | 0.5 ± 8.55 a | 8.7 ± 5.27 a | |
| 1/2 | LA1 | 41.7 ± 10.06 a | 19.3 ± 11.08 a | 13.22 ± 9.91 a | 13.4 ± 0.28 a |
| LA2 | 30.6 ± 11.04 a | 26.2 ± 8.94 a | 29.2 ± 11.20 a | 37.1 ± 9.67 a | |
| Verapamil | 147.3 ± 28.78 c | 145.5 ± 17.35 c | 129.3 ± 7.34 c | 128.4 ±10.89 c | |
| Chlorpromazine | 105.4 ± 9.02 c | 107.6 ± 9.03 c | 86.0 ± 3.86 b | 85.9 ± 9.91 b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anokwah, D.; Asante-Kwatia, E.; Asante, J.; Obeng-Mensah, D.; Danquah, C.A.; Amponsah, I.K.; Ameyaw, E.O.; Biney, R.P.; Obese, E.; Oberer, L.; et al. Antibacterial, Resistance Modulation, Anti-Biofilm Formation, and Efflux Pump Inhibition Properties of Loeseneriella africana (Willd.) N. Halle (Celastraceae) Stem Extract and Its Constituents. Microorganisms 2024, 12, 7. https://doi.org/10.3390/microorganisms12010007
Anokwah D, Asante-Kwatia E, Asante J, Obeng-Mensah D, Danquah CA, Amponsah IK, Ameyaw EO, Biney RP, Obese E, Oberer L, et al. Antibacterial, Resistance Modulation, Anti-Biofilm Formation, and Efflux Pump Inhibition Properties of Loeseneriella africana (Willd.) N. Halle (Celastraceae) Stem Extract and Its Constituents. Microorganisms. 2024; 12(1):7. https://doi.org/10.3390/microorganisms12010007
Chicago/Turabian StyleAnokwah, Daniel, Evelyn Asante-Kwatia, Jonathan Asante, Daniel Obeng-Mensah, Cynthia Amaning Danquah, Isaac Kingsley Amponsah, Elvis Ofori Ameyaw, Robert Peter Biney, Ernest Obese, Lukas Oberer, and et al. 2024. "Antibacterial, Resistance Modulation, Anti-Biofilm Formation, and Efflux Pump Inhibition Properties of Loeseneriella africana (Willd.) N. Halle (Celastraceae) Stem Extract and Its Constituents" Microorganisms 12, no. 1: 7. https://doi.org/10.3390/microorganisms12010007
APA StyleAnokwah, D., Asante-Kwatia, E., Asante, J., Obeng-Mensah, D., Danquah, C. A., Amponsah, I. K., Ameyaw, E. O., Biney, R. P., Obese, E., Oberer, L., Amoako, D. G., Abia, A. L. K., & Mensah, A. Y. (2024). Antibacterial, Resistance Modulation, Anti-Biofilm Formation, and Efflux Pump Inhibition Properties of Loeseneriella africana (Willd.) N. Halle (Celastraceae) Stem Extract and Its Constituents. Microorganisms, 12(1), 7. https://doi.org/10.3390/microorganisms12010007