

Assessment of Potential Anti-Methanogenic and Antimicrobial Activity of Ethyl Nitroacetate, α-Lipoic Acid, Taurine and L-Cysteinesulfinic Acid In Vitro
 , ,
, ,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Comparison of Antimicrobial Effects against Escherichia coli O157:H7 and Salmonella enterica Serovar Typhimurium
2.2. Comparison of Effects of Treatments on In Vitro Rumen Methane Production and Fermentation
2.3. Analytical
2.4. Statistics
3. Results
3.1. Antimicrobial Effects on E. coli and Salmonella
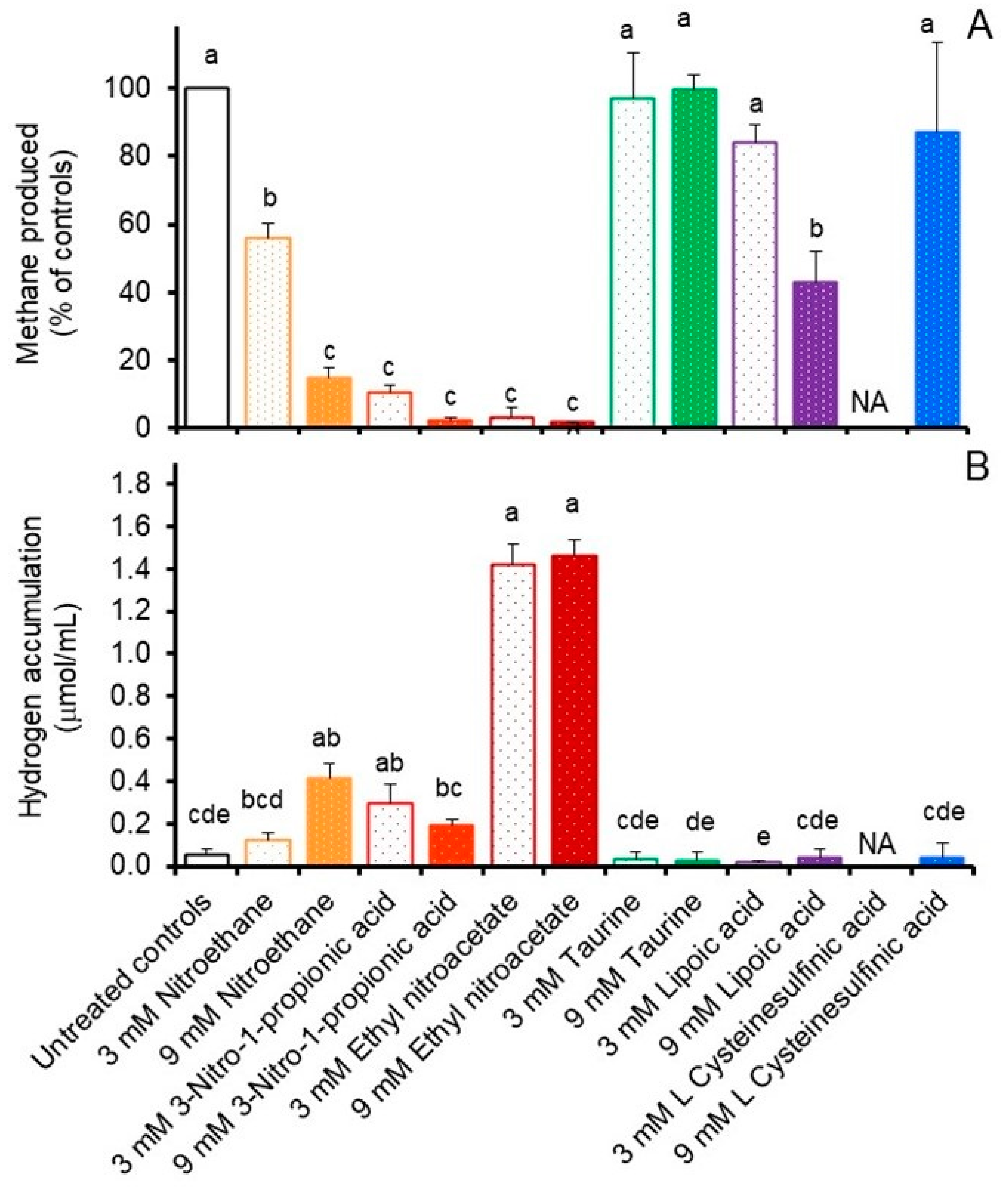
3.2. Comparative Effects of Inhibitors on In Vitro Rumen Methane Production and Fermentation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, K.A.; Johnson, D.E. Methane emissions from cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
- Van Nevel, C.J.; Demeyer, D.I. Control of rumen methanogenesis. Environ. Monit. Assess. 1996, 42, 73–97. [Google Scholar] [CrossRef]
- Anderson, R.C.; Jung, Y.S.; Oliver, C.E.; Horrocks, S.M.; Genovese, K.J.; Harvey, R.B.; Callaway, T.R.; Edrington, T.S.; Nisbet, D.J. Effects of nitrate or nitro-supplementation, with or without added chlorate, on Salmonella enterica serovar Typhimurium and Escherichia coli in swine feces. J. Food Prot. 2007, 70, 308–315. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bozic, A.K.; Anderson, R.C.; Callaway, T.R.; Nisbet, D.J.; Ricke, S.C.; Crandall, P.G.; O’Bryan, C.A. In vitro comparison of nitroethane, 2-nitro-1-propanol, lauric acid, LauricidinTM and the Hawaiian marine algae, Chaetoceros activity against anaerobically grown Staphylococcus aureus. Int. J. Appl. Res. Vet. Med. 2010, 8, 180–184. [Google Scholar]
- Bozic, A.K.; Anderson, R.C.; Ricke, S.C.; Crandall, P.G.; O’Bryan, C.A. Comparison of nitroethane, 2-nitro-1-propanol, lauric acid, LauricidinTM and the Hawaiian marine algae, Chaetoceros, for potential broad-spectrum control of anaerobically grown lactic acid bacteria. J. Environ. Sci. Health Part B 2012, 47, 269–272. [Google Scholar] [CrossRef] [PubMed]
- Božić, A.; Anderson, R.C.; Arzola-Alvarez, C.; Ruiz-Barrera, O.; Corral-Luna, A.; Castillo-Castillo, Y.; Arzola-Rubio, A.; Poole, T.L.; Harvey, R.B.; Hume, M.E.; et al. Inhibition of multidrug-resistant Staphylococci by sodium chlorate and select nitro- and medium chain fatty acid compounds. J. Appl. Microbiol. 2019, 126, 1508–1518. [Google Scholar] [CrossRef] [PubMed]
- Dimitrijevic, M.; Anderson, R.C.; Callaway, T.R.; Jung, Y.S.; Harvey, R.B.; Ricke, S.C.; Nisbet, D.J. Inhibitory effect of select nitrocompounds on growth and survivability of Listeria monocytogenes in vitro. J. Food Prot. 2006, 69, 1061–1065. [Google Scholar] [CrossRef]
- Horrocks, S.M.; Jung, Y.S.; Huwe, J.K.; Harvey, R.B.; Ricke, S.C.; Carstens, G.E.; Callaway, T.R.; Anderson, R.C.; Ramlachan, N.; Nisbet, D.J. Effects of short-chain nitrocompounds against Campylobacter jejuni and Campylobacter coli in vitro. J. Food Sci. 2007, 72, M50–M55. [Google Scholar] [CrossRef]
- Jung, Y.S.; Anderson, R.C.; Callaway, T.R.; Edrington, T.S.; Genovese, K.J.; Harvey, R.B.; Poole, T.L.; Nisbet, D.J. Inhibitory activity of 2-nitropropanol against select foodborne pathogens in vitro. Lett. Appl. Microbiol. 2004, 39, 471–476. [Google Scholar] [CrossRef]
- Jung, Y.S.; Anderson, R.C.; Edrington, T.S.; Genovese, K.J.; Byrd, J.A.; Callaway, T.R.; Nisbet, D.J. Experimental use of 2-nitropropanol for reduction of Salmonella Typhimurium in the ceca of broiler chicks. J. Food Prot. 2004, 67, 1945–1947. [Google Scholar] [CrossRef]
- Ruiz-Barrera, O.; Anderson, R.C.; Hume, M.E.; Corrales-Millan, J.; Castillo-Castillo, Y.; Corral-Luna, A.; Guevara-Valdez, J.L.; Salinas-Chavira, J.; Rodriguez-Muela, C.; Arzola-Alvarez, C. Short chain nitrocompounds as a treatment of layer hen manure and litter; effects on in vitro survivability of Salmonella, generic E. coli and nitrogen metabolism. J. Environ. Sci. Health Part B 2017, 52, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Beauchemin, K.A.; Dong, R. A review of 3-nitrooxypropanol for enteric methane mitigation from ruminant livestock. Animals 2021, 11, 3540. [Google Scholar] [CrossRef] [PubMed]
- Latham, E.A.; Anderson, R.C.; Pinchak, W.E.; Nisbet, D.J. Insights on alterations to the rumen ecosystem by nitrate and nitrocompounds. Front. Microbiol. 2016, 7, 228. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Krueger, N.A.; Stanton, T.B.; Callaway, T.R.; Edrington, T.S.; Harvey, R.B.; Jung, Y.S.; Nisbet, D.J. Effects of select nitrocompounds on in vitro ruminal fermentation during conditions of limiting or excess added reductant. Bioresour. Technol. 2008, 99, 8655–8661. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Ripley, L.H.; Bowman, J.G.P.; Callaway, T.R.; Genovese, K.J.; Beier, R.C.; Harvey, R.B.; Nisbet, D.J. Ruminal fermentation of anti-methanogenic nitrate- and nitro-containing forages in vitro. Front. Veter Sci. 2016, 3, 62. [Google Scholar] [CrossRef] [PubMed]
- Sawers, G. The hydrogenases and formate dehydrogenases of Escherichia coli. Antonie Van Leeuwenhoek 1994, 66, 57–88. [Google Scholar] [CrossRef] [PubMed]
- Pinske, C.; Sawers, R.G. Anaerobic formate and hydrogen metabolism. EcoSal Plus 2016, 7. [Google Scholar] [CrossRef]
- Angermaier, L.; Simon, H. On the reduction of aliphatic and aromatic nitro compounds by Clostridia, the role of ferredoxin and its stabilization. Hoppe-Seyler’s Z. Physiol. Chem. 1983, 364, 961–975. [Google Scholar] [CrossRef]
- Lamichhane-Khadka, R.; Benoit, S.; Miller-Parks, E.F.; Maier, R.J. Host hydrogen rather than that produced by the pathogen is important for Salmonella enterica serovar Typhimurium virulence. Infect. Immun. 2015, 83, 311–316. [Google Scholar] [CrossRef]
- Maier, L.; Barthel, M.; Stecher, B.; Maier, R.J.; Gunn, J.S.; Hardt, W.-D. Salmonella Typhimurium strain ATCC14028 requires H2-hydrogenases for growth in the gut, but not at systemic sites. PLoS ONE 2014, 9, e110187. [Google Scholar] [CrossRef]
- Maier, R.J.; Olczak, A.; Maier, S.; Soni, S.; Gunn, J. Respiratory hydrogen use by Salmonella enterica serovar Typhimurium is essential for virulence. Infect. Immun. 2014, 72, 6294–6299. [Google Scholar] [CrossRef] [PubMed]
- Francioso, A.; Baseggio Conrado, A.; Mosca, L.; Fontana, M. Chemistry and biochemistry of sulfur natural compounds: Key intermediates of metabolism and redox biology. Oxidative Med. Cell. Longev. 2020, 2020, 8294158. [Google Scholar] [CrossRef] [PubMed]
- Ruhee, R.T.; Roberts, L.A.; Ma, S.; Suzuki, K. Organosulfur compounds: A review of their anti-inflammatory effects in human health. Front. Nutr. 2020, 7, 64. [Google Scholar] [CrossRef] [PubMed]
- Bhatwalkar, S.B.; Mondal, R.; Krishna, S.B.N.; Adam, J.K.; Govender, P.; Anupam, R. Antibacterial properties of organosulfur compounds of garlic (Allium sativum). Front. Microbiol. 2021, 12, 613077. [Google Scholar] [CrossRef] [PubMed]
- Dufour, V.; Stahl, M.; Baysse, C. The antibacterial properties of isothiocyanates. Microbiology 2015, 161, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, S.; Kim, H.W. Effects and mechanisms of taurine as a therapeutic agent. Biomol. Ther. 2018, 26, 225–241. [Google Scholar] [CrossRef]
- Shi, C.; Sun, Y.; Zhang, X.; Zheng, Z.; Yang, M.; Ben, H.; Song, K.; Cao, Y.; Chen, Y.; Liu, X.; et al. Antimicrobial effect of lipoic acid against Cronobacter sakazakii. Food Control. 2016, 59, 352–358. [Google Scholar] [CrossRef]
- Majak, W.; Cheng, K.-J.; Hall, J.W. Enhanced degradation of 3-nitropropanol by ruminal microorganisms. J. Anim. Sci. 1986, 62, 1072–1080. [Google Scholar] [CrossRef][Green Version]
- Nisbet, D.J.; Callaway, T.R.; Edrington, T.S.; Anderson, R.C.; Poole, T.L. Effects of ionophores on Enterococcus faecalis and E. faecium growth in pure and mixed ruminal culture. Foodborne Pathog. Dis. 2008, 5, 193–198. [Google Scholar] [CrossRef]
- Leyendecker, S.A.; Callaway, T.R.; Anderson, R.C.; Nisbet, D.J. Technical note on a much simplified method for collecting ruminal fluid using a nylon paint strainer. J. Sci. Food Agric. 2004, 84, 387–389. [Google Scholar] [CrossRef]
- Allison, M.J.; Mayberry, W.R.; McSweeney, C.S.; Stahl, D.A. Synergistes jonesii, gen. nov., sp. nov.: A ruminal bacterium that degrades toxic pyridinediols. System. Appl. Microbiol. 1992, 15, 522–529. [Google Scholar] [CrossRef]
- Lambert, M.A.; Moss, C.W. Preparation and analysis of the butyl esters of short-chain volatile and non-volatile fatty acids. Advan. Chrom. 1972, 74, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Salanitro, J.P.; Muirhead, P.A. Quantitative method for the gas chromatographic analysis of short-chain monocarboxylic and dicarboxylic acids in fermentation media. Appl. Environ. Microbiol. 1975, 29, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Ungerfeld, E.M.; Rust, S.R.; Burnett, R. Use of some novel alternative electron sinks to inhibit ruminal methanogenesis. Reprod. Nutr. Dev. 2003, 43, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Chalupa, W. Manipulating rumen fermentation. J. Anim. Sci. 1977, 46, 585–599. [Google Scholar] [CrossRef]
- JMP®Pro, version 15.1; SAS Institute Inc.: Cary, NC, USA, 2020.
- Teng, P.-Y.; Kim, W.K. Roles of nitrocompounds in inhibition of foodborne bacteria, parasites, and methane production in economic animals. Animals 2021, 11, 923. [Google Scholar] [CrossRef]
- Correa, A.C.; Trachsel, J.; Allen, H.K.; Corral-Luna, A.; Gutierrez-Bañuelos, H.; Ochoa-Garcia, P.A.; Ruiz-Barrera, O.; Hume, M.E.; Callaway, T.R.; Harvey, R.B.; et al. Effect of sole or combined administration of nitrate and 3-nitro-1-propionic acid on fermentation and salmonella survivability in alfalfa-fed rumen cultures in vitro. Bioresour. Technol. 2017, 229, 69–77. [Google Scholar] [CrossRef]
- Ochoa-García, P.A.; Anderson, R.C.; Arévalos-Sánchez, M.M.; Rodríguez-Almeida, F.A.; Félix-Portillo, M.; Muro-Reyes, A.; Božić, A.K.; Arzola-Álvarez, C.; Corral-Luna, A. Astragallus mollissimus plant extract: A strategy to reduce ruminal methanogenesis. Trop. Anim. Health Prod. 2021, 53, 436. [Google Scholar] [CrossRef]
- Anderson, R.C.; Rasmussen, M.A. Use of a novel nitrotoxin-metabolizing bacterium to reduce ruminal methane production. Bioresour. Technol. 1998, 64, 89–95. [Google Scholar] [CrossRef]
- Anderson, R.C.; Callaway, T.R.; Van Kessel, J.S.; Jung, Y.S.; Edrington, T.S.; Nisbet, D.J. Effect of select nitrocompounds on ruminal fermentation; an initial look at their potential to reduce economic and environmental costs associated with ruminal methanogenesis. Bioresour. Technol. 2003, 90, 59–63. [Google Scholar] [CrossRef]
- Greening, C.; Geier, R.; Wang, C.; Woods, L.C.; Morales, S.E.; McDonald, M.J.; Rushton-Green, R.; Morgan, X.C.; Koike, S.; Leahy, S.C.; et al. Diverse hydrogen production and consumption pathways influence methane production in ruminants. ISME J. 2019, 13, 2617–2632. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Huwe, J.K.; Smith, D.J.; Stanton, T.B.; Krueger, N.A.; Callaway, T.R.; Edrington, T.S.; Harvey, R.B.; Nisbet, D.J. Effect of nitroethane, dimethyl-2-nitroglutarate and 2-nitro-methyl-propionate on ruminal methane production and hydrogen balance in vitro. Bioresour. Technol. 2010, 101, 5345–5349. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Rasmussen, M.A.; Jensen, N.S.; Allison, M.J. Denitrobacterium detoxificans gen. nov., sp. nov., a ruminal bacterium that respires on nitrocompounds. Int. J. Syst. Evol. Microbiol. 2000, 50, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Rasmussen, M.A.; Allison, M.J. Enrichment and isolation of a nitropropanol-metabolizing bacterium from the rumen. Appl. Environ. Microbiol. 1996, 62, 3885–3886. [Google Scholar] [CrossRef] [PubMed]
- Cronan, J.E.; Zhao, X.; Jiang, Y. Function, attachment and synthesis of lipoic acid in Escherichia coli. Adv. Microb. Physiol. 2005, 50, 103–146. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Du, Y.; Li, X.; Yao, C. Lipoic acid modified antimicrobial peptide with enhanced antimicrobial properties. Bioorganic Med. Chem. 2020, 28, 115682. [Google Scholar] [CrossRef] [PubMed]
- LibreTextsTM Chemistry. 18.8 Thiols and Sulfides; LibreTextsTM Chemistry: Davis, CA, USA; Available online: https://chem.libretexts.org/Bookshelves/Organic_Chemistry/Organic_Chemistry_(Morsch_et_al.)/18%253A_Ethers_and_Epoxides_Thiols_and_Sulfides/18.08%253A_Thiols_and_Sulfides (accessed on 21 September 2023).
- Zhang, S.; Liang, Q.; Li, M.; Zhao, G. Ruminal degradation of taurine and its effects on rumen fermentation in vitro. Fermentation 2023, 9, 43. [Google Scholar] [CrossRef]
- Biewenga, G.P.; Dorstijn, M.A.; Verhagen, J.V.; Haenen, G.R.; Bast, A. Reduction of lipoic acid by lipoamide dehydrogenase. Biochem. Pharmacol. 1996, 51, 233–238. [Google Scholar] [CrossRef]
- Hedderich, R.; Klimmek, O.; Kröger, A.; Dirmeier, R.; Keller, M.; Stetter, K.O. Anaerobic respiration with elemental sulfur and with disulfides. FEMS Microbiol. Rev. 1998, 22, 353–381. [Google Scholar] [CrossRef]
- Bal, M.A.; Ozturk, D. Effects of sulfur containing supplements on ruminal fermentation and microbial protein synthesis. Res. J. Anim. Vet. Sci. 2006, 1, 33–36. [Google Scholar]
- Hong, Y.; Gu, J.-D. Bacterial anaerobic respiration and electron transfer relevant to the biotransformation of pollutants. Int. Int. Biodeterior. Biodegrad. 2009, 63, 973–980. [Google Scholar] [CrossRef]
- Lie, T.J.; Pitta, T.; Leadbetter, E.R.; Godchaux, W., III; Leadbetter, J.R. Sulfonates: Novel electron acceptors in anaerobic respiration. Arch. Microbiol. 1996, 166, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Lie, T.J.; Godchaux, W.; Leadbetter, E.R. Sulfonates as terminal electron acceptors for growth of sulfite-reducing bacteria (Desulfitobacterium spp.) and sulfate-reducing bacteria: Effects of inhibitors of sulfidogenesis. Appl. Environ. Microbiol. 1999, 65, 4611–4617. [Google Scholar] [CrossRef] [PubMed]
- Stewart, L. Mineral Supplements for Beef Cattle; The University of Georgia: Athens, GA, USA, 2017. [Google Scholar]
- Mazumder, T.K.; Nishio, N.; Fukuzaki, S.; Nagai, S. Effect of sulfur-containing compounds on growth of Methanosarcina barkeri in defined medium. Appl. Environ. Microbiol. 1986, 52, 617–622. [Google Scholar] [CrossRef]
- Burrichter, A.G.; Dörr, S.; Bergmann, P.; Haiß, S.; Keller, A.; Fournier, C.; Franchini, P.; Isono, E.; Schleheck, D. Bacterial microcompartments for isethionate desulfonation in the taurine-degrading human-gut bacterium Bilophila wadsworthia. BMC Microbiol. 2021, 21, 340. [Google Scholar] [CrossRef]
- Kandylis, K. Toxicology of sulfur in ruminants: Review. J. Dairy Sci. 1984, 67, 2179–2187. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| None | Ethyl Nitroacetate | Taurine | α-Lipoic Acid | L-Cysteine- Sulfinic Acid | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 mM | 9 mM | 3 mM | 9 mM | 3 mM | 9 mM | 3 mM | 9 mM | p | SEM | ||
| Growth rate (h−1) | |||||||||||
| E. coli O157:H7 933 | 0.823 a | 0.639 bc | 0.527 c | 0.777 a | 0.740 ab | 0.568 c | 0.224 d | 0.809 a | 0.809 a | <0.0001 | 0.033 |
| S. Typhimurium DT104 | 0.886 a | 0.744 ab | 0.656 bc | 0.872 a | 0.920 a | 0.522 c | 0.195 d | 0.928 a | 0.888 a | <0.0001 | 0.048 |
| Maximum OD (600 nm) | |||||||||||
| E. coli O157:H7 933 | 0.448 a | 0.454 a | 0.447 a | 0.452 a | 0.456 a | 0.448 a | 0.337 b | 0.449 a | 0.451 a | <0.0001 | 0.006 |
| S. Typhimurium DT104 | 0.451 a | 0.451 a | 0.430 b | 0.451 a | 0.449 a | 0.445 a | 0.262 b | 0.449 a | 0.448 a | <0.0001 | 0.006 |
| Ending pH | |||||||||||
| E. coli O157:H7 933 | 6.45 b | 6.48 ab | 6.46 ab | 6.47 ab | 6.48 a | 6.49 a | 6.47 ab | 6.45 b | 6.40 c | <0.0001 | 0.006 |
| None | Ethyl Nitracetate | α-Lipoic Acid | Both at | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Measured Variable 1 | 3 mM | 9 mM | 3 mM | 9 mM | 3 mM | 9 mM | p | SEM | |
| Total gas (mL) | 6.8 | 3.5 | 4.2 | 4.2 | 5.2 | 5.2 | 3.8 | 0.8123 | 1.708 |
| Hydrogen (µmol/mL) | 0.14 | 1.23 | 0.66 | 1.17 | 0.23 | 1.43 | 0.84 | 0.2327 | 0.442 |
| Methane (µmol/mL) | 16.36 a | 0.53 b | 0.28 b | 5.63 b | 8.71 ab | 0.68 b | 0.40 b | 0.0006 | 1.928 |
| Total acids (µmol/mL) | 51.29 | 56.10 | 49.52 | 30.34 | 45.70 | 30.38 | 41.82 | 0.0664 | 6.680 |
| Acetate (µmol/mL) | 33.19 a | 33.04 a | 27.09 ab | 18.67 ab | 29.54 a | 16.13 ab | 23.51 ab | 0.0472 | 4.227 |
| Propionate (µmol/mL) | 11.85 | 14.90 | 16.30 | 7.90 | 10.17 | 9.43 | 12.42 | 0.0657 | 1.905 |
| Butyrate (µmol/mL) | 4.68 ab | 6.19 a | 4.83 ab | 2.78 b | 4.35 ab | 3.62 ab | 4.57 ab | 0.0257 | 0.596 |
| Valerate (µmol/mL) | 0.94 ab | 1.17 a | 0.72 bc | 0.70 bc | 0.94 ab | 0.82 bc | 0.56 c | 0.0012 | 0.073 |
| Isobutyrate (µmol/mL) | 0.27 | 0.34 | 0.24 | 0.12 | 0.28 | 0.15 | 0.28 | 0.0650 | 0.052 |
| Isovalerate (µmol/mL) | 0.35 | 0.46 | 0.33 | 0.18 | 0.41 | 0.23 | 0.47 | 0.0620 | 0.070 |
| Acetate to propionate ratio | 2.80 a | 2.22 bcd | 1.72 cd | 2.36 ab | 2.88 a | 1.66 d | 1.88 bcd | 0.0001 | 0.137 |
| None | Ethyl Nitracetate | α-Lipoic Acid | Both at | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Stoichiometric Estimate | 3 mM | 9 mM | 3 mM | 9 mM | 3 mM | 9 mM | p | SEM | |
| e− produced (µmol H2/mL) 1 | 99.79 | 109.28 | 91.97 | 58.46 | 89.48 | 58.65 | 79.41 | 0.0562 | 12.646 |
| e− consumed (µmol H2/mL) 1 | 99.61 a | 46.71 bc | 44.75 bc | 45.76bc | 65.03 ab | 30.06 c | 37.00 bc | 0.0040 | 9.203 |
| Observed e− recovery (%) | 101.58 a | 42.86 b | 48.26 b | 78.03ab | 73.40 ab | 55.05 b | 46.97 b | 0.0068 | 9.208 |
| Theoretical e− recovery (%) | 101.58 | 51.13 | 78.36 | 83.21 | 84.22 | 77.17 | 93.17 | 0.0734 | 10.764 |
| Hexose fermented (µmol/mL) 2 | 28.14 | 31.34 | 27.25 | 16.76 | 25.15 | 17.27 | 23.10 | 0.0614 | 3.601 |
| Fermentation efficiency (%) 2 | 74.72c | 76.77 bc | 79.26 ab | 76.19 bc | 74.52 c | 79.59 a | 78.28 ab | 0.0010 | 0.728 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levent, G.; Božić, A.; Petrujkić, B.T.; Callaway, T.R.; Poole, T.L.; Crippen, T.L.; Harvey, R.B.; Ochoa-García, P.; Corral-Luna, A.; Yeater, K.M.; et al. Assessment of Potential Anti-Methanogenic and Antimicrobial Activity of Ethyl Nitroacetate, α-Lipoic Acid, Taurine and L-Cysteinesulfinic Acid In Vitro. Microorganisms 2024, 12, 34. https://doi.org/10.3390/microorganisms12010034
Levent G, Božić A, Petrujkić BT, Callaway TR, Poole TL, Crippen TL, Harvey RB, Ochoa-García P, Corral-Luna A, Yeater KM, et al. Assessment of Potential Anti-Methanogenic and Antimicrobial Activity of Ethyl Nitroacetate, α-Lipoic Acid, Taurine and L-Cysteinesulfinic Acid In Vitro. Microorganisms. 2024; 12(1):34. https://doi.org/10.3390/microorganisms12010034
Chicago/Turabian StyleLevent, Gizem, Aleksandar Božić, Branko T. Petrujkić, Todd R. Callaway, Toni L. Poole, Tawni L. Crippen, Roger B. Harvey, Pedro Ochoa-García, Agustin Corral-Luna, Kathleen M. Yeater, and et al. 2024. "Assessment of Potential Anti-Methanogenic and Antimicrobial Activity of Ethyl Nitroacetate, α-Lipoic Acid, Taurine and L-Cysteinesulfinic Acid In Vitro" Microorganisms 12, no. 1: 34. https://doi.org/10.3390/microorganisms12010034
APA StyleLevent, G., Božić, A., Petrujkić, B. T., Callaway, T. R., Poole, T. L., Crippen, T. L., Harvey, R. B., Ochoa-García, P., Corral-Luna, A., Yeater, K. M., & Anderson, R. C. (2024). Assessment of Potential Anti-Methanogenic and Antimicrobial Activity of Ethyl Nitroacetate, α-Lipoic Acid, Taurine and L-Cysteinesulfinic Acid In Vitro. Microorganisms, 12(1), 34. https://doi.org/10.3390/microorganisms12010034