
Application of Fourier Transform Infrared Spectroscopy to Discriminate Two Closely Related Bacterial Species: Bacillus anthracis and Bacillus cereus Sensu Stricto

 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. IR Biotyper
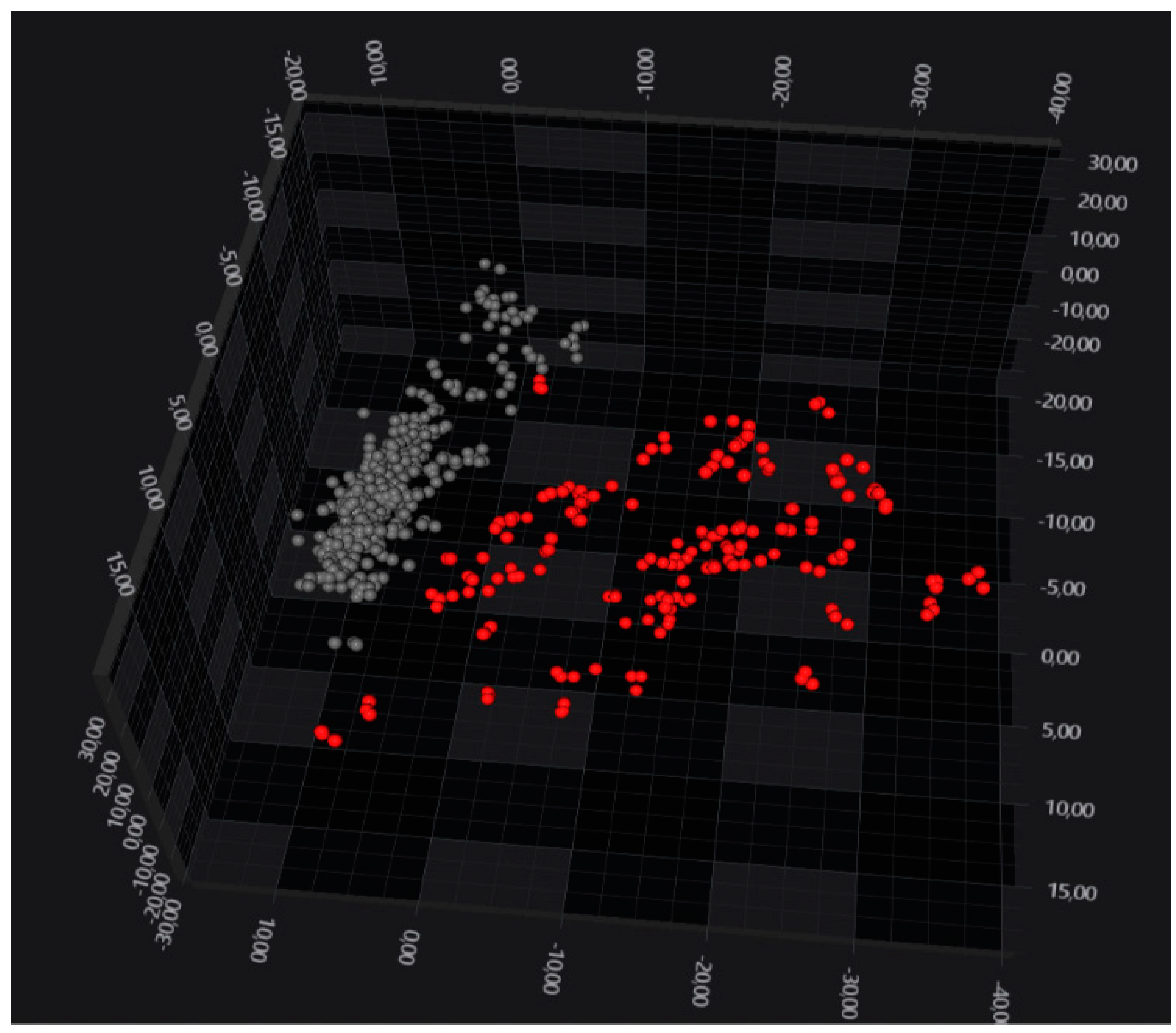
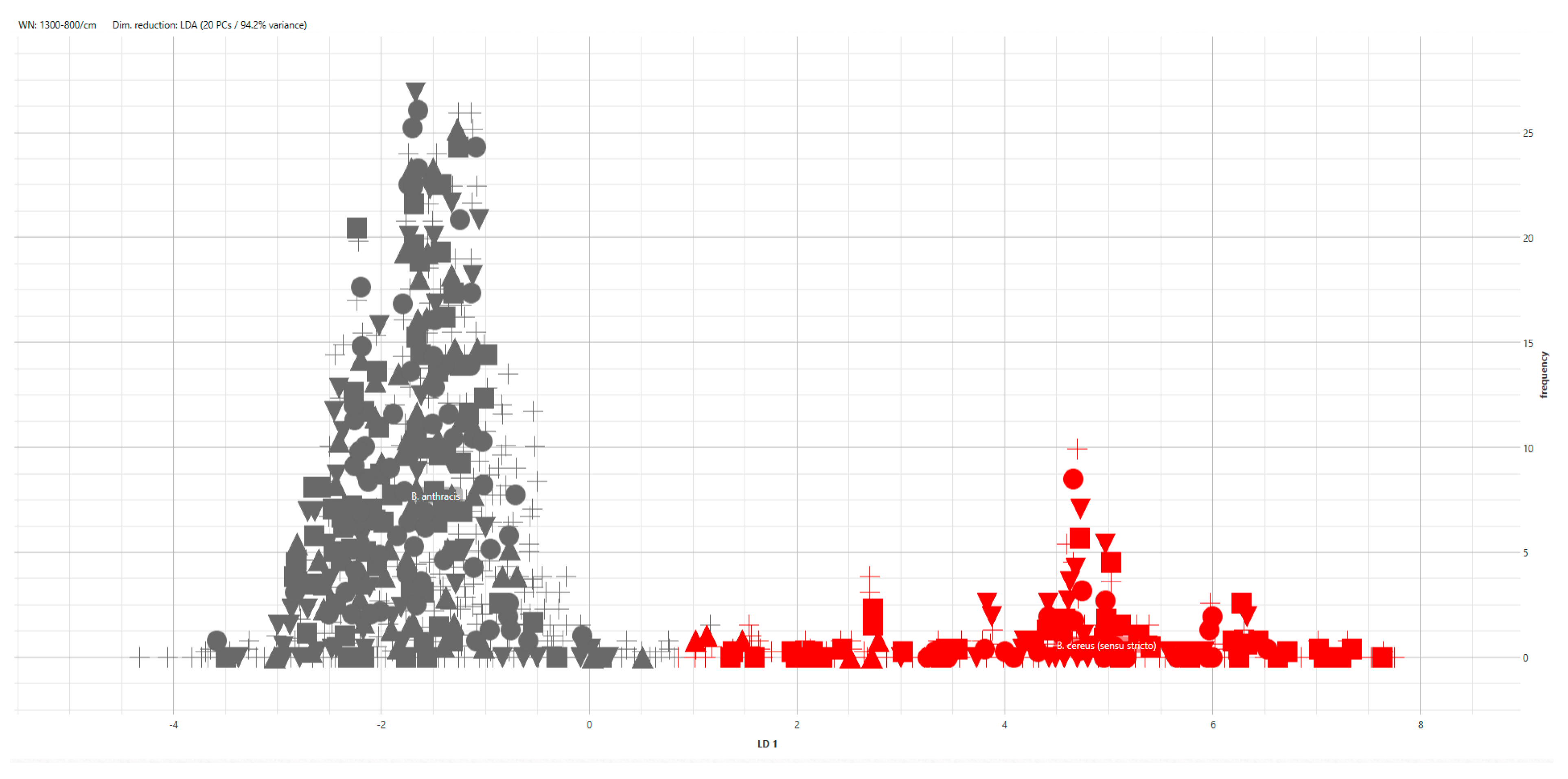
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Turnbull, P.C. Introduction: Anthrax history, disease and ecology. Curr. Top. Microbiol. Immunol. 2002, 271, 1–19. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Anthrax in Humans and Animals, 4th ed.; World Health Organization: Geneva, Switzerland, 2008. [Google Scholar]
- Pilo, P.; Frey, J. Pathogenicity, population genetics and dissemination of Bacillus anthracis. Infect. Genet. Evol. 2018, 64, 115–125. [Google Scholar] [CrossRef]
- Moayeri, M.; Leppla, S.H.; Vrentas, C.; Pomerantsev, A.P.; Liu, S. Anthrax Pathogenesis. Annu. Rev. Microbiol. 2015, 69, 185–208. [Google Scholar] [CrossRef] [PubMed]
- Hicks, C.W.; Sweeney, D.A.; Cui, X.; Li, Y.; Eichacker, P.Q. An overview of anthrax infection including the recently identified form of disease in injection drug users. Intensive Care Med. 2012, 38, 1092–1104. [Google Scholar] [CrossRef] [PubMed]
- Drobniewski, F.A. Bacillus cereus and related species. Clin. Microbiol. Rev. 1993, 6, 324–338. [Google Scholar] [CrossRef]
- Stenfors Arnesen, L.P.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef]
- Schoeni, J.L.; Wong, A.C.L. Bacillus cereus food poisoning and its toxins. J. Food Prot. 2005, 68, 636–648. [Google Scholar] [CrossRef]
- Manzulli, V.; Rondinone, V.; Buchicchio, A.; Serrecchia, L.; Cipolletta, D.; Fasanella, A.; Parisi, A.; Difato, L.; Iatarola, M.; Aceti, A.; et al. Discrimination of Bacillus cereus Group Members by MALDI-TOF Mass Spectrometry. Microorganisms 2021, 9, 1202. [Google Scholar] [CrossRef]
- Kamnev, A.A.; Tugarova, A.V. Specificities of the Fourier Transform Infrared Spectroscopic Methodology and Interpretation of Spectroscopic Data in Microbiological Analyses. J. Anal. Chem. 2023, 78, 1320–1332. [Google Scholar] [CrossRef]
- Novais, Â.; Freitas, A.R.; Rodrigues, C.; Peixe, L. Fourier transform infrared spectroscopy: Unlocking fundamentals and prospects for bacterial strain typing. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 38, 427–448. [Google Scholar] [CrossRef]
- Baker, M.J.; Trevisan, J.; Bassan, P.; Bhargava, R.; Butler, H.J.; Dorling, K.M.; Fielden, P.R.; Fogarty, S.W.; Fullwood, S.J.; Heys, K.A.; et al. Using Fourier transform IR spectroscopy to analyze biological materials. Nat. Protoc. 2014, 9, 1771–1791. [Google Scholar] [CrossRef] [PubMed]
- Uribe, G.; Salipante, S.J.; Curtis, L.; Lieberman, J.A.; Kurosawa, K.; Cookson, B.T.; Hoogestraat, D.; Stewart, M.K.; Olmstead, T.; Bourassa, L. Evaluation of Fourier transform-infrared spectroscopy (FT-IR) as a control measure for nosocomial outbreak investigations. J. Clin. Microbiol. 2023, 61, e0034723. [Google Scholar] [CrossRef] [PubMed]
- Tiquia-Arashiro, S.; Li, X.; Pokhrel, K.; Kassem, A.; Abbas, L.; Coutinho, O.; Kasperek, D.; Najaf, H.; Opara, S. Applications of Fourier Transform-Infrared spectroscopy in microbial cell biology and environmental microbiology: Advances, challenges, and future perspectives. Front. Microbiol. 2023, 14, 1304081. [Google Scholar] [CrossRef]
- Cordovana, M.; Mauder, N.; Join-Lambert, O.; Gravey, F.; LeHello, S.; Auzou, M.; Pitti, M.; Zoppi, S.; Buhl, M.; Steinmann, J.; et al. Machine learning-based typing of Salmonella enterica O-serogroups by the Fourier-Transform Infrared (FTIR) Spectroscopy-based IR Biotyper system. J. Microbiol. Methods 2022, 201, 106564. [Google Scholar] [CrossRef] [PubMed]
- Savini, F.; Romano, A.; Giacometti, F.; Indio, V.; Pitti, M.; Decastelli, L.; Devalle, P.L.; Gorrasi, I.S.R.; Miaglia, S.; Serraino, A. Investigation of a Staphylococcus aureus sequence type 72 food poisoning outbreak associated with food-handler contamination in Italy. Zoonoses Public Health 2023, 70, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.Y.; Kim, Y.A.; Lee, S.J.; Jung, W.W.; Kim, H.S.; Kim, S.S.; Kim, H.; Yong, D.; Lee, K. Performance Comparison Between Fourier-Transform Infrared Spectroscopy-based IR Biotyper and Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry for Strain Diversity. Ann. Lab. Med. 2023, 43, 174–179. [Google Scholar] [CrossRef]
- Azrad, M.; Matok, L.A.; Leshem, T.; Peretz, A. Comparison of FT-IR with whole-genome sequencing for identification of maternal-to-neonate transmission of antibiotic-resistant bacteria. J. Microbiol. Methods. 2022, 202, 106603. [Google Scholar] [CrossRef]
- Contreras, D.A.; Morgan, M.A. Surveillance diagnostic algorithm using real-time PCR assay and strain typing method development to assist with the control of C. auris amid COVID-19 pandemic. Front. Cell Infect. Microbiol. 2022, 12, 887754. [Google Scholar] [CrossRef]
- Kon, H.; Temkin, E.; Elmalih, P.; Keren-Paz, A.; Ben-David, D.; Najjar-Debbiny, R.; Gottesman, T.; Carmeli, Y. Analysis of four carbapenem-resistant Acinetobacter baumannii outbreaks using Fourier-transform infrared spectroscopy. Infect. Control Hosp. Epidemiol. 2023, 44, 991–993. [Google Scholar] [CrossRef]
- Hu, Y.; Zhou, H.; Lu, J.; Sun, Q.; Liu, C.; Zeng, Y.; Zhang, R. Evaluation of the IR Biotyper for Klebsiella pneumoniae typing and its potentials in hospital hygiene management. Microb. Biotechnol. 2021, 14, 1343–1352. [Google Scholar] [CrossRef]
- Deidda, F.; Cordovana, M.; BozziCionci, N.; Graziano, T.; Di Gioia, D.; Pane, M. In-process real-time probiotic phenotypic strain identity tracking: The use of Fourier transform infrared spectroscopy. Front. Microbiol. 2022, 13, 1052420. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhu, L.; Wang, X.; Li, J.; Tang, B. Evaluation of IR Biotyper for Lactiplantibacillus plantarum Typing and Its Application Potential in Probiotic Preliminary Screening. Front. Microbiol. 2022, 13, 823120. [Google Scholar] [CrossRef] [PubMed]
- Martak, D.; Valot, B.; Sauget, M.; Cholley, P.; Thouverez, M.; Bertrand, X.; Hocquet, D. Fourier-transform infrared spectroscopy can quickly type Gram negative bacilli responsible for hospital outbreaks. Front. Microbiol. 2019, 10, 1440. [Google Scholar] [CrossRef]
- Buron-Moles, G.; Chailyan, A.; Dolejs, I.; Forster, J.; Mikš, M.H. Uncovering carbohydrate metabolism through a genotype-phenotype association study of 56 lactic acid bacteria genomes. Appl. Microbiol. Biotechnol. 2019, 103, 3135–3152. [Google Scholar] [CrossRef]
- Sharma, A.; Lee, S.; Park, Y.S. Molecular typing tools for identifying and characterizing lactic acid bacteria: A review. Food Sci. Biotechnol. 2020, 29, 1301–1318. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manzulli, V.; Cordovana, M.; Serrecchia, L.; Rondinone, V.; Pace, L.; Farina, D.; Cipolletta, D.; Caruso, M.; Fraccalvieri, R.; Difato, L.M.; et al. Application of Fourier Transform Infrared Spectroscopy to Discriminate Two Closely Related Bacterial Species: Bacillus anthracis and Bacillus cereus Sensu Stricto. Microorganisms 2024, 12, 183. https://doi.org/10.3390/microorganisms12010183
Manzulli V, Cordovana M, Serrecchia L, Rondinone V, Pace L, Farina D, Cipolletta D, Caruso M, Fraccalvieri R, Difato LM, et al. Application of Fourier Transform Infrared Spectroscopy to Discriminate Two Closely Related Bacterial Species: Bacillus anthracis and Bacillus cereus Sensu Stricto. Microorganisms. 2024; 12(1):183. https://doi.org/10.3390/microorganisms12010183
Chicago/Turabian StyleManzulli, Viviana, Miriam Cordovana, Luigina Serrecchia, Valeria Rondinone, Lorenzo Pace, Donatella Farina, Dora Cipolletta, Marta Caruso, Rosa Fraccalvieri, Laura Maria Difato, and et al. 2024. "Application of Fourier Transform Infrared Spectroscopy to Discriminate Two Closely Related Bacterial Species: Bacillus anthracis and Bacillus cereus Sensu Stricto" Microorganisms 12, no. 1: 183. https://doi.org/10.3390/microorganisms12010183
APA StyleManzulli, V., Cordovana, M., Serrecchia, L., Rondinone, V., Pace, L., Farina, D., Cipolletta, D., Caruso, M., Fraccalvieri, R., Difato, L. M., Tolve, F., Vetritto, V., & Galante, D. (2024). Application of Fourier Transform Infrared Spectroscopy to Discriminate Two Closely Related Bacterial Species: Bacillus anthracis and Bacillus cereus Sensu Stricto. Microorganisms, 12(1), 183. https://doi.org/10.3390/microorganisms12010183