
Evaluation of Biofilm Production and Antifungal Susceptibility to Fluconazole in Clinical Isolates of Candida spp. in Both Planktonic and Biofilm Form

 , , , , , , ,
, , , , , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolates
2.2. Evaluation of Biofilm
2.2.1. Assessment of Biofilm Production Using Air-Dry Method
2.2.2. Assessment of Biofilm Production Using Crystal Violet Staining
2.2.3. Assessment of Metabolic Activity Using Alamar Blue Reduction Assay
2.3. MBEC50 and PMIC Evaluation
2.3.1. Fluconazole Stock Solution
2.3.2. Planktonic MICs (PMICs)
2.3.3. Minimum Biofilm Eradication Concentrations (MBECs)
2.4. Statistical Analysis
3. Results
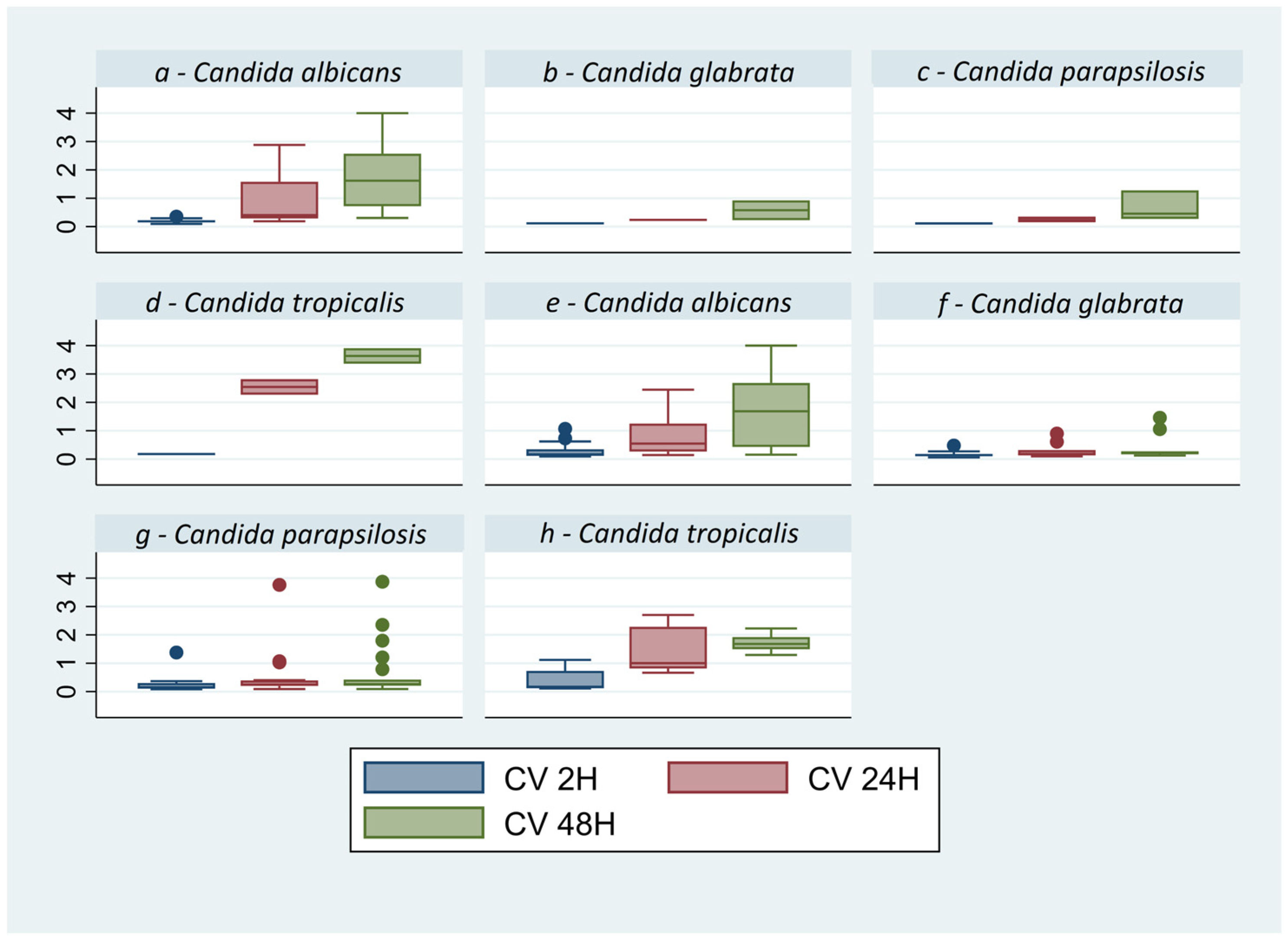
3.1. Biofilm Quantification Assays
3.2. Biofilm Metabolic Activity by Alamar Blue Reduction Assay
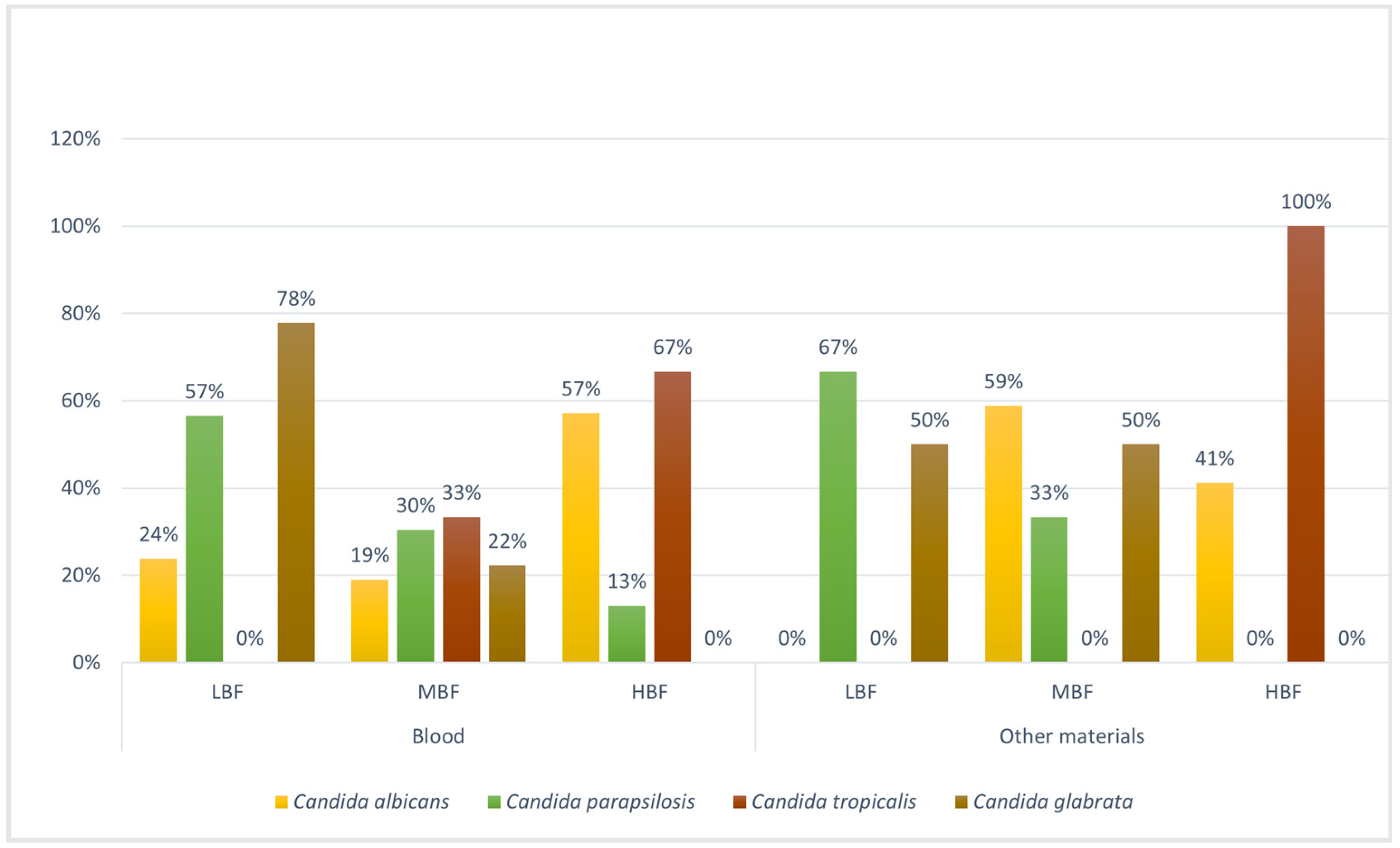
3.3. Planktonic and Biofilm Susceptibility Testing
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Branco, J.; Miranda, I.M.; Rodrigues, A.G. Candida parapsilosis Virulence and Antifungal Resistance Mechanisms: A Comprehensive Review of Key Determinants. J. Fungi 2023, 9, 80. [Google Scholar] [CrossRef] [PubMed]
- Melo, A.S.; Bizerra, F.C.; Freymüller, E.; Arthington-Skaggs, B.A.; Colombo, A.L. Biofilm production and evaluation of antifungal susceptibility amongst clinical Candida spp. isolates, including strains of the Candida parapsilosis complex. Med. Mycol. 2011, 49, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Munhoz-Alves, N.; Mimura, L.A.; Viero, R.M.; Bagagli, E.; Peron, J.P.; Sartori, A.; Fraga-Silva, T.F. Candida tropicalis Systemic Infection Redirects Leukocyte Infiltration to the Kidneys Attenuating Encephalomyelitis. J. Fungi 2021, 7, 757. [Google Scholar] [CrossRef] [PubMed]
- Modiri, M.; Khodavaisy, S.; Barac, A.; Dana, M.A.; Nazemi, L.; Aala, F.; Salehi, M.; Rezaie, S. Comparison of biofilm-producing ability of clinical isolates of Candida parapsilosis species complex. J. Mycol. Médicale 2019, 29, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Pammi, M.; Holland, L.; Butler, G.; Gacser, A.; Bliss, J.M. Candida parapsilosis is a Significant Neonatal Pathogen: A Systematic Review and Meta-Analysis. Pediatr. Infect. Dis. J. 2013, 32, e206. [Google Scholar] [CrossRef] [PubMed]
- Monfredini, P.M.; Souza, A.C.; Cavalheiro, R.P.; Siqueira, R.A.; Colombo, A.L. Clinical impact of Candida spp. biofilm production in a cohort of patients with candidemia. Med. Mycol. 2017, 56, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, K.F.; Zarnowski, R.; Sanchez, H.; Edward, J.A.; Reinicke, E.L.; Nett, J.E.; Mitchell, A.P.; Andes, D.R. Community participation in biofilm matrix assembly and function. Proc. Natl. Acad. Sci. USA 2015, 112, 4092–4097. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Rodrigues, C.F.; Araújo, D.; Rodrigues, M.E.; Henriques, M. Candida Species Biofilms’ Antifungal Resistance. J. Fungi 2017, 3, 8. [Google Scholar] [CrossRef] [PubMed]
- Casagrande Pierantoni, D.; Roscini, L.; Corte, L.; Bernardo, M.; Bassetti, M.; Tascini, C.; Cardinali, G. Qualitative and quantitative change of the tolerance to liposomal amphotericin B triggered by biofilm maturation in C. parapsilosis. Med. Mycol. 2020, 58, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Treatment for Invasive Candidiasis, Centers for Disease Control and Prevention (CDC). Available online: https://www.cdc.gov/fungal/diseases/candidiasis/invasive/treatment.html#:~:text=Treatment%20for%20Invasive%20Candidiasis&text=For%20most%20adults%2C%20the%20initial,be%20appropriate%20in%20certain%20situations (accessed on 14 December 2023).
- Trevijano-Contador, N.; Torres-Cano, A.; Carballo-González, C.; Puig-Asensio, M.; Martín-Gómez, M.T.; Jiménez-Martínez, E.; Romero, D.; Nuvials, F.X.; Olmos-Arenas, R.; Moretó-Castellsagué, M.C.; et al. Global Emer-gence of Resistance to Fluconazole and Voriconazole in Candida parapsilosis in Tertiary Hospitals in Spain during the COVID-19 Pandemic. Open Forum Infect. Dis. 2022, 9, ofac605. [Google Scholar] [CrossRef] [PubMed]
- Gulati, M.; Lohse, M.B.; Ennis, C.L.; Gonzalez, R.E.; Perry, A.M.; Bapat, P.; Arevalo, A.V.; Rodriguez, D.L.; Nobile, C.J. In Vitro Culturing and Screening of Candida albicans Biofilms. Curr. Protoc. Microbiol. 2018, 50, e60. [Google Scholar] [CrossRef] [PubMed]
- Repp, K.K.; Menor, S.A.; Pettit, R.K. Microplate Alamar blue assay for susceptibility testing of Candida albicans biofilms. Med. Mycol. 2007, 45, 603–607. [Google Scholar] [CrossRef] [PubMed]
- EUCAST. E.Def. 2023. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/AFST/Files/EUCAST_E.Def_7.4_Yeast_definitive_revised_2023.pdf (accessed on 18 October 2023).
- EUCAST. Eucast-Clinical Breakpoints for Fungi. 2020. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/AFST/Clinical_breakpoints/AFST_BP_v10.0_200204_updatd_links_200924.pdf (accessed on 10 October 2023).
- Marcos-Zambrano, L.J.; Escribano, P.; Bouza, E.; Guinea, J. Production of biofilm by Candida and non-Candida spp. isolates causing fungemia: Comparison of biomass production and metabolic activity and development of cut-off points. Int. J. Med. Microbiol. 2014, 304, 1192–1198. [Google Scholar] [CrossRef] [PubMed]
- Guinea, J.; Zaragoza, Ó.; Escribano, P.; Martín-Mazuelos, E.; Pemán, J.; Sánchez-Reus, F.; Cuenca-Estrella, M. Molecular identification and antifungal susceptibility of yeast isolates causing fungemia collected in a population-based study in Spain in 2010 and 2011. Antimicrob. Agents Chemother. 2014, 58, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Malinovská, Z.; Čonková, E.; Váczi, P. Biofilm Formation in Medically Important Candida Species. J. Fungi 2023, 9, 955. [Google Scholar] [CrossRef] [PubMed]
- Uppuluri, P.; Srinivasan, A.; Ramasubramanian, A.; Lopez-Ribot, J.L. Effects of Fluconazole, Amphotericin B, and Caspofungin on Candida albicans Biofilms under Conditions of Flow and on Biofilm Dispersion. ASM J. 2011, 55, 3591–3593. [Google Scholar] [CrossRef] [PubMed]
- Atiencia-Carrera, M.B.; Cabezas-Mera, F.S.; Tejera, E.; Machado, A. Prevalence of biofilms in Candida spp. bloodstream infections: A meta-analysis. PLoS ONE 2022, 17, e0263522. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Candida albicans | Candida parapsilosis | Candida glabrata | Candida tropicalis | Total | |
|---|---|---|---|---|---|
| Blood | 21 | 23 | 7 | 6 | 57 |
| Other materials | 17 | 3 | 4 | 2 | 26 |
| Total | 38 | 26 | 11 | 8 | 83 |
| 16 | 8 | 4 | 2 | 1 | 0.5 | 0.25 | 0.13 | 0.06 | 0.03 | CP | CN | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample 1 | ||||||||||||
| Sample 2 | ||||||||||||
| Sample 3 | ||||||||||||
| Sample 4 | ||||||||||||
| Sample 5 | ||||||||||||
| Sample 6 | ||||||||||||
| Sample 7 | ||||||||||||
| Sample 8 |
| Candida albicans OD ± SD | Candida parapsilosis OD ± SD | Candida glabrata OD ± SD | Candida tropicalis OD ± SD | |
|---|---|---|---|---|
| AIR-DRY 2H | 0.015 ± 0.025 | 0.010 ± 0.018 | 0.007 ± 0.008 | 0.006 ± 0.015 |
| AIR-DRY 24H | 0.069 ± 0.084 | 0.030 ± 0.064 | 0.010 ± 0.008 | 0.074 ± 0.025 |
| AIR-DRY 48H | 0.137 ± 0.121 | 0.051 ± 0.080 | 0.025 ± 0.038 | 0.134 ± 0.065 |
| CV 2H | 0.240 ± 0.202 | 0.219 ± 0.253 | 0.163 ± 0.119 | 0.345 ± 0.373 |
| CV 24H | 0.846 ± 0.785 | 0.446 ± 0.715 | 0.290 ± 0.247 | 1.694 ± 0.910 |
| CV 48H | 1.755 ± 1.210 | 0.636 ± 0.861 | 0.456 ± 0.462 | 2.196 ± 0.942 |
| AB 2H | 0.142 ± 0.213 | 0.154 ± 0.205 | 0.034 ± 0.040 | 0.103 ± 0.143 |
| AB 24H | 0.133 ± 0.127 | 0.052 ± 0.084 | 0.182 ± 0.257 | 0.114 ± 0.053 |
| AB 48H | 0.195 ± 0.158 | 0.129 ± 0.216 | 0.184 ± 0.260 | 0.150 ± 0.044 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marzucco, A.; Gatti, G.; Montanari, M.S.; Fantini, M.; Colosimo, C.; Tamburini, M.V.; Arfilli, V.; Morotti, M.; Schiavone, P.; Congestrì, F.; et al. Evaluation of Biofilm Production and Antifungal Susceptibility to Fluconazole in Clinical Isolates of Candida spp. in Both Planktonic and Biofilm Form. Microorganisms 2024, 12, 153. https://doi.org/10.3390/microorganisms12010153
Marzucco A, Gatti G, Montanari MS, Fantini M, Colosimo C, Tamburini MV, Arfilli V, Morotti M, Schiavone P, Congestrì F, et al. Evaluation of Biofilm Production and Antifungal Susceptibility to Fluconazole in Clinical Isolates of Candida spp. in Both Planktonic and Biofilm Form. Microorganisms. 2024; 12(1):153. https://doi.org/10.3390/microorganisms12010153
Chicago/Turabian StyleMarzucco, Anna, Giulia Gatti, Maria Sofia Montanari, Michela Fantini, Claudia Colosimo, Maria Vittoria Tamburini, Valentina Arfilli, Manuela Morotti, Pasqualina Schiavone, Francesco Congestrì, and et al. 2024. "Evaluation of Biofilm Production and Antifungal Susceptibility to Fluconazole in Clinical Isolates of Candida spp. in Both Planktonic and Biofilm Form" Microorganisms 12, no. 1: 153. https://doi.org/10.3390/microorganisms12010153
APA StyleMarzucco, A., Gatti, G., Montanari, M. S., Fantini, M., Colosimo, C., Tamburini, M. V., Arfilli, V., Morotti, M., Schiavone, P., Congestrì, F., Manera, M., Denicolò, A., Brandolini, M., Taddei, F., Grumiro, L., Zannoli, S., Dirani, G., De Pascali, A. M., Sambri, V., & Cricca, M. (2024). Evaluation of Biofilm Production and Antifungal Susceptibility to Fluconazole in Clinical Isolates of Candida spp. in Both Planktonic and Biofilm Form. Microorganisms, 12(1), 153. https://doi.org/10.3390/microorganisms12010153