

Dual-Temperature Microbiological Control of Cellular Products: A Potential Impact for Bacterial Screening of Platelet Concentrates?
Abstract
:1. Introduction
2. Material and Methods
2.1. PC Collection
2.2. Bacterial Strains
2.3. Spiking Experiments, Microbiological Cultivation, and Bacterial Identification
2.4. Statistical Analysis
3. Results
3.1. Growth Characteristic of Gram-Negatives
3.2. Growth Characteristic of Bacillus Species
3.3. Growth Characteristic of Staphylococcus Species
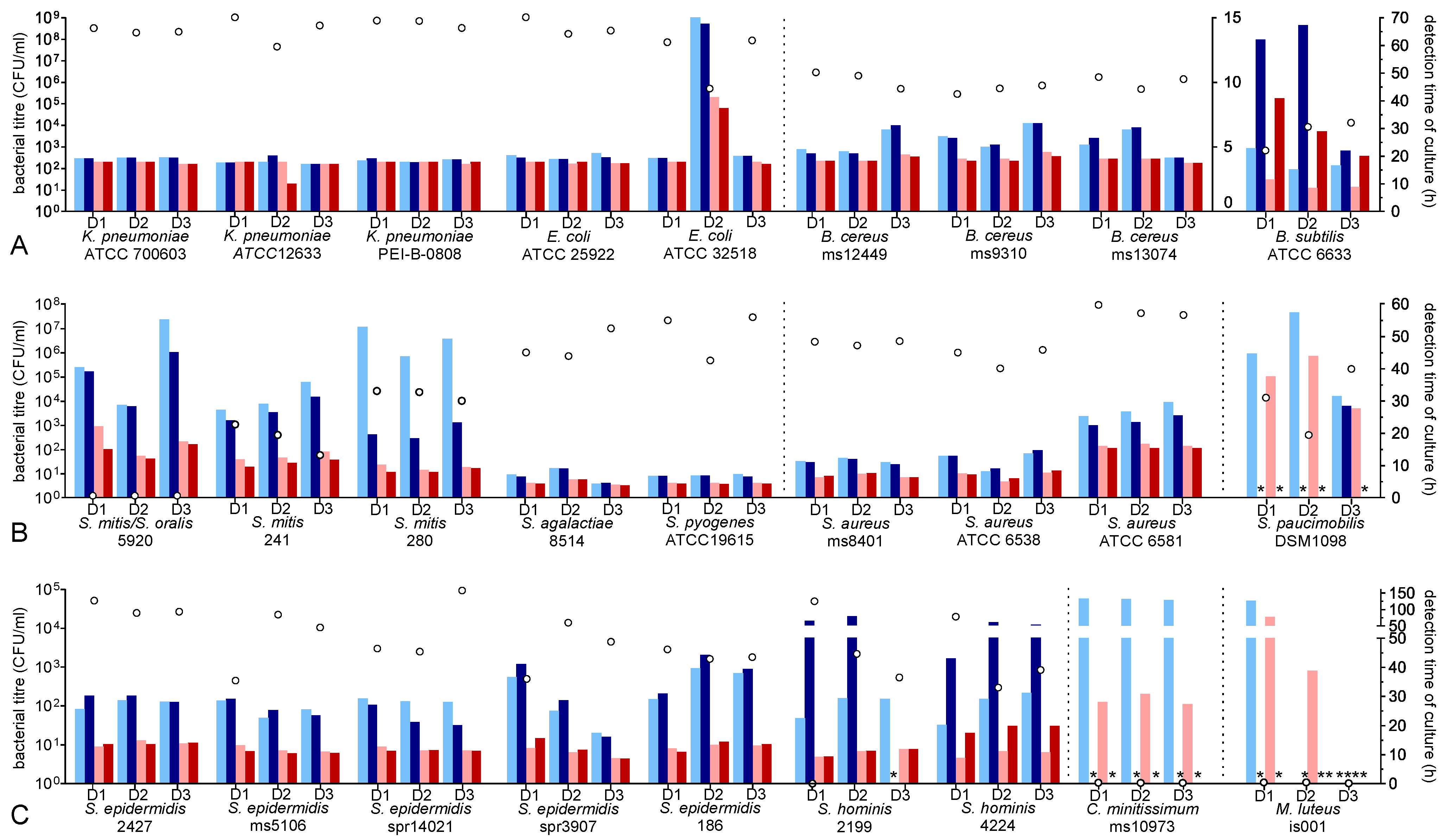
3.4. Growth Characteristic of Streptococcus Species
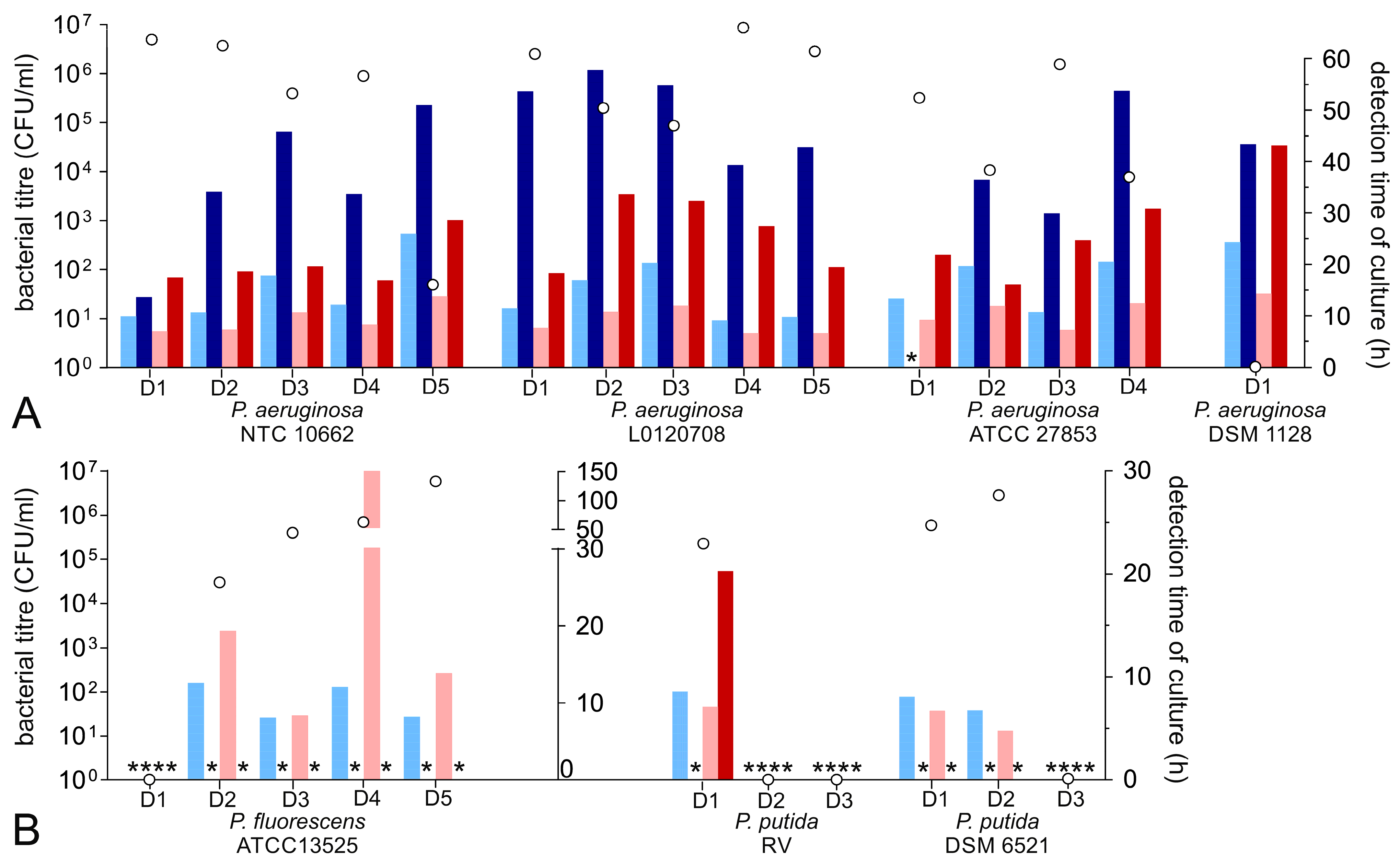
3.5. Growth Characteristic of Pseudomonas Species
3.6. Growth Characteristic of Other Species
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Pharmacopeia Chapter 2.6.27: Microbiological Examination of Cell-Based Preparations. 2021. Available online: https://pheur.edqm.eu (accessed on 13 September 2023).
- Störmer, M.; Wood, E.M.; Schurig, U.; Karo, O.; Spreitzer, I.; McDonald, C.P.; Montag, T. Bacterial safety of cell-based therapeutic preparations, focusing on haematopoietic progenitor cells. Vox Sang. 2014, 106, 285–296. [Google Scholar] [CrossRef]
- Arbeitskreis Blut. Votum 43: Aktualisierung des Votums 16—Mindestanforderungen an die mikrobiologische Kontrolle von Blutkomponenten zur Transfusion. Bundesgesundheitsblatt 2013, 56, 474–475. [Google Scholar] [CrossRef]
- Prax, M.; Bekeredjian-Ding, I.; Krut, O. Microbiological Screening of Platelet Concentrates in Europe. Transfus. Med. Hemother. 2019, 46, 76–86. [Google Scholar] [CrossRef]
- Funk, M.B.; Lohmann, A.; Guenay, S.; Henseler, O.; Heiden, M.; Hanschmann, K.M.; Keller-Stanislawski, B. Transfusion-Transmitted Bacterial Infections—Haemovigilance Data of German Blood Establishments (1997–2010). Transfus. Med. Hemother. 2011, 38, 266–271. [Google Scholar] [CrossRef]
- McDonald, C.; Allen, J.; Brailsford, S.; Roy, A.; Ball, J.; Moule, R.; Vasconcelos, M.; Morrison, R.; Pitt, T. Bacterial screening of platelet components by National Health Service Blood and Transplant, an effective risk reduction measure. Transfusion 2017, 57, 1122–1131. [Google Scholar] [CrossRef]
- Dickson, M.; Dinesh, D. Bacterial contamination of platelet concentrates produced in New Zealand. N. Z. Med. J. 2013, 126, 12–21. [Google Scholar]
- Dumont, L.J.; Kleinman, S.; Murphy, J.R.; Lippincott, R.; Schuyler, R.; Houghton, J.; Metzel, P. Screening of single-donor apheresis platelets for bacterial contamination: The PASSPORT study results. Transfusion 2010, 50, 589–599. [Google Scholar] [CrossRef]
- Murphy, W.G.; Foley, M.; Doherty, C.; Tierney, G.; Kinsella, A.; Salami, A.; Cadden, E.; Coakley, P. Screening platelet concentrates for bacterial contamination: Low numbers of bacteria and slow growth in contaminated units mandate an alternative approach to product safety. Vox Sang. 2008, 95, 13–19. [Google Scholar] [CrossRef]
- Vuk, T.; Barisic, M.; Hecimovic, A.; Rukavina, L.; Batarilo, I.; Sarlija, D.; Jukic, I. Bacterial contamination of blood products at the Croatian Institute of Transfusion Medicine: Results of eleven-year monitoring. Transfus Med 2012, 22, 432–439. [Google Scholar] [CrossRef]
- Walther-Wenke, G.; Daubener, W.; Heiden, M.; Hoch, J.; Hornei, B.; Volkers, P.; von Konig, C.H. Effect of Safety Measures on Bacterial Contamination Rates of Blood Components in Germany. Transfus. Med. Hemother. 2011, 38, 231–235. [Google Scholar] [CrossRef]
- Kleinman, S.H.; Kamel, H.T.; Harpool, D.R.; Vanderpool, S.K.; Custer, B.; Wiltbank, T.B.; Nguyen, K.A.; Tomasulo, P.A. Two-year experience with aerobic culturing of apheresis and whole blood-derived platelets. Transfusion 2006, 46, 1787–1794. [Google Scholar] [CrossRef]
- Orru, S.; Oberle, D.; Heiden, M.; Muller, S.; Krut, O.; Funk, M.B. Analysis of Transfusion-Transmitted Bacterial Infections according to German Hemovigilance Data (2011–2020). Transfus. Med. Hemother. 2023, 50, 144–153. [Google Scholar] [CrossRef]
- Lafeuillade, B.; Eb, F.; Ounnoughene, N.; Petermann, R.; Daurat, G.; Huyghe, G.; Vo Mai, M.P.; Caldani, C.; Rebibo, D.; Weinbreck, P. Residual risk and retrospective analysis of transfusion-transmitted bacterial infection reported by the French National Hemovigilance Network from 2000 to 2008. Transfusion 2015, 55, 636–646. [Google Scholar] [CrossRef]
- Satake, M.; Kozakai, M.; Matsumoto, M.; Matsubayashi, K.; Taira, R.; Goto, N. Platelet safety strategies in Japan: Impact of short shelf life on the incidence of septic reactions. Transfusion 2020, 60, 731–738. [Google Scholar] [CrossRef]
- Störmer, M.; Arroyo, A.; Brachert, J.; Carrero, H.; Devine, D.; Epstein, J.S.; Gabriel, C.; Gelber, C.; Goodrich, R.; Hanschmann, K.M.; et al. Establishment of the first International Repository for Transfusion-Relevant Bacteria Reference Strains: ISBT Working Party Transfusion-Transmitted Infectious Diseases (WP-TTID), Subgroup on Bacteria. Vox Sang. 2012, 102, 22–31. [Google Scholar] [CrossRef]
- Montag, T.; Stoermer, M.; Schurig, U.; Brachert, J.; Bubenzer, M.; Sicker, U.; Beshir, R.; Spreitzer, I.; Loeschner, B.; Bache, C.; et al. Probleme der mikrobiellen Sicherheit nei neuartigen Therapien. Bundesgesundheitsblatt 2010, 53, 45–51. [Google Scholar] [CrossRef]
- Montag, T.; Schurig, U. Einsatz von BacT/Alert versus BACTEC für die Sterilitätstestung. In Proceedings of the KOLT Meeting, Langen, Germany, 12 November 2009; Paul-Ehrlich Institute: Frankfurt, Germany, 2009. [Google Scholar]
- Brecher, M.E.; Hay, S.N. Bacterial contamination of blood components. Clin. Microbiol. Rev. 2005, 18, 195–204. [Google Scholar] [CrossRef]
- Palavecino, E.L.; Yomtovian, R.A.; Jacobs, M.R. Detecting bacterial contamination in platelet products. Clin. Lab. 2006, 52, 443–456. [Google Scholar]
- Günther, S.K.; Geiss, C.; Kaiser, S.J.; Mutters, N.T.; Günther, F. Microbiological Control of Cellular Products: The Relevance of the Cellular Matrix, Incubation Temperature, and Atmosphere for the Detection Performance of Automated Culture Systems. Transfus. Med. Hemother. 2020, 47, 254–263. [Google Scholar] [CrossRef]
- Walker, B.S.; Schmidt, R.L.; Moore, R.A.; White, S.K.; Fisher, M.A.; Metcalf, R.A. Bacterial culture time to detection in platelet components: An evidence synthesis and estimation of detection failures. Transfusion 2023, 63, 182–192. [Google Scholar] [CrossRef]
- Mastronardi, C.; Martincic, I.; Ramirez-Arcos, S. Implementation of a proficiency testing programme for bacterial screening in platelet preparations in Canada. Vox Sang. 2007, 93, 131–138. [Google Scholar] [CrossRef]
- Müller, T.H.; Montag, T.; Seltsam, A.W. Laboratory Evaluation of the Effectiveness of Pathogen Reduction Procedures for Bacteria. Transfus. Med. Hemother. 2011, 38, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, E.; Seghatchian, J. Bacterial contamination in blood components and preventative strategies: An overview. Transfus. Apher. Sci. Off. J. World Apher. Assoc. Off. J. Eur. Soc. Haemapheresis 2004, 31, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Kamel, H.; Ramirez-Arcos, S.; McDonald, C.; ISBT Transfusion-Transmitted Infectious Disease Bacterial Working Party Bacterial Subgroup. The international experience of bacterial screen testing of platelet components with automated microbial detection systems: An update. Vox Sang. 2022, 117, 647–655. [Google Scholar] [CrossRef]
- Schmidt, M.; Ramirez-Arcos, S.; Stiller, L.; McDonald, C.; ISBT Transfusion-Transmitted Infectious Diseases Working Party, Subgroup on Bacteria. Current status of rapid bacterial detection methods for platelet components: A 20-year review by the ISBT Transfusion-Transmitted Infectious Diseases Working Party Subgroup on Bacteria. Vox Sang. 2022, 117, 983–988. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Strain | Origin | Inoculum (CFU/PC) | Proliferation (n) | Detection Time | Figure | |||
|---|---|---|---|---|---|---|---|---|
| 25 °C (h (±SD)) | 35 °C (h (±SD)) | |||||||
| AE | AN | AE | AN | |||||
| Enterobacteriacae | ||||||||
| K. pneumoniae ATCC 700603 | RS | 11–14 | yes (3/3) | 3.94 (±0.18) | 4.01 (±0.22) | 3.77 (±0.08) | 3.60 (±0.55) | Figure 1A |
| K. pneumoniae ATCC 12633 | RS | |||||||
| K. pneumoniae PEI-B-0808 ** | RS | |||||||
| E. coli ATCC 25922 | RS | 6–12 | yes (3/3) | 4.24 (±0.19) | 4.14 (±0.10) | 3.77 (±0.07) | 3.76 (±0.08) | Figure 1A |
| E. coli ATCC 35218 | RS | |||||||
| Bacillus spp. | ||||||||
| B. cereus ms12449 | PC | 10–30 | yes (3/3) | 5.46 (±0.90) | 5.51 (±1.01) | 4.09 (±0.26) | 4.00 (±0.17) | Figure 1A |
| B. cereus ms9310 | PC | |||||||
| B. cereus ms13074 | PC | |||||||
| B. subtilis ATCC 6633 * | RS | 10 | yes (3/3) | 18.22 (±4.06) | 50.50 (±24.82) | 9.65 (±1.69) | 29.99 (±10.46) | Figure 1A |
| Staphylococcus spp. | ||||||||
| S. epidermidis 2427 | PC | 10–20 | yes (3/3) | 28.69 (±5.76) | 29.00 (±7.81) | 12.19 (±1.49) | 12.14 (±1.89) | Figure 1C |
| S. epidermidis ms5106 | PC | |||||||
| S. epidermidis spr14021 | PC | |||||||
| S. epidermidis ms6031 | PC | |||||||
| S. epidermidis spr3907 | PC | |||||||
| S. epidermidis 186 | PC | |||||||
| S. hominis 2199 | PC | 13–18 | yes (3/3) | 26.79 (±4.37) | 49.10 (±26.46) | 10.41 (±1.22) | 14.85 (±4.65) | Figure 1C |
| S. hominis 4224 | PC | |||||||
| S. aureus ms8401 | PC | 10 | yes (3/3) | 9.93 (±2.84) | 9.70 (±3.25) | 5.78 (±1.53) | 5.93 (±1.73) | Figure 1B |
| S. aureus ATCC 6538 | RS | |||||||
| S. aureus ATCC 6581 | RS | |||||||
| Streptococcus spp. | ||||||||
| S. mitis/S. oralis spr 5920 | PC | 22–56 | yes (3/3) | 41.35 (±13.29) | 37.35 (±8.54 | 17.38 (±4.58) | 14.49 (±2.30) | Figure 1A |
| S. mitis 241 | PC | 30.56 (±4.47) | 27.06 (±3.73) | 12.79 (±1.23) | 10.63 (±1.08) | |||
| S. mitis 280 | PC | 48.51 (±4.61) | 20.30 (±2.51) | 9.33 (±0.84) | 8.21 (±0.68) | |||
| S. pyogenes ATCC 19615 * | RS | 10 | yes (3/3) | 6.89 (±0.26) | 6.67 (±0.17) | 4.45 (±4.45) | 4.23 (±4.23) | Figure 1A |
| S. agalactiae 8514 | PC | 16 | yes (3/3) | 6.85 (±2.43) | 6.64 (±2.26) | 4.69 (±0.85) | 4.58 (±0.95) | Figure 1A |
| Strain | Origin | Inoculum (CFU/PC) | Proliferation (n) | Detection Time | Figure | |||
|---|---|---|---|---|---|---|---|---|
| 25 °C (h (±SD)) | 35 °C (h (±SD)) | |||||||
| AE | AN | AE | AN | |||||
| Pseudomonas spp. | ||||||||
| P. aeruginosa ATCC 9027 | RS | 12 | no (5/5) | negative (5/5) | none | |||
| P. aeruginosa DSM 1128 | RS | 24 | no (5/5) | 24.23 | 43.25 | 14.25 | 43.05 | Figure 2A |
| P. aeruginosa ATCC 27853 | RS | 24 | yes (4/5) | 16.11 (±4.81) | 30.02 (±22.40) | 10.21 (±2.42) | 23.40 (±6.13) | Figure 2A |
| P. aeruginosa NTC 10662 | RS | 10 | yes (5/5) | 15.38 (±6.71) | 35.72 (±14.39) | 9.48 (±2.85) | 20.32 (±4.80) | Figure 2A |
| P. aeruginosa L0120708 | RS | 21 | yes (5/5) | 13.60 (±4.88) | 49.75 (±8.11) | 8.81 (±2.49) | 26.31 (±7.12) | Figure 2A |
| P. fluorescens DSM 6147 | RS | 54 | no (3/3) | negative (3/3) | none | |||
| P. fluorescens DSM 50091 | RS | 12 | no (3/3) | negative (3/3) | none | |||
| P. fluorescens DSM 50415 | RS | 67 | no (3/3) | negative (3/3) | none | |||
| P. fluorescens ATCC 13525 | RS | 30–35 | yes (4/5) | 10.92 (±3.62) | negative | 42.74 (±71.02) | negative | Figure 2B |
| P. putida DSM 6521 | RS | 24 | yes (2/5) | 7.81 (±0.93) | negative | 6.20 (±1.25) | (20.27, 1 bottle) | Figure 2B |
| P. putida RV | P | 24 | yes (1/3) | Figure 2B | ||||
| P. putida DSM 50257 | RS | 25 | no (3/3) | negative (3/3) | none | |||
| P. putida ATCC 12633 | RS | 30–42 | no (3/3) | negative (3/3) | none | |||
| Others | ||||||||
| C. acnes spr4694 | PC | 10–48 | no (3/3) | statistical analysis not reliable due to heterogenous results | none | |||
| C. acnes spr14472 | PC | |||||||
| C. acnes spr4991 | PC | |||||||
| C. acnes ATCC 11827 | RS | |||||||
| S. paucimobilis DSM 1098 | RS | 10 | yes (3/3) | 44.41 (±13.0) | (28.35, 1 bottle) | 36.25 (±8.15) | negative | Figure 1A |
| C. minitissimum ms10973 | PC | 30 | no (3/3) | 122.8 (±2.08) | negative | 28.64 (±1.86) | negative | Figure 1C |
| M. lutens is001 | P | 16 | no (3/3) | statistical analysis not reliable due to heterogenous results | Figure 1C | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vollmer, T.; Knabbe, C.; Dreier, J. Dual-Temperature Microbiological Control of Cellular Products: A Potential Impact for Bacterial Screening of Platelet Concentrates? Microorganisms 2023, 11, 2350. https://doi.org/10.3390/microorganisms11092350
Vollmer T, Knabbe C, Dreier J. Dual-Temperature Microbiological Control of Cellular Products: A Potential Impact for Bacterial Screening of Platelet Concentrates? Microorganisms. 2023; 11(9):2350. https://doi.org/10.3390/microorganisms11092350
Chicago/Turabian StyleVollmer, Tanja, Cornelius Knabbe, and Jens Dreier. 2023. "Dual-Temperature Microbiological Control of Cellular Products: A Potential Impact for Bacterial Screening of Platelet Concentrates?" Microorganisms 11, no. 9: 2350. https://doi.org/10.3390/microorganisms11092350
APA StyleVollmer, T., Knabbe, C., & Dreier, J. (2023). Dual-Temperature Microbiological Control of Cellular Products: A Potential Impact for Bacterial Screening of Platelet Concentrates? Microorganisms, 11(9), 2350. https://doi.org/10.3390/microorganisms11092350