
Host-Derived Extracellular Vesicles in Blood and Tissue Human Protozoan Infections


Abstract
:1. Introduction
1.1. Extracellular Vesicles: Important Definitions
1.2. Generalities of EVs in Parasitic Diseases
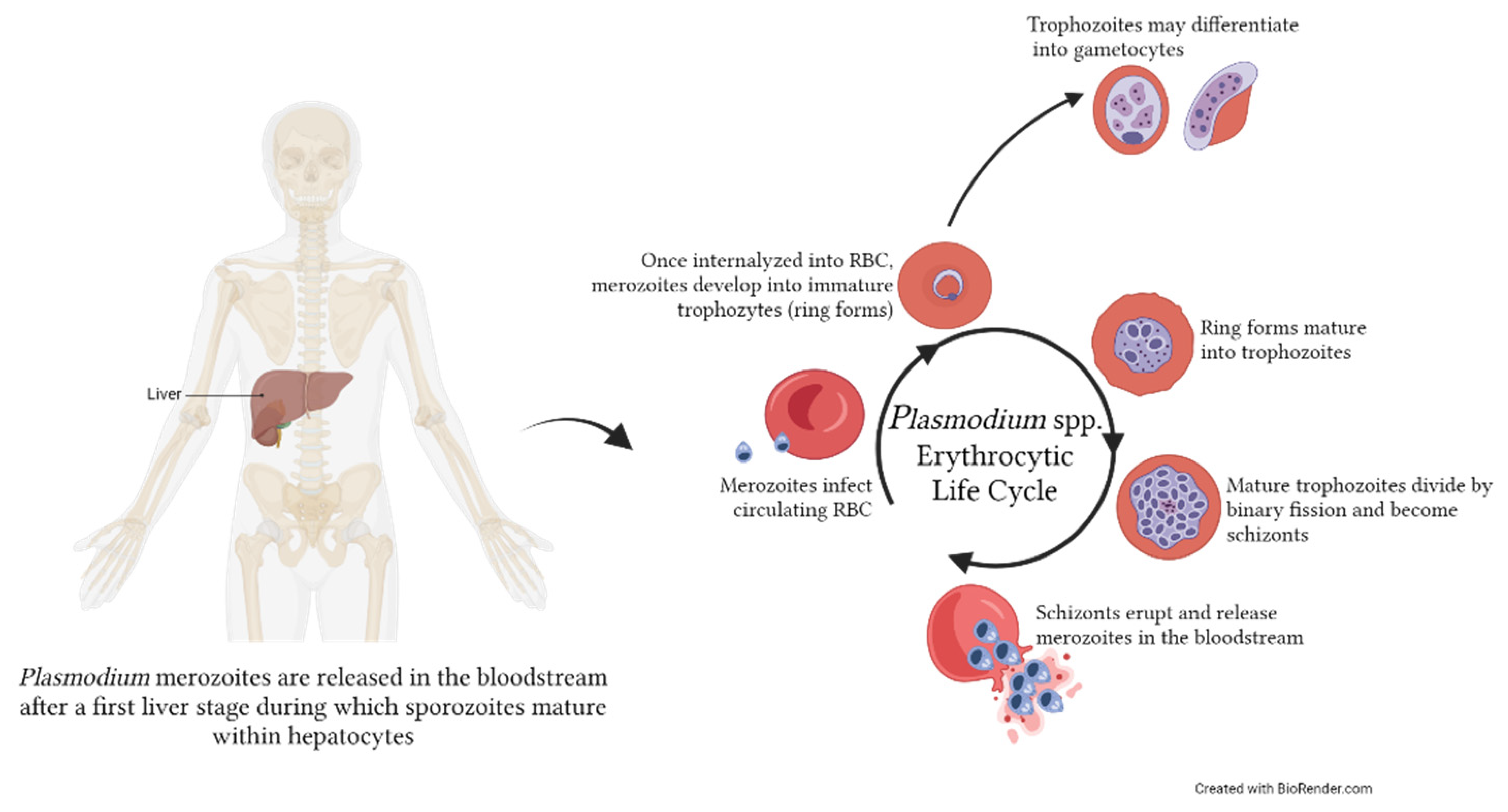
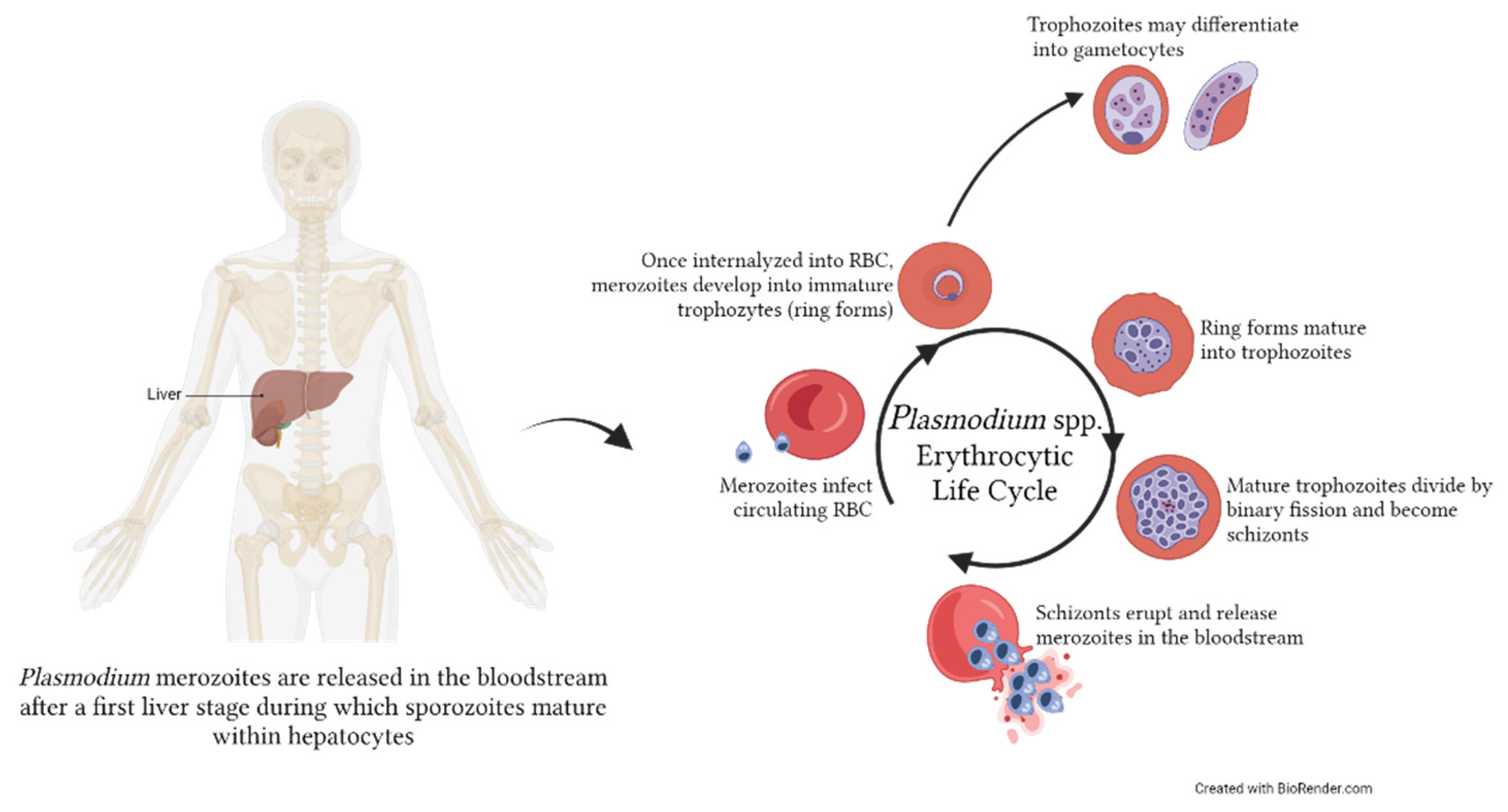
2. Malaria
2.1. Surface Markers of Circulating EVs during Malaria
2.1.1. Clinical Malaria
2.1.2. Experimental Malaria
2.2. Circulating EV Cargo: Phenotypic Characterisation and Functional Properties
2.2.1. MVs Protein Cargo
2.2.2. EXOs Protein Cargo
2.2.3. EXOs miRNA Cargo
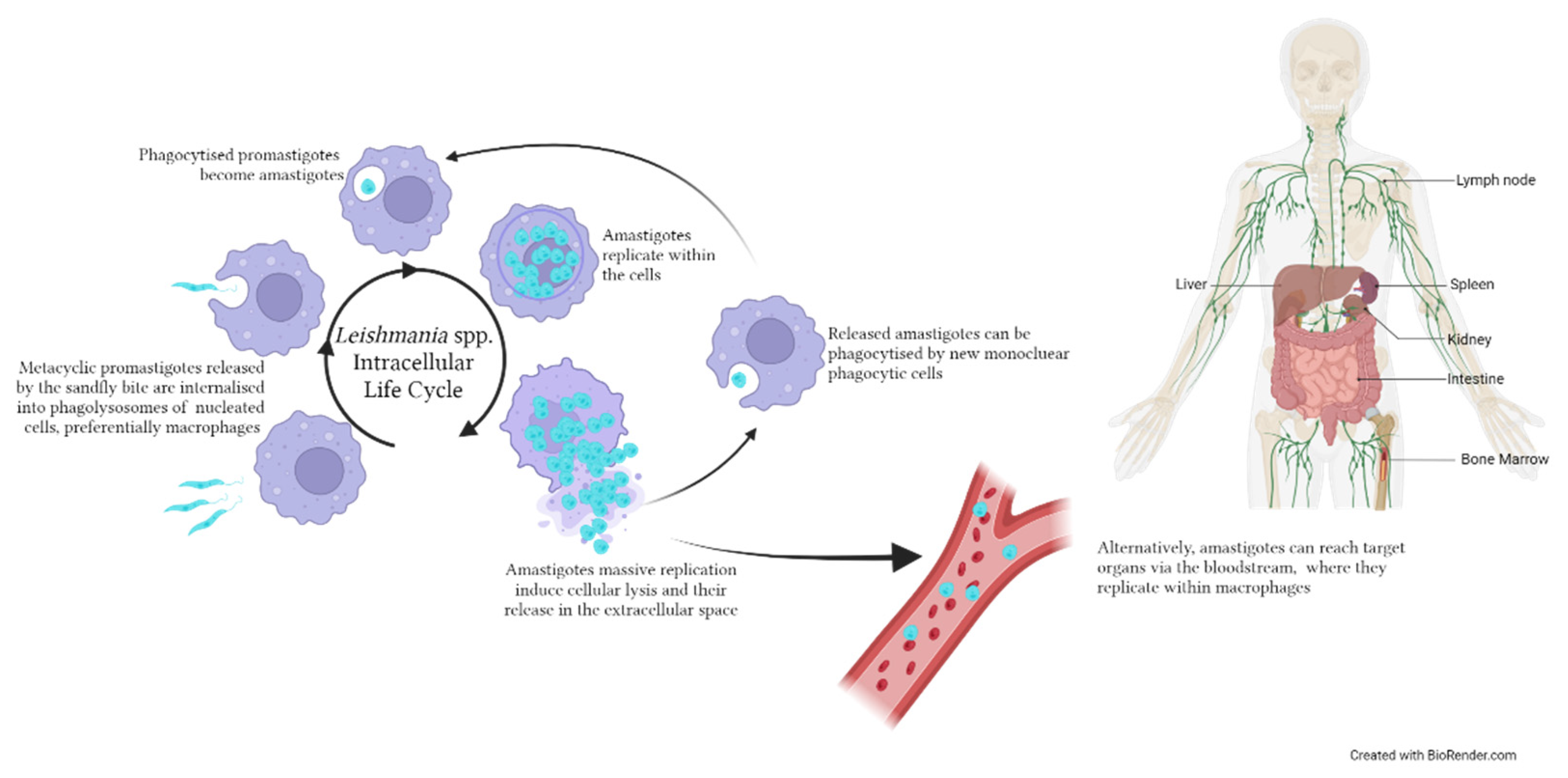
3. Leishmaniasis
Host-Derived Circulating EVs: The Case of Dogs and Mice
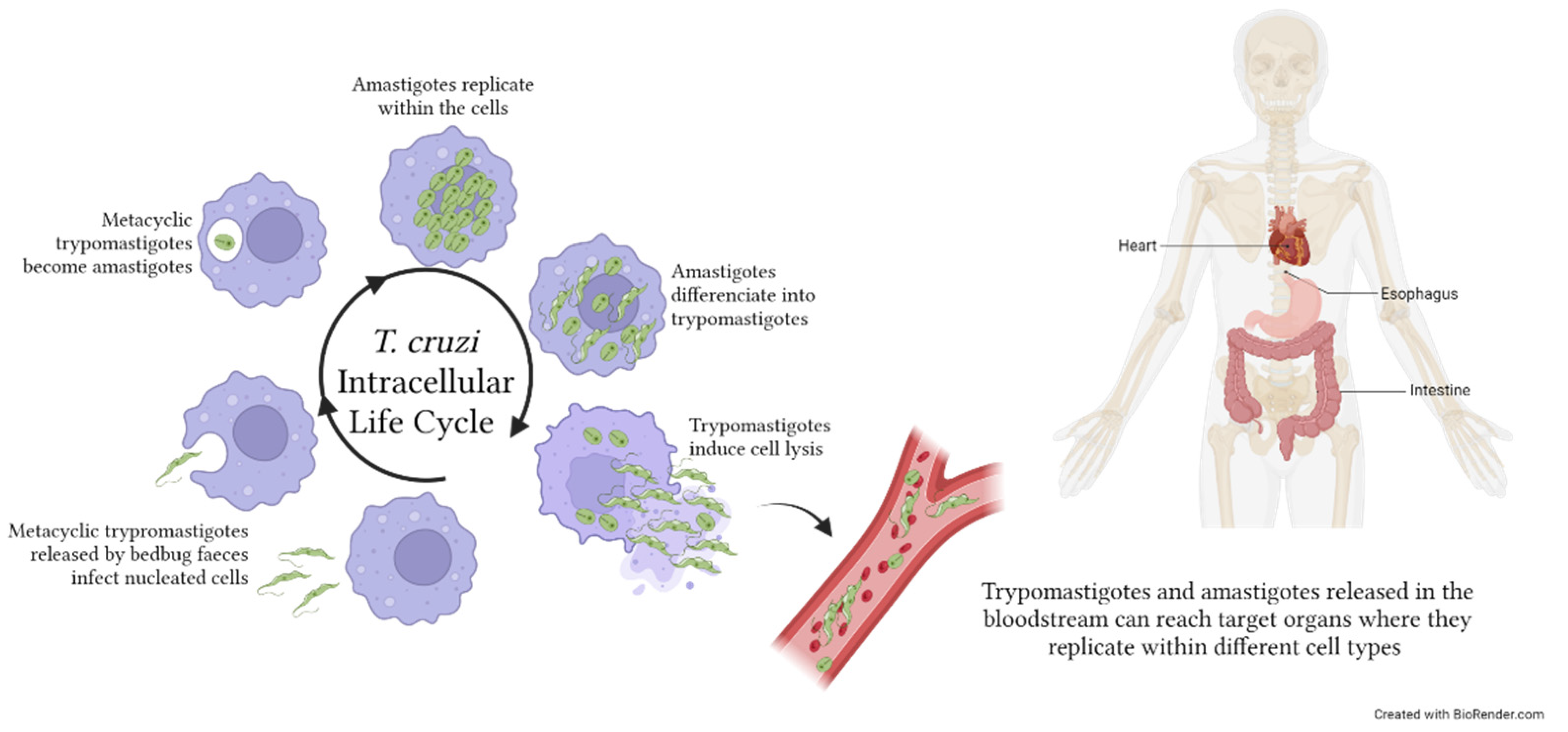
4. Chagas Disease
Circulating EVs in Chagas Disease: Cargo and Functional Properties
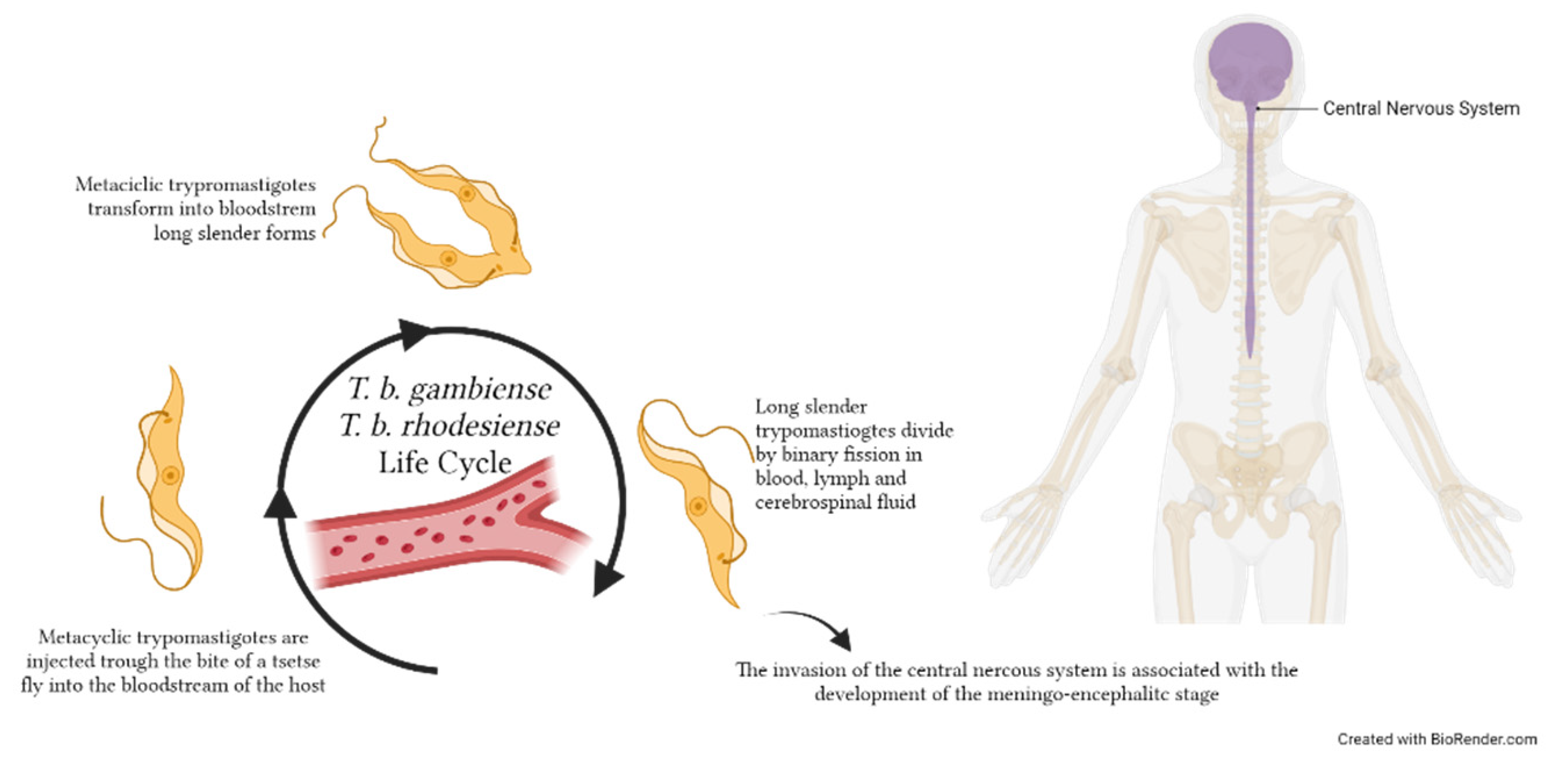
5. Circulating EVs in Human African Trypanosomiasis (HAT): Evidence in CSF
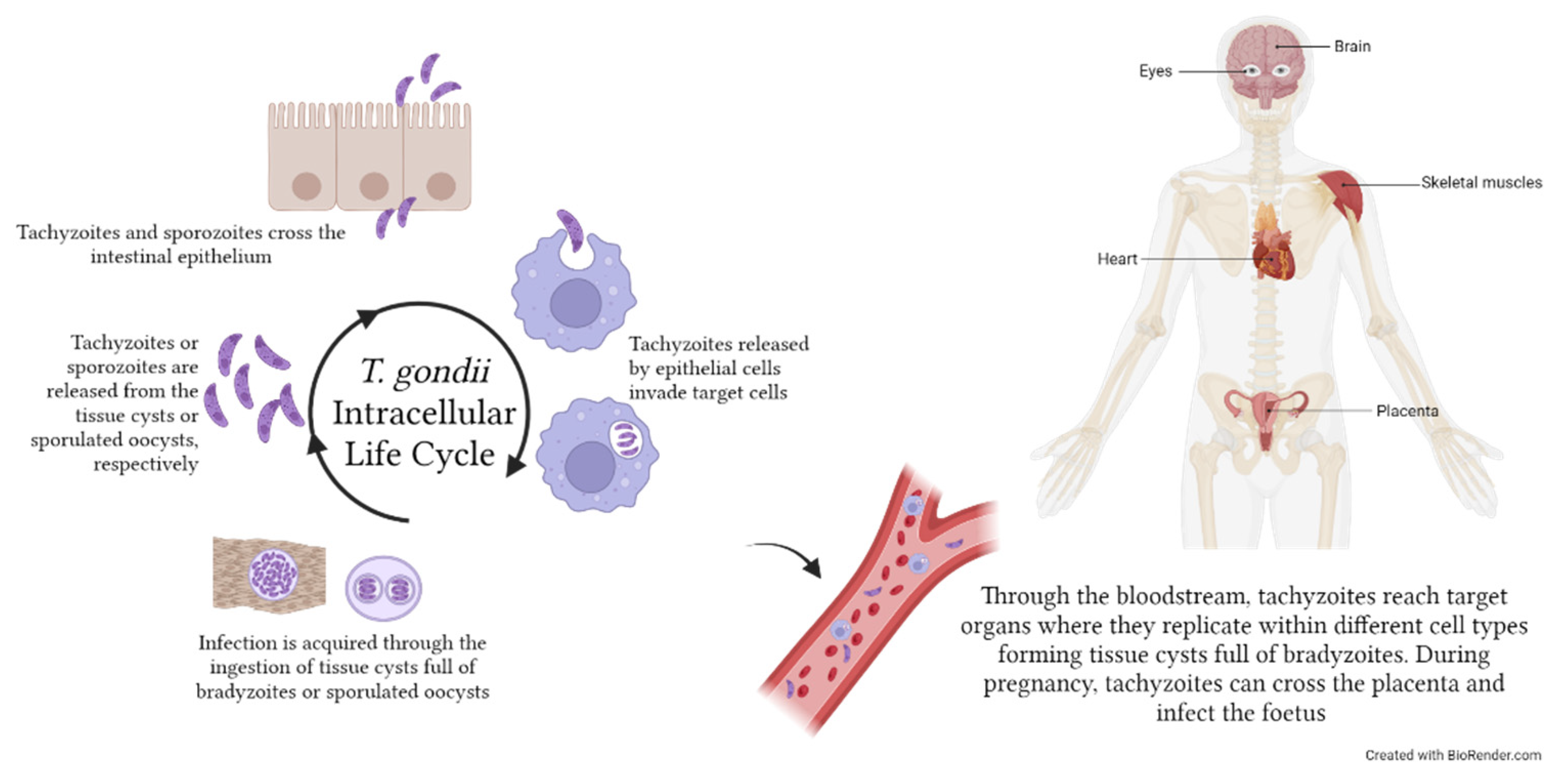
6. Toxoplasmosis
Circulating EVs in Toxoplasmosis: Evidence in Serum and CSF
7. Concluding Remarks and Future Challenges
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marti, M.; Johnson, P.J. Emerging roles for extracellular vesicles in parasitic infections. Curr. Opin. Microbiol. 2016, 32, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Schorey, J.S.; Harding, C.V. Extracellular vesicles and infectious diseases: New complexity to an old story. J. Clin. Investig. 2016, 126, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Coakley, G.; Maizels, R.M.; Buck, A.H. Exosomes and Other Extracellular Vesicles: The New Communicators in Parasite Infections. Trends Parasitol. 2015, 31, 477–489. [Google Scholar] [CrossRef]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular Vesicles: Unique Intercellular Delivery Vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Bazzan, E.; Tine, M.; Casara, A.; Biondini, D.; Semenzato, U.; Cocconcelli, E.; Balestro, E.; Damin, M.; Radu, C.M.; Turato, G.; et al. Critical Review of the Evolution of Extracellular Vesicles’ Knowledge: From 1946 to Today. Int. J. Mol. Sci. 2021, 22, 6417. [Google Scholar] [CrossRef] [PubMed]
- Lotvall, J.; Hill, A.F.; Hochberg, F.; Buzas, E.I.; Di Vizio, D.; Gardiner, C.; Gho, Y.S.; Kurochkin, I.V.; Mathivanan, S.; Quesenberry, P.; et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: A position statement from the International Society for Extracellular Vesicles. J. Extracell. Vesicles 2014, 3, 26913. [Google Scholar] [CrossRef]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Buzas, E.I. The roles of extracellular vesicles in the immune system. Nat. Rev. Immunol. 2023, 23, 236–250. [Google Scholar] [CrossRef]
- Thery, C.; Duban, L.; Segura, E.; Veron, P.; Lantz, O.; Amigorena, S. Indirect activation of naive CD4+ T cells by dendritic cell-derived exosomes. Nat. Immunol. 2002, 3, 1156–1162. [Google Scholar] [CrossRef]
- Admyre, C.; Johansson, S.M.; Paulie, S.; Gabrielsson, S. Direct exosome stimulation of peripheral human T cells detected by ELISPOT. Eur. J. Immunol. 2006, 36, 1772–1781. [Google Scholar] [CrossRef]
- Walker, J.D.; Maier, C.L.; Pober, J.S. Cytomegalovirus-infected human endothelial cells can stimulate allogeneic CD4+ memory T cells by releasing antigenic exosomes. J. Immunol. 2009, 182, 1548–1559. [Google Scholar] [CrossRef]
- Qazi, K.R.; Gehrmann, U.; Domange Jordo, E.; Karlsson, M.C.; Gabrielsson, S. Antigen-loaded exosomes alone induce Th1-type memory through a B-cell-dependent mechanism. Blood 2009, 113, 2673–2683. [Google Scholar] [CrossRef] [PubMed]
- Atayde, V.D.; Hassani, K.; da Silva Lira Filho, A.; Borges, A.R.; Adhikari, A.; Martel, C.; Olivier, M. Leishmania exosomes and other virulence factors: Impact on innate immune response and macrophage functions. Cell. Immunol. 2016, 309, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.; Malik, M.I.; Hameed, M.; Zhou, J. Research progress on the composition and function of parasite-derived exosomes. Acta Trop. 2019, 196, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Mantel, P.Y.; Marti, M. The role of extracellular vesicles in Plasmodium and other protozoan parasites. Cell. Microbiol. 2014, 16, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Garcez, E.M.; Gomes, N.; Moraes, A.S.; Pogue, R.; Uenishi, R.H.; Hecht, M.; Carvalho, J.L. Extracellular vesicles in the context of chagas disease—A systematic review. Acta Trop. 2023, 242, 106899. [Google Scholar] [CrossRef]
- Opadokun, T.; Rohrbach, P. Extracellular vesicles in malaria: An agglomeration of two decades of research. Malar. J. 2021, 20, 442. [Google Scholar] [CrossRef]
- Wang, X.; Chen, J.; Zheng, J. The state of the art of extracellular vesicle research in protozoan infection. Front. Genet. 2022, 13, 941561. [Google Scholar] [CrossRef]
- Torrecilhas, A.C.; Soares, R.P.; Schenkman, S.; Fernandez-Prada, C.; Olivier, M. Extracellular Vesicles in Trypanosomatids: Host Cell Communication. Front. Cell. Infect. Microbiol. 2020, 10, 602502. [Google Scholar] [CrossRef]
- Dong, G.; Wagner, V.; Minguez-Menendez, A.; Fernandez-Prada, C.; Olivier, M. Extracellular vesicles and leishmaniasis: Current knowledge and promising avenues for future development. Mol. Immunol. 2021, 135, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Carrera-Bravo, C.; Koh, E.Y.; Tan, K.S.W. The roles of parasite-derived extracellular vesicles in disease and host-parasite communication. Parasitol. Int. 2021, 83, 102373. [Google Scholar] [CrossRef]
- de Souza, W.; Barrias, E.S. Membrane-bound extracellular vesicles secreted by parasitic protozoa: Cellular structures involved in the communication between cells. Parasitol. Res. 2020, 119, 2005–2023. [Google Scholar] [CrossRef] [PubMed]
- Cortes-Serra, N.; Gualdron-Lopez, M.; Pinazo, M.J.; Torrecilhas, A.C.; Fernandez-Becerra, C. Extracellular Vesicles in Trypanosoma cruzi Infection: Immunomodulatory Effects and Future Perspectives as Potential Control Tools against Chagas Disease. J. Immunol. Res. 2022, 2022, 5230603. [Google Scholar] [CrossRef] [PubMed]
- WHO. World Malaria Report 2022. 2022. Available online: https://www.who.int/publications/i/item/9789240064898 (accessed on 31 July 2023).
- Conroy, A.L.; Datta, D.; John, C.C. What causes severe malaria and its complications in children? Lessons learned over the past 15 years. BMC Med. 2019, 17, 52. [Google Scholar] [CrossRef] [PubMed]
- Severe malaria. Trop. Med. Int. Health 2014, 19 (Suppl. S1), 7–131. [CrossRef] [PubMed]
- WHO. Guidelines for the Treatment of Malaria, 3rd ed.; WHO: Geneva, Switzerland, 2015; Available online: https://www.afro.who.int/publications/guidelines-treatment-malaria-third-edition (accessed on 31 July 2023).
- Wassmer, S.C.; Grau, G.E. Severe malaria: What’s new on the pathogenesis front? Int. J. Parasitol. 2017, 47, 145–152. [Google Scholar] [CrossRef]
- Cowman, A.F.; Healer, J.; Marapana, D.; Marsh, K. Malaria: Biology and Disease. Cell 2016, 167, 610–624. [Google Scholar] [CrossRef]
- Anstey, N.M.; Grigg, M.J.; Rajahram, G.S.; Cooper, D.J.; William, T.; Kho, S.; Barber, B.E. Knowlesi malaria: Human risk factors, clinical spectrum, and pathophysiology. Adv. Parasitol. 2021, 113, 1–43. [Google Scholar]
- Kurtovic, L.; Boyle, M.J.; Opi, D.H.; Kennedy, A.T.; Tham, W.H.; Reiling, L.; Chan, J.A.; Beeson, J.G. Complement in malaria immunity and vaccines. Immunol. Rev. 2020, 293, 38–56. [Google Scholar] [CrossRef]
- Debs, S.; Cohen, A.; Hosseini-Beheshti, E.; Chimini, G.; Hunt, N.H.; Grau, G.E.R. Interplay of extracellular vesicles and other players in cerebral malaria pathogenesis. Biochim. Et Biophys. Acta Gen. Subj. 2019, 1863, 325–331. [Google Scholar] [CrossRef]
- Muppidi, P.; Wright, E.; Wassmer, S.C.; Gupta, H. Diagnosis of cerebral malaria: Tools to reduce Plasmodium falciparum associated mortality. Front. Cell. Infect. Microbiol. 2023, 13, 1090013. [Google Scholar] [CrossRef]
- Ghazanfari, N.; Mueller, S.N.; Heath, W.R. Cerebral Malaria in Mouse and Man. Front. Immunol. 2018, 9, 2016. [Google Scholar] [CrossRef]
- Sierro, F.; Grau, G.E.R. The Ins and Outs of Cerebral Malaria Pathogenesis: Immunopathology, Extracellular Vesicles, Immunometabolism, and Trained Immunity. Front. Immunol. 2019, 10, 830. [Google Scholar] [CrossRef]
- Olatunde, A.C.; Cornwall, D.H.; Roedel, M.; Lamb, T.J. Mouse Models for Unravelling Immunology of Blood Stage Malaria. Vaccines 2022, 10, 1525. [Google Scholar] [CrossRef]
- Combes, V.; Taylor, T.E.; Juhan-Vague, I.; Mege, J.L.; Mwenechanya, J.; Tembo, M.; Grau, G.E.; Molyneux, M.E. Circulating endothelial microparticles in malawian children with severe falciparum malaria complicated with coma. JAMA 2004, 291, 2542–2544. [Google Scholar]
- Pankoui Mfonkeu, J.B.; Gouado, I.; Fotso Kuate, H.; Zambou, O.; Amvam Zollo, P.H.; Grau, G.E.; Combes, V. Elevated cell-specific microparticles are a biological marker for cerebral dysfunctions in human severe malaria. PLoS ONE 2010, 5, e13415. [Google Scholar] [CrossRef]
- Nantakomol, D.; Dondorp, A.M.; Krudsood, S.; Udomsangpetch, R.; Pattanapanyasat, K.; Combes, V.; Grau, G.E.; White, N.J.; Viriyavejakul, P.; Day, N.P.; et al. Circulating red cell-derived microparticles in human malaria. J. Infect. Dis. 2011, 203, 700–706. [Google Scholar] [CrossRef]
- Antwi-Baffour, S.; Malibha-Pinchbeck, M.; Stratton, D.; Jorfi, S.; Lange, S.; Inal, J. Plasma mEV levels in Ghanain malaria patients with low parasitaemia are higher than those of healthy controls, raising the potential for parasite markers in mEVs as diagnostic targets. J. Extracell. Vesicles 2020, 9, 1697124. [Google Scholar] [CrossRef]
- Jung, A.L.; Moller Jorgensen, M.; Baek, R.; Artho, M.; Griss, K.; Han, M.; Bertrams, W.; Greulich, T.; Koczulla, R.; Hippenstiel, S.; et al. Surface proteome of plasma extracellular vesicles as mechanistic and clinical biomarkers for malaria. Infection 2023. Online ahead of print. [Google Scholar] [CrossRef]
- Campos, F.M.; Franklin, B.S.; Teixeira-Carvalho, A.; Filho, A.L.; de Paula, S.C.; Fontes, C.J.; Brito, C.F.; Carvalho, L.H. Augmented plasma microparticles during acute Plasmodium vivax infection. Malar. J. 2010, 9, 327. [Google Scholar] [CrossRef]
- Toda, H.; Diaz-Varela, M.; Segui-Barber, J.; Roobsoong, W.; Baro, B.; Garcia-Silva, S.; Galiano, A.; Gualdrón-López, M.; Almeida, A.C.G.; Brito, M.A.M.; et al. Plasma-derived extracellular vesicles from Plasmodium vivax patients signal spleen fibroblasts via NF-kB facilitating parasite cytoadherence. Nat. Commun. 2020, 11, 2761. [Google Scholar] [CrossRef] [PubMed]
- Piguet, P.F.; Kan, C.D.; Vesin, C. Thrombocytopenia in an animal model of malaria is associated with an increased caspase-mediated death of thrombocytes. Apoptosis Int. J. Program. Cell Death 2002, 7, 91–98. [Google Scholar] [CrossRef] [PubMed]
- El-Assaad, F.; Wheway, J.; Hunt, N.H.; Grau, G.E.; Combes, V. Production, fate and pathogenicity of plasma microparticles in murine cerebral malaria. PLoS Pathog. 2014, 10, e1003839. [Google Scholar] [CrossRef]
- Faille, D.; Combes, V.; Mitchell, A.J.; Fontaine, A.; Juhan-Vague, I.; Alessi, M.C.; Chimini, G.; Fusai, T.; Grau, G.E. Platelet microparticles: A new player in malaria parasite cytoadherence to human brain endothelium. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2009, 23, 3449–3458. [Google Scholar] [CrossRef] [PubMed]
- Martianez-Vendrell, X.; Skjefte, M.; Sikka, R.; Gupta, H. Factors Affecting the Performance of HRP2-Based Malaria Rapid Diagnostic Tests. Trop. Med. Infect. Dis. 2022, 7, 265. [Google Scholar] [CrossRef]
- Kavanaugh, M.J.; Azzam, S.E.; Rockabrand, D.M. Malaria Rapid Diagnostic Tests: Literary Review and Recommendation for a Quality Assurance, Quality Control Algorithm. Diagnostics 2021, 11, 768. [Google Scholar] [CrossRef]
- Craig, A.G.; Khairul, M.F.; Patil, P.R. Cytoadherence and severe malaria. Malays. J. Med. Sci. MJMS 2012, 19, 5–18. [Google Scholar]
- Cunningham, D.A.; Lin, J.W.; Brugat, T.; Jarra, W.; Tumwine, I.; Kushinga, G.; Ramesar, J.; Franke-Fayard, B.; Langhorne, J. ICAM-1 is a key receptor mediating cytoadherence and pathology in the Plasmodium chabaudi malaria model. Malar. J. 2017, 16, 185. [Google Scholar] [CrossRef]
- Combes, V.; Coltel, N.; Alibert, M.; van Eck, M.; Raymond, C.; Juhan-Vague, I.; Grau, G.E.; Chimini, G. ABCA1 gene deletion protects against cerebral malaria: Potential pathogenic role of microparticles in neuropathology. Am. J. Pathol. 2005, 166, 295–302. [Google Scholar] [CrossRef]
- Antwi-Baffour, S.; Adjei, J.K.; Agyemang-Yeboah, F.; Annani-Akollor, M.; Kyeremeh, R.; Asare, G.A.; Gyan, B. Proteomic analysis of microparticles isolated from malaria positive blood samples. Proteome Sci. 2016, 15, 5. [Google Scholar] [CrossRef]
- Tiberti, N.; Latham, S.L.; Bush, S.; Cohen, A.; Opoka, R.O.; John, C.C.; Juillard, A.; Grau, G.E.; Combes, V. Exploring experimental cerebral malaria pathogenesis through the characterisation of host-derived plasma microparticle protein content. Sci. Rep. 2016, 6, 37871. [Google Scholar] [CrossRef] [PubMed]
- Martin-Jaular, L.; Nakayasu, E.S.; Ferrer, M.; Almeida, I.C.; Del Portillo, H.A. Exosomes from Plasmodium yoelii-infected reticulocytes protect mice from lethal infections. PLoS ONE 2011, 6, e26588. [Google Scholar] [CrossRef]
- Gualdron-Lopez, M.; Flannery, E.L.; Kangwanrangsan, N.; Chuenchob, V.; Fernandez-Orth, D.; Segui-Barber, J.; Royo, F.; Falcón-Pérez, J.M.; Fernandez-Becerra, C.; Lacerda, M.V.G.; et al. Characterization of Plasmodium vivax Proteins in Plasma-Derived Exosomes From Malaria-Infected Liver-Chimeric Humanized Mice. Front. Microbiol. 2018, 9, 1271. [Google Scholar] [CrossRef] [PubMed]
- Aparici-Herraiz, I.; Gualdron-Lopez, M.; Castro-Cavadia, C.J.; Carmona-Fonseca, J.; Yasnot, M.F.; Fernandez-Becerra, C.; Del Portillo, H.A. Antigen Discovery in Circulating Extracellular Vesicles From Plasmodium vivax Patients. Front. Cell. Infect. Microbiol. 2021, 11, 811390. [Google Scholar] [CrossRef] [PubMed]
- Esteves, S.; Lima, C.; Costa, I.; Osorio, H.; Fernandez-Becerra, C.; Santarem, N.; Cordeiro-da-Silva, A. Characterization and Proteomic Analysis of Plasma EVs Recovered from Healthy and Diseased Dogs with Canine Leishmaniosis. Int. J. Mol. Sci. 2023, 24, 5490. [Google Scholar] [CrossRef] [PubMed]
- Cortes-Serra, N.; Mendes, M.T.; Mazagatos, C.; Segui-Barber, J.; Ellis, C.C.; Ballart, C.; Garcia-Alvarez, A.; Gallego, M.; Gascon, J.; Almeida, I.C.; et al. Plasma-Derived Extracellular Vesicles as Potential Biomarkers in Heart Transplant Patient with Chronic Chagas Disease. Emerg. Infect. Dis. 2020, 26, 1846–1851. [Google Scholar] [CrossRef]
- Ketprasit, N.; Cheng, I.S.; Deutsch, F.; Tran, N.; Imwong, M.; Combes, V.; Palasuwan, D. The characterization of extracellular vesicles-derived microRNAs in Thai malaria patients. Malar. J. 2020, 19, 285. [Google Scholar] [CrossRef]
- Cohen, A.; Zinger, A.; Tiberti, N.; Grau, G.E.R.; Combes, V. Differential plasma microvesicle and brain profiles of microRNA in experimental cerebral malaria. Malar. J. 2018, 17, 192. [Google Scholar] [CrossRef]
- Kusakisako, K.; Nakao, R.; Katakura, K. Detection of parasite-derived tRNA and rRNA fragments in the peripheral blood of mice experimentally infected with Leishmania donovani and Leishmania amazonensis using next-generation sequencing analysis. Parasitol. Int. 2023, 93, 102716. [Google Scholar] [CrossRef]
- Ballinas-Verdugo, M.A.; Jimenez-Ortega, R.F.; Martinez-Martinez, E.; Rivas, N.; Contreras-Lopez, E.A.; Carbo, R.; Sanchez, F.; Bojalil, R.; Marquez-Velasco, R.; Sanchez-Munoz, F.; et al. Circulating miR-146a as a possible candidate biomarker in the indeterminate phase of Chagas disease. Biol. Res. 2021, 54, 21. [Google Scholar] [CrossRef] [PubMed]
- da Cruz, A.B.; Maia, M.M.; Pereira, I.S.; Taniwaki, N.N.; Namiyama, G.M.; Telles, J.P.M.; Vidal, J.E.; Spegiorin, L.; Brandao de Mattos, C.C.; Mattos, L.C.; et al. Human extracellular vesicles and correlation with two clinical forms of toxoplasmosis. PLoS ONE 2020, 15, e0229602. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, F.; Baur, A. Biogenesis and Functions of Exosomes and Extracellular Vesicles. Methods Mol. Biol. 2016, 1448, 201–216. [Google Scholar]
- Martin-Jaular, L.; de Menezes-Neto, A.; Monguio-Tortajada, M.; Elizalde-Torrent, A.; Diaz-Varela, M.; Fernandez-Becerra, C.; Borras, F.E.; Montoya, M.; Del Portillo, H.A. Spleen-Dependent Immune Protection Elicited by CpG Adjuvanted Reticulocyte-Derived Exosomes from Malaria Infection Is Associated with Changes in T cell Subsets’ Distribution. Front. Cell Dev. Biol. 2016, 4, 131. [Google Scholar] [CrossRef] [PubMed]
- Mantel, P.Y.; Hoang, A.N.; Goldowitz, I.; Potashnikova, D.; Hamza, B.; Vorobjev, I.; Ghiran, I.; Toner, M.; Irimia, D.; Ivanov, A.R.; et al. Malaria-infected erythrocyte-derived microvesicles mediate cellular communication within the parasite population and with the host immune system. Cell Host Microbe 2013, 13, 521–534. [Google Scholar] [CrossRef]
- Regev-Rudzki, N.; Wilson, D.W.; Carvalho, T.G.; Sisquella, X.; Coleman, B.M.; Rug, M.; Bursac, D.; Angrisano, F.; Gee, M.; Hill, A.F.; et al. Cell-cell communication between malaria-infected red blood cells via exosome-like vesicles. Cell 2013, 153, 1120–1133. [Google Scholar] [CrossRef]
- Couper, K.N.; Barnes, T.; Hafalla, J.C.; Combes, V.; Ryffel, B.; Secher, T.; Grau, G.E.; Riley, E.M.; de Souza, J.B. Parasite-derived plasma microparticles contribute significantly to malaria infection-induced inflammation through potent macrophage stimulation. PLoS Pathog. 2010, 6, e1000744. [Google Scholar] [CrossRef]
- Mantel, P.Y.; Hjelmqvist, D.; Walch, M.; Kharoubi-Hess, S.; Nilsson, S.; Ravel, D.; Ribeiro, M.; Grüring, C.; Ma, S.; Padmanabhan, P.; et al. Infected erythrocyte-derived extracellular vesicles alter vascular function via regulatory Ago2-miRNA complexes in malaria. Nat. Commun. 2016, 7, 12727. [Google Scholar] [CrossRef]
- van Eijndhoven, M.A.; Zijlstra, J.M.; Groenewegen, N.J.; Drees, E.E.; van Niele, S.; Baglio, S.R.; Koppers-Lalic, D.; van der Voorn, H.; Libregts, S.F.; Wauben, M.H.; et al. Plasma vesicle miRNAs for therapy response monitoring in Hodgkin lymphoma patients. JCI Insight 2016, 1, e89631. [Google Scholar] [CrossRef]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef]
- El-Assaad, F.; Hempel, C.; Combes, V.; Mitchell, A.J.; Ball, H.J.; Kurtzhals, J.A.; Hunt, N.H.; Mathys, J.M.; Grau, G.E. Differential microRNA expression in experimental cerebral and noncerebral malaria. Infect. Immun. 2011, 79, 2379–2384. [Google Scholar] [CrossRef] [PubMed]
- Chamnanchanunt, S.; Kuroki, C.; Desakorn, V.; Enomoto, M.; Thanachartwet, V.; Sahassananda, D.; Sattabongkot, J.; Jenwithisuk, R.; Fucharoen, S.; Svasti, S.; et al. Downregulation of plasma miR-451 and miR-16 in Plasmodium vivax infection. Exp. Parasitol. 2015, 155, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Gupta, H.; Sahu, P.K.; Pattnaik, R.; Mohanty, A.; Majhi, M.; Mohanty, A.K.; Pirpamer, L.; Hoffmann, A.; Mohanty, S.; Wassmer, S.C. Plasma levels of hsa-miR-3158-3p microRNA on admission correlate with MRI findings and predict outcome in cerebral malaria. Clin. Transl. Med. 2021, 11, e396. [Google Scholar] [CrossRef]
- Gupta, H.; Wassmer, S.C. Harnessing the Potential of miRNAs in Malaria Diagnostic and Prevention. Front. Cell. Infect. Microbiol. 2021, 11, 793954. [Google Scholar] [CrossRef] [PubMed]
- Vickers, K.C.; Shoucri, B.M.; Levin, M.G.; Wu, H.; Pearson, D.S.; Osei-Hwedieh, D.; Collins, F.S.; Remaley, A.T.; Sethupathy, P. MicroRNA-27b is a regulatory hub in lipid metabolism and is altered in dyslipidemia. Hepatology 2013, 57, 533–542. [Google Scholar] [CrossRef]
- Akhoundi, M.; Kuhls, K.; Cannet, A.; Votypka, J.; Marty, P.; Delaunay, P.; Sereno, D. A Historical Overview of the Classification, Evolution, and Dispersion of Leishmania Parasites and Sandflies. PLoS Neglected Trop. Dis. 2016, 10, e0004349. [Google Scholar] [CrossRef]
- WHO. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 20 June 2023).
- Almeida, F.S.; Vanderley, S.E.R.; Comberlang, F.C.; Andrade, A.G.; Cavalcante-Silva, L.H.A.; Silva, E.D.S.; Palmeira, P.H.S.; Amaral, I.; Keesen, T.S.L. Leishmaniasis: Immune Cells Crosstalk in Macrophage Polarization. Trop. Med. Infect. Dis. 2023, 8, 276. [Google Scholar] [CrossRef]
- Costa-da-Silva, A.C.; Nascimento, D.O.; Ferreira, J.R.M.; Guimaraes-Pinto, K.; Freire-de-Lima, L.; Morrot, A.; Decote-Ricardo, D.; Filardy, A.A.; Freire-de-Lima, C.G. Immune Responses in Leishmaniasis: An Overview. Trop. Med. Infect. Dis. 2022, 7, 54. [Google Scholar] [CrossRef]
- Silverman, J.M.; Clos, J.; de’Oliveira, C.C.; Shirvani, O.; Fang, Y.; Wang, C.; Foster, L.J.; Reiner, N.E. An exosome-based secretion pathway is responsible for protein export from Leishmania and communication with macrophages. J. Cell Sci. 2010, 123 Pt 6, 842–852. [Google Scholar] [CrossRef]
- Rossi, I.V.; Ferreira Nunes, M.A.; Vargas-Otalora, S.; da Silva Ferreira, T.C.; Cortez, M.; Ramirez, M.I. Extracellular Vesicles during TriTryps infection: Complexity and future challenges. Mol. Immunol. 2021, 132, 172–183. [Google Scholar] [CrossRef]
- Silverman, J.M.; Reiner, N.E. Leishmania exosomes deliver preemptive strikes to create an environment permissive for early infection. Front. Cell. Infect. Microbiol. 2011, 1, 26. [Google Scholar] [CrossRef] [PubMed]
- Douanne, N.; Dong, G.; Amin, A.; Bernardo, L.; Blanchette, M.; Langlais, D.; Olivier, M.; Fernandez-Prada, C. Leishmania parasites exchange drug-resistance genes through extracellular vesicles. Cell Rep. 2022, 40, 111121. [Google Scholar] [CrossRef]
- Soto-Serna, L.E.; Diupotex, M.; Zamora-Chimal, J.; Ruiz-Remigio, A.; Delgado-Dominguez, J.; Cervantes-Sarabia, R.B.; Mendez-Bernal, A.; Escalona-Montano, A.R.; Aguirre-Garcia, M.M.; Becker, I. Leishmania mexicana: Novel Insights of Immune Modulation through Amastigote Exosomes. J. Immunol. Res. 2020, 2020, 8894549. [Google Scholar] [CrossRef]
- Hassani, K.; Olivier, M. Immunomodulatory impact of leishmania-induced macrophage exosomes: A comparative proteomic and functional analysis. PLoS Neglected Trop. Dis. 2013, 7, e2185. [Google Scholar] [CrossRef] [PubMed]
- Gioseffi, A.; Hamerly, T.; Van, K.; Zhang, N.; Dinglasan, R.R.; Yates, P.A.; Kima, P.E. Leishmania-infected macrophages release extracellular vesicles that can promote lesion development. Life Sci. Alliance 2020, 3, e202000742. [Google Scholar] [CrossRef] [PubMed]
- Schnitzer, J.K.; Berzel, S.; Fajardo-Moser, M.; Remer, K.A.; Moll, H. Fragments of antigen-loaded dendritic cells (DC) and DC-derived exosomes induce protective immunity against Leishmania major. Vaccine 2010, 28, 5785–5793. [Google Scholar] [CrossRef]
- Toledo, M.D.S.; Cronemberger-Andrade, A.; Barbosa, F.M.C.; Reis, N.F.C.; Dupin, T.V.; Soares, R.P.; Torrecilhas, A.C.; Xander, P. Effects of extracellular vesicles released by peritoneal B-1 cells on experimental Leishmania (Leishmania) amazonensis infection. J. Leukoc. Biol. 2020, 108, 1803–1814. [Google Scholar] [CrossRef]
- Cronemberger-Andrade, A.; Aragao-Franca, L.; de Araujo, C.F.; Rocha, V.J.; Borges-Silva Mda, C.; Figueira, C.P.; Oliveira, P.R.; de Freitas, L.A.; Veras, P.S.; Pontes-de-Carvalho, L. Extracellular vesicles from Leishmania-infected macrophages confer an anti-infection cytokine-production profile to naive macrophages. PLoS Neglected Trop. Dis. 2014, 8, e3161. [Google Scholar] [CrossRef]
- da Cruz, A.B.; Carneiro, F.M.; Maia, M.M.; Pereira, I.S.; Taniwaki, N.N.; Namiyama, G.M.; Gava, R.; Hiramoto, R.M.; Pereira-Chioccola, V.L. Dogs with canine visceral leishmaniasis have a boost of extracellular vesicles and miR-21-5p up-expression. Parasite Immunol. 2023, 45, e13004. [Google Scholar] [CrossRef]
- Chowdhury, I.H.; Koo, S.J.; Gupta, S.; Liang, L.Y.; Bahar, B.; Silla, L.; Nunez-Burgos, J.; Barrientos, N.; Zago, M.P.; Garg, N.J. Gene Expression Profiling and Functional Characterization of Macrophages in Response to Circulatory Microparticles Produced during Trypanosoma cruzi Infection and Chagas Disease. J. Innate Immun. 2017, 9, 203–216. [Google Scholar] [CrossRef]
- Ramirez, M.I.; Deolindo, P.; de Messias-Reason, I.J.; Arigi, E.A.; Choi, H.; Almeida, I.C.; Evans-Osses, I. Dynamic flux of microvesicles modulate parasite-host cell interaction of Trypanosoma cruzi in eukaryotic cells. Cell. Microbiol. 2017, 19, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Madeira, R.P.; Dal’Mas Romera, L.M.; de Cassia Buck, P.; Mady, C.; Ianni, B.M.; Torrecilhas, A.C. New Biomarker in Chagas Disease: Extracellular Vesicles Isolated from Peripheral Blood in Chronic Chagas Disease Patients Modulate the Human Immune Response. J. Immunol. Res. 2021, 2021, 6650670. [Google Scholar] [CrossRef] [PubMed]
- Dozio, V.; Lejon, V.; Mumba Ngoyi, D.; Buscher, P.; Sanchez, J.C.; Tiberti, N. Cerebrospinal Fluid-Derived Microvesicles From Sleeping Sickness Patients Alter Protein Expression in Human Astrocytes. Front. Cell. Infect. Microbiol. 2019, 9, 391. [Google Scholar] [CrossRef] [PubMed]
- Maia, M.M.; da Cruz, A.B.; Pereira, I.S.; Taniwaki, N.N.; Namiyama, G.M.; Pereira-Chioccola, V.L. Characterization of murine extracellular vesicles and Toxoplasma gondii infection. Parasite Immunol. 2021, 43, e12869. [Google Scholar] [CrossRef]
- Perez-Molina, J.A.; Molina, I. Chagas disease. Lancet 2018, 391, 82–94. [Google Scholar] [CrossRef]
- Guarner, J. Chagas disease as example of a reemerging parasite. Semin. Diagn. Pathol. 2019, 36, 164–169. [Google Scholar] [CrossRef]
- WHO. Ending the Neglect to Attain the Sustainable Development Goals: A Road Map for Neglected Tropical Diseases 2021–2030. 2021. Available online: https://www.who.int/publications/i/item/9789240010352 (accessed on 31 July 2023).
- Burleigh, B.A.; Woolsey, A.M. Cell signalling and Trypanosoma cruzi invasion. Cell. Microbiol. 2002, 4, 701–711. [Google Scholar] [CrossRef]
- Boldt, A.B.; Luz, P.R.; Messias-Reason, I.J. MASP2 haplotypes are associated with high risk of cardiomyopathy in chronic Chagas disease. Clin. Immunol. 2011, 140, 63–70. [Google Scholar] [CrossRef]
- Corsten, M.F.; Dennert, R.; Jochems, S.; Kuznetsova, T.; Devaux, Y.; Hofstra, L.; Wagner, D.R.; Staessen, J.A.; Heymans, S.; Schroen, B. Circulating MicroRNA-208b and MicroRNA-499 reflect myocardial damage in cardiovascular disease. Circ. Cardiovasc. Genet. 2010, 3, 499–506. [Google Scholar] [CrossRef]
- Cestari, I.; Ansa-Addo, E.; Deolindo, P.; Inal, J.M.; Ramirez, M.I. Trypanosoma cruzi immune evasion mediated by host cell-derived microvesicles. J. Immunol. 2012, 188, 1942–1952. [Google Scholar] [CrossRef]
- Buscher, P.; Cecchi, G.; Jamonneau, V.; Priotto, G. Human African trypanosomiasis. Lancet 2017, 390, 2397–2409. [Google Scholar] [CrossRef] [PubMed]
- Pays, E.; Radwanska, M.; Magez, S. The Pathogenesis of African Trypanosomiasis. Annu. Rev. Pathol. 2023, 18, 19–45. [Google Scholar] [CrossRef] [PubMed]
- Franco, J.R.; Cecchi, G.; Paone, M.; Diarra, A.; Grout, L.; Kadima Ebeja, A.; Simarro, P.P.; Zhao, W.; Argaw, D. The elimination of human African trypanosomiasis: Achievements in relation to WHO road map targets for 2020. PLoS Neglected Trop. Dis. 2022, 16, e0010047. [Google Scholar] [CrossRef] [PubMed]
- WHO. Control and Surveillance of Human African Trypanosomiasis: Report of a WHO Expert Committee. 2013. Available online: https://apps.who.int/iris/handle/10665/95732 (accessed on 31 July 2023).
- Kennedy, P.G.E.; Rodgers, J. Clinical and Neuropathogenetic Aspects of Human African Trypanosomiasis. Front. Immunol. 2019, 10, 39. [Google Scholar] [CrossRef] [PubMed]
- Atyame Nten, C.M.; Sommerer, N.; Rofidal, V.; Hirtz, C.; Rossignol, M.; Cuny, G.; Peltier, J.B.; Geiger, A. Excreted/secreted proteins from trypanosome procyclic strains. J. Biomed. Biotechnol. 2010, 2010, 212817. [Google Scholar] [CrossRef] [PubMed]
- Szempruch, A.J.; Sykes, S.E.; Kieft, R.; Dennison, L.; Becker, A.C.; Gartrell, A.; Martin, W.J.; Nakayasu, E.S.; Almeida, I.C.; Hajduk, S.L.; et al. Extracellular Vesicles from Trypanosoma brucei Mediate Virulence Factor Transfer and Cause Host Anemia. Cell 2016, 164, 246–257. [Google Scholar] [CrossRef]
- Eliaz, D.; Kannan, S.; Shaked, H.; Arvatz, G.; Tkacz, I.D.; Binder, L.; Waldman Ben-Asher, H.; Okalang, U.; Chikne, V.; Cohen-Chalamish, S.; et al. Exosome secretion affects social motility in Trypanosoma brucei. PLoS Pathog. 2017, 13, e1006245. [Google Scholar] [CrossRef]
- Dias-Guerreiro, T.; Palma-Marques, J.; Mourata-Goncalves, P.; Alexandre-Pires, G.; Valerio-Bolas, A.; Gabriel, A.; Nunes, T.; Antunes, W.; Fonseca, I.P.D.; Sousa-Silva, M.; et al. African Trypanosomiasis: Extracellular Vesicles Shed by Trypanosoma brucei brucei Manipulate Host Mononuclear Cells. Biomedicines 2021, 9, 1056. [Google Scholar] [CrossRef]
- Smith, N.C.; Goulart, C.; Hayward, J.A.; Kupz, A.; Miller, C.M.; van Dooren, G.G. Control of human toxoplasmosis. Int. J. Parasitol. 2021, 51, 95–121. [Google Scholar] [CrossRef]
- Gubbels, M.J.; White, M.; Szatanek, T. The cell cycle and Toxoplasma gondii cell division: Tightly knit or loosely stitched? Int. J. Parasitol. 2008, 38, 1343–1358. [Google Scholar] [CrossRef]
- Pappas, G.; Roussos, N.; Falagas, M.E. Toxoplasmosis snapshots: Global status of Toxoplasma gondii seroprevalence and implications for pregnancy and congenital toxoplasmosis. Int. J. Parasitol. 2009, 39, 1385–1394. [Google Scholar] [CrossRef] [PubMed]
- Molan, A.; Nosaka, K.; Hunter, M.; Wang, W. Global status of Toxoplasma gondii infection: Systematic review and prevalence snapshots. Trop. Biomed. 2019, 36, 898–925. [Google Scholar] [PubMed]
- Xiao, J.; Yolken, R.H. Strain hypothesis of Toxoplasma gondii infection on the outcome of human diseases. Acta Physiol. 2015, 213, 828–845. [Google Scholar] [CrossRef]
- Dubey, J.P.; Murata, F.H.A.; Cerqueira-Cezar, C.K.; Kwok, O.C.H.; Villena, I. Congenital toxoplasmosis in humans: An update of worldwide rate of congenital infections. Parasitology 2021, 148, 1406–1416. [Google Scholar] [CrossRef] [PubMed]
- Schluter, D.; Barragan, A. Advances and Challenges in Understanding Cerebral Toxoplasmosis. Front. Immunol. 2019, 10, 242. [Google Scholar] [CrossRef]
- Gomez-Chavez, F.; Murrieta-Coxca, J.M.; Caballero-Ortega, H.; Morales-Prieto, D.M.; Markert, U.R. Host-pathogen interactions mediated by extracellular vesicles in Toxoplasma gondii infection during pregnancy. J. Reprod. Immunol. 2023, 158, 103957. [Google Scholar] [CrossRef]
- Olajide, J.S.; Cai, J. Perils and Promises of Pathogenic Protozoan Extracellular Vesicles. Front. Cell. Infect. Microbiol. 2020, 10, 371. [Google Scholar] [CrossRef]
- Ozturk, E.A.; Caner, A. Liquid Biopsy for Promising Non-invasive Diagnostic Biomarkers in Parasitic Infections. Acta Parasitol. 2022, 67, 1–17. [Google Scholar] [CrossRef]
- Drurey, C.; Coakley, G.; Maizels, R.M. Extracellular vesicles: New targets for vaccines against helminth parasites. Int. J. Parasitol. 2020, 50, 623–633. [Google Scholar] [CrossRef]
- Santos, P.; Almeida, F. Exosome-Based Vaccines: History, Current State, and Clinical Trials. Front. Immunol. 2021, 12, 711565. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Population | Infecting Pathogen | EV Source | Technique for EVs Analysis | Main Observations of the Study | Ref. |
|---|---|---|---|---|---|
| Children with malaria (Malawi) | P. falciparum | Plasma | FC: CD51+ E-MV |
| [38] |
| Children with malaria (Cameroon) | P. falciparum | Plasma | FC: AnV+ total MV CD51+ and CD105+ E-MV CD41+ P-MV CD235a+ R-MV CD11b+ M-MV CD3+ Ly-MV |
| [39] |
| Adults with malaria (Thailand) | P. falciparum P. vivax P. malariae | Plasma | FC: AnV+ total MV CD235a+ R-MV |
| [40] |
| Adults and children with malaria (Ghana) | P. falciparum | Plasma | High-speed centrifugation NTA TEM WB FC: AnV+ total MV |
| [41] |
| Adults (travellers) with malaria | P. falciparum | Plasma | EVs microarray targeting 40 antigens |
| [42] |
| Patients with malaria | P. vivax | Plasma | High-speed centrifugation FC: AnV+ total MV CD144+ E-MV CD41a+ P-MV CD235a+ R-MV CD14+ M-MV CD45+ L-MV |
| [43] |
| Patients with malaria (Brazil and Colombia) | P. vivax | Plasma | SEC NTA Bead-based FC Proteomics |
| [44] |
| C57BL/6J mice | P. berghei ANKA | Plasma | FC: Anti-platelet antibody |
| [45] |
| CBA mice | P. berghei ANKA P. berghei K173 | Plasma | FC: AnV+ total MV CD105+ E-MV CD41+ P-MV CD235a+ R-MV CD11b+ M-MV CD45+ L-MV |
| [46] |
| Study Population | Infecting Pathogen | EV Source | Technique for EVs Analysis | # ID, Host-Derived | # ID, Parasite- Derived | Main Observations of the Study | Ref. |
|---|---|---|---|---|---|---|---|
| Adults and children with malaria (Ghana) | P. falciparum | Plasma | High-speed centrifugation FC: AnV+ total MV LC-MS/MS | Patients: 1729 Controls: 234 Culture SN: 333 | 29, P. falciparum |
| [53] |
| DBA/1 and C57BL/6 mice | P. berghei ANKA | Plasma | High-speed centrifugation FC SEM LC-MS/MS WB | 368 | 2, P. berghei |
| [54] |
| BALB/c mice | P. yoelii lethal and non-lethal strains | Plasma | Differential UC TEM Bead-based FC LC-MS/MS | 259 | 31, P. yoelii |
| [55] |
| FRG huEP chimeric mice | P. vivax | Plasma | SEC Bead-based FC (CD5L+) NTA Cryo-EM LC-MS/MS | 290, H. sapiens 234, M. musculus | 17, P. vivax |
| [56] |
| Malaria patients (Brazil and Colombia) | P. vivax | Plasma | SEC NTA Bead-based FC LC-MS/MS | 533 | 20, P. vivax |
| [44] |
| Malaria patients (Colombia and travellers) | P. vivax | Plasma | UC + direct immunoaffinity capture (CD71+) WB LC-MS/MS Bead-based Luminex assay | 440 | 48, P. vivax |
| [57] |
| Dogs | L. infantum | Plasma | SEC Bead-based FC NTA (CD5L, CD71 and CD9) TEM LC-MS/MS | 1148 | 12, L. infantum |
| [58] |
| Case report: CCD adult patient | T. cruzi | Plasma | SEC NTA Bead-based FC LC-MS/MS | 338 | 12, T. cruzi (1 with high confidence) |
| [59] |
| Study Population | Infecting Pathogen | EV Source | Technique for EVs Analysis | # Match, Host-Derived | # Match, Parasite-Derived | Main Observations of the Study | Ref. |
|---|---|---|---|---|---|---|---|
| Malaria patients (Thailand) | P. falciparum P. vivax | Plasma | UC RT-qPCR | 5 | na |
| [60] |
| CBA mice | P. berghei ANKA P. yoelii | Plasma | High-speed centrifugation miRNA OpenArray RT-qPCR | 431 quantified | na |
| [61] |
| BALB/c mice | L. donovani L. amazonensis | Plasma | Total EXOs isolation kit Small RNA seq | 12,268,998 reads in L. donovani-mice 11,410,173 reads in L. amazonensis-mice | L. donovani hits: 126 in L. donovani-mice 119 in L. amazonensis-mice |
| [62] |
| CD1 mice | T. cruzi | Plasma | UC NTA RT-qPCR | 3 | na |
| [63] |
| Patients with toxoplasmosis (gestational and cerebral + HIV) | T. gondii | Serum CSF | UC NTA TEM WB RT-qPCR | 5 | na |
| [64] |
| Study Population | Pathogen | EV Source | Technique for EVs Analysis | Main Observations of the Study | Ref. |
|---|---|---|---|---|---|
| Dogs | L. infantum | Serum | UC TEM NTA WB |
| [92] |
| Symtomatic and asymptomatic CD patients C57BL/6 mice | T. cruzi | Plasma | High-speed centrifugation FC (AnV, CD14, CD61, CD62E, troponin, CD4 CD8) |
| [93] |
| CCD patients | T. cruzi | Plasma | FC |
| [94] |
| CCD patients | T. cruzi | Plasma | UC NTA |
| [95] |
| HAT patients | T. b. gambiense | CSF | FC (AnV, CTV, CD105, NSE and CD45) High-speed centrifugation TEM |
| [96] |
| A/Sn inbred mice | T. gondii | Serum | UC NTA TEM WB |
| [97] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiberti, N.; Longoni, S.S.; Combes, V.; Piubelli, C. Host-Derived Extracellular Vesicles in Blood and Tissue Human Protozoan Infections. Microorganisms 2023, 11, 2318. https://doi.org/10.3390/microorganisms11092318
Tiberti N, Longoni SS, Combes V, Piubelli C. Host-Derived Extracellular Vesicles in Blood and Tissue Human Protozoan Infections. Microorganisms. 2023; 11(9):2318. https://doi.org/10.3390/microorganisms11092318
Chicago/Turabian StyleTiberti, Natalia, Silvia Stefania Longoni, Valéry Combes, and Chiara Piubelli. 2023. "Host-Derived Extracellular Vesicles in Blood and Tissue Human Protozoan Infections" Microorganisms 11, no. 9: 2318. https://doi.org/10.3390/microorganisms11092318
APA StyleTiberti, N., Longoni, S. S., Combes, V., & Piubelli, C. (2023). Host-Derived Extracellular Vesicles in Blood and Tissue Human Protozoan Infections. Microorganisms, 11(9), 2318. https://doi.org/10.3390/microorganisms11092318