

Prevalence of Aminoglycoside Resistance Genes in Clinical Isolates of Pseudomonas aeruginosa from Taif, Saudi Arabia—An Emergence Indicative Study

Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of P. aeruginosa Isolates
2.2. Antimicrobial Susceptibility Testing (AST)

{kind=link}
{kind=link}
| Primer/Gene | Sequence | PCR Condition | Amplicon Size (pb) | References |
|---|---|---|---|---|
| VIC/algD | F: TTCCCTCGCAGAGAAAACATC R: CCTGGTTGATCAGGTCGATCT | Initial denaturation at 95 °C for 15 min, then 30 cycles of 95 °C for 1 min, 58 °C for 1 min and 72 °C for 5 min and one cycle of final elongation at 72 °C | 520 | [14] |
| GyrA | F: TTATGCCATGAGCGAGCTGGGCAACGACT R: AACCGTTGACCAGCAGGTTGGGAATCTT | Initial denaturation at 95 °C for 15 min, then 35 cycles of 95 °C for 1 min, 57 °C for 1 min, and 72 °C for 5 min, and one cycle of final elongation at 72 °C. | 365 | [2] |
| Aph(3′)-VI | F: ATGGAATTGCCCAATATTATT R: TCAATTCAATTCATCAAGTTT | Initial denaturation at 95 °C for 15 min, then 30 cycles of 95 °C for 1 min, 55 °C for 1 min, and 72 °C for 5 min, and one cycle of final elongation at 72 °C. | 780 | [15] |
| aac(3′)-II/aac(3′)-II | F: ATATCGCGATGCATACGCGG R: GACGGCCTCTAACCGGAAGG | 877 | ||
| aac(6′)-Ib/aac(6′)-Ib | F: TTGCGATGCTCTATGAGTGGCTA R: CTCGAATGCCTGGCGTGTTT | 472 | ||
| aac(6′)-II/aac(6′)-II | F: CGACCATTTCATGTCC R: GAAGGCTTGTCGTGTTT | 542 | ||
| ant(3″)-I/ant(3″)-I/ | F: CATCATGAGGGAAGCGGTG R: GACTACCTTGGTGATCTCG | 787 | ||
| ArmA/ArmA | F: CCGAAATGACAGTTCCTATC R: GAAAATGAGTGCCTTGGAGG | 846 | ||
| rmtB/rmtB | F: ATGAACATCAACGATGCCCTC R: CCTTCTGATTGGCTTATCCA | Initial denaturation at 95 °C for 15 min, then 30 cycles of 95 °C for 1min, 60 °C for 1 min, and 72 °C for 5 min, and one cycle of final elongation at 72 °C. | 769 | [15] |
2.3. Genomic DNA Extraction
2.4. PCR and Gel Electrophoresis
2.5. DNA Sequencing
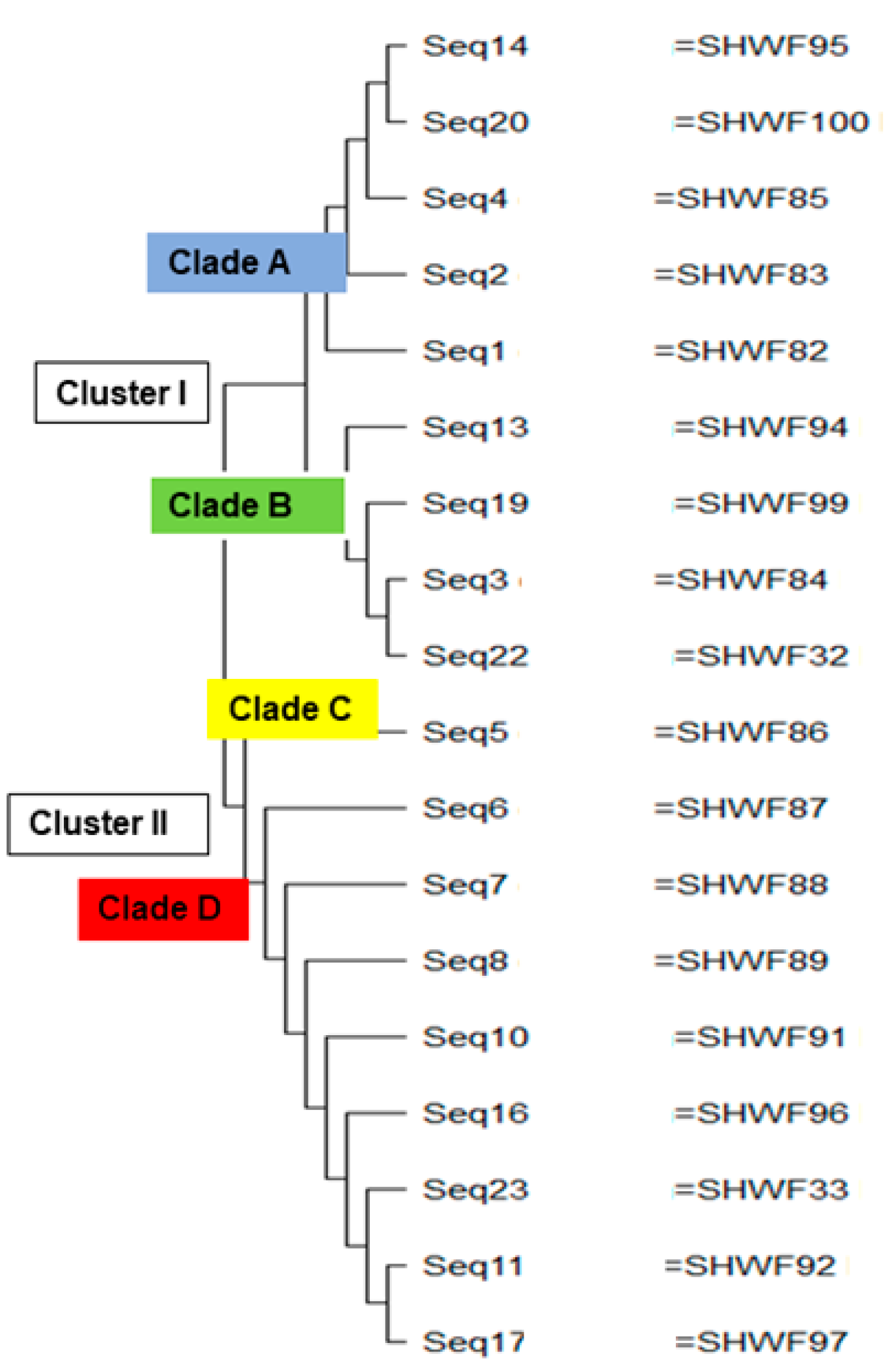
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Rawson, T.M.; Ming, D.; Ahmad, R.; Moore, L.S.P.; Holmes, A.H. Antimicrobial use, drug-resistant infections and COVID-19. Nat. Rev. Microbiol. 2020, 18, 409–410. [Google Scholar] [CrossRef]
- Nguyen, K.V.; Nguyen, T.V.; Nguyen, H.T.T.; Le, D.V. Mutations in the gyrA, parC, and mexR genes provide functional insights into the fluoroquinolone-resistant Pseudomonas aeruginosa isolated in Vietnam. Infect Drug Resist. 2018, 28, 275–282. [Google Scholar] [CrossRef]
- Atassi, G.; Medernach, R.; Scheetz, M.; Nozick, S.; Rhodes, N.J.; Murphy-Belcaster, M.; Murphy, K.R.; Alisoltani, A.; Ozer, E.A.; Hauser, A.R. Genomics of Aminoglycoside Resistance in Pseudomonas aeruginosa Bloodstream Infections at a United States Academic Hospital. Microbiol. Spectr. 2023, 11, e0508722. [Google Scholar] [CrossRef]
- Yang, W.; Hu, F. Research Updates of Plasmid-Mediated Aminoglycoside Resistance 16S rRNA Methyltransferase. Antibiotics 2022, 11, 906. [Google Scholar] [CrossRef]
- Savaş, L.; Duran, N.; Savaş, N.; Önlen YOcak, S. The prevalence and resistance patterns of Pseudomonas aeruginosa in intensive care units in a university hospital. Turk. J. Med Sci. 2005, 35, 317–322. [Google Scholar]
- Asghar, A.H.; Ahmed, O.B. Prevalence of aminoglycoside resistance genes in Pseudomonas aeruginosa isolated from a tertiary care hospital in Makkah, KSA. Clin. Pract. 2018, 15, 438–441. [Google Scholar] [CrossRef]
- Ramirez, M.S.; Nikolaidis, N.; Tolmasky, M.E. Rise and dissemination of aminoglycoside resistance: The aac(6′)-Ib paradigm. Front. Microbiol. 2013, 4, 121. [Google Scholar] [CrossRef]
- Dubois, V.; Poirel, L.; Marie, C.; Arpin, C.; Nordmann, P.; Quentin, C. Molecular characterization of a novel class 1 integron containing blaGES-1 and a fused product of aac(3)-Ib/aac(6″)-Ib″ gene cassettes in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2002, 46, 638–645. [Google Scholar] [CrossRef]
- Yamane, K.; Doi, Y.; Yokoyama, K.; Yagi, T.; Kurokawa, H.; Shibata, N.; Shibayama, K.; Kato, H.; Arakawa, Y. Genetic Environments of the rmtA Gene in Pseudomonas aeruginosa Clinical Isolates. Antimicrob. Agents Chemother. 2004, 48, 2069–2074. [Google Scholar] [CrossRef]
- Zuhuang, M.; Qin, L. Genes of 16S rRNA Methylase and Aminoglycoside-modifying Enzymes in Pan-drug Resistant Pseudomonas aeruginosa. Chin. J. Nosocomiol. 2006; in press. [Google Scholar]
- Cho, Y.J.; Moon, D.C.; Jin, J.S.; Choi, C.H.; Lee, Y.C.; Lee, J.C. Genetic basis of resistance to aminoglycosides in Acinetobacter spp. and spread of armA in Acinetobacter baumannii sequence group 1 in Korean hospitals. Diagn. Microbiol. Infect. Dis. 2009, 64, 185–190. [Google Scholar] [CrossRef]
- El-Badawy, M.F.; El-Far, S.W.; Althobaiti, S.S.; Abou-Elazm, F.I.A.; Shohayeb, M.M. The First Egyptian Report Showing the Co-Existence of blaNDM-25, blaOXA-23, blaOXA-181, and blaGES-1 Among Carbapenem-Resistant K. pneumoniae Clinical Isolates Genotyped by BOX-PCR. Infect. Drug Resist. 2020, 13, 1237–1250. [Google Scholar] [CrossRef] [PubMed]
- Wayne, P. CLSI Performance Standards for Antimicrobial Susceptibility Testing; CLSI Document Clinical Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2017. [Google Scholar]
- El-Badawy, M.F.; Alrobaian, M.M.; Shohayeb, M.M.; Abdelwahab, S.F. Investigation of six plasmid-mediated quinolone resistance genes among clinical isolates of pseudomonas: A genotypic study in Saudi Arabia. Infect. Drug Resist. 2019, 12, 915–923. [Google Scholar] [CrossRef] [PubMed]
- El-Badawy, M.F.; Tawakol, W.M.; El-Far, S.W.; Maghrabi, I.A.; Al-Ghamdi, S.A.; Mansy, M.S.; Ashour, M.S.; Shohayeb, M.M. Molecular Identification of Aminoglycoside-Modifying Enzymes and Plasmid-Mediated Quinolone Resistance Genes among Klebsiella pneumoniae Clinical Isolates Recovered from Egyptian Patients. Int. J. Microbiol. 2017, 2017, 8050432. [Google Scholar] [CrossRef] [PubMed]
- Bradford, P.A.; Sanders, C.C. Use of a predictor panel to evaluate susceptibility test methods proposed for piperacillin-tazobactam. Antimicrob. Agents Chemother. 1993, 37, 2578–2583. [Google Scholar] [CrossRef]
- Mayrhofer, S.; Domig, K.J.; Mair, C.; Zitz, U.; Huys, G.; Kneifel, W.J. Comparison of broth microdilution, Etest, and agar disk diffusion methods for antimicrobial susceptibility testing of Lactobacillus acidophilus group members. Appl. Environ. Microbiol. 2008, 74, 3745–3748. [Google Scholar] [CrossRef] [PubMed]
- Greco, R.; Panetta, V.; Della Rocca, M.T.; Durante, A.; Di Caprio, G.; Maggi, P. Profile of Co-Infection Prevalence and Antibiotics Use among COVID-19 Patients. Pathogens 2022, 11, 1250. [Google Scholar] [CrossRef]
- Cayci, Y.T.; Ulker, K.H.; Birinci, A. Evaluation of three different methods for susceptibility testing of gentamicin in carbapenem-resistant Enterobacterales. Infez. Med. 2021, 29, 568–573. [Google Scholar] [CrossRef]
- Batool, S.; Almaghaslah, D.; Alqahtani, A.; Almanasef, M.; Alasmari, M.; Vasudevan, R.; Attique, S.; Riaz, F. Aetiology and antimicrobial susceptibility pattern of bacterial isolates in community acquired pneumonia patients at Asir region, Saudi Arabia. Int. J. Clin. Pract. 2020, 75, e13667. [Google Scholar] [CrossRef]
- Yezli, S.; Shibl, A.M.; Livermore, D.M.; Memish, Z.A. Prevalence and antimicrobial resistance among Gram-negative pathogens in Saudi Arabia. J. Chemother. 2014, 26, 257–272. [Google Scholar] [CrossRef]
- O’Donnell, J.N.; Bidell, M.R.; Lodise, T.P. Approach to the Treatment of Patients with Serious Multidrug-Resistant Pseudomonas aeruginosa Infections. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2020, 40, 952–969. [Google Scholar] [CrossRef]
- Ahmed, M.A.S.; Khan, F.A.; Sultan, A.A.; Söderquist, B.; Ibrahim, E.B.; Jass, J.; Omrani, A.S. β-lactamase-mediated resistance in MDR-Pseudomonas aeruginosa from Qatar. Antimicrob. Resist. Infect. Control. 2020, 9, 170. [Google Scholar] [CrossRef]
- Helmy, O.M.; Kashef, M.T. Different phenotypic and molecular mechanisms associated with multidrug resistance in Gram-negative clinical isolates from Egypt. Infect. Drug Resist. 2017, 10, 479–498. [Google Scholar] [CrossRef]
- Lila, G.; Mulliqi-Osmani, G.; Bajrami, R.; Kurti, A.; Azizi, E.; Raka, L. The prevalence and resistance patterns of Pseudomonas aeruginosa in a tertiary care hospital in Kosovo. Infez. Med. 2017, 25, 21–26. [Google Scholar]
- Al-Tawfiq, J.A. Occurrence and antimicrobial resistance pattern of inpatient and outpatient isolates of Pseudomonas aeruginosa in a Saudi Arabian hospital: 1998–2003. Int. J. Infect. Dis. 2007, 11, 109–114. [Google Scholar] [CrossRef]
- Poole, K. Aminoglycoside Resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2005, 49, 479–487. [Google Scholar] [CrossRef]
- Panahi, T.; Asadpour, L.; Ranji, N. Distribution of aminoglycoside resistance genes in clinical isolates of Pseudomonas aeruginosa in north of Iran. Gene Rep. 2020, 21, 100929. [Google Scholar] [CrossRef]
- Torres, C.; Perlin, M.H.; Baquero, F.; Lerner, D.L.; Lerner, S.A. High-level amikacin resistance in Pseudomonas aeruginosa associated with a 3′-phosphotransferase with high affinity for amikacin. Int. J. Antimicrob. Agents 2000, 15, 257–263. [Google Scholar] [CrossRef]
- Teixeira, B.; Rodulfo, H.; Carreño, N.; Guzmán, M.; Salazar, E.; De Donato, M. Aminoglycoside resistance genes in Pseudomonas aeruginosa isolates from Cumana. Rev. Inst. Med. Trop. São Paulo 2016, 58, 13. [Google Scholar] [CrossRef]
- Vaziri, F.; Peerayeh, S.N.; Nejad, Q.B.; Farhadian, A. The prevalence of aminoglycoside-modifying enzyme genes (aac(6′)-I, aac(6′)-II, ant(2″)-I, aph(3′)-VI) in Pseudomonas aeruginosa. Clinics 2011, 66, 1519–1522. [Google Scholar]
- Miller, G.H.; Sabatelli, F.J.; Hare, R.S.; Glupczynski, Y.; Mackey, P.; Shlaes, D.; Shimizu, K.; Shaw, K.J.; Aminoglycoside Resistance Study Groups. The most frequent aminoglycoside resistance mechanisms—Changes with time and geographic area: A reflection of aminoglycoside usage patterns? Clin. Infect. Dis. 1997, 24, S46–S62. [Google Scholar] [CrossRef]
- Poonsuk, K.; Tribuddharat, C.; Chuanchuen, R. Aminoglycoside resistance mechanisms in Pseudomonas aeruginosa isolates from non-cystic fibrosis patients in Thailand. Can. J. Microbiol. 2013, 59, 51–56. [Google Scholar] [CrossRef]
- Thacharodi, A.; Lamont, I.L. Aminoglycoside resistance in Pseudomonas aeruginosa: The contribution of the MexXY-OprM efflux pump varies between isolates. J. Med. Microbiol. 2022, 71, 001551. [Google Scholar] [CrossRef]
- Meoli, A.; Ciavola, L.; Rahman, S.; Masetti, M.; Toschetti, T.; Morini, R.; Canto, G.D.; Auriti, C.; Caminiti, C.; Castagnola, E.; et al. Prevention of Surgical Site Infections in Neonates and Children: Non-Pharmacological Measures of Prevention. Antibiotics 2022, 11, 863. [Google Scholar] [CrossRef] [PubMed]
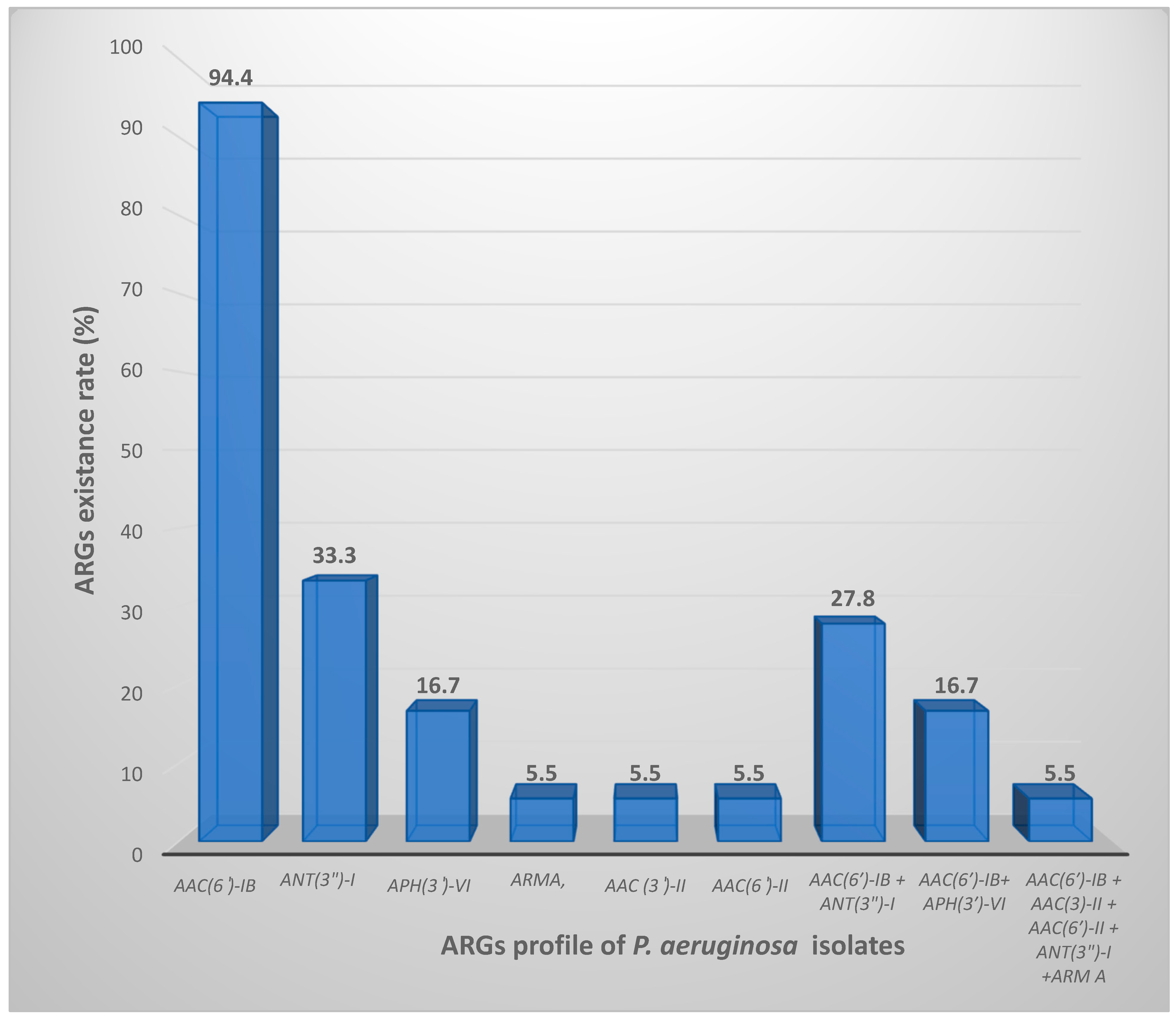
| Antibiotic/Method | Disk Diffusion No. (%) of Isolates | Broth Microdilution MIC (µg/mL) | Resistant Rate (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Resistant (R) | Intermediate (I) | Susceptible (S) | 1024–256 | 128 | 64 | 32 | 16 | 8 | 4 | 2 | 1 | 0.5 | ||
| Gentamicin (GM) | 29 | - | 71 | 4 | - | 3 | 4 | 18 | 2 | 7 | 6 | 6 | 50 | 31% |
| Amikacin (AK) | 18 | 6 | 76 | 3 | 5 | 10 | 5 | 29 | 1 | 3 | 6 | - | 38 | 18% |
| Tobramycin (TM) | 18 | 3 | 79 | 1 | 11 | 6 | 3 | 10 | 5 | 15 | 9 | 29 | 11 | 18% |
| Ceftazidime (CTZ) | 9 | 10 | 81 | 2 | 2 | 1 | 5 | 15 | 26 | 6 | 7 | 14 | 22 | 10% |
| Imipenem (IMP) | 29 | - | 71 | 2 | 1 | 1 | 7 | 21 | 7 | 28 | 7 | 6 | 20 | 32% |
| Piperacillin/Tazobactam * (TZP) | 13 | - | 87 | 6 | 7 | 5 | 6 | 6 | 9 | 10 | 10 | 21 | 20 | 13% |
| Ciprofloxacin (CIP) | 27 | - | 73 | - | - | - | 1 | 2 | 6 | 20 | 1 | 18 | 52 | 29% |
| Isolate | aph(3)-VI (780 bp) | Arm A (846 bp) | aac(6*)-Ib (472 bp) | aac(6*)-II (542 bp) | aac(3*)-II (877 bp) | rmtB (769 bp) | ant(3**)-I (787 bp) |
|---|---|---|---|---|---|---|---|
| # 1 | + | - | + | - | - | - | - |
| # 2 | - | - | + | - | - | - | - |
| # 3 | - | - | + | - | - | - | - |
| # 4 | - | - | - | - | - | - | - |
| # 5 | - | - | + | - | - | - | - |
| # 6 | - | - | + | - | - | - | + |
| # 7 | - | - | + | - | - | - | - |
| # 8 | + | - | + | - | - | - | - |
| # 10 | - | - | + | - | - | - | + |
| # 11 | - | - | + | - | - | - | - |
| # 13 | + | - | + | - | - | - | - |
| # 14 | - | - | + | - | - | - | - |
| # 16 | - | - | + | - | - | - | + |
| # 17 | - | + | + | + | + | - | + |
| # 19 | - | - | + | - | - | - | + |
| # 20 | - | - | + | - | - | - | + |
| # 22 | - | - | + | - | - | - | - |
| # 23 | - | - | + | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Far, S.W.; Abukhatwah, M.W. Prevalence of Aminoglycoside Resistance Genes in Clinical Isolates of Pseudomonas aeruginosa from Taif, Saudi Arabia—An Emergence Indicative Study. Microorganisms 2023, 11, 2293. https://doi.org/10.3390/microorganisms11092293
El-Far SW, Abukhatwah MW. Prevalence of Aminoglycoside Resistance Genes in Clinical Isolates of Pseudomonas aeruginosa from Taif, Saudi Arabia—An Emergence Indicative Study. Microorganisms. 2023; 11(9):2293. https://doi.org/10.3390/microorganisms11092293
Chicago/Turabian StyleEl-Far, Shaymaa W., and Mohamed W. Abukhatwah. 2023. "Prevalence of Aminoglycoside Resistance Genes in Clinical Isolates of Pseudomonas aeruginosa from Taif, Saudi Arabia—An Emergence Indicative Study" Microorganisms 11, no. 9: 2293. https://doi.org/10.3390/microorganisms11092293
APA StyleEl-Far, S. W., & Abukhatwah, M. W. (2023). Prevalence of Aminoglycoside Resistance Genes in Clinical Isolates of Pseudomonas aeruginosa from Taif, Saudi Arabia—An Emergence Indicative Study. Microorganisms, 11(9), 2293. https://doi.org/10.3390/microorganisms11092293