

Transcriptomic Analysis of the Response of the Toxic Dinoflagellate Prorocentrum lima to Phosphorous Limitation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Culture
2.2. Toxin Analysis
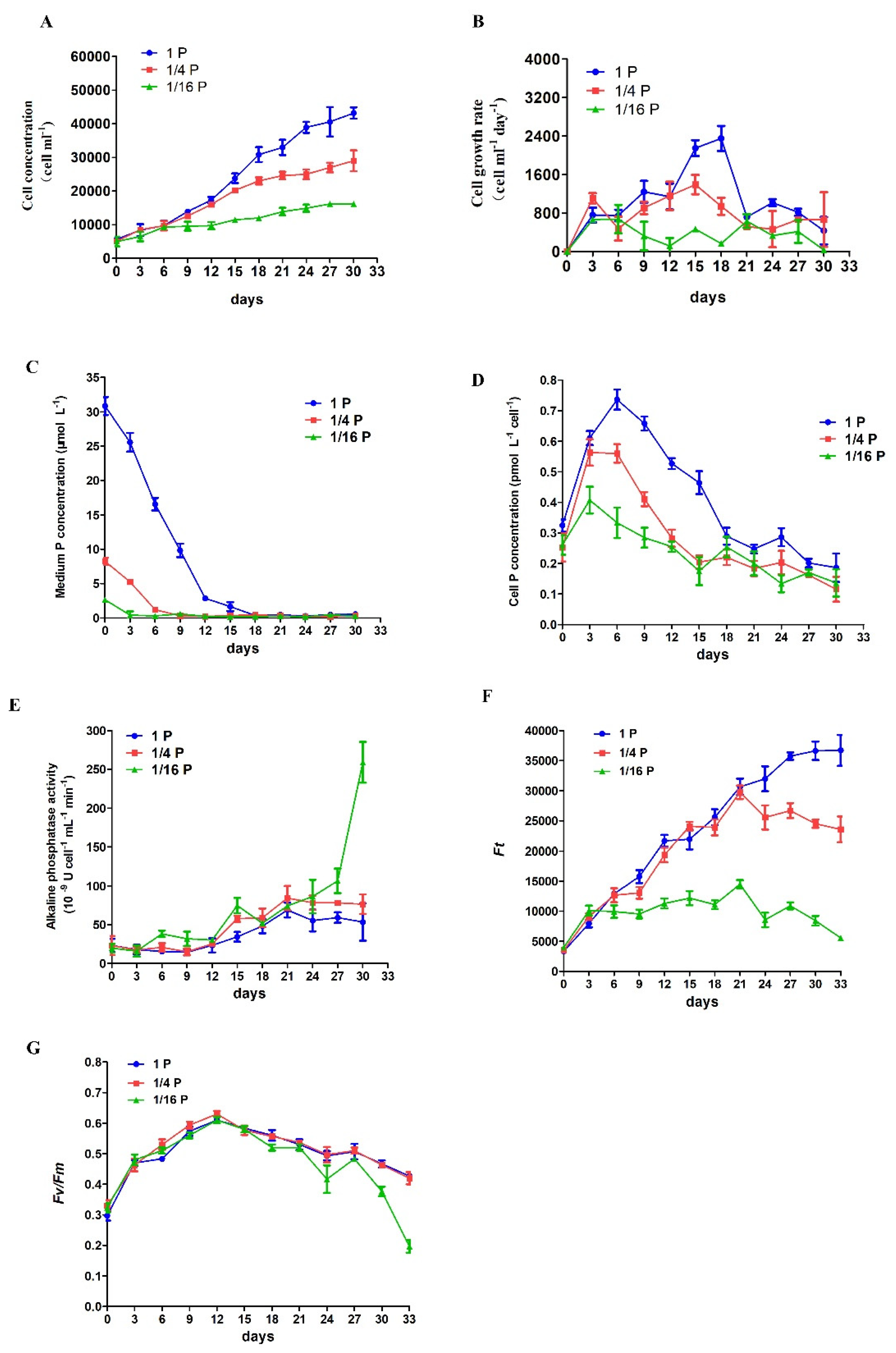
2.3. Analysis of Physiological Parameters
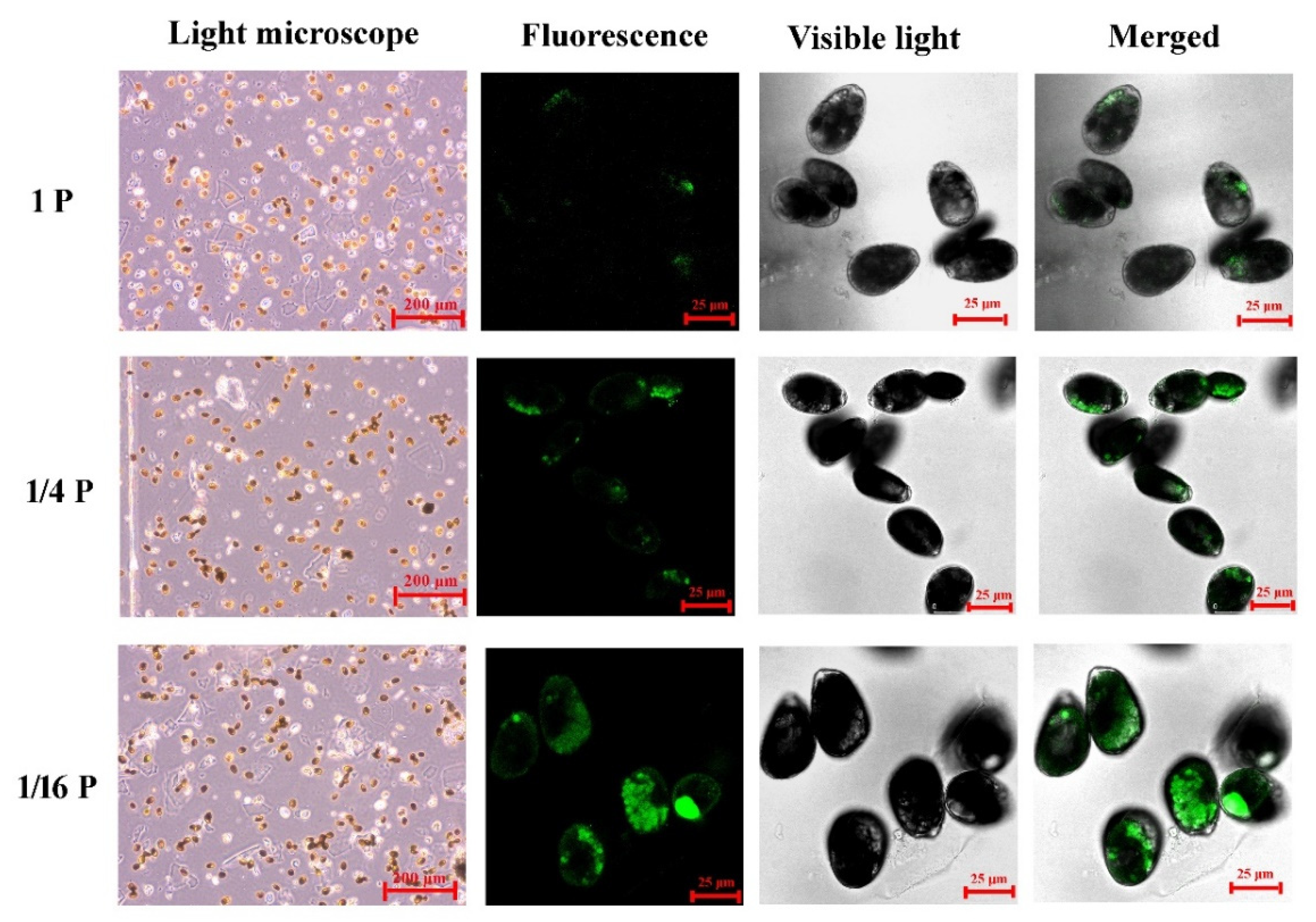
2.4. Confocal Laser Scanning Microscope (CLSM) Analysis
2.5. Lipid Analysis
2.6. Starch Content Measurements
2.7. RNA Extraction, Sequencing, and Transcriptome Analysis
2.8. RT-qPCR
2.9. Statistical Analysis
3. Results
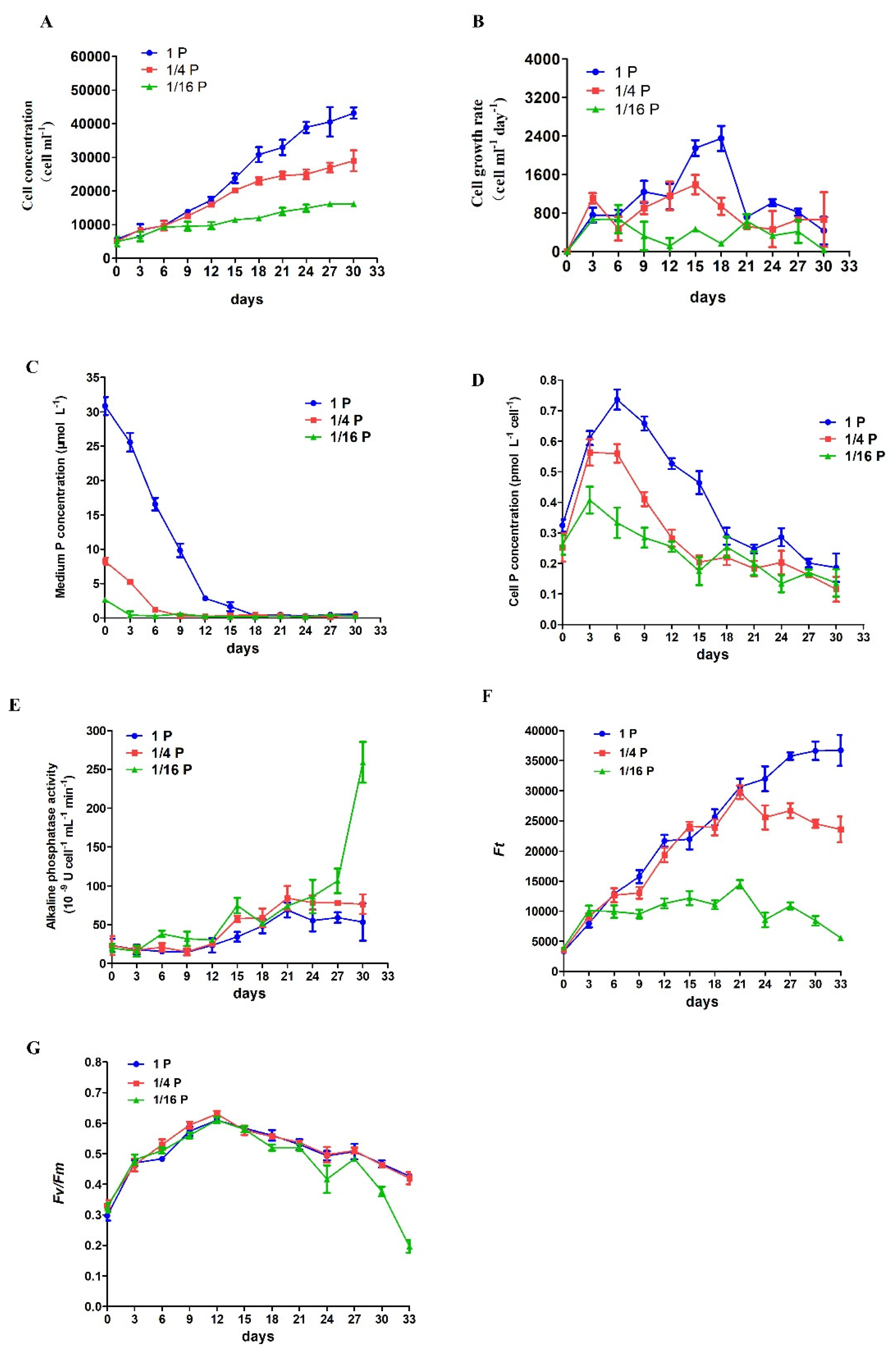
3.1. Physiological Responses of P. lima under Phosphate Limitation
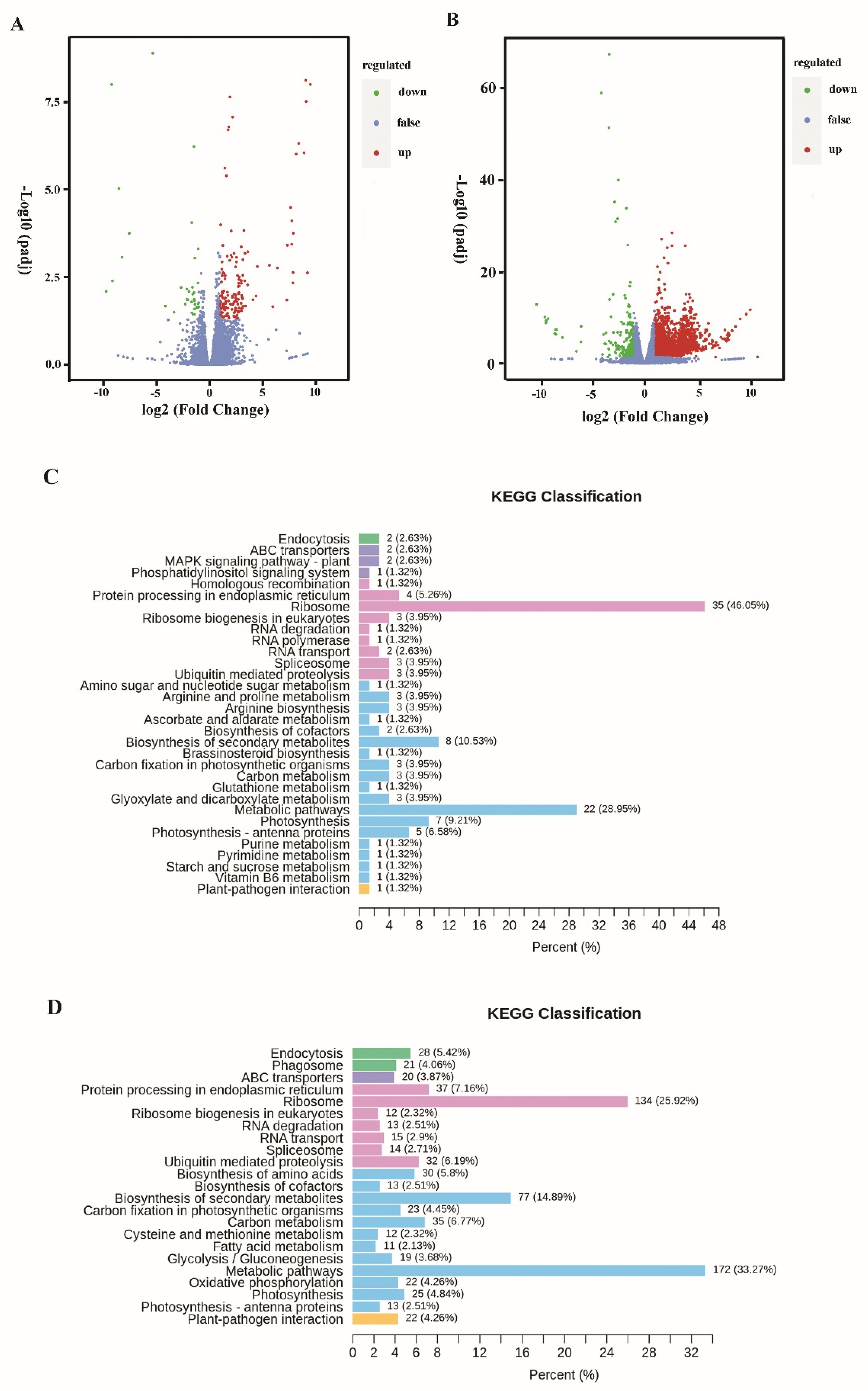
3.2. Transcriptomic Analysis
3.3. Transcriptomic Analysis of Differential Gene Expression
3.4. RT-qPCR Validation of Differentially Expressed Genes
3.5. Lipid and Starch Accumulation under P-Limited Conditions
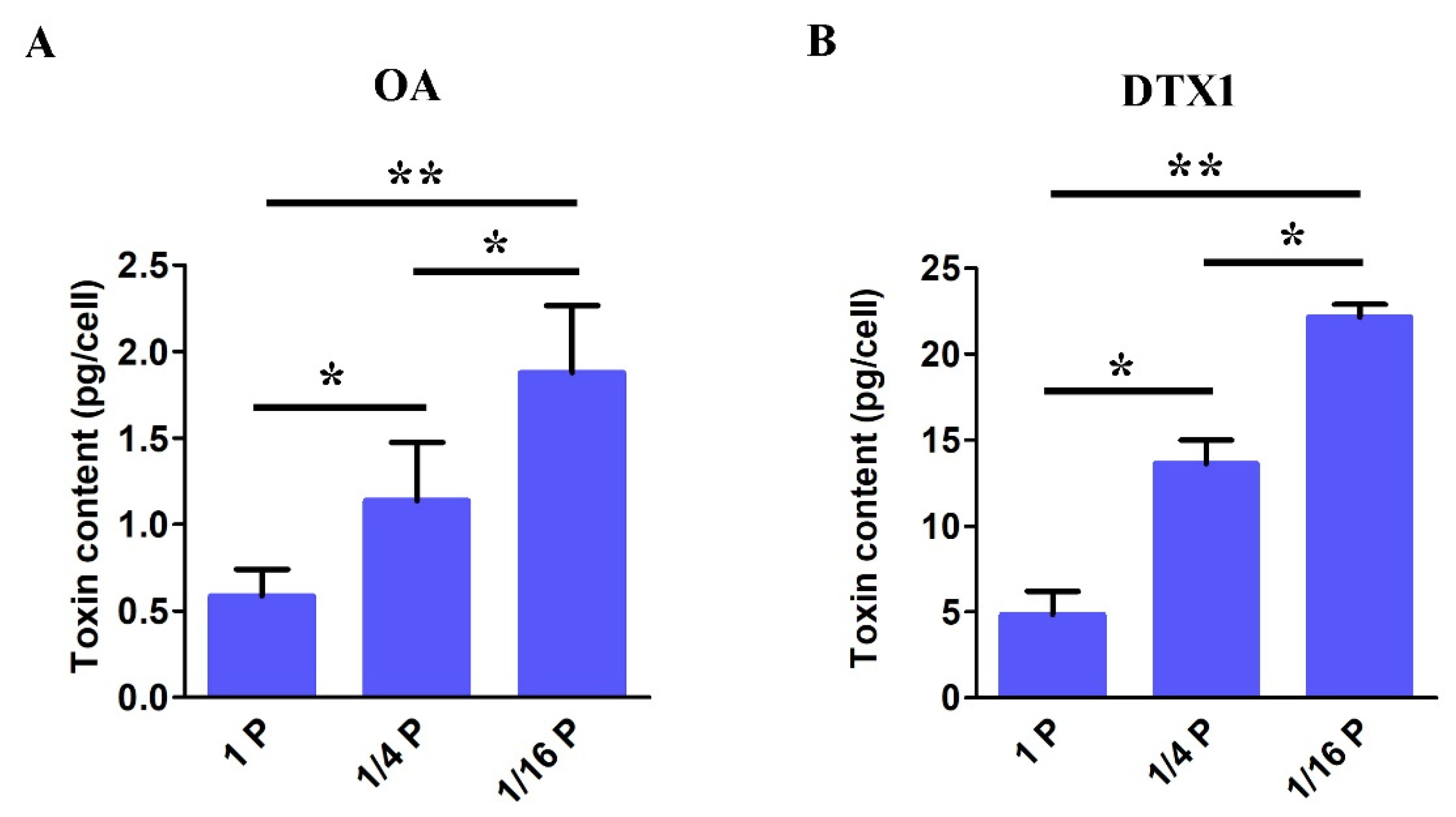
3.6. DSP Toxin Content in P. lima under P-Limited Conditions
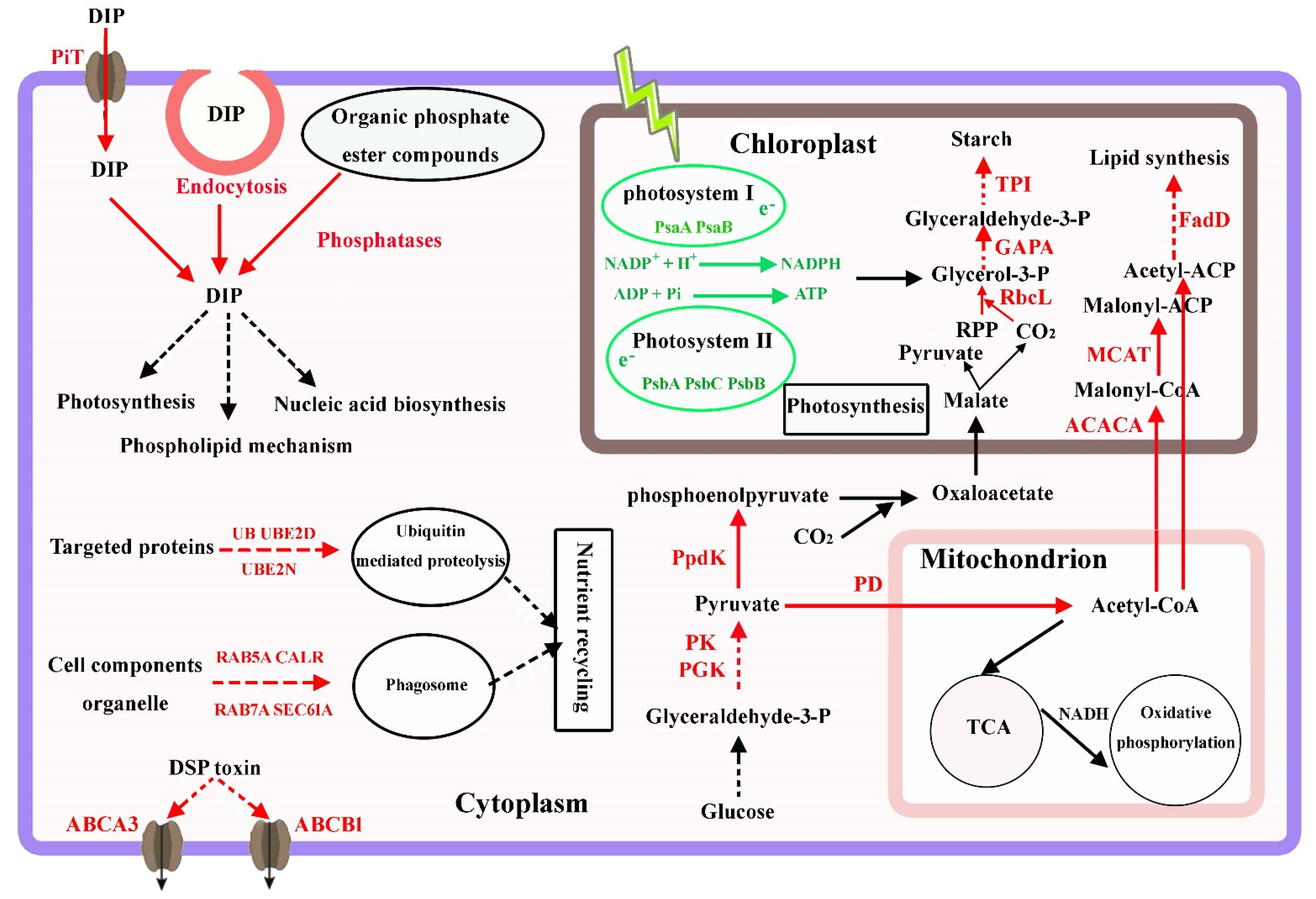
4. Discussion
4.1. Carbon Metabolism and Photosynthesis under P-Limited Conditions
4.2. Phosphate Transport, Utilization, and Homeostasis
4.3. Lipid and Starch Accumulation under P-Limited Conditions
4.4. DSP Toxins Production in P. lima
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Morse, D.; Tse, S.P.K.; Lo, S.C.L. Exploring dinoflagellate biology with high-throughput proteomics. Harmful Algae 2018, 75, 16–26. [Google Scholar] [CrossRef]
- Wan, X.; Yao, G.; Wang, K.; Bao, S.; Han, P.; Wang, F.; Song, T.; Jiang, H. Transcriptomic analysis of polyketide synthesis in dinoflagellate, Prorocentrum lima. Harmful Algae 2023, 123, 102391. [Google Scholar] [CrossRef]
- Gong, W.; Browne, J.; Hall, N.; Schruth, D.; Paerl, H.; Marchetti, A. Molecular insights into a dinoflagellate bloom. ISME J. 2017, 11, 439–452. [Google Scholar] [CrossRef]
- Zhang, S.F.; Yuan, C.J.; Chen, Y.; Lin, L.; Wang, D.Z. Transcriptomic response to changing ambient phosphorus in the marine dinoflagellate Prorocentrum donghaiense. Sci. Total Environ. 2019, 692, 1037–1047. [Google Scholar] [CrossRef]
- Bravo, I.; Fern’andez, M.L.; Ramilo, I.; Martınez, A. Toxin composition of the toxic dinoflagellate Prorocentrum lima isolated from different locations along the Galician coast (NW Spain). Toxicon 2001, 39, 1537–1545. [Google Scholar] [CrossRef]
- Nascimento, S.M.; Purdie, D.A.; Morris, S. Morphology, toxin composition and pigment content of Prorocentrum lima strains isolated from a coastal lagoon in southern UK. Toxicon 2005, 45, 633–649. [Google Scholar] [CrossRef]
- Oyeku, O.G.; Mandal, S.K. Taxonomy and okadaic acid production of a strain of Prorocentrum lima (Dinophyceae) isolated from the Bay of Bengal, North Indian Ocean. Toxicon 2021, 196, 32–43. [Google Scholar] [CrossRef]
- Koike, K.; Sato, S.; Yamaji, M.; Nagahama, Y.; Kotaki, Y.; Ogata, T.; Kodama, M. Occurrence of okadaic acid-producing Prorocentrum lima on the Sanriku coast, northern Japan. Toxicon 1998, 36, 2039–2042. [Google Scholar] [CrossRef]
- Zhang, H.; Li, Y.; Cen, J.; Wang, H.; Cui, L.; Dong, Y.; Lu, S. Morphotypes of Prorocentrum lima (Dinophyceae) from Hainan Island, South China Sea: Morphological and molecular characterization. Phycologia 2015, 54, 503–516. [Google Scholar] [CrossRef]
- Lawrence, J.E.; Cembella, A.D.; Ross, N.W.; Wright, J.L. Cross-reactivity of an anti-okadaic acid antibody to dinophysistoxin-4 (DTX-4), dinophysistoxin-5 (DTX-5), and an okadaic acid diol ester. Toxicon 1998, 36, 1193–1196. [Google Scholar] [CrossRef]
- Gayoso, A.N.; Dover, S.; Morton, S.; Busman, M.; Moeller, P.; Fulco, V.K.; Maranda, L. Diarrhetic shellfish poisoning associated with Prorocentrum lima (dinophyceae) in Patagonian Gulfs (Argentina). J. Shellfish Res. 2002, 21, 461–463. [Google Scholar]
- Ingarao, C.; Lanciani, G.; Verri, C.; Pagliani, T. First record of Prorocentrum lima (Dinophyceae) inside harbor areas and along the Abruzzo region coast, W Adriatic. Mar. Pollut. Bull. 2009, 58, 596–600. [Google Scholar] [CrossRef] [PubMed]
- Turkoglu, M. First harmful algal bloom record of tycoplanktonic dinoflagellate Prorocentrum lima (Ehrenberg) F. Stein, 1878 in the Dardanelles (Turkish Straits System, Turkey). J. Coast. Life Med. 2016, 4, 765–774. [Google Scholar]
- D’Agostino, V.C.; Krock, B.; Degrati, M.; Sastre, V.; Santinelli, N.; Krohn, T.; Hoffmeyer, M.S. Occurrence of toxigenic microalgal species and phycotoxin accumulation in mesozooplankton in Northern Patagonian Gulfs, Argentina. Environ. Toxicol. Chem. 2019, 38, 2209–2223. [Google Scholar] [CrossRef]
- Gobler, C.J.; Doherty, O.M.; Hattenrath-Lehmann, T.K.; Griffith, A.W.; Kang, Y.; Litaker, R.W. Ocean warming since 1982 has expanded the niche of toxic algal blooms in the North Atlantic and North Pacific oceans. Proc. Natl. Acad. Sci. USA 2017, 114, 4975–4980. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Liu, X.; Irwin, A.J.; Laws, E.A.; Wang, L.; Chen, B.; Zeng, Y.; Huang, B. Warming and eutrophication combine to restructure diatoms and dinoflagellates. Water Res. 2018, 128, 206–216. [Google Scholar] [CrossRef]
- Varkitzi, I.; Pagou, K.; Graneli, E.; Hatzianestis, I.; Pyrgaki, C.; Pavlidou, A.; Montesanto, B.; Economou-Amilli, A. Unbalanced N: P ratios and nutrient stress controlling growth and toxin production of the harmful dinoflagellate Prorocentrum lima (Ehrenberg) Dodge. Harmful Algae 2010, 9, 304–311. [Google Scholar] [CrossRef]
- Lai, J.; Yu, Z.; Song, X.; Cao, X.; Han, X. Responses of the growth and biochemical composition of Prorocentrum donghaiense to different nitrogen and phosphorus concentrations. J. Exp. Mar. Biol. Ecol. 2011, 405, 6–17. [Google Scholar] [CrossRef]
- Hou, D.Y.; Mao, X.T.; Gu, S.; Li, H.Y.; Liu, J.S.; Yang, W.D. Systems-level analysis of metabolic mechanism following nitrogen limitation in benthic dinoflagellate Prorocentrum lima. Algal Res. 2018, 33, 389–398. [Google Scholar] [CrossRef]
- Cui, G.; Liew, Y.J.; Konciute, M.K.; Zhan, Y.; Hung, S.H.; Thistle, J.; Gastoldi, L.; Schmidt-Roach, S.; Dekker, J.; Aranda, M. Nutritional control regulates symbiont proliferation and life history in coral-dinoflagellate symbiosis. BMC Biol. 2022, 20, 103. [Google Scholar] [CrossRef]
- Vanucci, S.; Guerrini, F.; Milandri, A.; Pistocchi, R. Effects of different levels of N and P-deficiency on cell yield, okadaic acid, DTX-1, protein and carbohydrate dynamics in the benthic dinoflagellate Prorocentrum lima. Harmful Algae 2010, 9, 590–599. [Google Scholar] [CrossRef]
- Hou, D.Y.; Liang, J.J.; Zou, C.; Li, H.Y.; Liu, J.S.; Yang, W.D. MRP functional activity and character in the dinoflagellate Prorocentrum lima. J. Appl. Phycol. 2016, 28, 1667–1676. [Google Scholar] [CrossRef]
- Wang, D.Z.; Zhang, H.; Zhang, Y.; Zhang, S.F. Marine dinoflagellate proteomics: Current status and future perspectives. J. Proteom. 2014, 105, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Dyhrman, S.T. Nutrients and their acquisition: Phosphorus physiology in microalgae. Physiol. Microalgae 2016, 22, 155–183. [Google Scholar]
- Lin, S.; Litaker, R.W.; Sunda, W.G. Phosphorus physiological ecology and molecular mechanisms in marine phytoplankton. J Phycol. 2016, 52, 10–36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, G.; Wang, Y.; Guo, C.; Zhou, J. Physiological and molecular responses of Prorocentrum donghaiense to dissolved inorganic phosphorus limitation. Mar. Pollut. Bull. 2018, 129, 562–572. [Google Scholar] [CrossRef]
- Zhang, S.F.; Yuan, C.J.; Chen, Y.; Chen, X.H.; Li, D.X.; Liu, J.L.; Lin, L.; Wang, D.Z. Comparative transcriptomic analysis reveals novel insights into the adaptive response of Skeletonema costatum to changing ambient phosphorus. Front. Microbiol. 2016, 7, 1476. [Google Scholar] [CrossRef]
- Luo, H.; Lin, X.; Li, L.; Zhang, C.; Lin, S. Transcriptomic and physiological analyses of the dinoflagellate Karenia mikimotoi reveal non-alkaline phosphatase-based molecular machinery of ATP utilisation. Environ. Microbiol. 2017, 19, 4506–4518. [Google Scholar] [CrossRef]
- Dyhrman, S.T.; Jenkins, B.D.; Rynearson, T.A.; Saito, M.A.; Mercier, M.L.; Alexander, H.; Whitney, L.P.; Drzewianowski, A.; Bulygin, V.V.; Bertrand, E.M.; et al. The transcriptome and proteome of the diatom Thalassiosira pseudonana reveal a diverse phosphorus stress response. PLoS ONE 2012, 7, e33768. [Google Scholar] [CrossRef]
- Orchard, E.D.; Benitez-Nelson, C.R.; Pellechia, P.J.; Lomas, M.W.; Dyhrman, S.T. Polyphosphate in Trichodesmium from the low-phosphorus Sargasso Sea. Limnol. Oceanogr. 2010, 55, 2161–2169. [Google Scholar] [CrossRef]
- Diaz, J.; Ingall, E.; Benitez-Nelson, C.; Paterson, D.; de Jonge, M.D.; McNulty, I.; Brandes, J.A. Marine polyphosphate: A key player in geologic phosphorus sequestration. Science 2008, 320, 652–655. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Lin, X.; Li, L.; Li, M.; Palenik, B.; Lin, S. Transcriptomic and microRNAomic profiling reveals multi-faceted mechanisms to cope with phosphate stress in a dinoflagellate. ISME J. 2017, 11, 2209–2218. [Google Scholar] [PubMed]
- Jauzein, C.; Labry, C.; Youenou, A.; Quéré, J.; Delmas, D.; Collos, Y. Growth and phosphorus uptake by the toxic dinoflagellate Alexandrium catenella (dinophyceae) in response to phosphate limitation. J. Phycol. 2010, 46, 926–936. [Google Scholar] [CrossRef]
- Martin, P.; Van Mooy, B.A.S.; Heithoff, A.; Dyhrman, S.T. Phosphorus supply drives rapid turnover of membrane phospholipids in the diatom Thalassiosira pseudonana. ISME J. 2011, 5, 1057–1060. [Google Scholar] [CrossRef]
- Van Mooy, B.A.; Fredricks, H.F.; Pedler, B.E.; Dyhrman, S.T.; Karl, D.M.; Koblízek, M.; Lomas, M.W.; Mincer, T.J.; Moore, L.R.; Moutin, T.; et al. Phytoplankton in the ocean use non-phosphorus lipids in response to phosphorus scarcity. Nature 2009, 458, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.F.; Chen, Y.; Xie, Z.X.; Zhang, H.; Lin, L.; Wang, D.Z. Unraveling the molecular mechanism of the response to changing ambient phosphorus in the dinoflagellate Alexandrium catenella with quantitative proteomics. J. Proteom. 2019, 196, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Ding, N.; Huertas, R.; Torres-Jerez, I.; Liu, W.; Watson, B.; Scheible, W.R.; Udvardi, M. Transcriptional, metabolic, physiological and developmental responses of switchgrass to phosphorus limitation. Plant Cell Environ. 2021, 44, 186–202. [Google Scholar] [CrossRef]
- Chen, Z.; Song, J.; Li, X.; Arango, J.; Cardoso, J.A.; Rao, I.; Schultze-Kraft, R.; Peters, M.; Mo, X.; Liu, G. Physiological responses and transcriptomic changes reveal the mechanisms underlying adaptation of Stylosanthes guianensis to phosphorus deficiency. BMC Plant Biol. 2021, 21, 466. [Google Scholar] [CrossRef]
- Wu, Q.; Yang, L.; Liang, H.; Yin, L.; Chen, D.; Shen, P. Integrated analyses reveal the response of peanut to phosphorus deficiency on phenotype, transcriptome and metabolome. BMC Plant Biol. 2022, 22, 524. [Google Scholar] [CrossRef]
- Nagahama, Y.; Murray, S.; Tomaru, A.; Fukuyo, Y. Species boundaries in the toxic dinoflagellate Prorocentrum lima (Dinophyceae, Prorocentrales), based on morphological and phylogenetic characters. J. Phycol. 2011, 47, 178–189. [Google Scholar] [CrossRef]
- Ben-Gharbia, H.; Yahia, O.K.; Amzil, Z.; Chom’erat, N.; Abadie, E.; Masseret, E.; Sibat, M.; Zmerli Triki, H.; Nouri, H.; Laabir, M. Toxicity and growth assessments of three thermophilic benthic dinoflagellates (Ostreopsis cf. ovata, Prorocentrum lima and Coolia monotis) developing in the southern mediterranean basin. Toxins 2016, 8, 297. [Google Scholar] [CrossRef]
- Cembella, A.D.; Dur’an-Riveroll, L.M.; Tarazona-Janampa, U.I.; Okolodkov, Y.B.; García-Sandoval, R.; Krock, B.; Hörstmann, C.; John, U. Phylogeography and diversity among populations of the toxigenic benthic dinoflagellate Prorocentrum from coastal reef systems in Mexico. Front. Mar. Sci. 2021, 8, 716669. [Google Scholar] [CrossRef]
- From, N.; Richardson, K.; Mousing, E.A.; Jensen, P.E. Removing the light history signal from normalized variable fluorescence (Fv/Fm) measurements on marine phytoplankton. Limnol. Oceanogr. Meth. 2014, 12, 776–783. [Google Scholar] [CrossRef]
- Gajic, G.; Mitrovic, M.; Pavlovic, P.; Stevanovic, B.; Djurdjevic, L.; Kostic, O. An assessment of the tolerance of Ligustrum ovalifolium Hassk. to traffic-generated Pb using physiological and biochemical markers. Ecotox. Environ. Saf. 2009, 72, 1090–1101. [Google Scholar] [CrossRef]
- Roberts, K.; Granum, E.; Leegood, R.C.; Raven, J.A. Carbon acquisition by diatoms. Photosynth. Res. 2007, 93, 79–88. [Google Scholar]
- Moseley, J.L.; Chang, C.W.; Grossman, A.R. Genome-based approaches to understanding phosphorus deprivation responses and PSR1 control in Chlamydomonas reinhardtii. Eukaryot. Cell 2006, 5, 26–44. [Google Scholar] [CrossRef] [PubMed]
- Mo, X.; Liu, G.; Zhang, Z.; Lu, X.; Liang, C.; Tian, J. Mechanisms Underlying Soybean Response to Phosphorus Deficiency through Integration of Omics Analysis. Int. J. Mol. Sci. 2022, 23, 4592. [Google Scholar] [CrossRef]
- Jonsson, C.M.; Paraiba, L.C.; Aoyama, H. Metals and linear alkylbenzene sulphonate as inhibitors of the algae Pseudokirchneriella subcapitata acid phosphatase activity. Ecotoxicology 2009, 18, 610–619. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, J.; Shi, L.; Wang, Q.; Zhang, P.; Wang, H.; Liu, J.; Li, H.; Li, L.; Li, X.; et al. Identification of core genes associated with different phosphorus levels in quinoa seedlings by weighted gene co-expression network analysis. BMC Genom. 2023, 24, 399. [Google Scholar] [CrossRef]
- Deng, S.; Li, J.; Du, Z.; Wu, Z.; Yang, J.; Cai, H.; Wu, G.; Xu, F.; Huang, Y.; Wang, S.; et al. Rice ACID PHOSPHATASE 1 regulates Pi stress adaptation by maintaining intracellular Pi homeostasis. Plant Cell Environ. 2022, 45, 191–205. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, H.; Chu, S.; Li, H.; Chi, Y.; Triebwasser-Freese, D.; Lv, H.; Yu, D. Integrating QTL mapping and transcriptomics identifies candidate genes underlying QTLs associated with soybean tolerance to low-phosphorus stress. Plant Mol. Biol. 2017, 93, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Richmond, G.S.; Smith, T.K. Phospholipases A1. Int. J. Mol. Sci. 2011, 12, 588–612. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J. Phospholipase D and phosphatidic acid in plant defence response: From protein-protein and lipid-protein interactions to hormone signalling. J. Exp. Bot. 2015, 66, 1721–1736. [Google Scholar] [CrossRef] [PubMed]
- Pleskot, R.; Li, J.; Žárský, V.; Potocký, M.; Staiger, C.J. Regulation of cytoskeletal dynamics by phospholipase D and phosphatidic acid. Trends Plant Sci. 2013, 18, 496–504. [Google Scholar] [CrossRef]
- Frohman, M.A. The phospholipase D super family as therapeutic targets. Trends Pharmacol. Sci. 2015, 36, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Collinet, C.; Stöter, M.; Bradshaw, C.R.; Samusik, N.; Rink, J.C.; Kenski, D.; Habermann, B.; Buchholz, F.; Henschel, R.; Mueller, M.S.; et al. Systems survey of endocytosis by multiparametric image analysis. Nature 2010, 464, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Li, R.; Pan, J.; Ding, Z.; Lin, J. Endocytosis and its regulation in plants. Trends Plant Sci. 2015, 20, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Naramoto, S.; Kleine-Vehn, J.; Robert, S.; Fujimoto, M.; Dainobu, T.; Paciorek, T.; Ueda, T.; Nakano, A.; Van Montagu, M.C.; Fukuda, H.; et al. ADP-ribosylation factor machinery mediates endocytosis in plant cells. Proc. Natl. Acad. Sci. USA 2010, 107, 21890–21895. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.X.; Kozmikova, I.; Ono, H.; Nossa, C.W.; Kozmik, Z.; Putnam, N.H.; Yu, J.K.; Holland, L.Z. Conserved noncoding elements in the most distant genera of cephalochordates: The goldilocks principle. Genome Biol. Evol. 2016, 8, 2387–2405. [Google Scholar] [CrossRef]
- Urwin, H.; Ghazi-Noori, S.; Collinge, J.; Isaacs, A. The role of CHMP2B in frontotemporal dementia. Biochem. Soc. Trans. 2009, 37, 208–212. [Google Scholar] [CrossRef]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef]
- Xin, L.; Hu, H.Y.; Ke, G.; Sun, Y.X. Effects of different nitrogen and phosphorus concentrations on the growth, nutrient uptake, and lipid accumulation of a freshwater microalga Scenedesmus sp. Bioresour. Technol. 2010, 101, 5494–5500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.M.; Chen, H.; He, C.L.; Wang, Q. Nitrogen starvation induced oxidative stress in an oil-producing green alga Chlorella sorokiniana C3. PLoS ONE 2013, 8, e69225. [Google Scholar] [CrossRef] [PubMed]
- Reitan, K.I.; Rainuzzo, J.R.; Olsen, Y. Effect of nutrient limitation on fatty acid and lipid content of marine microalgae. J. Phycol. 1994, 30, 972–979. [Google Scholar] [CrossRef]
- Lacour, T.; Sciandra, A.; Talec, A.; Mayzaud, P.; Bernard, O. Neutral lipid and carbohydrate productivities as a response to nitrogen status in Isochrysis sp. (T-ISO.; Haptophyceae): Starvation versus limitation. J. Phycol. 2012, 48, 647–656. [Google Scholar] [CrossRef]
- Li, Y.; Horsman, M.; Wang, B.; Wu, N.; Lan, C.Q. Effects of nitrogen sources on cell growth and lipid accumulation of green alga Neochloris oleoabundans. Appl. Microbiol. Biot. 2008, 81, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.; Deng, Z.; Fan, L.; Hu, Z. Lipid accumulation and growth characteristics of Chlorella zofingiensis under different nitrate and phosphate concentrations. J. Biosci. Bioeng. 2012, 114, 405–410. [Google Scholar] [CrossRef]
- Khozin-Goldberg, I.; Cohen, Z. The effect of phosphate starvation on the lipid and fatty acid composition of the fresh water eustigmatophyte Monodus subterraneus. Phytochemistry 2006, 67, 696–701. [Google Scholar] [CrossRef]
- Yao, C.H.; Ai, J.N.; Cao, X.P.; Xue, S. Characterization of cell growth and starch production in the marine green microalga Tetraselmis subcordiformis under extracellular phosphorus-deprived and sequentially phosphorus-replete conditions. Appl. Microbiol. Biot. 2013, 97, 6099–6110. [Google Scholar] [CrossRef]
- Jiang, Q.; Zhao, L.; Dai, J.; Wu, Q. Analysis of autophagy genes in microalgae: Chlorella as a potential model to study mechanism of autophagy. PLoS ONE 2012, 7, e41826. [Google Scholar] [CrossRef]
- McLachlan, J.L.; Marr, J.C.; Conlon-Keily, A.; Adamson, A. Effects of nitrogen concentration and cold temperature on DSP-toxin concentrations in the dinoflagellate Prorocentrum lima (Prorocentrales, Dinophyceae). Nat. Toxins 1994, 2, 263–270. [Google Scholar] [CrossRef]
- Wang, S.; Chen, J.; Li, Z.; Wang, Y.; Fu, B.; Han, X.; Zheng, L. Cultivation of the benthic microalga Prorocentrum lima for the production of diarrhetic shellfish poisoning toxins in a vertical flat photobioreactor. Bioresour. Technol. 2015, 179, 243–248. [Google Scholar] [CrossRef]
- Lee, T.C.; Fong, F.L.; Ho, K.C.; Lee, F.W. The mechanism of diarrhetic shellfish poisoning toxin production in Prorocentrum spp.: Physiological and molecular perspectives. Toxins 2016, 8, 272. [Google Scholar] [CrossRef]
- Aquino-Cruz, A.; Purdie, D.A.; Morris, S. Effect of increasing sea water temperature on the growth and toxin production of the benthic dinoflagellate Prorocentrum lima. Hydrobiologia 2018, 813, 103–122. [Google Scholar] [CrossRef]
- Hashimoto, K.; Uchida, H.; Nishimura, T.; Oikawa, H.; Funaki, H.; Honma, C.; Yamaguchi, H.; Suzuki, T.; Adachi, M. Determination of optimal culture conditions for toxin production by a Prorocentrum lima complex strain with high diarrhetic shellfish toxins yield. Harmful Algae 2021, 103, 102025. [Google Scholar] [CrossRef] [PubMed]
- Stukkens, Y.; Bultreys, A.; Grec, S.; Trombik, T.; Vanham, D.; Boutry, M. NpPDR1, a pleiotropic drug resistance-type ATP-binding cassette transporter from Nicotiana plumbaginifolia, plays a major role in plant pathogen defense. Plant Physiol. 2005, 139, 341–352. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Clean Reads | GC Content (%) | Q20 Percentage (%) | Number of Unigenes | BLASTx Analysis Percentage (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| KEGG | NR | Swiss-Prot | TrEMBL | KOG | GO | Pfam | ||||||
| 1P | 110,831,089 | 81,618,516 | 58.86 | 97.48 | 88,186 | 15.63 | 22.46 | 11.73 | 45.91 | 15.02 | 35.42 | 43.44 |
| 1/4P | 110,497,380 | 83,366,479 | 58.87 | 97.57 | 90,706 | 17.97 | 24.61 | 14.19 | 47.42 | 17.38 | 37.21 | 45.00 |
| 1/16P | 115,006,749 | 90,073,946 | 58.54 | 97.71 | 91,890 | 19.03 | 25.58 | 15.30 | 48.09 | 18.45 | 38.02 | 45.72 |
| Average Data | 112,111,739 | 85,019,647 | 58.76 | 97.59 | 90,261 | 17.54 | 24.22 | 13.74 | 47.14 | 16.95 | 36.88 | 44.72 |
| Functional Classification | Gene | Description | Unigene ID | log2 (Fold Change) (1/16P vs. 1P) |
|---|---|---|---|---|
| Ribosome | RP-S20e | Small subunit ribosomal protein S20e | Cluster-1527.83547 | 3.92 |
| RP-S11e | Small subunit ribosomal protein S11e | Cluster-1527.1298 | 3.56 | |
| RP-L34e | Large subunit ribosomal protein L34e | Cluster-1527.8476 | 3.77 | |
| RP-S5e | Small subunit ribosomal protein S5e | Cluster-1527.76135 | 3.65 | |
| RP-L10e | Large subunit ribosomal protein L10e | Cluster-5599.0 | 7.70 | |
| Stress–shock proteins | Hsp 70 | Heat shock 70 kDa protein, mitochondrial-like | Cluster-1527.8250 | 4.34 |
| Hsp 90 | Heat shock protein 90 | Cluster-1527.77292 | 5.04 | |
| Hsp 83 | Heat shock protein 83 | Cluster-1527.9757 | 4.57 | |
| HSPA1s | Heat shock 70 kDa protein 1/2/6/8 | Cluster-1527.86746 | 4.97 | |
| Photosynthesis system | PsbA | Photosystem II P680 reaction center D1 protein | Cluster-1527.39156 | −2.94 |
| PsbC | Photosystem II CP43 chlorophyll apoprotein | Cluster-1527.45732 | −3.30 | |
| PsbB | Photosystem II CP47 chlorophyll apoprotein | Cluster-1527.50539 | −2.51 | |
| PsaA | Photosystem I P700 chlorophyll a apoprotein A1 | Cluster-1527.41047 | −2.44 | |
| PsaB | Photosystem I P700 chlorophyll a apoprotein A2 | Cluster-1527.52290 | −2.79 | |
| PetB | Cytochrome b6 | Cluster-1527.15944 | −3.60 | |
| PetD | Cytochrome b6-f complex subunit 4 | Cluster-1527.82828 | −3.59 | |
| PetF | Ferredoxin | Cluster-1527.21762 | −8.44 | |
| Carbon fixation | PpdK | Pyruvate orthophosphate dikinase | Cluster-1527.73147 | 1.25 |
| RbcL | Ribulose bisphosphate carboxylase | Cluster-1527.41437 | 1.95 | |
| GAPA | Glyceraldehyde-3-phosphate dehydrogenase | Cluster-1527.43164 | 2.54 | |
| TPI | Triosephosphate isomerase | Cluster-1527.53219 | 1.05 | |
| Phagosome | RAB5A | Ras-related protein Rab-5A | Cluster-1527.16544 | 5.36 |
| CALR | Calreticulin | Cluster-5481.0 | 4.07 | |
| RAB7A | Ras-related protein Rab-7A | Cluster-1527.86789 | 4.29 | |
| SEC61A | Protein transport protein SEC61 subunit alpha | Cluster-5772.0 | 3.57 | |
| RAC1 | Ras-related C3 botulinum toxin substrate 1 | Cluster-1527.76712 | 3.78 | |
| Endocytosis | ARF1/2 | ADP-ribosylation factor 1/2 | Cluster-1527.8157 | 4.11 |
| SMAP | Stromal membrane-associated protein | Cluster-1527.1659 | 4.74 | |
| HSPA1s | Heat shock 70 kDa protein 1/2/6/8 | Cluster-1527.86746 | 4.97 | |
| RAB11A | Ras-related protein Rab-11A | Cluster-6525.0 | 4.52 | |
| CHMP4A/B | Charged multivesicular body protein 4A/B | Cluster-9796.0 | 4.04 | |
| CHMP3 | Charged multivesicular body protein 3 | Cluster-6541.0 | 4.52 | |
| Glycolysis, oxidative phosphorylation | PK | Pyruvate kinase | Cluster-1527.60580 | 0.91 |
| PD | Pyruvate dehydrogenase | Cluster-1527.62164 | 1.46 | |
| PGK | Phosphoglycerate kinase | Cluster-1527.83027 | 4.04 | |
| COX5B | Cytochrome c oxidase subunit 5b | Cluster-1527.419 | 4.42 | |
| SDHA | Succinate dehydrogenase (ubiquinone) Flavoprotein subunit | Cluster-1527.79011 | 4.92 | |
| ATP5B | F-type H+-transporting ATPase subunit beta | Cluster-1527.10945 | 4.34 | |
| ABC transporters | ABCA3 | ATP-binding cassette, subfamily A (ABC1), member 3 | Cluster-1527.71271 | 1.35 |
| ABCB1 | ATP-binding cassette, subfamily B (MDR/TAP), member 1 | Cluster-1527.63439 | 1.81 | |
| ABCC1 | ATP-binding cassette, subfamily C (CFTR/MRP), member 1 | Cluster-1527.10718 | 1.64 | |
| phosphate transporters | MPT | Mitochondrial phosphate transporter | Cluster-1527.80953 | 3.72 |
| TPT | Triose-phosphate transporter family | Cluster-1527.55083 | 1.41 | |
| Fatty acid biosynthesis | ACACA | Acetyl-CoA carboxylase/biotin carboxylase 1 | Cluster-1527.38700 | 2.00 |
| MCAT | Malonyl coenzyme A-acyl carrier protein transacylase | Cluster-1527.31766 | 1.16 | |
| FadD | Long-chain acyl-CoA synthetase | Cluster-4976.0 | 3.70 | |
| Ubiquitin-mediated proteolysis | UB | Ubiquitin family protein | Cluster-1527.11208 | 4.41 |
| UBE2D | Ubiquitin-conjugating enzyme E2 D | Cluster-1527.86049 | 4.35 | |
| UBE2N | Ubiquitin-conjugating enzyme E2 N | Cluster-1527.9310 | 4.67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, X.; Yao, G.; Wang, K.; Liu, Y.; Wang, F.; Jiang, H. Transcriptomic Analysis of the Response of the Toxic Dinoflagellate Prorocentrum lima to Phosphorous Limitation. Microorganisms 2023, 11, 2216. https://doi.org/10.3390/microorganisms11092216
Wan X, Yao G, Wang K, Liu Y, Wang F, Jiang H. Transcriptomic Analysis of the Response of the Toxic Dinoflagellate Prorocentrum lima to Phosphorous Limitation. Microorganisms. 2023; 11(9):2216. https://doi.org/10.3390/microorganisms11092216
Chicago/Turabian StyleWan, Xiukun, Ge Yao, Kang Wang, Yanli Liu, Fuli Wang, and Hui Jiang. 2023. "Transcriptomic Analysis of the Response of the Toxic Dinoflagellate Prorocentrum lima to Phosphorous Limitation" Microorganisms 11, no. 9: 2216. https://doi.org/10.3390/microorganisms11092216
APA StyleWan, X., Yao, G., Wang, K., Liu, Y., Wang, F., & Jiang, H. (2023). Transcriptomic Analysis of the Response of the Toxic Dinoflagellate Prorocentrum lima to Phosphorous Limitation. Microorganisms, 11(9), 2216. https://doi.org/10.3390/microorganisms11092216