

Whole-Genome Sequencing of Mycobacterium tuberculosis Isolates from Diabetic and Non-Diabetic Patients with Pulmonary Tuberculosis

 ,
,  ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Sample Processing
2.3. DNA Quality Check
2.4. DNA Library Preparation Protocol
2.5. Sequencing Protocol
2.6. Bioinformatics Analysis
2.7. Statistical Analysis
3. Results
3.1. Patient Characteristics
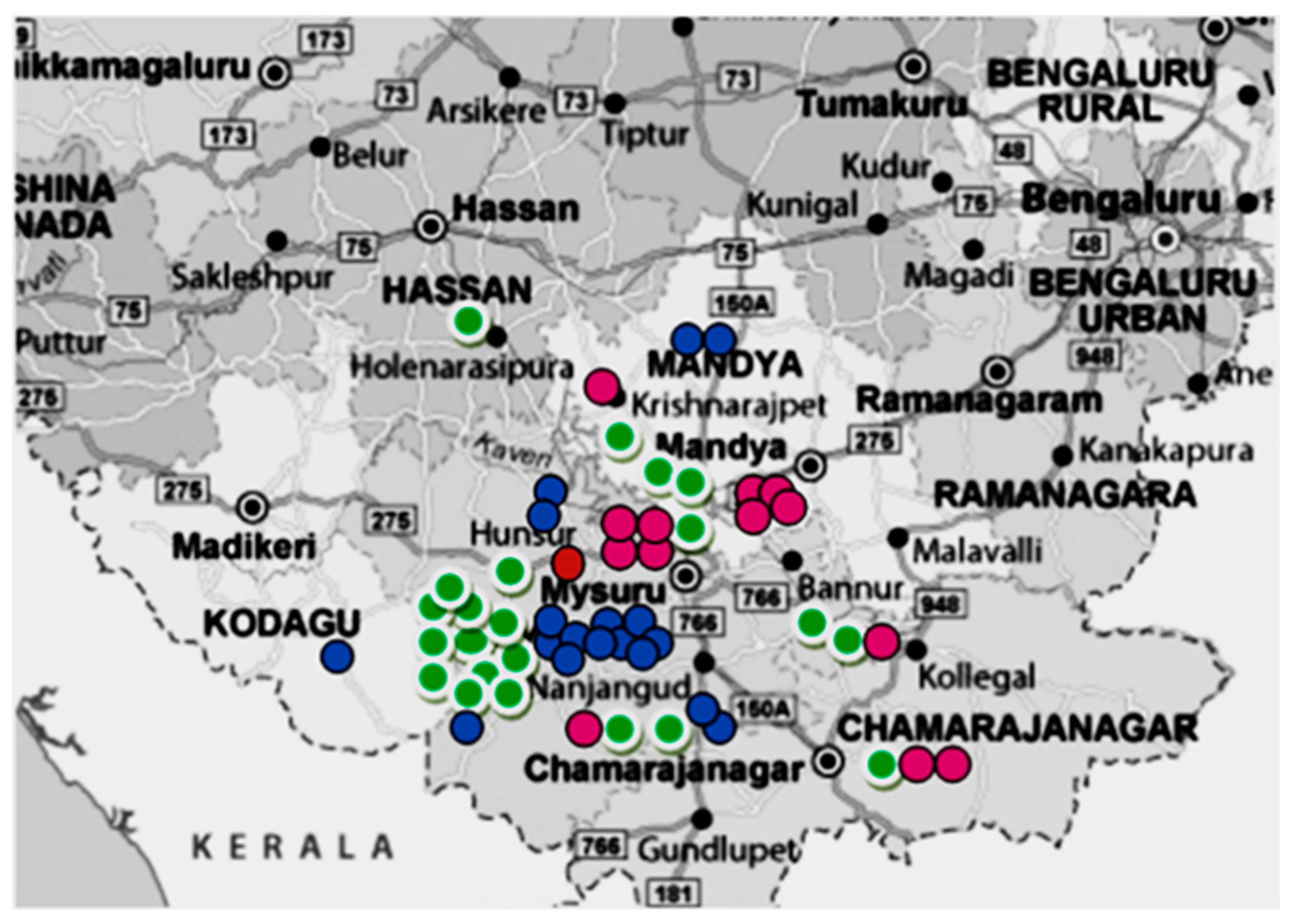
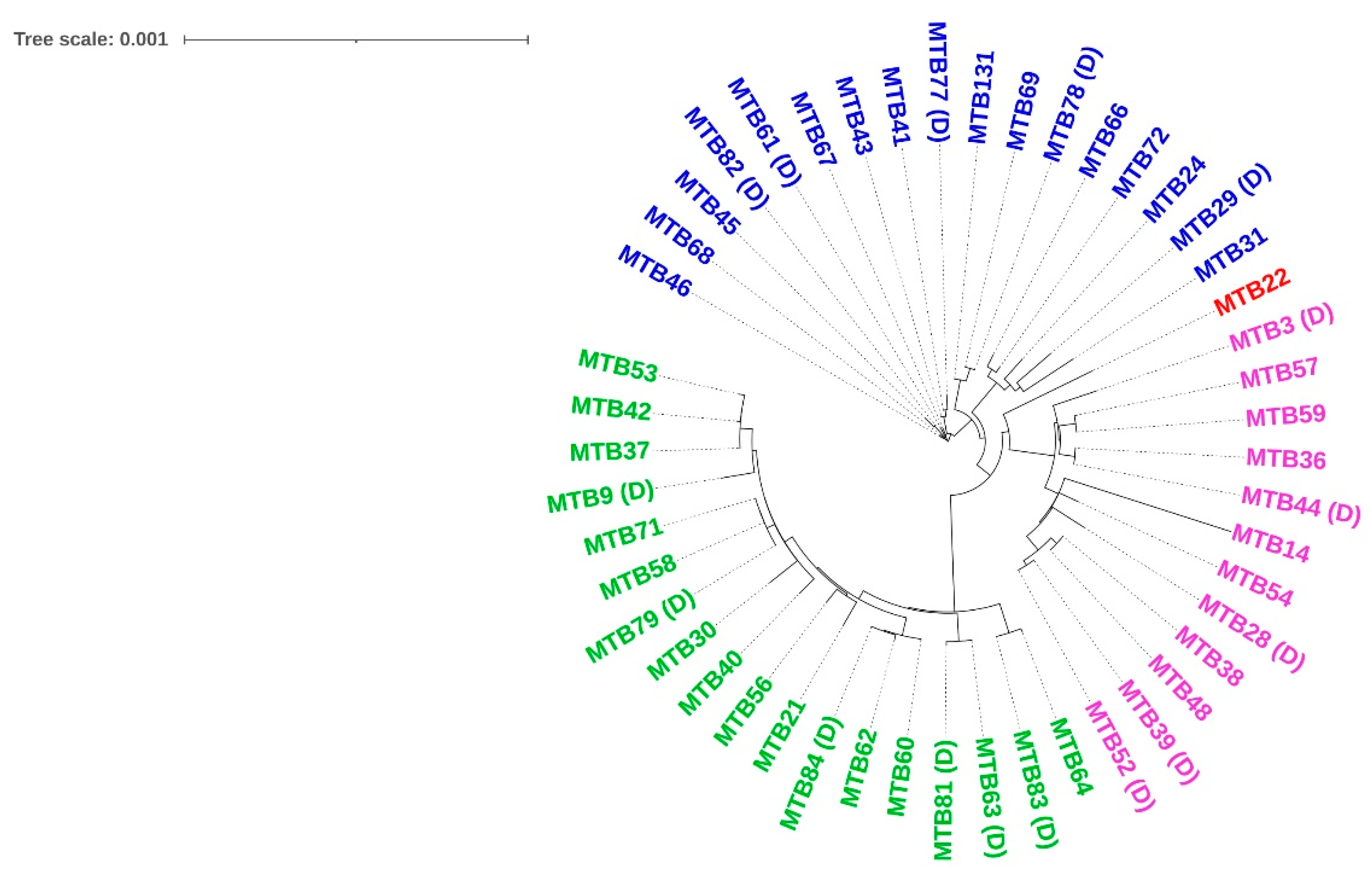
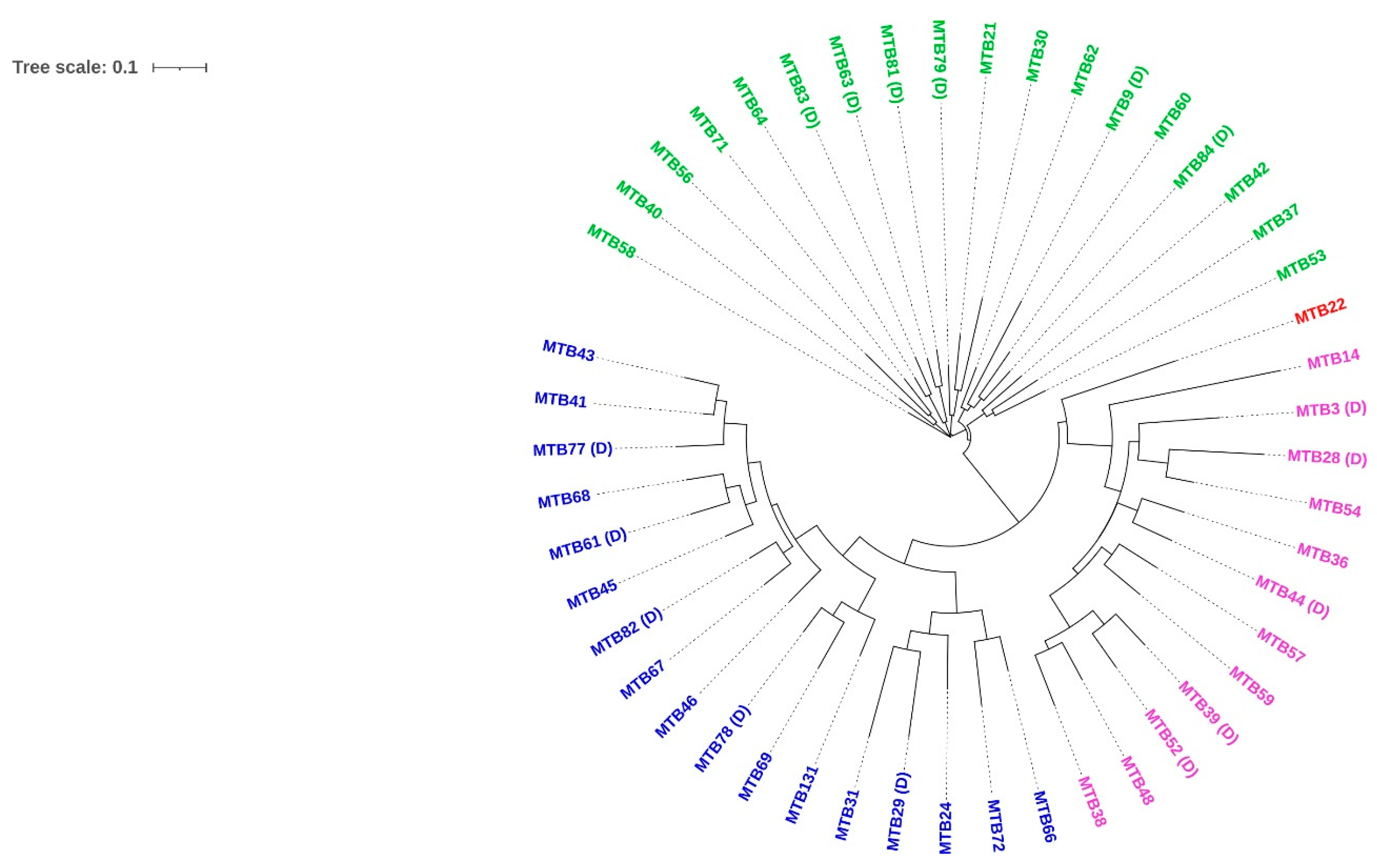
3.2. M. tuberculosis Genotypes
3.3. M. tuberculosis Drug Resistance
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO TB Report. 2019. Available online: http://library1.nida.ac.th/termpaper6/sd/2554/19755.pdf (accessed on 15 May 2020).
- Restrepo, B.I. Diabetes and tuberculosis. In Understanding the Host Immune Response against Mycobacterium tuberculosis Infection; Springer: Berlin/Heidelberg, Germany, 2018; pp. 1–21. [Google Scholar]
- Pan, S.C.; Ku, C.C.; Kao, D.; Ezzati, M.; Fang, C.T.; Lin, H.H. Effect of diabetes on tuberculosis control in 13 countries with high tuberculosis: A modelling study. Lancet Diabetes Endocrinol. 2015, 3, 323–330. Available online: https://pubmed.ncbi.nlm.nih.gov/25754415/ (accessed on 9 April 2021). [CrossRef] [PubMed]
- Hensel, R.L.; Kempker, R.R.; Tapia, J.; Oladele, A.; Blumberg, H.M.; Magee, M.J. Increased risk of latent tuberculous infection among persons with pre-diabetes and diabetes mellitus. Int. J. Tuberc. Lung Dis. 2016, 20, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Division Welfare CT. India TB Report 2019; Welfare, Ministry of Health and Family: New Delhi, India, 2019; p. 244. [Google Scholar]
- Dooley, K.E.; Chaisson, R.E. Tuberculosis and diabetes mellitus: Convergence of two epidemics. Lancet Infect. Dis. 2009, 9, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Hu, M.; Gao, F. Diabetes and pulmonary tuberculosis: A global overview with special focus on the situation in Asian countries with high TB-DM burden. Glob. Health Action 2017, 10, 1264702. [Google Scholar] [CrossRef]
- A Baker, M.; Harries, A.D.; Jeon, C.Y.; E Hart, J.; Kapur, A.; Lönnroth, K.; Ottmani, S.-E.; Goonesekera, S.D.; Murray, M.B. The impact of diabetes on tuberculosis treatment outcomes: A systematic review. BMC Med. 2011, 9, 81. [Google Scholar] [CrossRef]
- Barron, M.M.; Shaw, K.M.; Bullard, K.M.K.; Ali, M.K.; Magee, M.J. Diabetes is associated with increased prevalence of latent tuberculosis infection: Findings from the National Health and Nutrition Examination Survey, 2011–2012. Diabetes Res. Clin. Pract. 2018, 139, 366–379. Available online: http://www.diabetesresearchclinicalpractice.com/article/S0168822717316261/fulltext (accessed on 9 April 2021). [CrossRef]
- Lee, M.R.; Huang, Y.P.; Kuo, Y.T.; Luo, C.H.; Shih, Y.J.; Shu, C.C.; Wang, J.Y.; Ko, J.C.; Yu, C.J.; Lin, H.H. Diabetes Mellitus and Latent Tuberculosis Infection: A Systemic Review and Metaanalysis. Clin. Infect. Dis-Eases Off. Publ. Infect. Dis. Soc. Am. 2017, 64, 719–727. [Google Scholar] [CrossRef]
- Restrepo, B.I.; Schlesinger, L.S. Host-pathogen interactions in tuberculosis patients with type 2 diabetes mellitus Blanca. Tuberculosis 2013, 93, S10–S14. [Google Scholar] [CrossRef]
- Jiménez-Corona, M.E.; Cruz-Hervert, L.P.; García-García, L.; Ferreyra-Reyes, L.; Delgado-Sánchez, G.; Bobadilla-Del-Valle, M.; Canizales-Quintero, S.; Ferreira-Guerrero, E.; Báez-Saldaña, R.; Téllez-Vázquez, N.; et al. Association of diabetes and tuberculosis: Impact on treatment and post-treatment outcomes. Thorax 2013, 68, 214–220. [Google Scholar] [CrossRef]
- Lopez, K.; Arriaga, M.B.; Aliaga, J.G.; Barreda, N.N.; Sanabria, O.M.; Huang, C.C.; Zhang, Z.; García-de-la-Guarda, R.; Lecca, L.; Calçada Carvalho, A.C.; et al. Dysglycemia is associated with Mycobacterium tuberculosis lineages in tuberculosis patients of North Lima—Peru. PLoS ONE 2021, 16, e0243184. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Iwai, H.; Kato-Miyazawa, M.; Kirikae, T.; Miyoshi-Akiyama, T. CASTB (the comprehensive analysis server for the Mycobacterium tuberculosis complex): A publicly accessible web server for epidemiological analyses, drug-resistance prediction and phylogenetic com-par-ison of clinical isolates. Tuberculosis 2015, 95, 843–844. [Google Scholar] [CrossRef] [PubMed]
- Feuerriegel, S.; Schleusener, V.; Beckert, P.; Kohl, T.A.; Miotto, P.; Cirillo, D.M.; Cabibbe, A.M.; Niemann, S.; Fellenberg, K. PhyResSE: A Web Tool Delineating Mycobacterium tuberculosis Antibiotic Resistance and Lineage from Whole-Genome Sequencing Data. J. Clin. Microbiol. 2015, 53, 1908-14. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- India Tb Report National Tuberculosis 2021 Central TB Division-Directorate General of Helath Services, New Delhi. Available online: https://tbcindia.gov.in/showfile.php?lid=3587 (accessed on 20 January 2022).
- Manson, A.L.; Abeel, T.; Galagan, J.E.; Sundaramurthi, J.C.; Salazar, A.; Gehrmann, T.; Shanmugam, S.K.; Palaniyandi, K.; Narayanan, S.; Swaminathan, S.; et al. Mycobacterium tuberculosis Whole Genome Sequences from Southern India Suggest Novel Resistance Mechanisms and the Need for Region-Specific Diagnostics. Clin. Infect. Dis. 2017, 64, 1494–1501. [Google Scholar] [CrossRef] [PubMed]
- Reed, M.B.; Pichler, V.K.; McIntosh, F.; Mattia, A.; Fallow, A.; Masala, S.; Domenech, P.; Zwerling, A.; Thibert, L.; Menzies, D.; et al. Major Mycobacterium tuberculosis Lineages Associate with Patient Country of Origin. J. Clin. Microbiol. 2009, 47, 1119–1128. [Google Scholar] [CrossRef]
- Advani, J.; Verma, R.; Chatterjee, O.; Pachouri, P.K.; Upadhyay, P.; Singh, R.; Yadav, J.; Naaz, F.; Ravikumar, R.; Buggi, S.; et al. Whole ge-nome sequencing of Mycobacterium tuberculosis clinical isolates from India reveals ge-netic heterogeneity and region-specific variations that might affect drug susceptibility. Front. Microbiol. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Arora, J.; Singh, U.B.; Suresh, N.; Rana, T.; Porwal, C.; Kaushik, A.; Pande, J.N. Infection, Genetics and Evolution Characterization of predominant Mycobacterium tuberculosis strains from dif-ferent subpopulations of India. Infect. Genet. Evol. 2009, 9, 832–839. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, D.; Martinez, L.; Lu, P.; Zhu, L.; Lu, W.; Wang, J. Mycobacterium tuberculosis Bei-jing genotype strains and unfavourable treatment outcomes: A systematic review and meta-analysis. Clin. Microbiol. Infect. 2020, 26, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.R.; Pradhan, J.; Bhutia, R.; Dadul, P.; Sarkar, A.; Gohain, N.; Narain, K. Molecular diversity of Mycobacterium tuberculosis complex in Sikkim, India and prediction of dominant spoligotypes using artificial intelligence. Sci. Rep. 2021, 11, 7365. [Google Scholar] [CrossRef] [PubMed]
- Mburu, J.W.; Kingwara, L.; Esther, M.; Andrew, N. Molecular clustering of patients with Mycobacterium tuberculosis strains cultured from the diabetic and non-diabetic newly diagnosed TB positive cases. J. Clin. Tuberc. Other Mycobact. Dis. 2018, 12, 21–26. [Google Scholar] [CrossRef]
- Blanco-Guillot, F.; Delgado-Sánchez, G.; Mongua-Rodríguez, N.; Cruz-Hervert, P.; Ferreyra-Reyes, L.; Ferreira-Guerrero, E.; Yanes-Lane, M.; Montero-Campos, R.; Bobadilla-Del-Valle, M.; Torres-González, P.; et al. Molecular clustering of patients with diabetes and pulmonary tuberculosis: A systematic review and meta-analysis. PLoS ONE 2017, 12, e018467512. [Google Scholar] [CrossRef]
- Gagneux, S. Ecology and evolution of Mycobacterium tuberculosis. Nat. Rev. Microbiol. 2018, 16, 202–213. [Google Scholar] [CrossRef]
- Micheni, L.N.; Kassaza, K.; Kinyi, H.; Ntulume, I.; Bazira, J. Diversity of Mycobacterium tuberculosis Complex Lineages Associated with Pulmonary Tuberculosis in Southwestern, Uganda. Tuberc. Res. Treat. 2021, 2021, 5588339. Available online: http://europepmc.org/abstract/MED/34306752 (accessed on 20 January 2022). [CrossRef] [PubMed]
- Mogashoa, T.; Melamu, P.; Ley, S.D.; Streicher, E.M.; Iketleng, T.; Kelentse, N.; Mupfumi, L.; Mokomane, M.; Kgwaadira, B.; Novitsky, V.; et al. Genetic diversity of Mycobacterium tuberculosis strains circulating in Botswana. PLoS ONE 2019, 14, e0216306. [Google Scholar] [CrossRef] [PubMed]
- Rutaihwa, L.K.; Sasamalo, M.; Jaleco, A.; Hella, J.; Kingazi, A.; Kamwela, L.; Kingalu, A.; Malewo, B.; Shirima, R.; Doetsch, A.; et al. Insights into the genetic diversity of Mycobacterium tuberculosis in Tanzania. PLoS ONE 2019, 14, e0206334. [Google Scholar] [CrossRef]
- Taye, H.; Alemu, K.; Mihret, A.; Ayalew, S.; Hailu, E.; Wood, J.L.N.; Shkedy, Z.; Berg, S.; Aseffa, A. Epidemiology of Mycobacterium tuberculosis lineages and strain clustering within urban and pe-ri-urban settings in Ethiopia. PLoS ONE 2021, 16, e0253480. [Google Scholar] [CrossRef]
- Monteserin, J.; Paul, R.; Gravina, E.; Reniero, A.; Hernandez, T.; Mazzeo, E.; Togneri, A.; Simboli, N.; López, B.; Couvin, D.; et al. Genotypic diversity of Mycobacterium tuberculosis in Buenos Aires, Argentina. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2018, 62, 1–7. [Google Scholar] [CrossRef]
- Santos-Lazaro, D.; Gavilan, R.G.; Solari, L.; Vigo, A.N.; Puyen, Z.M. Whole genome analysis of extensively drug resistant Mycobacterium tuberculosis strains in Peru. Sci. Rep. 2021, 11, 9493. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Corrales, L.; Tovar-Aguirre, O.L.; Galeano-Vanegas, N.F.; Castaño Jiménez, P.A.; Martínez-Vega, R.A.; Maldonado-Londoño, C.E.; Hernández-Botero, J.S.; Siller-López, F. Phylogenomic analysis and Mycobacterium tuberculosis antibiotic resistance prediction by whole-genome sequencing from clinical isolates of Caldas, Colombia. PLoS ONE 2021, 16, e0258402. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; He, G.; Wang, S.; Lin, S.; Chen, J.; Zhang, W. Evaluation of Whole Genome Sequence Method to Diagnose Resistance of 13 Anti-tuberculosis Drugs and Characterize Resistance Genes in Clinical Multi-Drug Resistance Mycobacterium tuberculosis Isolates From China. Front. Microbiol. 2019, 10, 1741. Available online: https://www.frontiersin.org/article/10.3389/fmicb.2019.01741 (accessed on 19 April 2021). [CrossRef] [PubMed]
- Wu, B.; Zhu, W.; Wang, Y.; Wang, Q.; Zhou, L.; Liu, Z.; Bi, L.; Barun, M.; Kreiswirth, B.N.; Chen, L.; et al. Genetic composition and evolution of the prevalent Mycobacterium tuberculosis lineages 2 and 4 in the Chinese and Zhejiang Province populations. Cell Biosci. 2021, 11, 162. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, J.; Chen, L.; Wang, W.; Yu, F.; Xiong, H. Whole-genome sequencing of Mycobacterium tuberculosis for prediction of drug resistance. Epidemiol. Infect. 2022, 150, e22. [Google Scholar] [CrossRef]
- Mizukoshi, F.; Kobayashi, N.; Kirikae, F.; Ohta, K.; Tsuyuguchi, K.; Yamada, N.; Inoue, Y.; Horiba, M.; Kawata, N.; Ichinose, A.; et al. Molecular Epidemiology of Drug-Resistant Mycobacterium tuberculosis in Japan. mSphere 2021, 6, e0097820. [Google Scholar] [CrossRef]
- Maeda, S.; Hijikata, M.; Hang, N.T.L.; Thuong, P.H.; Huan, H.V.; Hoang, N.P.; Hung, N.V.; Cuong, V.C.; Miyabayashi, A.; Seto, S.; et al. Genotyping of Mycobacterium tuberculosis spreading in Hanoi, Vietnam using conventional and whole genome sequencing methods. Infect. Genet. Evol. 2020, 78, 104107. [Google Scholar] [CrossRef]
- Smittipat, N.; Miyahara, R.; Juthayothin, T.; Billamas, P.; Dokladda, K.; Imsanguan, W.; Intralawan, D.; Rukseree, K.; Jaitrong, S.; Chaiyasirinroje, B.; et al. Indo-Oceanic Mycobacterium tuberculosis strains from Thailand associated with higher mortality. Int. J. Tuberc. Lung Dis. Off. J. Int. Union Against Tuberc. Lung Dis. 2019, 23, 972–979. [Google Scholar] [CrossRef]
- Phelan, J.E.; Lim, D.R.; Mitarai, S.; de Sessions, P.F.; Tujan, M.A.A.; Reyes, L.T.; Medado, I.A.P.; Palparan, A.G.; Naim, A.N.M.; Jie, S.; et al. Mycobacterium tuberculosis whole genome sequencing provides insights into the Manila strain and drug-resistance mutations in the Philippines. Sci. Rep. 2019, 9, 9305. [Google Scholar] [CrossRef]
- Bainomugisa, A.; Meumann, E.M.; Rajahram, G.S.; Ong, R.T.; Coin, L.; Paul, D.C.; William, T.; Coulter, C.; Ralph, A.P. Genomic epidemiology of tuberculosis in eastern Malaysia: Insights for strengthening public health responses. Microb Genom. 2021, 7, 000573. [Google Scholar] [CrossRef]
- Manson, A.L.; Cohen, K.A.; Abeel, T.; Desjardins, C.A.; Armstrong, D.T.; Barry, C.E., 3rd; Brand, J. Genomic analysis of globally diverse Mycobacterium tuberculosis strains provides insights into the emergence and spread of multidrug resistance. Nat. Genet. 2017, 49, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.V.; Singh, S.; Yadav, A.; Kushwah, S.; Yadav, R.; Sai, D.K.; Chauhan, D.S. Genetic variability in multidrug-resistant Mycobacterium tuberculosis isolates from patients with pulmonary tuberculosis in North India. BMC Microbiol. 2021, 21, 123. [Google Scholar] [CrossRef] [PubMed]
- Suzana, S.; Shanmugam, S.; Uma Devi, K.R.; Swarna Latha, P.N.; Michael, J.S. Spoligotyping of Mycobacterium tuberculosis isolates at a tertiary care hospital in India. Trop. Med. Int. Health TM IH 2017, 22, 703–707. [Google Scholar] [CrossRef]
- Tejashree, A.; Mahesh, P.A.; Krishna Karthik, M.; Nirmala Azeem, A.; Reddy, R.H.R.; Ravichandra, C.; Nagaraja, S.B. Era of TB elimination: Growing need to understand diversities of Mycobacterium tuberculosis lineages! Indian J. Tuberc. 2022, 69, 79–84. [Google Scholar] [CrossRef]
- Gupta, A.; Sinha, P.; Nema, V.; Gupta, P.K.; Chakraborty, P.; Kulkarni, S.; Rastogi, N.; Anupurba, S. Detection of Beijing strains of MDR M. tuberculosis and their association with drug resistance mu-ta-tions in katG, rpoB, and embB genes. BMC Infect. Dis. 2020, 20, 752. [Google Scholar] [CrossRef]
- Prakash, R.; Gupta, R.; Sharma, P.; Jain, S.; Chauhan, D.S.; Katoch, V.M.; Tiwari, P.K. Genotypic diversity of Mycobacterium tuberculosis isolates from North-Central Indian population. Pathog. Glob. Health 2019, 113, 39–48. [Google Scholar] [CrossRef]
- Gupta, R.; Amrathlal, R.S.; Prakash, R.; Jain, S.; Tiwari, P.K. Spoligotyping, phenotypic and genotypic characterization of katG, rpoB gene of M. tuberculosis isolates from Sahariya tribe of Madhya Pradesh India. J. Infect. Public Health 2019, 12, 395–402. [Google Scholar] [CrossRef]
- Desikan, P.; Chauhan, D.S.; Sharma, P.; Panwalkar, N.; Chourey, M.; Patidar, M.L.; Yadav, P.; Chandrasekaran, V.; Ohri, B.S. Genetic diversity of Mycobacterium tuberculosis isolates from central India. Indian J. Med. Res. 2016, 143, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Sankar, M.M.; Kumar, P.; Couvin, D.; Rastogi, N.; Singh, S. Genetic Diversity and Drug Susceptibility Profile of Mycobacterium tuberculosis Isolated from Different Regions of India. J. Infect. 2015, 71, 207–219. Available online: http://www.journalofinfection.com/article/S0163445315001218/fulltext (accessed on 20 December 2020). [CrossRef]
- Kandhakumari, G.; Stephen, S.; Sivakumar, S.; Narayanan, S. Spoligotype patterns of Mycobacterium tuberculosis isolated from extra pulmonary tuberculosis patients in Puducherry, India. Indian J. Med. Microbiol. 2015, 33, 267–270. [Google Scholar] [CrossRef]
- Varma-Basil, M.; Kumar, S.; Arora, J.; Angrup, A.; Zozio, T.; Banavaliker, J.N.; Singh, U.B.; Rastogi, N.; Bose, M. Comparison of spoligotyping, mycobacterial interspersed repetitive units typing and IS6110-RFLP in a study of genotypic diversity of Mycobacterium tuberculosis in Delhi, North India. Mem. Do Inst. Oswaldo Cruz 2011, 106, 524–535. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.; Rodrigues, C.; Ashavaid, T.F.; Lalvani, A.; Udwadia, Z.F.; Mehta, A. High incidence of the Beijing genotype among multidrug-resistant isolates of Mycobacterium tuberculosis in a tertiary care center in Mumbai, India. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2005, 40, 881–886. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kulkarni, S.; Sola, C.; Filliol, I.; Rastogi, N.; Kadival, G. Spoligotyping of Mycobacterium tuberculosis isolates from patients with pulmonary tuberculosis in Mumbai, India. Res. Microbiol. 2005, 156, 588–596. [Google Scholar] [CrossRef]
- Sankar, M.M.; Singh, J.; Diana, S.C.A.; Singh, S. Molecular characterization of Mycobacterium tuberculosis isolates from North Indian patients with extrapulmonary tuberculosis. Tuberculosis 2013, 93, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.K.; Iravatham, C.C.; Moni, B.H.; Kumar, A.; Archana, B.V.; Majid, M.; Priyadarshini, Y.; Rani, P.S.; Valluri, V.; Hasnain, S.E.; et al. Modern and ancestral genotypes of Mycobacterium tuberculosis from Andhra Pradesh, India. PLoS ONE 2011, 6, e27584. [Google Scholar] [CrossRef]
- Rahman, S.M.M.; Rahman, A.; Nasrin, R.; Ather, M.F.; Ferdous, S.S.; Ahmed, S.; Uddin, M.K.M.; Khatun, R.; Sarker, M.S.; Mahmud, A.M.; et al. Molecular Epidemiology and Genetic Diversity of Multidrug-Resistant Mycobacterium tuberculosis Isolates in Bangladesh. Microbiol. Spectr. 2022, 10, e0184821. [Google Scholar] [CrossRef]
- Zenteno-Cuevas, R.; Munro-Rojas, D.; Pérez-Martínez, D.; Fernandez-Morales, E.; Jimenez-Ruano, A.C.; Montero, H.; Escobar, L.; de Igartua, E.; Trigos, Á.; Fuentes-Dominguez, J. Genetic diversity and drug susceptibility of My-co-bacterium tuberculosis in a city with a high prevalence of drug resistant tuberculosis from Southeast of Mexico. BMC Infect. Dis. 2021, 21, 1202. [Google Scholar] [CrossRef]
- Ndungu, P.W.; Kariuki, S.; Revathi, G.; Ng, Z.; Niemann, S. Mycobacteria Interspersed Repetitive Units-Variable Number of Tandem Repeat, Spoligotyping and Drug Re-sistance of Isolates from Pulmonary Tuberculosois Patients in Kenya. Adv. Microbiol. 2017, 7, 205–216. [Google Scholar] [CrossRef]
- Fernandes, T.; Osório, C.; Sousa, M.J.; Franco-Duarte, R. Contributions of Adaptive Laboratory Evolution towards the Enhancement of the Biotechnological Potential of Non-Conventional Yeast Species. J. Fungi 2023, 9, 186. [Google Scholar] [CrossRef]
- Franco-Duarte, R.; Bessa, D.; Gonçalves, F.; Martins, R.; Silva-Ferreira, A.C.; Schuller, D.; Sampaio, P.; Pais, C. Genomic and transcriptomic analysis of Saccharomyces cerevisiae isolates with focus in succinic acid production. FEMS Yeast Res. 2017, 17, fox057. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Characteristics | Total | DM | Non-DM | p-Value |
|---|---|---|---|---|
| Gender | ||||
| Male | 40 | 13 (23.63%) | 27 (49.09%) | 0.33 (NS) |
| Female | 15 | 7 (12.72%) | 8 (14.54%) | |
| Age (years) | ||||
| <30 | 10 | 0 | 10 (18.18%) | 0.014 (S) |
| 30–49 | 24 | 7 (12.72%) | 17 (30.90%) | |
| 50–69 | 16 | 11 (20%) | 5 (9.09%) | |
| 70+ | 5 | 2 (3.63%) | 3 (5.45%) | |
| Education | ||||
| Primary school | 12 | 3 (5.45%) | 9 (16.36%) | 0.64 (NS) |
| Middle school | 2 | 0 | 2 (3.63%) | |
| Secondary school | 10 | 5 (9.09%) | 5 (9.09%) | |
| Pre-University | 5 | 2 (3.63%) | 3 (5.45%) | |
| Undergraduate | 2 | 2 (3.63%) | 0 | |
| No qualification | 24 | 8 (14.54%) | 16 (29.09%) | |
| Occupation | ||||
| Student | 3 | 0 | 3 (5.45%) | 0.87 (NS) |
| Agriculturist | 11 | 6 (10.90%) | 5 (9.09%) | |
| Daily-wages laborer | 6 | 3 (5.45%) | 3 (5.45%) | |
| Business | 9 | 3 (5.45%) | 6 (10.90%) | |
| Housewife | 5 | 2 (3.63% | 3 (5.45%) | |
| Unknown/unemployed | 21 | 6 (10.90%) | 15 (27.27%) | |
| Co-morbidities apart from DM | ||||
| Hypertension | 2 | 1 (5.26%) | 1 (5.26%) | 0.31 (NS) |
| Asthma | 7 | 1 (5.26%) | 6 (31.57%) | 0.91 (NS) |
| Chronic obstructive pulmonary disease | 1 | 1 (5.26%) | 0 | 1 (NS) |
| Ischemic heart disease | 2 | 1(5.26%) | 1 (5.26%) | 0.69 (NS) |
| Smoker | 7 | 4 (21.05%) | 3 (15.78%) | 0.22 (NS) |
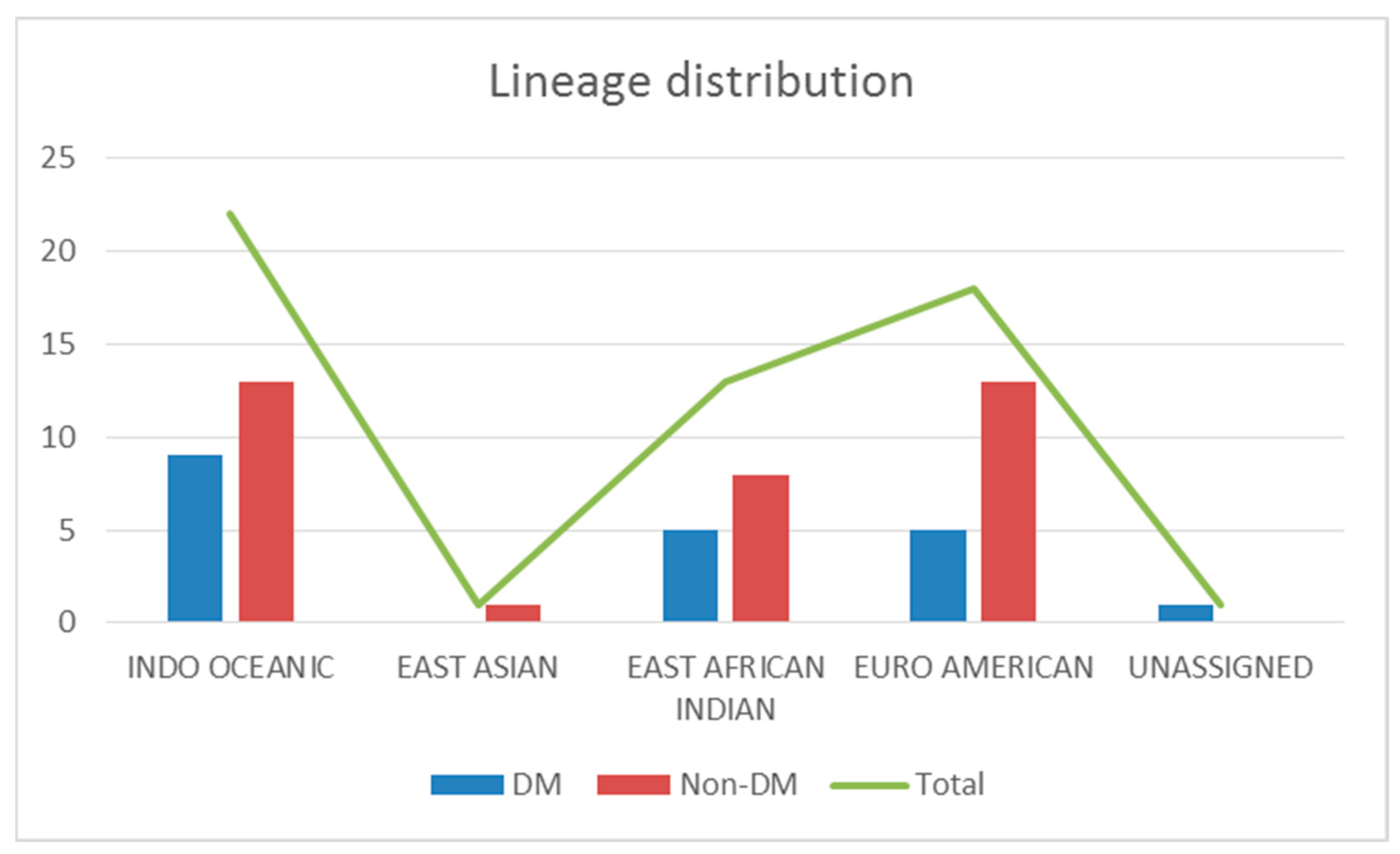
| Lineage | Charmrajnagar | Hassan | Hd Kote | Hunsur | Kodagu | Kr Pete | Mandya | Mysore | Nanjangud | Srirangapatana | T.narsipura | Total | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ND | DM | ND | DM | ND | DM | ND | DM | ND | DM | ND | DM | ND | DM | ND | DM | ND | DM | ND | DM | ND | DM | ||
| Euro American | 1 | 2 | 1 | 1 | 1 | 6 | 4 | 2 | 18 | ||||||||||||||
| East Asian | 1 | 1 | |||||||||||||||||||||
| East African Indian | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 1 | 1 | 13 | |||||||||||||
| Indo Oceanic | 1 | 1 | 1 | 3 | 8 | 3 | 2 | 1 | 2 | 22 | |||||||||||||
| 1 | 2 | 1 | 0 | 1 | 1 | 2 | 0 | 1 | 0 | 1 | 0 | 3 | 6 | 17 | 9 | 5 | 0 | 0 | 1 | 3 | 0 | ||
| Total | 3 | 1 | 2 | 2 | 1 | 1 | 9 | 26 | 5 | 1 | 3 | ||||||||||||
| Main Spoligotype | Region of Difference | Total | DM | Non-DM | p-Value |
|---|---|---|---|---|---|
| CAS | RD 750 | 12 | 6 | 6 | NS |
| EAI | RD 239 | 22 | 8 | 14 | 0.31 (NS) |
| Beijing | RD 181 | 1 | 0 | 1 | NS |
| T | RD 182 | 5 | 1 | 4 | 0.16 (NS) |
| T | RD 219 | 4 | 1 | 3 | 0.30 (NS) |
| LAM | RD 219 | 10 | 3 | 7 | |
| Unassigned | - | 1 | 1 | - | 0.16 (NS) |
| Drug resistance | |||||
| Not detected | 46 | 17 | 29 | 0.83 (NS) | |
| Isoniazid | 3 | 1 | 2 | 1 (NS) | |
| Streptomycin | 2 | 0 | 2 | NS | |
| Isoniazid + Ethambutol | 1 | 1 | 0 | NS | |
| Isoniazid + Streptomycin | 1 | 0 | 1 | NS | |
| Ethionamide + Streptomycin | 2 | 1 | 1 | 1 (NS) | |
| MDR | 0 | 0 | 0 | NS | |
| Total | 55 | 20 | 35 | ||
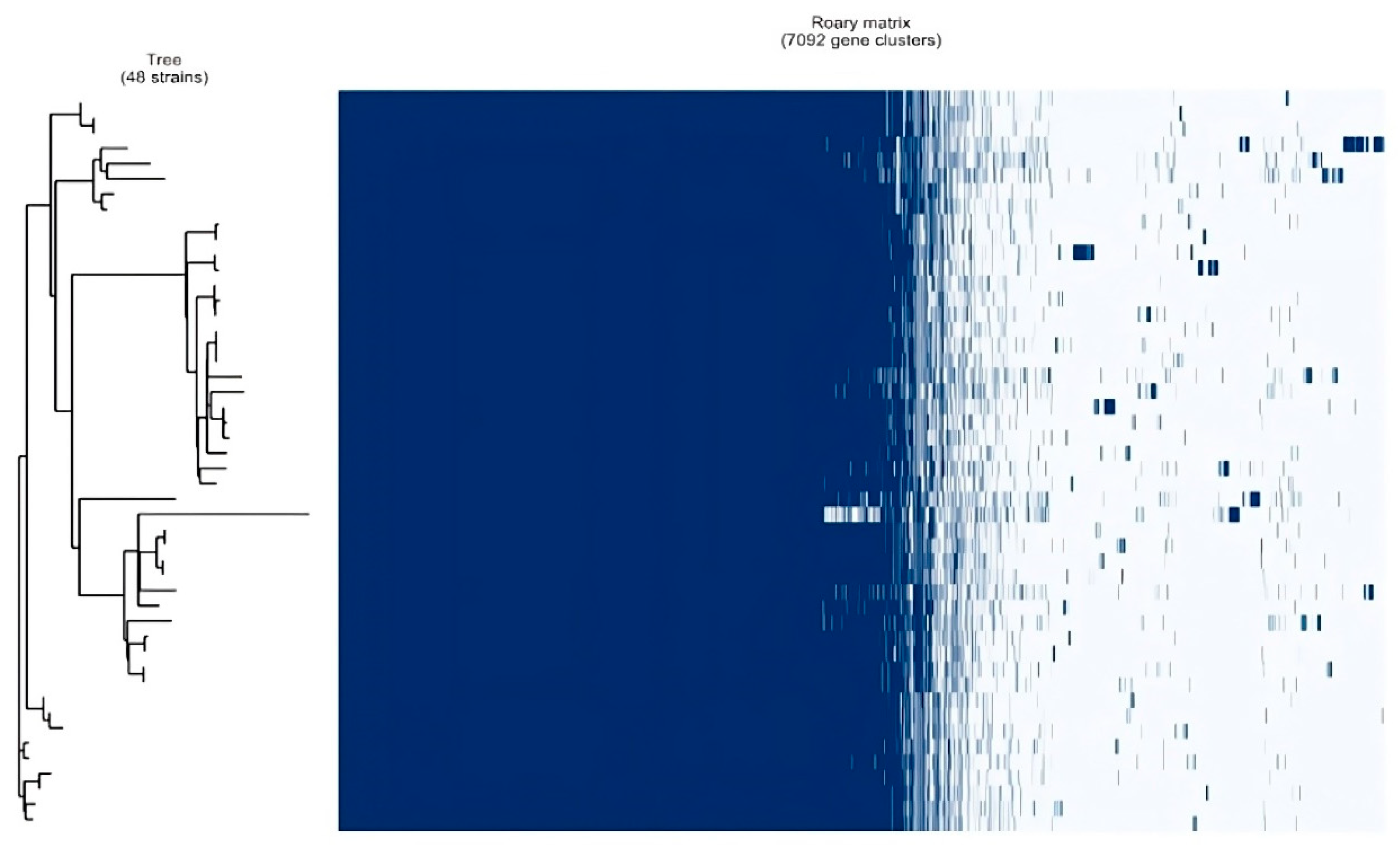
| Genes | Total Number | |
|---|---|---|
| Core genes | (99% <= strains <= 100%) | 3283 |
| Soft core genes | (95% <= strains < 99%) | 435 |
| Shell genes | (15% <= strains < 95%) | 575 |
| Cloud genes | (0% <= strains < 15%) | 2799 |
| Total genes | (0% <= strains <= 100%) | 7092 |
| An Overview of Various Studies Across the World: | |||||
|---|---|---|---|---|---|
| Study Conducted in | Year | Predominant Lineage | Predominant Spoligotype | Method | Reference in Discussion |
| Uganda, East Africa | 2021 | Euro-American (74.2%) | - | SNP typing by PCR | 30 |
| Botswana, South Africa | 2019 | Euro-American (81.9%) | LAM (33%), T (16%) | Spoligotyping and MIRU-VNTR | 31 |
| Tanzania, East Africa | 2019 | Euro-American (42.5%) | - | SNP typing by PCR | 32 |
| Ethiopia, East Africa | 2021 | Euro-American (61.6%) | - | LSP typing by PCR | 33 |
| Argentina, S.America | 2018 | Euro-American (99%) | T (35.9%), LAM (33.2%) | Spoligotyping | 34 |
| Peru, South America | 2021 | Euro-American (91.2%) | LAM (26.47%), Harleem (23.5%) | WGS | 35 |
| Colombia, S.America | 2021 | Euro-American (100%) | - | WGS | 36 |
| China | 2017 | East Asian (42.1%) | - | LSP typing and MIRU-VNTR | 37 |
| China | 2021 | East Asian (74.38%) | - | WGS | 38 |
| Shanghai | 2022 | East Asian (97.4%) | - | WGS | 39 |
| Japan | 2021 | East Asian (78.3%) | - | WGS | 40 |
| Vietnam | 2019 | East Asian (57.2%) | - | WGS | 41 |
| Thailand | 2019 | East Asian (44.6%), Indo-Oceanic (40%) | - | WGS | 42 |
| Philippines | 2019 | Indo-Oceanic (80.3%) | - | WGS | 43 |
| Malaysia | 2021 | Indo-Oceanic (93.8%) | - | WGS | 44 |
| South India (NIT, Delhi) | 2017 | Indo Oceanic (70%) | - | WGS | 45 |
| North India (JALMA, UP) | 2021 | East-African Indian (66.25%) | CAS (65%), Beijing (14.1%) | Spoligotyping | 46 |
| Study conducted in | Year | Predominant Spoligotype | Method | Reference | |
| CMC Vellore, South India | 2017 | North Indian isolates—Beijing (23.4%), South Indian isolates—(EAI 43%) | Spoligotyping | 47 | |
| JSSMC, Mysore, South India | 2022 | EAI (46%) | Spoligotyping | 48 | |
| Sikkim, North India | 2021 | Beijing type (62.41%) | Spoligotyping and MIRU-VNTR | 26 | |
| Varanasi, North India | 2020 | Beijing type (19.95%) | Spoligotyping | 49 | |
| JALMA, Agra, North India | 2019 | EAI (51%), CAS (19%) | Spoligotyping and MIRU-VNTR | 50 | |
| Madhya Pradesh, Central India | 2019 | CAS | Spoligotyping and RFLP | 51 | |
| Bhopal, MP, Central India | 2016 | CAS (70%), EAI (30%) | Spoligotyping | 52 | |
| AIIMS, Delhi, North India | 2015 | CAS (35.4%), EAI (24.2%) | Spoligotyping | 53 | |
| Pondicherry, South India | 2015 | EAI (41.8%) | Spoligotyping | 54 | |
| Delhi University, North India | 2011 | CAS | Spoligotyping and RFLP | 55 | |
| Hinduja Hospital, Mumbai | 2005 | Beijing type (35%) | RFLP | 56 | |
| Baba Atomic research, Mumbai | 2005 | CAS (30%), EAI (17%) | Spoligotyping | 57 | |
| AIIMS, Delhi, North India | 2012 | CAS (57.27%) | Spoligotyping | 58 | |
| Andhra Pradesh, South India | 2011 | CAS (40%), EAI (38%) | Spoligotyping | 59 | |
| Bangladesh | 2022 | Beijing type (38%) | Spoligotyping | 60 | |
| Mexico | 2021 | H (32%), T (23%) | Spoligotyping and MIRU-VNTR | 61 | |
| Kenya | 2017 | CAS (28.9%), LAM & Beijing (17.6%) | Spoligotyping and MIRU-VNTR | 62 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shankaregowda, R.; Allegretti, Y.H.; Sumana, M.N.; Rao, M.R.; Raphael, E.; Mahesh, P.A.; Riley, L.W. Whole-Genome Sequencing of Mycobacterium tuberculosis Isolates from Diabetic and Non-Diabetic Patients with Pulmonary Tuberculosis. Microorganisms 2023, 11, 1881. https://doi.org/10.3390/microorganisms11081881
Shankaregowda R, Allegretti YH, Sumana MN, Rao MR, Raphael E, Mahesh PA, Riley LW. Whole-Genome Sequencing of Mycobacterium tuberculosis Isolates from Diabetic and Non-Diabetic Patients with Pulmonary Tuberculosis. Microorganisms. 2023; 11(8):1881. https://doi.org/10.3390/microorganisms11081881
Chicago/Turabian StyleShankaregowda, Ranjitha, Yuan Hu Allegretti, Mahadevaiah Neelambike Sumana, Morubagal Raghavendra Rao, Eva Raphael, Padukudru Anand Mahesh, and Lee W. Riley. 2023. "Whole-Genome Sequencing of Mycobacterium tuberculosis Isolates from Diabetic and Non-Diabetic Patients with Pulmonary Tuberculosis" Microorganisms 11, no. 8: 1881. https://doi.org/10.3390/microorganisms11081881
APA StyleShankaregowda, R., Allegretti, Y. H., Sumana, M. N., Rao, M. R., Raphael, E., Mahesh, P. A., & Riley, L. W. (2023). Whole-Genome Sequencing of Mycobacterium tuberculosis Isolates from Diabetic and Non-Diabetic Patients with Pulmonary Tuberculosis. Microorganisms, 11(8), 1881. https://doi.org/10.3390/microorganisms11081881