

Insights and Perspectives on the Role of Proteostasis and Heat Shock Proteins in Fungal Infections

 ,
,  and
and
Abstract
1. Introduction
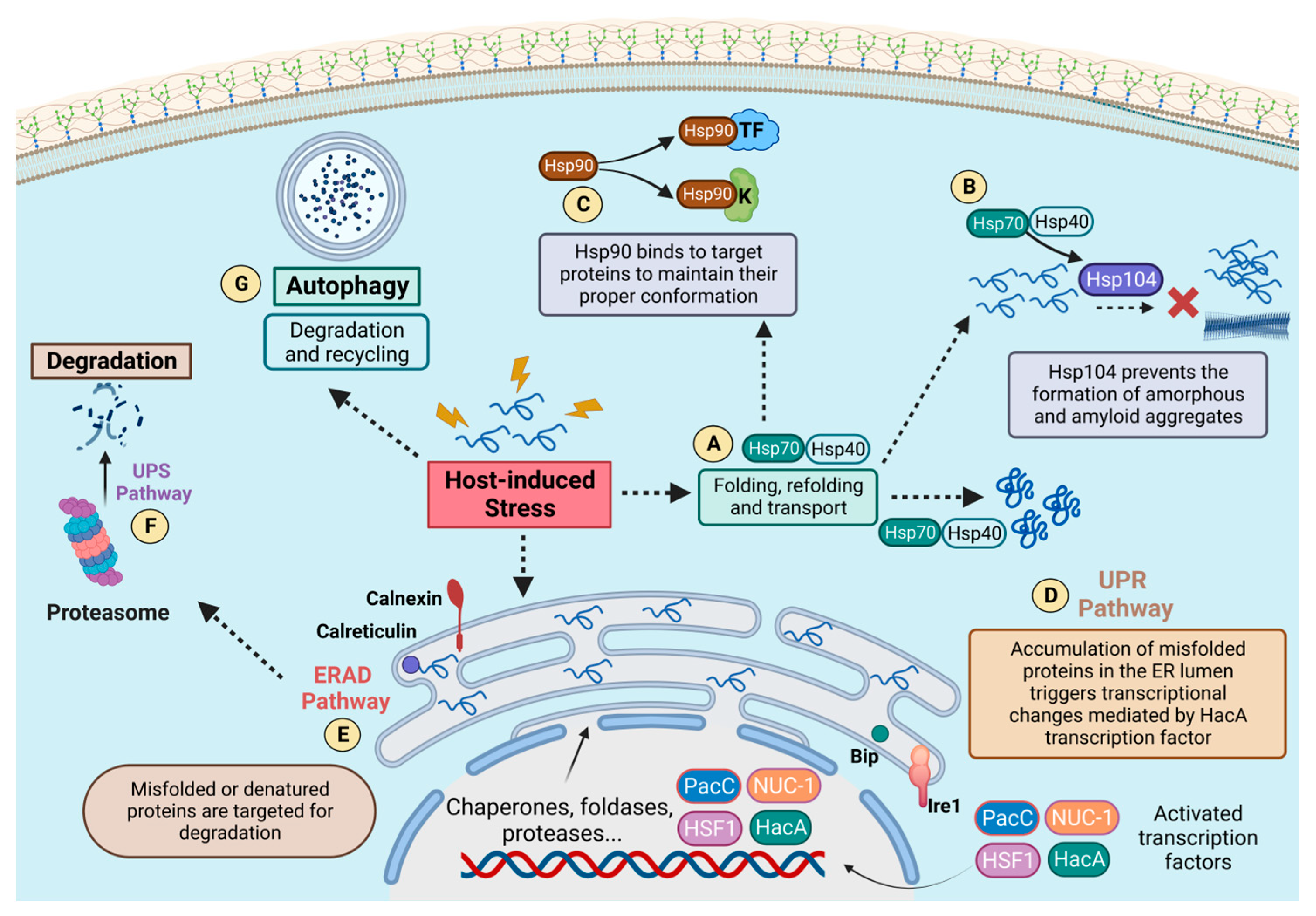
2. Proteostasis Circuits and Pathogenesis
2.1. Proteostasis during Infection
2.2. HSPs and Host–Pathogen Interactions
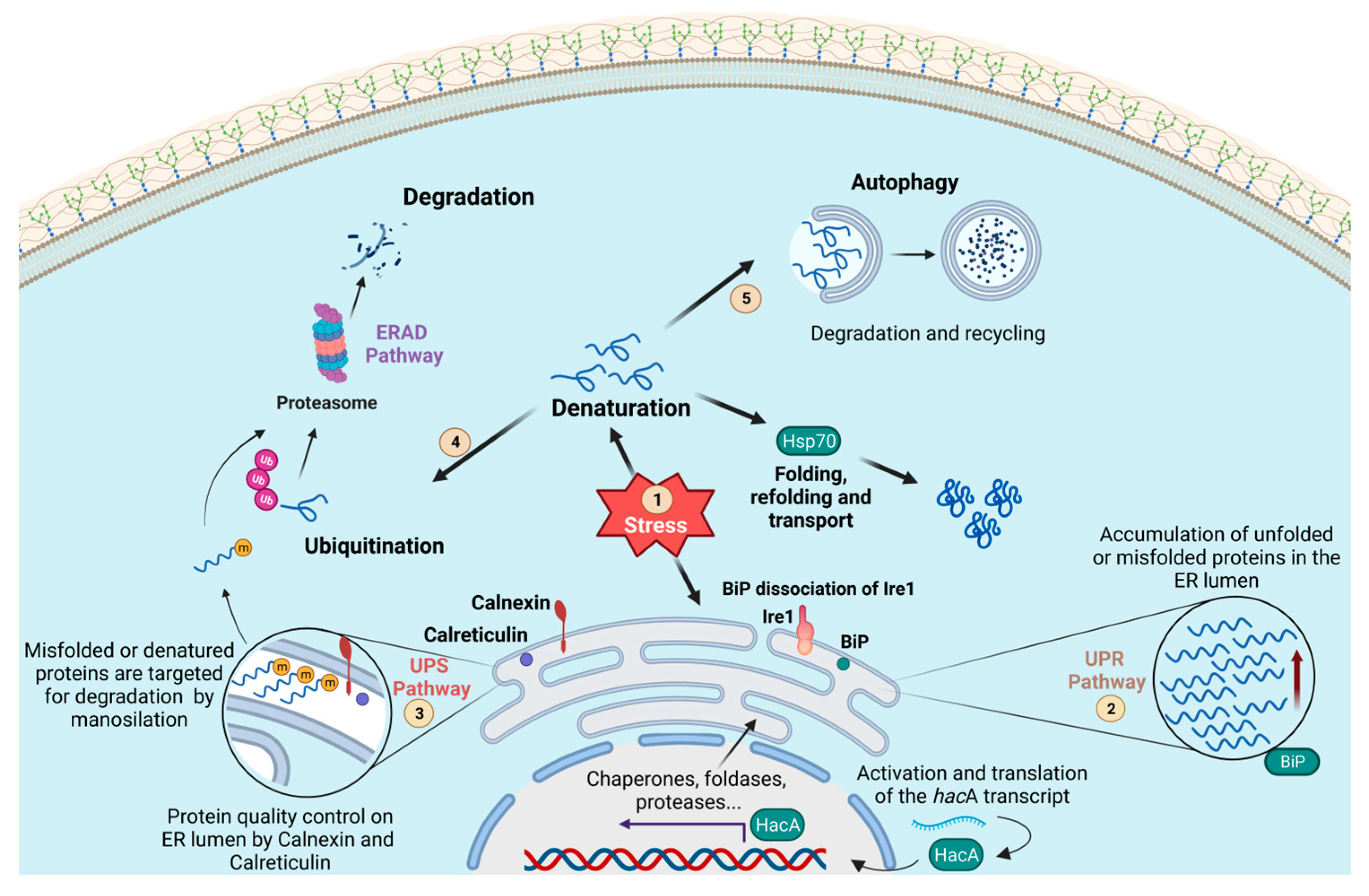
2.3. Protein Degradation and The Unfolded Protein Response
3. Heat Shock Proteins
3.1. HSP Paralogs
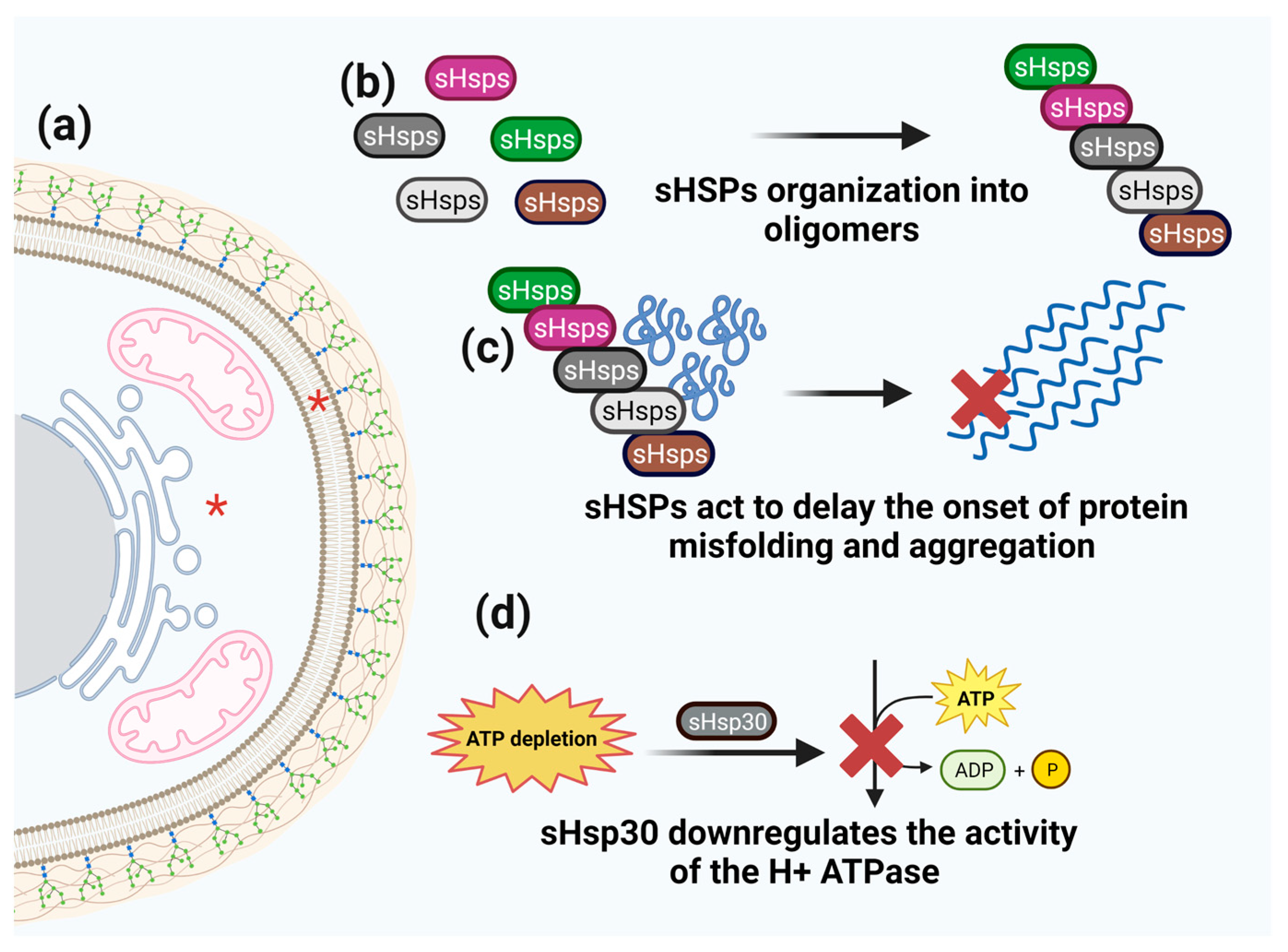
3.2. Small HSPs
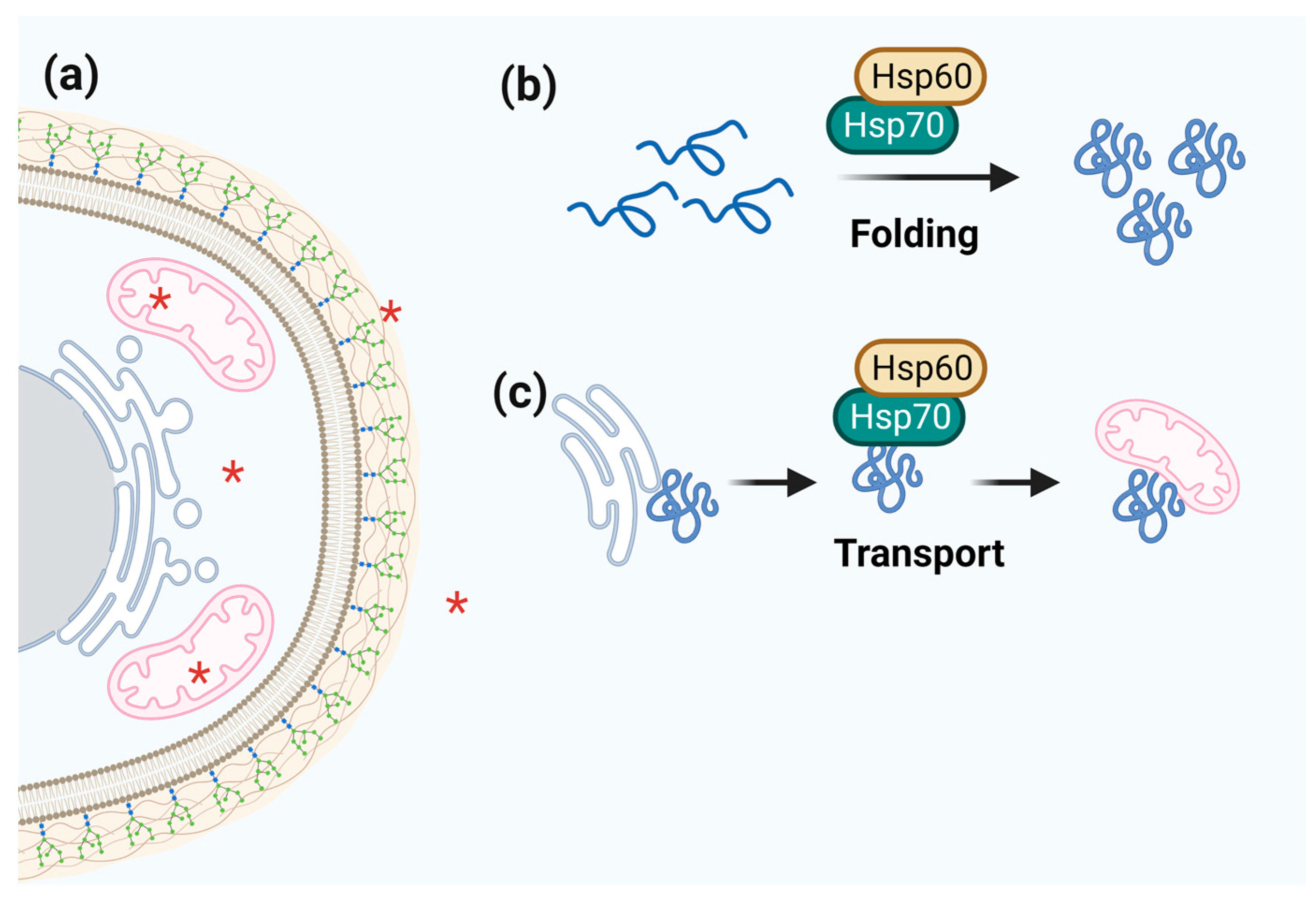
3.3. Hsp60
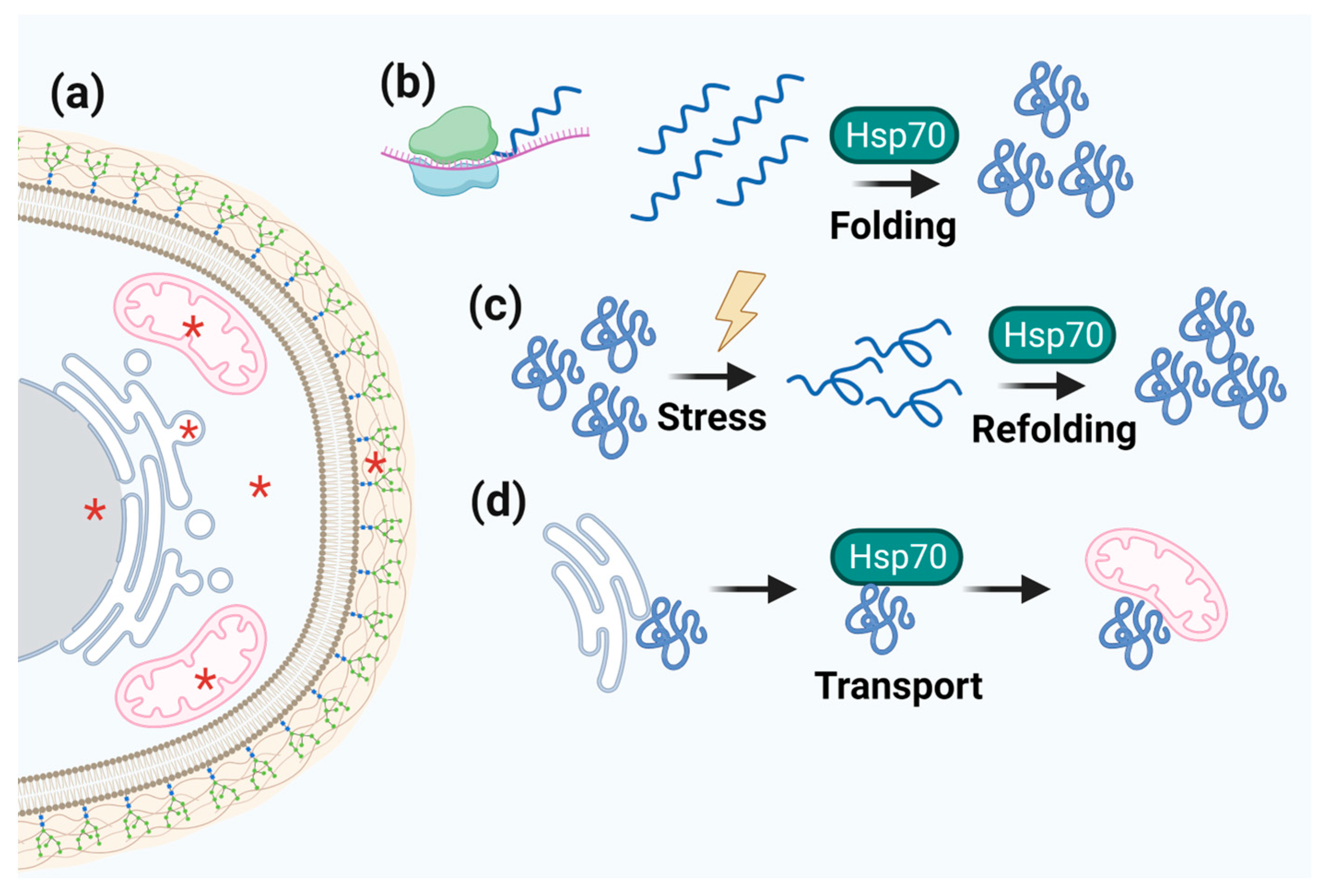
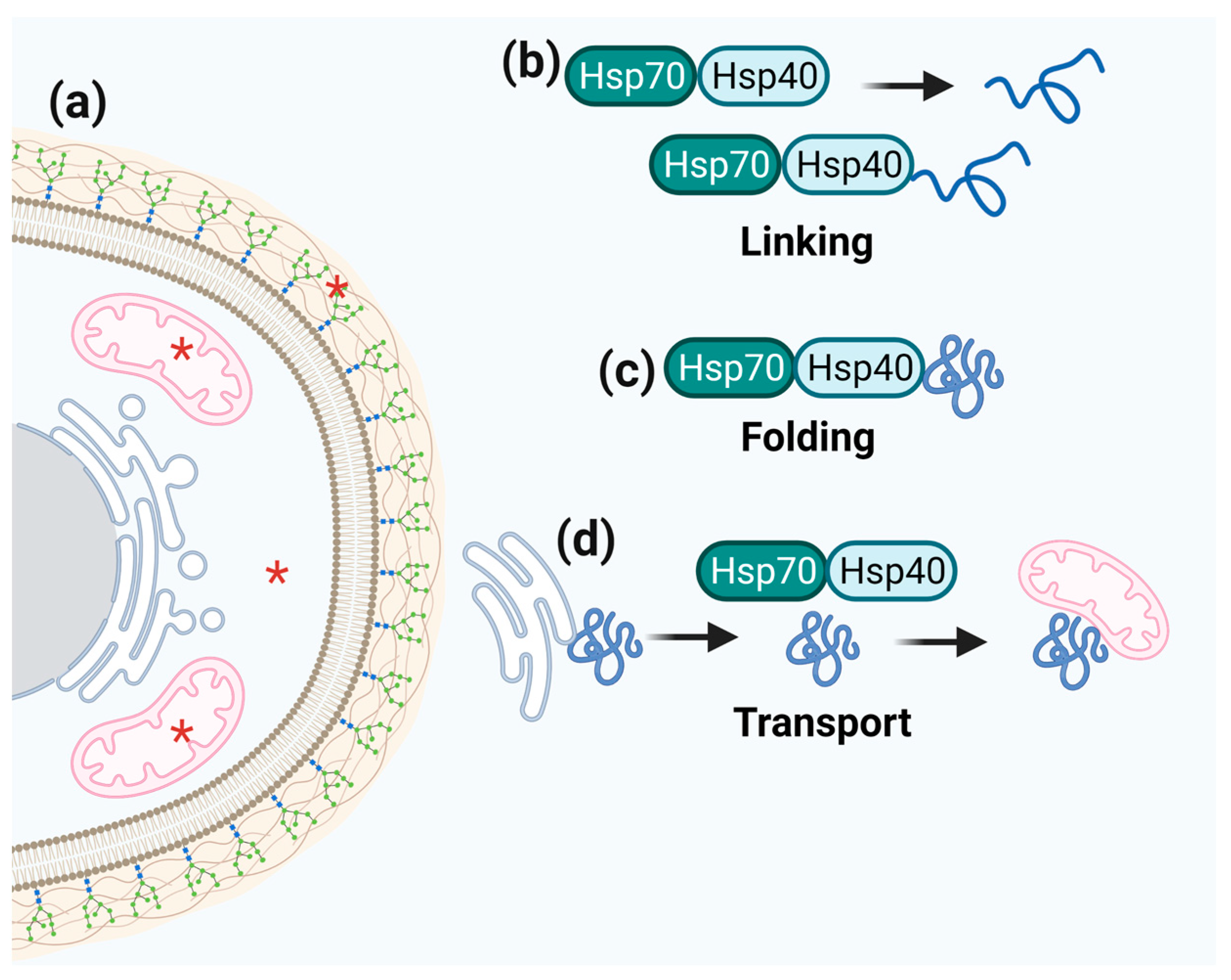
3.4. Hsp40/Hsp70
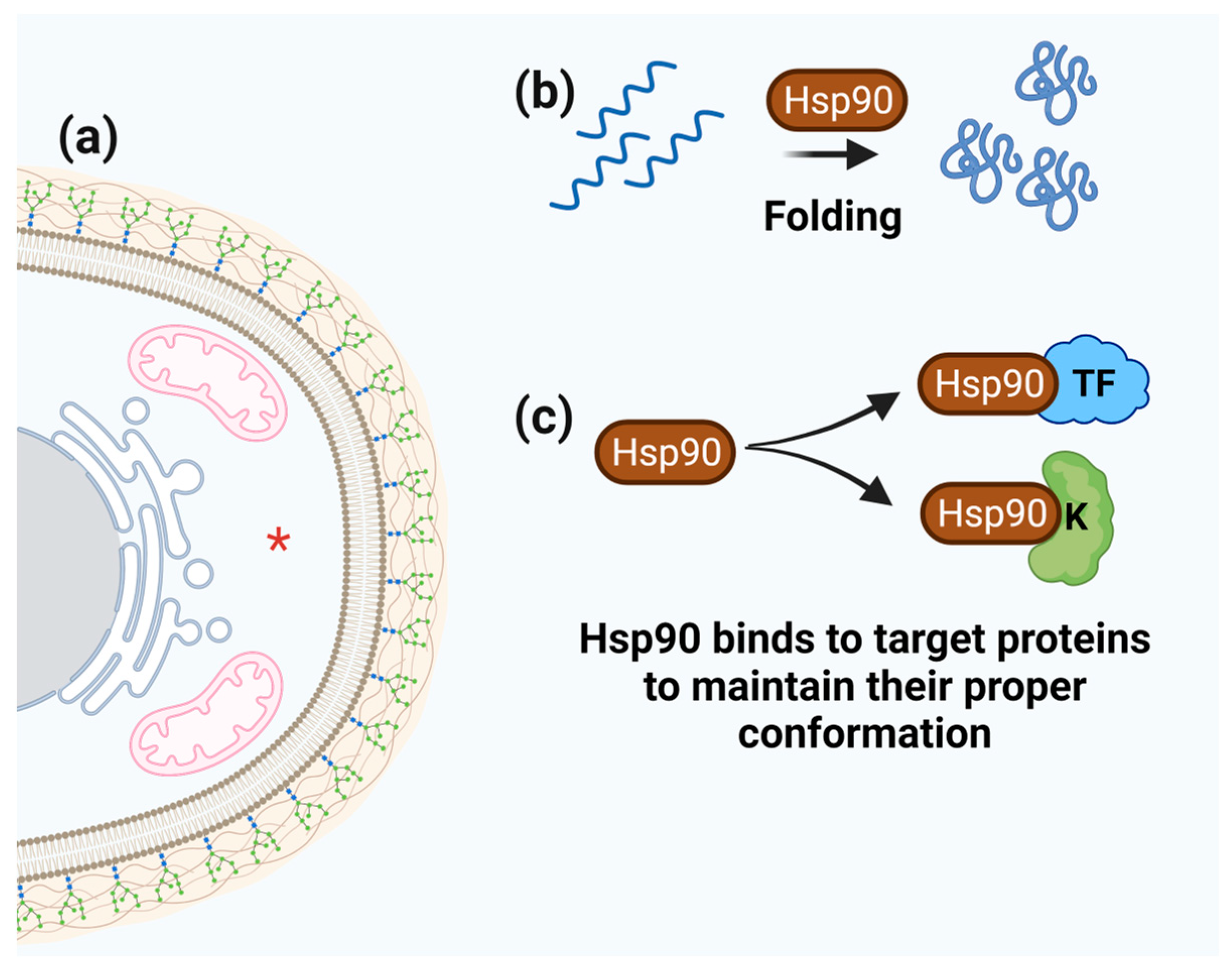
3.5. Hsp90
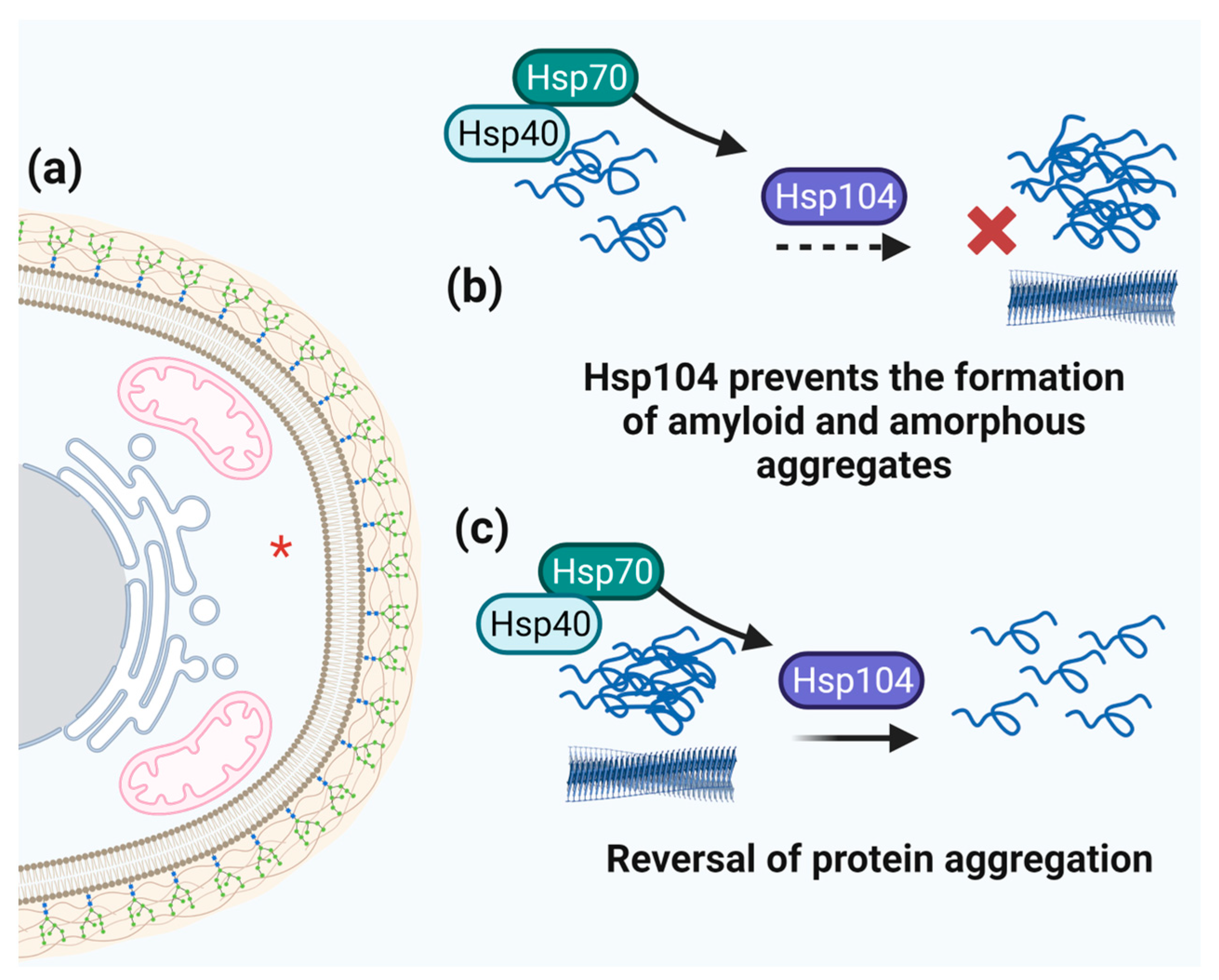
3.6. Hsp104 and Aggregation
4. Antifungal Therapy
5. Conclusions and Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv113. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous Emergence of Multidrug-Resistant Candida auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clin. Infect. Dis. 2017, 64, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Karunarathna, S.C.; Maharachchikumbura, S.S.N.; Ariyawansa, H.A.; Shenoy, B.D.; Jeewon, R. Editorial: Emerging Fungal Plant Pathogens. Front. Cell Infect. Microbiol. 2021, 11, 765549. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E. Fusarium Mycotoxins: Chemistry, Genetics, and Biology; American Phytopathological Society (APS Press): Algiers, Algeria, 2006. [Google Scholar]
- Hossain, S.; Lash, E.; Veri, A.O.; Cowen, L.E. Functional connections between cell cycle and proteostasis in the regulation of Candida albicans morphogenesis. Cell Rep. 2021, 34, 108781. [Google Scholar] [CrossRef]
- Smith, M.H.; Ploegh, H.L.; Weissman, J.S. Road to ruin: Targeting proteins for degradation in the endoplasmic reticulum. Science 2011, 334, 1086–1090. [Google Scholar] [CrossRef]
- Amm, I.; Sommer, T.; Wolf, D.H. Protein quality control and elimination of protein waste: The role of the ubiquitin-proteasome system. Biochim. Biophys. Acta 2014, 1843, 182–196. [Google Scholar] [CrossRef]
- Le Breton, L.; Mayer, M.P. A model for handling cell stress. eLife 2016, 5, e22850. [Google Scholar] [CrossRef]
- Kampinga, H.H.; Craig, E.A. The HSP70 chaperone machinery: J proteins as drivers of functional specificity. Nat. Rev. Mol. Cell Biol. 2010, 11, 579–592. [Google Scholar] [CrossRef]
- Morano, K.A.; Grant, C.M.; Moye-Rowley, W.S. The response to heat shock and oxidative stress in Saccharomyces cerevisiae. Genetics 2012, 190, 1157–1195. [Google Scholar] [CrossRef]
- Jacob, T.R.; Peres, N.T.; Martins, M.P.; Lang, E.A.; Sanches, P.R.; Rossi, A.; Martinez-Rossi, N.M. Heat Shock Protein 90 (Hsp90) as a Molecular Target for the Development of Novel Drugs against the Dermatophyte Trichophyton rubrum. Front. Microbiol. 2015, 6, 1241. [Google Scholar] [CrossRef]
- Ahn, S.G.; Thiele, D.J. Redox regulation of mammalian heat shock factor 1 is essential for Hsp gene activation and protection from stress. Genes. Dev. 2003, 17, 516–528. [Google Scholar] [CrossRef] [PubMed]
- Hahn, J.S.; Neef, D.W.; Thiele, D.J. A stress regulatory network for co-ordinated activation of proteasome expression mediated by yeast heat shock transcription factor. Mol. Microbiol. 2006, 60, 240–251. [Google Scholar] [CrossRef]
- Beekman, C.N.; Ene, I.V. Short-term evolution strategies for host adaptation and drug escape in human fungal pathogens. PLoS Pathog. 2020, 16, e1008519. [Google Scholar] [CrossRef] [PubMed]
- Eastman, A.J.; He, X.; Qiu, Y.; Davis, M.J.; Vedula, P.; Lyons, D.M.; Park, Y.D.; Hardison, S.E.; Malachowski, A.N.; Osterholzer, J.J.; et al. Cryptococcal heat shock protein 70 homolog Ssa1 contributes to pulmonary expansion of Cryptococcus neoformans during the afferent phase of the immune response by promoting macrophage M2 polarization. J. Immunol. 2015, 194, 5999–6010. [Google Scholar] [CrossRef] [PubMed]
- Lamoth, F.; Juvvadi, P.R.; Fortwendel, J.R.; Steinbach, W.J. Heat shock protein 90 is required for conidiation and cell wall integrity in Aspergillus fumigatus. Eukaryot. Cell 2012, 11, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Wirk, B. Heat shock protein inhibitors for the treatment of fungal infections. Recent. Pat. Antiinfect. Drug Discov. 2011, 6, 38–44. [Google Scholar] [CrossRef]
- Köhler, J.R.; Hube, B.; Puccia, R.; Casadevall, A.; Perfect, J.R. Fungi that infect humans. Microbiol. Spectr. 2017, 5, 8. [Google Scholar] [CrossRef]
- Alagar Boopathy, L.R.; Jacob-Tomas, S.; Alecki, C.; Vera, M. Mechanisms tailoring the expression of heat shock proteins to proteostasis challenges. J. Biol. Chem. 2022, 298, 101796. [Google Scholar] [CrossRef]
- Mayer, F.L.; Wilson, D.; Jacobsen, I.D.; Miramón, P.; Slesiona, S.; Bohovych, I.M.; Brown, A.J.; Hube, B. Small but crucial: The novel small heat shock protein Hsp21 mediates stress adaptation and virulence in Candida albicans. PLoS ONE 2012, 7, e38584. [Google Scholar] [CrossRef]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef]
- Brown, A.J.; Budge, S.; Kaloriti, D.; Tillmann, A.; Jacobsen, M.D.; Yin, Z.; Ene, I.V.; Bohovych, I.; Sandai, D.; Kastora, S.; et al. Stress adaptation in a pathogenic fungus. J. Exp. Biol. 2014, 217, 144–155. [Google Scholar] [CrossRef]
- Burnie, J.P.; Carter, T.L.; Hodgetts, S.J.; Matthews, R.C. Fungal heat-shock proteins in human disease. FEMS Microbiol. Rev. 2006, 30, 53–88. [Google Scholar] [CrossRef]
- Ost, K.S.; O’Meara, T.R.; Huda, N.; Esher, S.K.; Alspaugh, J.A. The Cryptococcus neoformans alkaline response pathway: Identification of a novel rim pathway activator. PLoS Genet. 2015, 11, e1005159. [Google Scholar] [CrossRef] [PubMed]
- Esher, S.K.; Zaragoza, O.; Alspaugh, J.A. Cryptococcal pathogenic mechanisms: A dangerous trip from the environment to the brain. Mem. Inst. Oswaldo Cruz 2018, 113, e180057. [Google Scholar] [CrossRef]
- Thompson, S.A. Molecular cloning and deletion of the gene encoding aspergillopepsin A from Aspergillus awamori. Gene 1990, 96, 313. [Google Scholar]
- Mattern, I.E.; van Noort, J.M.; van den Berg, P.; Archer, D.B.; Roberts, I.N.; van den Hondel, C.A. Isolation and characterization of mutants of Aspergillus niger deficient in extracellular proteases. Mol. Gen. Genet. 1992, 234, 332–336. [Google Scholar] [CrossRef]
- Kitano, H.; Kataoka, K.; Furukawa, K.; Hara, S. Specific expression and temperature-dependent expression of the acid protease-encoding gene (pepA) in Aspergillus oryzae in solid-state culture (Rice-Koji). J. Biosci. Bioeng. 2002, 93, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Cruz, A.H.; Santos, R.S.; Silva, P.M.; Silva, E.M.; Mendes, N.S.; Martinez-Rossi, N.M. Ambient pH sensing in filamentous fungi: Pitfalls in elucidating regulatory hierarchical signaling networks. Iubmb Life 2013, 65, 930–935. [Google Scholar] [CrossRef] [PubMed]
- Denison, S.H. pH regulation of gene expression in fungi. Fungal Genet. Biol. 2000, 29, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.; Barata-Antunes, C.; Casal, M.; Brown, A.J.P.; Van Dijck, P.; Paiva, S. Adapting to survive: How Candida overcomes host-imposed constraints during human colonization. PLoS Pathog. 2020, 16, e1008478. [Google Scholar] [CrossRef]
- Brown, S.M.; Campbell, L.T.; Lodge, J.K. Cryptococcus neoformans, a fungus under stress. Curr. Opin. Microbiol. 2007, 10, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.A.; Lu, J.P.; Liu, X.H.; Rehman, A.; Lin, F.C. Multifunction of autophagy-related genes in filamentous fungi. Microbiol. Res. 2012, 167, 339–345. [Google Scholar] [CrossRef]
- Voigt, O.; Pöggeler, S. Self-eating to grow and kill: Autophagy in filamentous ascomycetes. Appl. Microbiol. Biotechnol. 2013, 97, 9277–9290. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.H.; Xu, F.; Snyder, J.H.; Shi, H.B.; Lu, J.P.; Lin, F.C. Autophagy in plant pathogenic fungi. Semin. Cell Dev. Biol. 2016, 57, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.M.; Li, L.; Wu, M.; Liang, S.; Shi, H.B.; Liu, X.H.; Lin, F.C. Current opinions on autophagy in pathogenicity of fungi. Virulence 2019, 10, 481–489. [Google Scholar] [CrossRef]
- Dokladny, K.; Myers, O.B.; Moseley, P.L. Heat shock response and autophagy—Cooperation and control. Autophagy 2015, 11, 200–213. [Google Scholar] [CrossRef]
- Metzenberg, R.L. Implications of some genetic control mechanisms in Neurospora. Microbiol. Rev. 1979, 43, 361–383. [Google Scholar] [CrossRef]
- Peleg, Y.; Addison, R.; Aramayo, R.; Metzenberg, R.L. Translocation of Neurospora crassa transcription factor NUC-1 into the nucleus is induced by phosphorus limitation. Fungal Genet. Biol. 1996, 20, 185–191. [Google Scholar] [CrossRef]
- Gras, D.E.; Persinoti, G.F.; Peres, N.T.; Martinez-Rossi, N.M.; Tahira, A.C.; Reis, E.M.; Prade, R.A.; Rossi, A. Transcriptional profiling of Neurospora crassa Δmak-2 reveals that mitogen-activated protein kinase MAK-2 participates in the phosphate signaling pathway. Fungal Genet. Biol. 2013, 60, 140–149. [Google Scholar] [CrossRef]
- Martinez-Rossi, N.M.; Peres, N.T.; Rossi, A. Pathogenesis of Dermatophytosis: Sensing the Host Tissue. Mycopathologia 2017, 182, 215–227. [Google Scholar] [CrossRef]
- Ferreira-Nozawa, M.S.; Nozawa, S.R.; Martinez-Rossi, N.M.; Rossi, A. The dermatophyte Trichophyton rubrum secretes an EDTA-sensitive alkaline phosphatase on high-phosphate medium. Braz. J. Microbiol. 2003, 34, 161–164. [Google Scholar] [CrossRef]
- Martinez-Rossi, N.M.; Persinoti, G.F.; Peres, N.T.; Rossi, A. Role of pH in the pathogenesis of dermatophytoses. Mycoses 2012, 55, 381–387. [Google Scholar] [CrossRef]
- Gastebois, A.; Clavaud, C.; Aimanianda, V.; Latgé, J.P. Aspergillus fumigatus: Cell wall polysaccharides, their biosynthesis and organization. Future Microbiol. 2009, 4, 583–595. [Google Scholar] [CrossRef] [PubMed]
- Sharafi, G.; Khosravi, A.R.; Vahedi, G.; Yahyaraeyat, R.; Abbasi, T. A comparative study of the timecourse of the expression of the thermo-inducible HSP70 gene in clinical and environmental isolates of Aspergillus fumigatus. Mol. Med. Rep. 2016, 13, 4513–4521. [Google Scholar] [CrossRef] [PubMed]
- Latgé, J.P.; Beauvais, A.; Chamilos, G. The Cell Wall of the Human Fungal Pathogen Aspergillus fumigatus: Biosynthesis, Organization, Immune Response, and Virulence. Annu. Rev. Microbiol. 2017, 71, 99–116. [Google Scholar] [CrossRef]
- van de Veerdonk, F.L.; Gresnigt, M.S.; Romani, L.; Netea, M.G.; Latgé, J.P. Aspergillus fumigatus morphology and dynamic host interactions. Nat. Rev. Microbiol. 2017, 15, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Rocha, M.C.; Minari, K.; Fabri, J.H.T.M.; Kerkaert, J.D.; Gava, L.M.; da Cunha, A.F.; Cramer, R.A.; Borges, J.C.; Malavazi, I. Aspergillus fumigatus Hsp90 interacts with the main components of the cell wall integrity pathway and cooperates in heat shock and cell wall stress adaptation. Cell Microbiol. 2021, 23, e13273. [Google Scholar] [CrossRef]
- Samalova, M.; Carr, P.; Bromley, M.; Blatzer, M.; Moya-Nilges, M.; Latgé, J.P.; Mouyna, I. GPI Anchored Proteins in Aspergillus fumigatus and Cell Wall Morphogenesis. Curr. Top. Microbiol. Immunol. 2020, 425, 167–186. [Google Scholar] [CrossRef]
- Horianopoulos, L.C.; Kronstad, J.W. Chaperone Networks in Fungal Pathogens of Humans. J. Fungi 2021, 7, 209. [Google Scholar] [CrossRef]
- Schrettl, M.; Haas, H. Iron homeostasis—Achilles’ heel of Aspergillus fumigatus? Curr. Opin. Microbiol. 2011, 14, 400–405. [Google Scholar] [CrossRef]
- Misslinger, M.; Hortschansky, P.; Brakhage, A.A.; Haas, H. Fungal iron homeostasis with a focus on Aspergillus fumigatus. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118885. [Google Scholar] [CrossRef]
- Brantl, V.; Boysen, J.M.; Yap, A.; Golubtsov, E.; Ruf, D.; Heinekamp, T.; Straßburger, M.; Dichtl, K.; Haas, H.; Hillmann, F.; et al. Peroxiredoxin Asp f3 Is Essential for Aspergillus fumigatus To Overcome Iron Limitation during Infection. mBio 2021, 12, e0097621. [Google Scholar] [CrossRef]
- Zhai, P.; Shi, L.; Zhong, G.; Jiang, J.; Zhou, J.; Chen, X.; Dong, G.; Zhang, L.; Li, R.; Song, J. The OxrA Protein of Aspergillus fumigatus Is Required for the Oxidative Stress Response and Fungal Pathogenesis. Appl. Environ. Microbiol. 2021, 87, e0112021. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Fan, Y.; Cao, P.; Tan, K. Multifaceted roles of HSF1 in cell death: A state-of-the-art review. Biochim. Biophys. Acta Rev. Cancer 2021, 1876, 188591. [Google Scholar] [CrossRef] [PubMed]
- Fabri, J.H.T.M.; Rocha, M.C.; Fernandes, C.M.; Persinoti, G.F.; Ries, L.N.A.; da Cunha, A.F.; Goldman, G.H.; Del Poeta, M.; Malavazi, I. The Heat Shock Transcription Factor HsfA Is Essential for Thermotolerance and Regulates Cell Wall Integrity in. Front. Microbiol. 2021, 12, 656548. [Google Scholar] [CrossRef] [PubMed]
- Agita, A.; Alsagaff, M.T. Inflammation, Immunity, and Hypertension. Acta Med. Indones. 2017, 49, 158–165. [Google Scholar] [PubMed]
- Zou, X.; Wei, Y.; Jiang, S.; Xu, F.; Wang, H.; Zhan, P.; Shao, X. ROS Stress and Cell Membrane Disruption are the Main Antifungal Mechanisms of 2-Phenylethanol against. J. Agric. Food Chem. 2022, 70, 14468–14479. [Google Scholar] [CrossRef]
- Huarte-Bonnet, C.; Juárez, M.P.; Pedrini, N. Oxidative stress in entomopathogenic fungi grown on insect-like hydrocarbons. Curr. Genet. 2015, 61, 289–297. [Google Scholar] [CrossRef]
- Niforou, K.; Cheimonidou, C.; Trougakos, I.P. Molecular chaperones and proteostasis regulation during redox imbalance. Redox Biol. 2014, 2, 323–332. [Google Scholar] [CrossRef]
- Ikwegbue, P.C.; Masamba, P.; Oyinloye, B.E.; Kappo, A.P. Roles of Heat Shock Proteins in Apoptosis, Oxidative Stress, Human Inflammatory Diseases, and Cancer. Pharmaceuticals 2017, 11, 2. [Google Scholar] [CrossRef]
- Li, X.S.; Sun, J.N.; Okamoto-Shibayama, K.; Edgerton, M. Candida albicans cell wall ssa proteins bind and facilitate import of salivary histatin 5 required for toxicity. J. Biol. Chem. 2006, 281, 22453–22463. [Google Scholar] [CrossRef] [PubMed]
- Nimrichter, L.; De Souza, M.M.; Del Poeta, M.; Nosanchuk, J.D.; Joffe, L.; Tavares, P.D.M.; Rodrigues, M.L. Extracellular Vesicle-Associated Transitory Cell Wall Components and Their Impact on the Interaction of Fungi with Host Cells. Front. Microbiol. 2016, 7, 1034. [Google Scholar] [CrossRef] [PubMed]
- Rollenhagen, C.; Mamtani, S.; Ma, D.; Dixit, R.; Eszterhas, S.; Lee, S.A. The Role of Secretory Pathways in. J. Fungi 2020, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Raggam, R.B.; Salzer, H.J.; Marth, E.; Heiling, B.; Paulitsch, A.H.; Buzina, W. Molecular detection and characterisation of fungal heat shock protein 60. Mycoses 2011, 54, e394–e399. [Google Scholar] [CrossRef]
- Moura, Á.; Oliveira, D.S.L.; Paredes, V.; Rocha, L.B.; Oliveira, F.F.M.; Lessa, G.M.; Riasco-Palacios, J.F.; Casadevall, A.; Albuquerque, P.; Felipe, M.S.S.; et al. Paracoccidioides HSP90 Can Be Found in the Cell Surface and Is a Target for Antibodies with Therapeutic Potential. J. Fungi 2020, 6, 193. [Google Scholar] [CrossRef]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef]
- Cheon, S.A.; Jung, K.W.; Chen, Y.L.; Heitman, J.; Bahn, Y.S.; Kang, H.A. Unique evolution of the UPR pathway with a novel bZIP transcription factor, Hxl1, for controlling pathogenicity of Cryptococcus neoformans. PLoS Pathog. 2011, 7, e1002177. [Google Scholar] [CrossRef]
- Cheon, S.A.; Jung, K.W.; Bahn, Y.S.; Kang, H.A. The unfolded protein response (UPR) pathway in Cryptococcus. Virulence 2014, 5, 341–350. [Google Scholar] [CrossRef]
- Krishnan, K.; Askew, D.S. The fungal UPR: A regulatory hub for virulence traits in the mold pathogen Aspergillus fumigatus. Virulence 2014, 5, 334–340. [Google Scholar] [CrossRef]
- Saloheimo, M.; Valkonen, M.; Penttilä, M. Activation mechanisms of the HAC1-mediated unfolded protein response in filamentous fungi. Mol. Microbiol. 2003, 47, 1149–1161. [Google Scholar] [CrossRef]
- Hernández-Elvira, M.; Torres-Quiroz, F.; Escamilla-Ayala, A.; Domínguez-Martin, E.; Escalante, R.; Kawasaki, L.; Ongay-Larios, L.; Coria, R. The Unfolded Protein Response Pathway in the Yeast. Cells 2018, 7, 106. [Google Scholar] [CrossRef]
- Kimata, Y.; Ishiwata-Kimata, Y.; Ito, T.; Hirata, A.; Suzuki, T.; Oikawa, D.; Takeuchi, M.; Kohno, K. Two regulatory steps of ER-stress sensor Ire1 involving its cluster formation and interaction with unfolded proteins. J. Cell Biol. 2007, 179, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Bitencourt, T.A.; Lang, E.A.; Sanches, P.R.; Peres, N.T.; Oliveira, V.M.; Fachin, A.L.; Rossi, A.; Martinez-Rossi, N.M. HacA Governs Virulence Traits and Adaptive Stress Responses in Trichophyton rubrum. Front. Microbiol. 2020, 11, 193. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Krishnan, K.; Richie, D.L.; Aimanianda, V.; Hartl, L.; Grahl, N.; Powers-Fletcher, M.V.; Zhang, M.; Fuller, K.K.; Nierman, W.C.; et al. HacA-independent functions of the ER stress sensor IreA synergize with the canonical UPR to influence virulence traits in Aspergillus fumigatus. PLoS Pathog. 2011, 7, e1002330. [Google Scholar] [CrossRef] [PubMed]
- Richie, D.L.; Hartl, L.; Aimanianda, V.; Winters, M.S.; Fuller, K.K.; Miley, M.D.; White, S.; McCarthy, J.W.; Latgé, J.P.; Feldmesser, M.; et al. A role for the unfolded protein response (UPR) in virulence and antifungal susceptibility in Aspergillus fumigatus. PLoS Pathog. 2009, 5, e1000258. [Google Scholar] [CrossRef] [PubMed]
- Haynes, C.M.; Titus, E.A.; Cooper, A.A. Degradation of misfolded proteins prevents ER-derived oxidative stress and cell death. Mol. Cell 2004, 15, 767–776. [Google Scholar] [CrossRef]
- Hirsch, C.; Gauss, R.; Horn, S.C.; Neuber, O.; Sommer, T. The ubiquitylation machinery of the endoplasmic reticulum. Nature 2009, 458, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Pisa, R.; Rapoport, T.A. Disulfide-crosslink analysis of the ubiquitin ligase Hrd1 complex during endoplasmic reticulum-associated protein degradation. J. Biol. Chem. 2022, 298, 102373. [Google Scholar] [CrossRef]
- Kästle, M.; Grune, T. Interactions of the proteasomal system with chaperones: Protein triage and protein quality control. Prog. Mol. Biol. Transl. Sci. 2012, 109, 113–160. [Google Scholar] [CrossRef] [PubMed]
- Fachin, A.L.; Petrucelli, M.F.; Bitencourt, T.A.; Nishimura, F.G.; Segura, G.G.; Cantelli, B.A.M.; de Abreu, M.H.; Marins, M. Transcriptome of host–dermatophyte interactions using infection models. In Dermatophytes and Dermatophytoses; Springer: Berlin/Heidelberg, Germany, 2021; pp. 161–179. [Google Scholar]
- Wimalasena, T.T.; Enjalbert, B.; Guillemette, T.; Plumridge, A.; Budge, S.; Yin, Z.; Brown, A.J.; Archer, D.B. Impact of the unfolded protein response upon genome-wide expression patterns, and the role of Hac1 in the polarized growth, of Candida albicans. Fungal Genet. Biol. 2008, 45, 1235–1247. [Google Scholar] [CrossRef]
- Cox, J.S.; Shamu, C.E.; Walter, P. Transcriptional induction of genes encoding endoplasmic reticulum resident proteins requires a transmembrane protein kinase. Cell 1993, 73, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Altenhoff, A.M.; Glover, N.M.; Dessimoz, C. Inferring Orthology and Paralogy. Methods Mol. Biol. 2019, 1910, 149–175. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V. Orthologs, paralogs, and evolutionary genomics. Annu. Rev. Genet. 2005, 39, 309–338. [Google Scholar] [CrossRef] [PubMed]
- da Silva, S.P.; Borges-Walmsley, M.I.; Pereira, I.S.; Soares, C.M.; Walmsley, A.R.; Felipe, M.S. Differential expression of an hsp70 gene during transition from the mycelial to the infective yeast form of the human pathogenic fungus Paracoccidioides brasiliensis. Mol. Microbiol. 1999, 31, 1039–1050. [Google Scholar] [CrossRef]
- Sun, J.N.; Solis, N.V.; Phan, Q.T.; Bajwa, J.S.; Kashleva, H.; Thompson, A.; Liu, Y.; Dongari-Bagtzoglou, A.; Edgerton, M.; Filler, S.G. Host cell invasion and virulence mediated by Candida albicans Ssa1. PLoS Pathog. 2010, 6, e1001181. [Google Scholar] [CrossRef]
- Li, X.S.; Reddy, M.S.; Baev, D.; Edgerton, M. Candida albicans Ssa1/2p is the cell envelope binding protein for human salivary histatin 5. J. Biol. Chem. 2003, 278, 28553–28561. [Google Scholar] [CrossRef]
- Neves-da-Rocha, J.; Bitencourt, T.A.; Oliveira, V.M.; Sanches, P.R.; Rossi, A.; Martinez-Rossi, N.M. Alternative Splicing in Heat Shock Protein Transcripts as a Mechanism of Cell Adaptation in Trichophyton rubrum. Cells 2019, 8, 1206. [Google Scholar] [CrossRef]
- Seidl, V.; Song, L.; Lindquist, E.; Gruber, S.; Koptchinskiy, A.; Zeilinger, S.; Schmoll, M.; Martínez, P.; Sun, J.; Grigoriev, I.; et al. Transcriptomic response of the mycoparasitic fungus Trichoderma atroviride to the presence of a fungal prey. BMC Genom. 2009, 10, 567. [Google Scholar] [CrossRef]
- Mota, T.M.; Oshiquiri, L.H.; Lopes, É.; Barbosa Filho, J.R.; Ulhoa, C.J.; Georg, R.C. Hsp genes are differentially expressed during Trichoderma asperellum self-recognition, mycoparasitism and thermal stress. Microbiol. Res. 2019, 227, 126296. [Google Scholar] [CrossRef]
- Avrova, A.O.; Venter, E.; Birch, P.R.; Whisson, S.C. Profiling and quantifying differential gene transcription in Phytophthora infestans prior to and during the early stages of potato infection. Fungal Genet. Biol. 2003, 40, 4–14. [Google Scholar] [CrossRef]
- Brunt, S.A.; Borkar, M.; Silver, J.C. Regulation of hsp90 and hsp70 genes during antheridiol-induced hyphal branching in the oomycete Achlya ambisexualis. Fungal Genet. Biol. 1998, 24, 310–324. [Google Scholar] [CrossRef] [PubMed]
- Janowska, M.K.; Baughman, H.E.R.; Woods, C.N.; Klevit, R.E. Mechanisms of Small Heat Shock Proteins. Cold Spring Harb. Perspect. Biol. 2019, 11, a034025. [Google Scholar] [CrossRef]
- Sun, Y.; MacRae, T.H. Small heat shock proteins: Molecular structure and chaperone function. Cell Mol. Life Sci. 2005, 62, 2460–2476. [Google Scholar] [CrossRef] [PubMed]
- Bakthisaran, R.; Tangirala, R.; Rao, C.M. Small heat shock proteins: Role in cellular functions and pathology. Biochim. Biophys. Acta 2015, 1854, 291–319. [Google Scholar] [CrossRef] [PubMed]
- van Montfort, R.L.; Basha, E.; Friedrich, K.L.; Slingsby, C.; Vierling, E. Crystal structure and assembly of a eukaryotic small heat shock protein. Nat. Struct. Biol. 2001, 8, 1025–1030. [Google Scholar] [CrossRef]
- Kampinga, H.H.; Garrido, C. HSPBs: Small proteins with big implications in human disease. Int. J. Biochem. Cell Biol. 2012, 44, 1706–1710. [Google Scholar] [CrossRef]
- Wu, J.; Wang, M.; Zhou, L.; Yu, D. Small heat shock proteins, phylogeny in filamentous fungi and expression analyses in Aspergillus nidulans. Gene 2016, 575, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A. Small heat shock proteins (HSP12, HSP20 and HSP30) play a role in Ustilago maydis pathogenesis. FEMS Microbiol. Lett. 2014, 361, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Motta, H.; Catarina Vieira Reuwsaat, J.; Daidrê Squizani, E.; da Silva Camargo, M.; Wichine Acosta Garcia, A.; Schrank, A.; Henning Vainstein, M.; Christian Staats, C.; Kmetzsch, L. The small heat shock protein Hsp12.1 has a major role in the stress response and virulence of Cryptococcus gattii. Fungal Genet. Biol. 2023, 165, 103780. [Google Scholar] [CrossRef]
- Taricani, L.; Feilotter, H.E.; Weaver, C.; Young, P.G. Expression of hsp16 in response to nucleotide depletion is regulated via the spc1 MAPK pathway in Schizosaccharomyces pombe. Nucleic Acids Res. 2001, 29, 3030–3040. [Google Scholar] [CrossRef]
- Glatz, A.; Pilbat, A.M.; Németh, G.L.; Vince-Kontár, K.; Jósvay, K.; Hunya, Á.; Udvardy, A.; Gombos, I.; Péter, M.; Balogh, G.; et al. Involvement of small heat shock proteins, trehalose, and lipids in the thermal stress management in Schizosaccharomyces pombe. Cell Stress. Chaperones 2016, 21, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Thakur, R.; Shankar, J. Role of Heat-Shock Proteins in Cellular Function and in the Biology of Fungi. Biotechnol. Res. Int. 2015, 2015, 132635. [Google Scholar] [CrossRef]
- Ajdidi, A.; Sheehan, G.; Kavanagh, K. Exposure of Aspergillus fumigatus to Atorvastatin Leads to Altered Membrane Permeability and Induction of an Oxidative Stress Response. J. Fungi 2020, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Piper, P.W.; Ortiz-Calderon, C.; Holyoak, C.; Coote, P.; Cole, M. Hsp30, the integral plasma membrane heat shock protein of Saccharomyces cerevisiae, is a stress-inducible regulator of plasma membrane H(+)-ATPase. Cell Stress. Chaperones 1997, 2, 12–24. [Google Scholar] [CrossRef]
- Kamo, K.; Takabatake, A.; Inoue, Y.; Izawa, S. Temperature dependent N-glycosylation of plasma membrane heat shock protein Hsp30p in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2012, 420, 119–123. [Google Scholar] [CrossRef]
- Miura, T.; Minegishi, H.; Usami, R.; Abe, F. Systematic analysis of HSP gene expression and effects on cell growth and survival at high hydrostatic pressure in Saccharomyces cerevisiae. Extremophiles 2006, 10, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Behzadi, E.; Behzadi, P.; Sirmatel, F. Identification of 30-kDa heat shock protein gene in Trichophyton rubrum. Mycoses 2009, 52, 234–238. [Google Scholar] [CrossRef]
- Maranhão, F.C.; Paião, F.G.; Martinez-Rossi, N.M. Isolation of transcripts over-expressed in human pathogen Trichophyton rubrum during growth in keratin. Microb. Pathog. 2007, 43, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Rossi, N.M.; Peres, N.T.A.; Bitencourt, T.A.; Martins, M.P.; Rossi, A. State-of-the-Art Dermatophyte Infections: Epidemiology Aspects, Pathophysiology, and Resistance Mechanisms. J. Fungi 2021, 7, 629. [Google Scholar] [CrossRef]
- Borges, C.L.; Bailão, A.M.; Báo, S.N.; Pereira, M.; Parente, J.A.; de Almeida Soares, C.M. Genes potentially relevant in the parasitic phase of the fungal pathogen Paracoccidioides brasiliensis. Mycopathologia 2011, 171, 1–9. [Google Scholar] [CrossRef]
- Plesofsky-Vig, N.; Brambl, R. Heat shock response of Neurospora crassa: Protein synthesis and induced thermotolerance. J. Bacteriol. 1985, 162, 1083–1091. [Google Scholar] [CrossRef]
- Bernardo, R.T.; Cunha, D.V.; Wang, C.; Pereira, L.; Silva, S.; Salazar, S.B.; Schröder, M.S.; Okamoto, M.; Takahashi-Nakaguchi, A.; Chibana, H.; et al. The CgHaa1-Regulon Mediates Response and Tolerance to Acetic Acid Stress in the Human Pathogen Candida glabrata. G3 Genes Genomes Genet. 2017, 7, 1–18. [Google Scholar] [CrossRef]
- Vanittanakom, N.; Pongpom, M.; Praparattanapan, J.; Cooper, C.R.; Sirisanthana, T. Isolation and expression of heat shock protein 30 gene from Penicillium marneffei. Sabouraudia 2009, 47, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Freitas, J.S.; Silva, E.M.; Leal, J.; Gras, D.E.; Martinez-Rossi, N.M.; Dos Santos, L.D.; Palma, M.S.; Rossi, A. Transcription of the Hsp30, Hsp70, and Hsp90 heat shock protein genes is modulated by the PalA protein in response to acid pH-sensing in the fungus Aspergillus nidulans. Cell Stress. Chaperones 2011, 16, 565–572. [Google Scholar] [CrossRef]
- Long, K.H.; Gomez, F.J.; Morris, R.E.; Newman, S.L. Identification of heat shock protein 60 as the ligand on Histoplasma capsulatum that mediates binding to CD18 receptors on human macrophages. J. Immunol. 2003, 170, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, F.F.; Pitangui, N.S.; Peron, G.; Valim, C.X.R.; Mazucato, V.M.; Oliveira, A.F.; Silva, T.A.D.; Roque-Barreira, M.C.; Panunto-Castelo, A. Expression of Hsp60 and its cell location in Paracoccidioides brasiliensis. Rev. Inst. Med. Trop. Sao Paulo 2020, 62, e29. [Google Scholar] [CrossRef] [PubMed]
- Cleare, L.G.; Zamith-Miranda, D.; Nosanchuk, J.D. Heat Shock Proteins in Histoplasma and Paracoccidioides. Clin. Vaccine Immunol. 2017, 24, e00221-17. [Google Scholar] [CrossRef] [PubMed]
- Böttinger, L.; Oeljeklaus, S.; Guiard, B.; Rospert, S.; Warscheid, B.; Becker, T. Mitochondrial heat shock protein (Hsp) 70 and Hsp10 cooperate in the formation of Hsp60 complexes. J. Biol. Chem. 2015, 290, 11611–11622. [Google Scholar] [CrossRef]
- García-Carnero, L.C.; Salinas-Marín, R.; Lozoya-Pérez, N.E.; Wrobel, K.; Wrobel, K.; Martínez-Duncker, I.; Niño-Vega, G.A.; Mora-Montes, H.M. The heat shock protein 60 and Pap1 participate in the Sporothrix schenckii-host interaction. J. Fungi 2021, 7, 960. [Google Scholar] [CrossRef]
- Verghese, J.; Abrams, J.; Wang, Y.; Morano, K.A. Biology of the heat shock response and protein chaperones: Budding yeast (Saccharomyces cerevisiae) as a model system. Microbiol. Mol. Biol. Rev. 2012, 76, 115–158. [Google Scholar] [CrossRef]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef]
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 chaperone network. Nat. Rev. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef] [PubMed]
- Keri, G.; Toth, I. Molecular Pathomechanisms and New Trends in Drug Research; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Faust, O.; Abayev-Avraham, M.; Wentink, A.S.; Maurer, M.; Nillegoda, N.B.; London, N.; Bukau, B.; Rosenzweig, R. HSP40 proteins use class-specific regulation to drive HSP70 functional diversity. Nature 2020, 587, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Shankar, J. Hsp70 in fungi: Evolution, function and vaccine candidate. HSP70 Human. Dis. Disord. 2018, 14, 381–400. [Google Scholar]
- Roy, A.; Tamuli, R. Heat shock proteins and the calcineurin-crz1 signaling regulate stress responses in fungi. Arch. Microbiol. 2022, 204, 240. [Google Scholar] [CrossRef] [PubMed]
- Steen, B.R.; Zuyderduyn, S.; Toffaletti, D.L.; Marra, M.; Jones, S.J.; Perfect, J.R.; Kronstad, J. Cryptococcus neoformans gene expression during experimental cryptococcal meningitis. Eukaryot. Cell 2003, 2, 1336–1349. [Google Scholar] [CrossRef]
- Yang, J.; Liu, M.; Liu, X.; Yin, Z.; Sun, Y.; Zhang, H.; Zheng, X.; Wang, P.; Zhang, Z. Heat-Shock Proteins MoSsb1, MoSsz1, and MoZuo1 Attenuate MoMkk1-Mediated Cell-Wall Integrity Signaling and Are Important for Growth and Pathogenicity of Magnaporthe oryzae. Mol. Plant Microbe Interact. 2018, 31, 1211–1221. [Google Scholar] [CrossRef]
- Dourado, I.B.; Batista, W.L.; Longo, L.V.; Mortara, R.A.; Puccia, R. Dual localization of Mdj1 in pathogenic fungi varies with growth temperature. Med. Mycol. 2014, 52, 187–195. [Google Scholar] [CrossRef]
- Batista, W.L.; Matsuo, A.L.; Ganiko, L.; Barros, T.F.; Veiga, T.R.; Freymüller, E.; Puccia, R. The Pb MDJ1 gene belongs to a conserved MDJ1/LON locus in thermodimorphic pathogenic fungi and encodes a heat shock protein that localizes to both the mitochondria and cell wall of Paracoccidioides brasiliensis. Eukaryot. Cell 2006, 5, 379–390. [Google Scholar] [CrossRef]
- Rezaie, S.; Ban, J.; Mildner, M.; Poitschek, C.; Brna, C.; Tschachler, E. Characterization of a cDNA clone, encoding a 70 kDa heat shock protein from the dermatophyte pathogen Trichophyton rubrum. Gene 2000, 241, 27–33. [Google Scholar] [CrossRef]
- Martinez-Rossi, N.M.; Bitencourt, T.A.; Peres, N.T.A.; Lang, E.A.S.; Gomes, E.V.; Quaresemin, N.R.; Martins, M.P.; Lopes, L.; Rossi, A. Dermatophyte Resistance to Antifungal Drugs: Mechanisms and Prospectus. Front. Microbiol. 2018, 9, 1108. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Zhang, W.; Wang, L.; Yang, J.; Liu, T.; Peng, J.; Leng, W.; Chen, L.; Li, R.; Jin, Q. Transcriptional profiles of the response to ketoconazole and amphotericin B in Trichophyton rubrum. Antimicrob. Agents Chemother. 2007, 51, 144–153. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cowen, L.E.; Lindquist, S. Hsp90 potentiates the rapid evolution of new traits: Drug resistance in diverse fungi. Science 2005, 309, 2185–2189. [Google Scholar] [CrossRef]
- Schopf, F.H.; Biebl, M.M.; Buchner, J. The HSP90 chaperone machinery. Nat. Rev. Mol. Cell Biol. 2017, 18, 345–360. [Google Scholar] [CrossRef]
- Shapiro, R.S.; Uppuluri, P.; Zaas, A.K.; Collins, C.; Senn, H.; Perfect, J.R.; Heitman, J.; Cowen, L.E. Hsp90 orchestrates temperature-dependent Candida albicans morphogenesis via Ras1-PKA signaling. Curr. Biol. 2009, 19, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Senn, H.; Shapiro, R.S.; Cowen, L.E. Cdc28 provides a molecular link between Hsp90, morphogenesis, and cell cycle progression in Candida albicans. Mol. Biol. Cell 2012, 23, 268–283. [Google Scholar] [CrossRef]
- Mishra, S.; Singh, S.; Misra, K. Restraining Pathogenicity in Candida albicans by Taxifolin as an Inhibitor of Ras1-pka Pathway. Mycopathologia 2017, 182, 953–965. [Google Scholar] [CrossRef]
- Diezmann, S.; Leach, M.D.; Cowen, L.E. Functional Divergence of Hsp90 Genetic Interactions in Biofilm and Planktonic Cellular States. PLoS ONE 2015, 10, e0137947. [Google Scholar] [CrossRef]
- Robbins, N.; Uppuluri, P.; Nett, J.; Rajendran, R.; Ramage, G.; Lopez-Ribot, J.L.; Andes, D.; Cowen, L.E. Hsp90 governs dispersion and drug resistance of fungal biofilms. PLoS Pathog. 2011, 7, e1002257. [Google Scholar] [CrossRef]
- Becherelli, M.; Tao, J.; Ryder, N.S. Involvement of heat shock proteins in Candida albicans biofilm formation. J. Mol. Microbiol. Biotechnol. 2013, 23, 396–400. [Google Scholar] [CrossRef]
- Cordeiro, R.A.; Evangelista, A.J.J.; Serpa, R.; Marques, F.J.F.; Melo, C.V.S.; Oliveira, J.S.; Franco, J.D.S.; Alencar, L.P.; Bandeira, T.J.P.G.; Brilhante, R.S.N.; et al. Inhibition of heat-shock protein 90 enhances the susceptibility to antifungals and reduces the virulence of Cryptococcus neoformans/Cryptococcus gattii species complex. Microbiology 2016, 162, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Tatu, U. Heat shock protein 90 localizes to the surface and augments virulence factors of Cryptococcus neoformans. PLoS Negl. Trop. Dis. 2017, 11, e0005836. [Google Scholar] [CrossRef] [PubMed]
- O’Meara, T.R.; Cowen, L.E. Hsp90-dependent regulatory circuitry controlling temperature-dependent fungal development and virulence. Cell Microbiol. 2014, 16, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Crunden, J.L.; Diezmann, S. Hsp90 interaction networks in fungi-tools and techniques. FEMS Yeast Res. 2021, 21, foab054. [Google Scholar] [CrossRef]
- Fu, C.; Beattie, S.R.; Jezewski, A.J.; Robbins, N.; Whitesell, L.; Krysan, D.J.; Cowen, L.E. Genetic analysis of Hsp90 function in Cryptococcus neoformans highlights key roles in stress tolerance and virulence. Genetics 2022, 220, iyab164. [Google Scholar] [CrossRef]
- Leach, M.D.; Klipp, E.; Cowen, L.E.; Brown, A.J. Fungal Hsp90: A biological transistor that tunes cellular outputs to thermal inputs. Nat. Rev. Microbiol. 2012, 10, 693–704. [Google Scholar] [CrossRef]
- Lachowiec, J.; Lemus, T.; Borenstein, E.; Queitsch, C. Hsp90 promotes kinase evolution. Mol. Biol. Evol. 2015, 32, 91–99. [Google Scholar] [CrossRef]
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nature 2011, 475, 324–332. [Google Scholar] [CrossRef]
- Raskatov, J.A.; Teplow, D.B. Using chirality to probe the conformational dynamics and assembly of intrinsically disordered amyloid proteins. Sci. Rep. 2017, 7, 12433. [Google Scholar] [CrossRef]
- Cox, D.; Raeburn, C.; Sui, X.; Hatters, D.M. Protein aggregation in cell biology: An aggregomics perspective of health and disease. Semin. Cell Dev. Biol. 2020, 99, 40–54. [Google Scholar] [CrossRef]
- Doyle, S.M.; Wickner, S. Hsp104 and ClpB: Protein disaggregating machines. Trends Biochem. Sci. 2009, 34, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Bukau, B.; Weissman, J.; Horwich, A. Molecular chaperones and protein quality control. Cell 2006, 125, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Glover, J.R.; Lindquist, S. Hsp104, Hsp70, and Hsp40: A novel chaperone system that rescues previously aggregated proteins. Cell 1998, 94, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.M.; Egan, M.J. Autophagy machinery promotes the chaperone-mediated formation and compartmentalization of protein aggregates during appressorium development by the rice blast fungus. Mol. Biol. Cell 2020, 31, 2298–2305. [Google Scholar] [CrossRef]
- Xia, H.; Chen, L.; Fan, Z.; Peng, M.; Zhao, J.; Chen, W.; Li, H.; Shi, Y.; Ding, S. Heat Stress Tolerance Gene. Front. Microbiol. 2021, 12, 695535. [Google Scholar] [CrossRef]
- Chen, Y.R.; Ziv, I.; Swaminathan, K.; Elias, J.E.; Jarosz, D.F. Protein aggregation and the evolution of stress resistance in clinical yeast. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2021, 376, 20200127. [Google Scholar] [CrossRef]
- Iñigo, M.; Del Pozo, J.L. Fungal biofilms: From bench to bedside. Rev. Esp. Quimioter. 2018, 31 (Suppl. S1), 35–38. [Google Scholar]
- Lagree, K.; Mitchell, A.P. Fungal Biofilms: Inside out. Microbiol. Spectr. 2017, 5, 10–1128. [Google Scholar] [CrossRef]
- Mourer, T.; El Ghalid, M.; Pehau-Arnaudet, G.; Kauffmann, B.; Loquet, A.; Brûlé, S.; Cabral, V.; d’Enfert, C.; Bachellier-Bassi, S. The Pga59 cell wall protein is an amyloid forming protein involved in adhesion and biofilm establishment in the pathogenic yeast Candida albicans. NPJ Biofilms Microbiomes 2023, 9, 6. [Google Scholar] [CrossRef]
- Cao, Y.Y.; Cao, Y.B.; Xu, Z.; Ying, K.; Li, Y.; Xie, Y.; Zhu, Z.Y.; Chen, W.S.; Jiang, Y.Y. cDNA microarray analysis of differential gene expression in Candida albicans biofilm exposed to farnesol. Antimicrob. Agents Chemother. 2005, 49, 584–589. [Google Scholar] [CrossRef]
- Fiori, A.; Kucharíková, S.; Govaert, G.; Cammue, B.P.; Thevissen, K.; Van Dijck, P. The heat-induced molecular disaggregase Hsp104 of Candida albicans plays a role in biofilm formation and pathogenicity in a worm infection model. Eukaryot. Cell 2012, 11, 1012–1020. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Upadhyay, A.K.; Bhat, M.K. Inhibition of Hsp27 and Hsp40 potentiates 5-fluorouracil and carboplatin mediated cell killing in hepatoma cells. Cancer Biol. Ther. 2009, 8, 2106–2113. [Google Scholar] [CrossRef] [PubMed]
- Cappello, F.; Conway de Macario, E.; Marasà, L.; Zummo, G.; Macario, A.J. Hsp60 expression, new locations, functions and perspectives for cancer diagnosis and therapy. Cancer Biol. Ther. 2008, 7, 801–809. [Google Scholar] [CrossRef]
- Nitzsche, B.; Höpfner, M.; Biersack, B. Synthetic Small Molecule Modulators of Hsp70 and Hsp40 Chaperones as Promising Anticancer Agents. Int. J. Mol. Sci. 2023, 24, 4083. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Park, S.; Shin, I. Synthesis of an Hsp70 inhibitor and its assessment of lysosomal membrane permeabilization. STAR Protoc. 2021, 2, 100349. [Google Scholar] [CrossRef] [PubMed]
- Whitesell, L.; Lindquist, S.L. HSP90 and the chaperoning of cancer. Nat. Rev. Cancer 2005, 5, 761–772. [Google Scholar] [CrossRef]
- Samanta, S.; Yang, S.; Debnath, B.; Xue, D.; Kuang, Y.; Ramkumar, K.; Lee, A.S.; Ljungman, M.; Neamati, N. The Hydroxyquinoline Analogue YUM70 Inhibits GRP78 to Induce ER Stress-Mediated Apoptosis in Pancreatic Cancer. Cancer Res. 2021, 81, 1883–1895. [Google Scholar] [CrossRef]
- Martin, S.; Lamb, H.K.; Brady, C.; Lefkove, B.; Bonner, M.Y.; Thompson, P.; Lovat, P.E.; Arbiser, J.L.; Hawkins, A.R.; Redfern, C.P. Inducing apoptosis of cancer cells using small-molecule plant compounds that bind to GRP78. Br. J. Cancer 2013, 109, 433–443. [Google Scholar] [CrossRef]
- Raymundo, D.P.; Doultsinos, D.; Guillory, X.; Carlesso, A.; Eriksson, L.A.; Chevet, E. Pharmacological Targeting of IRE1 in Cancer. Trends Cancer 2020, 6, 1018–1030. [Google Scholar] [CrossRef]
- Du, R.; Sullivan, D.K.; Azizian, N.G.; Liu, Y.; Li, Y. Inhibition of ERAD synergizes with FTS to eradicate pancreatic cancer cells. BMC Cancer 2021, 21, 237. [Google Scholar] [CrossRef]
- Wójcik, C.; DeMartino, G.N. Intracellular localization of proteasomes. Int. J. Biochem. Cell Biol. 2003, 35, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Fricker, L.D. Proteasome inhibitor drugs. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 457–476. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.-B.; Xue, C. The ubiquitin-proteasome system and F-box proteins in pathogenic fungi. Mycobiology 2011, 39, 243–248. [Google Scholar] [CrossRef]
- Sharma, C.; Seo, Y.H. Small Molecule Inhibitors of HSF1-Activated Pathways as Potential Next-Generation Anticancer Therapeutics. Molecules 2018, 23, 2757. [Google Scholar] [CrossRef]
- Demirovic, D.; de Toda, I.M.; Nizard, C.; Rattan, S.I. Differential translocation of heat shock factor-1 after mild and severe stress to human skin fibroblasts undergoing aging in vitro. J. Cell Commun. Signal 2014, 8, 333–339. [Google Scholar] [CrossRef]
- Pincus, D. Regulation of Hsf1 and the heat shock response. In HSF1 and Molecular Chaperones in Biology and Cancer; Springer: Berlin/Heidelberg, Germany, 2020; pp. 41–50. [Google Scholar]
- Behl, A.; Mishra, P.C. Structural insights into the binding mechanism of Plasmodium falciparum exported Hsp40-Hsp70 chaperone pair. Comput. Biol. Chem. 2019, 83, 107099. [Google Scholar] [CrossRef]
- Blatzer, M.; Blum, G.; Jukic, E.; Posch, W.; Gruber, P.; Nagl, M.; Binder, U.; Maurer, E.; Sarg, B.; Lindner, H.; et al. Blocking Hsp70 enhances the efficiency of amphotericin B treatment against resistant Aspergillus terreus strains. Antimicrob. Agents Chemother. 2015, 59, 3778–3788. [Google Scholar] [CrossRef] [PubMed]
- Kartsonis, N.A.; Nielsen, J.; Douglas, C.M. Caspofungin: The first in a new class of antifungal agents. Drug Resist. Updat. 2003, 6, 197–218. [Google Scholar] [CrossRef]
- Kanafani, Z.A.; Perfect, J.R. Resistance to antifungal agents: Mechanisms and clinical impact. Clin. Infect. Dis. 2008, 46, 120–128. [Google Scholar] [CrossRef]
- Odom, A.; Del Poeta, M.; Perfect, J.; Heitman, J. The immunosuppressant FK506 and its nonimmunosuppressive analog L-685,818 are toxic to Cryptococcus neoformans by inhibition of a common target protein. Antimicrob. Agents Chemother. 1997, 41, 156–161. [Google Scholar] [CrossRef]
- de Andrade, I.B.; Corrêa-Junior, D.; Alves, V.; Figueiredo-Carvalho, M.H.G.; Santos, M.V.; Almeida, M.A.; Valdez, A.F.; Nimrichter, L.; Almeida-Paes, R.; Frases, S. Cyclosporine Affects the Main Virulence Factors of. J. Fungi 2023, 9, 487. [Google Scholar] [CrossRef]
- Rossato, L.; Venturini, T.P.; de Azevedo, M.I.; Santurio, J.M.; Alves, S.H. In vitro activity of immunosuppressive agents against Cryptococcus neoformans. Enferm. Infecc. Microbiol. Clin. 2022, 40, 86–88. [Google Scholar] [CrossRef]
- Cowen, L.E.; Singh, S.D.; Köhler, J.R.; Collins, C.; Zaas, A.K.; Schell, W.A.; Aziz, H.; Mylonakis, E.; Perfect, J.R.; Whitesell, L.; et al. Harnessing Hsp90 function as a powerful, broadly effective therapeutic strategy for fungal infectious disease. Proc. Natl. Acad. Sci. USA 2009, 106, 2818–2823. [Google Scholar] [CrossRef] [PubMed]
- Lamoth, F.; Juvvadi, P.R.; Gehrke, C.; Asfaw, Y.G.; Steinbach, W.J. Transcriptional activation of heat shock protein 90 mediated via a proximal promoter region as trigger of caspofungin resistance in Aspergillus fumigatus. J. Infect. Dis. 2014, 209, 473–481. [Google Scholar] [CrossRef]
- Matthews, R.C.; Rigg, G.; Hodgetts, S.; Carter, T.; Chapman, C.; Gregory, C.; Illidge, C.; Burnie, J. Preclinical assessment of the efficacy of mycograb, a human recombinant antibody against fungal HSP90. Antimicrob. Agents Chemother. 2003, 47, 2208–2216. [Google Scholar] [CrossRef] [PubMed]
- Hodgetts, S.; Nooney, L.; Al-Akeel, R.; Curry, A.; Awad, S.; Matthews, R.; Burnie, J. Efungumab and caspofungin: Pre-clinical data supporting synergy. J. Antimicrob. Chemother. 2008, 61, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- Karwa, R.; Wargo, K.A. Efungumab: A novel agent in the treatment of invasive candidiasis. Ann. Pharmacother. 2009, 43, 1818–1823. [Google Scholar] [CrossRef]
- Rutherford, S.L.; Lindquist, S. Hsp90 as a capacitor for morphological evolution. Nature 1998, 396, 336–342. [Google Scholar] [CrossRef]
- Piłsyk, S.; Mieczkowski, A.; Golan, M.P.; Wawrzyniak, A.; Kruszewska, J.S. Internalization of the. Int. J. Mol. Sci. 2020, 21, 7727. [Google Scholar] [CrossRef]
- Torrente, M.P.; Castellano, L.M.; Shorter, J. Suramin inhibits Hsp104 ATPase and disaggregase activity. PLoS ONE 2014, 9, e110115. [Google Scholar] [CrossRef]
- Milan, R.; Alois, R.; Josef, C.; Jana, B.; Evzen, W. Recombinant protein and DNA vaccines derived from hsp60 Trichophyton mentagrophytes control the clinical course of trichophytosis in bovine species and guinea-pigs. Mycoses 2004, 47, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Gomez, F.J.; Allendoerfer, R.; Deepe, G.S. Vaccination with recombinant heat shock protein 60 from Histoplasma capsulatum protects mice against pulmonary histoplasmosis. Infect. Immun. 1995, 63, 2587–2595. [Google Scholar] [CrossRef] [PubMed]
- Erives, A.J.; Fassler, J.S. Metabolic and chaperone gene loss marks the origin of animals: Evidence for Hsp104 and Hsp78 chaperones sharing mitochondrial enzymes as clients. PLoS ONE 2015, 10, e0117192. [Google Scholar] [CrossRef]
- Durie, C.L.; Lin, J.; Scull, N.W.; Mack, K.L.; Jackrel, M.E.; Sweeny, E.A.; Castellano, L.M.; Shorter, J.; Lucius, A.L. Hsp104 and Potentiated Variants Can Operate as Distinct Nonprocessive Translocases. Biophys. J. 2019, 116, 1856–1872. [Google Scholar] [CrossRef] [PubMed]
- Ami, D.; Natalello, A.; Lotti, M.; Doglia, S.M. Why and how protein aggregation has to be studied in vivo. Microb. Cell Fact. 2013, 12, 17. [Google Scholar] [CrossRef] [PubMed]
- Shorter, J.; Lindquist, S. Prions as adaptive conduits of memory and inheritance. Nat. Rev. Genet. 2005, 6, 435–450. [Google Scholar] [CrossRef]
- Tyedmers, J.; Madariaga, M.L.; Lindquist, S. Prion switching in response to environmental stress. PLoS Biol. 2008, 6, e294. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteostasis Components | Function | Cellular Location | Biological Process Involved in Fungi | Targeting Drugs | References |
|---|---|---|---|---|---|
| Small HSPs | Prevent the onset of protein misfolding and aggregation and have the ability to form HSP oligomers. | Cytosol close to and on the cell membrane. | Stress response following nucleotide depletion or DNA damage; tolerance to thermal/cold, osmotic, and oxidative stress; phagocytosis and virulence. | Apatorsen (OGX-427); Quercetin (Qctn). | [96,98,99,101,102,165] |
| Hsp60 | Auxiliary to Hsp70 in the folding and transport of client proteins through the cell. | Especially in mitochondria but also in the cytosol, the cell wall, and the extracellular space. | Ligands of CD11/CD18 of human macrophages; immunodominant antigens resulting in humoral and cellular responses. | Mefipristone (RU486); Pyrazolopirimidine (EC3016); Mizoribine; Avrainvillamide; Epolactaene; Suvanine. | [65,117,118,119,166] |
| Hsp70 | Centralized function in proteostasis circuits. Folding and assembly of newly synthesized proteins, refolding of denatured or aggregated proteins, protein transport, and degradation mediated by ERAD and the UPR. | All major cell compartments: cytosol, nucleus, ER, and mitochondria. | Binding to human salivary histatin 5; germination, conidiation, and sporulation; interaction with the MAPK pathway. | Pifithrin-µ (2-PES); KNK437; Methylene Blue; Imidazole derivates (apoptozole); Pyrrhoricin, Oncocin and derivates; KLR-70; Adenosine derivates (VER-155008); S1g-2 andS1g-6; Benzimidazole class; 2,5′-thiodipyrimidine (YK-5). | [123,127,128,130,167,168] |
| Hsp40 | Linker between the substrate and Hsp70; Pairs with Hsp70 to promote protein refolding and degradation. | Cytosol, cell wall, and mitochondria. | Host–pathogen interaction, growth, and yeast–mycelium transition. | KNK437; Quercetin (Qcnt); Plumbagin derivates (PLIHZ and PLTFBH). | [80,123,131,132] |
| Hsp90 | Controls the activity of regulatory proteins such as transcription factors and kinases, all of which participate in a wide range of cellular processes. | Cytosol. | Regulation of the cell cycle and yeast-to-hyphae transition; hyphal growth, reproduction, and development at high temperatures (dependent on Ras1 signaling system); biofilms and host cell attachment; colonization of keratinized tissues. | Benzoquinone Ansamycin class (Geldanamycin, 17-AAG, 17-DMAG, Ganetespib); Trichostatin A; Enfungumab; Radicicol; Novobiocin derivates; Deguelin. | [80,136,169] |
| Hsp104 | Prevention and reversal of protein aggregation assisted by Hsp70 and Hsp40. | Cytosol. | Biofilm development; crosstalk with the autophagy mechanism. | Suramin; Guanidine Hydrochloride; NSC 34931; NSC 71948; Hexachlorophene. | [154,157,158] |
| UPR Pathway | Triggered by ER stress caused by the accumulation of toxic misfolded proteins; upregulation of genes that support ER function to rebalance proteostasis. | Endoplasmic Reticulum. | Response and adaptation to environmental stress; virulence; nutrient adaptation and host invasion; keratinolytic capacity; thermotolerance; growth in host molecules and resistance to antifungals. | GRP78: Hyfroxyquinolines; Epigallocatechin gallate (EGCG), and Honokiol (HNK) IRE1: Kinase-biding RNase-attenuators (KIRAs); MKC analogues, and salicylaldehydes ERAD: Eeyarestatin I ER Stress: Dithiothreitol (DTT) and Tunicamycin. | [67,69,170,171,172,173] |
| Autophagy | Recycling of intracellular components. | Autophagosomes. | Mechanism to cope with nutrient deprivation; related to the expression of HSPs. | VPS34: PIK-III mTOR: AZD8055 Lysosomal Function: Bafilomycin A. | [5,33,34,35,36,37] |
| Proteasome | Multicatalytic complexes that cleave proteins into peptides. | Cytosol and nucleus. | F-box proteins from the UPS are related to the following: pseudo-hyphal production, dimorphic switch, cell membrane integrity, sexual sporulation, carbon utilization, signal transduction, and nutrient sensing. | Bortezomib (Velcade); Carfilzomib (Kyprolis); Ixazomib (Ninlaro); Mg-132; Lactacystin. | [174,175,176] |
| HSF1 | A transcription factor that binds to heat shock elements (HSE) in the promoter regions of HSP and related genes, activating the heat shock response (HSR). | Cytosol and nucleus. | Virulence; cell wall integrity; stress response. | Natural Inhibitors: Quercetin, Stresgenin B, Triplotide; Cantharidin; Fisetin; Rocaglamide A; CL-43; 2,4-Bis(4-hydroxybenzyl)Phenol. Synthetic Inhibitors: KNK437; NZ-28; KRIBB11; PW3405; IHSF115; 4,-6-Disubstituted Pyrimidine; CCT251236; α-Acyl Amino Carboxamides. | [56,177,178,179] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neves-da-Rocha, J.; Santos-Saboya, M.J.; Lopes, M.E.R.; Rossi, A.; Martinez-Rossi, N.M. Insights and Perspectives on the Role of Proteostasis and Heat Shock Proteins in Fungal Infections. Microorganisms 2023, 11, 1878. https://doi.org/10.3390/microorganisms11081878
Neves-da-Rocha J, Santos-Saboya MJ, Lopes MER, Rossi A, Martinez-Rossi NM. Insights and Perspectives on the Role of Proteostasis and Heat Shock Proteins in Fungal Infections. Microorganisms. 2023; 11(8):1878. https://doi.org/10.3390/microorganisms11081878
Chicago/Turabian StyleNeves-da-Rocha, João, Maria J. Santos-Saboya, Marcos E. R. Lopes, Antonio Rossi, and Nilce M. Martinez-Rossi. 2023. "Insights and Perspectives on the Role of Proteostasis and Heat Shock Proteins in Fungal Infections" Microorganisms 11, no. 8: 1878. https://doi.org/10.3390/microorganisms11081878
APA StyleNeves-da-Rocha, J., Santos-Saboya, M. J., Lopes, M. E. R., Rossi, A., & Martinez-Rossi, N. M. (2023). Insights and Perspectives on the Role of Proteostasis and Heat Shock Proteins in Fungal Infections. Microorganisms, 11(8), 1878. https://doi.org/10.3390/microorganisms11081878