
Sequencing the Genomes of the First Terrestrial Fungal Lineages: What Have We Learned?

 ,
,  , , , ,
, , , ,  ,
,  , , , ,
, , , ,  , add
Show full author list
, add
Show full author list
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
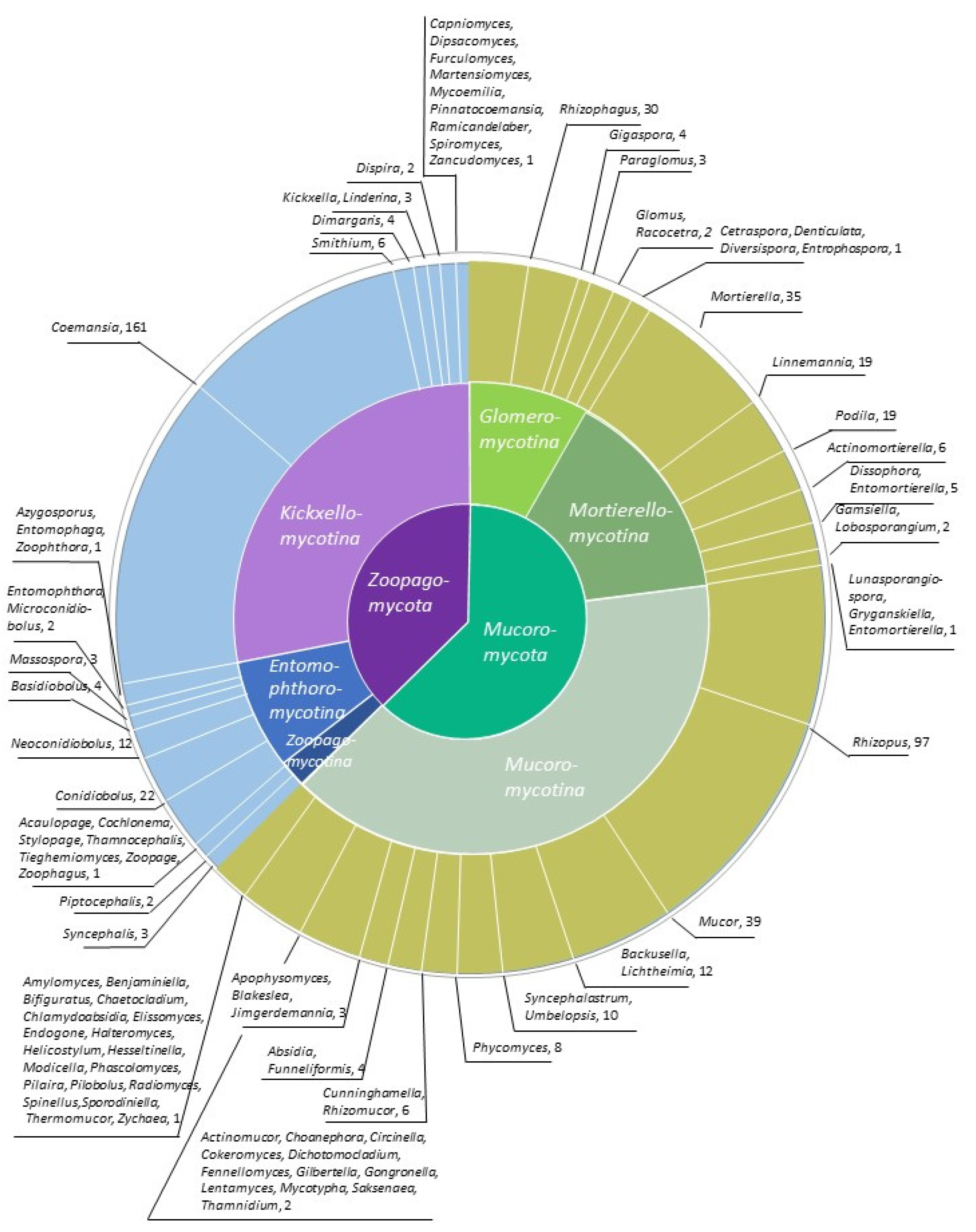
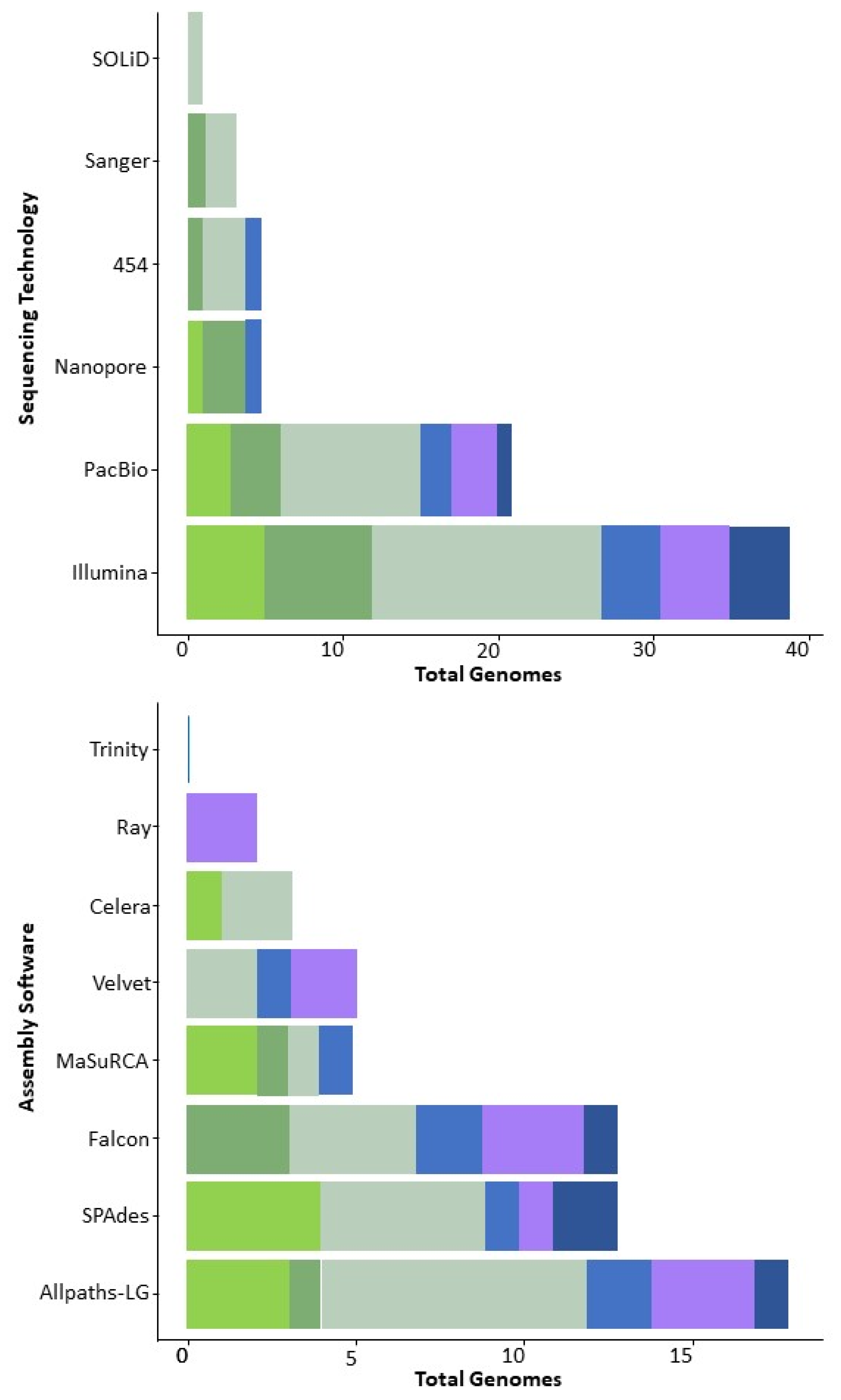
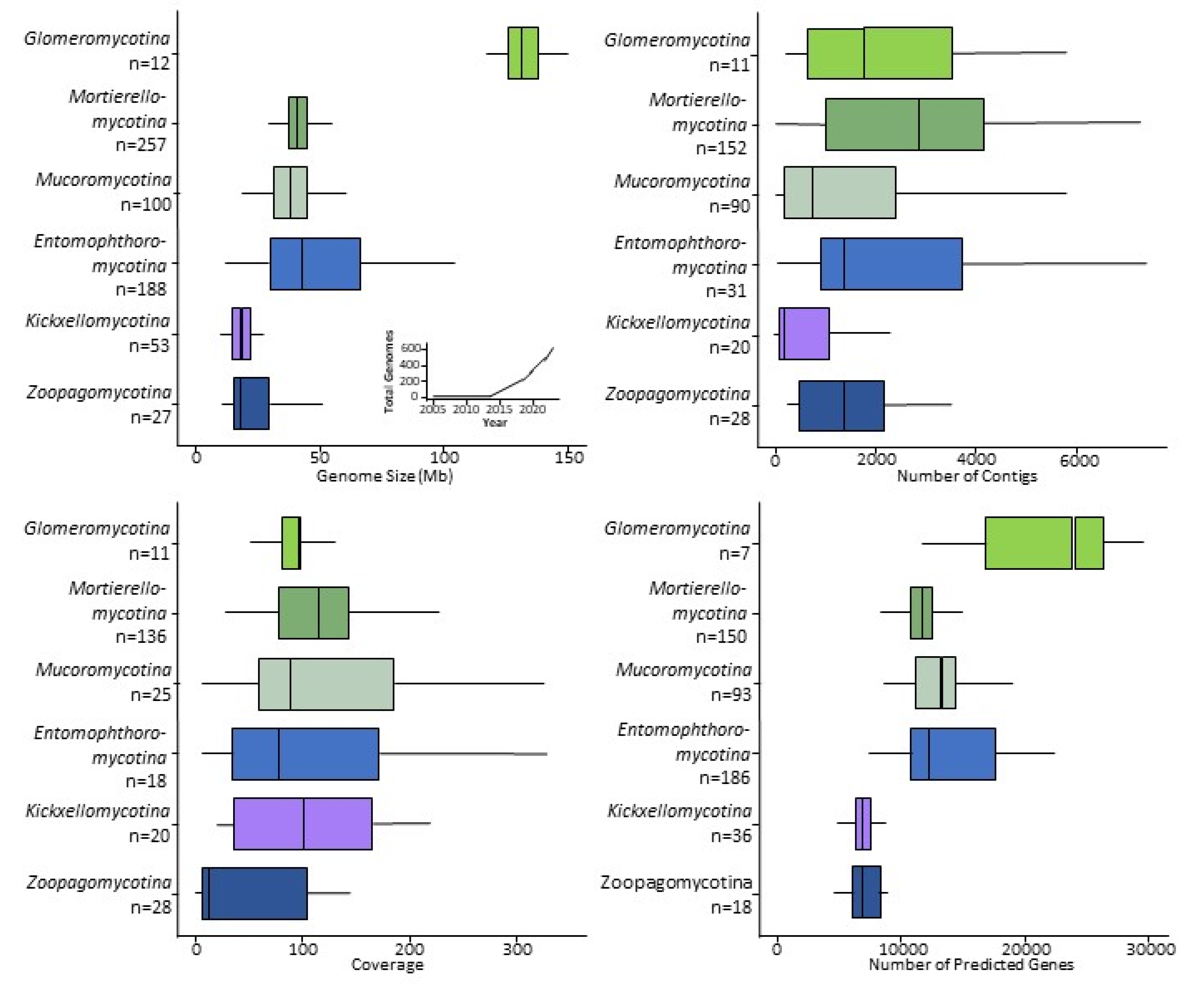
2. Sequencing the Zygomycetes
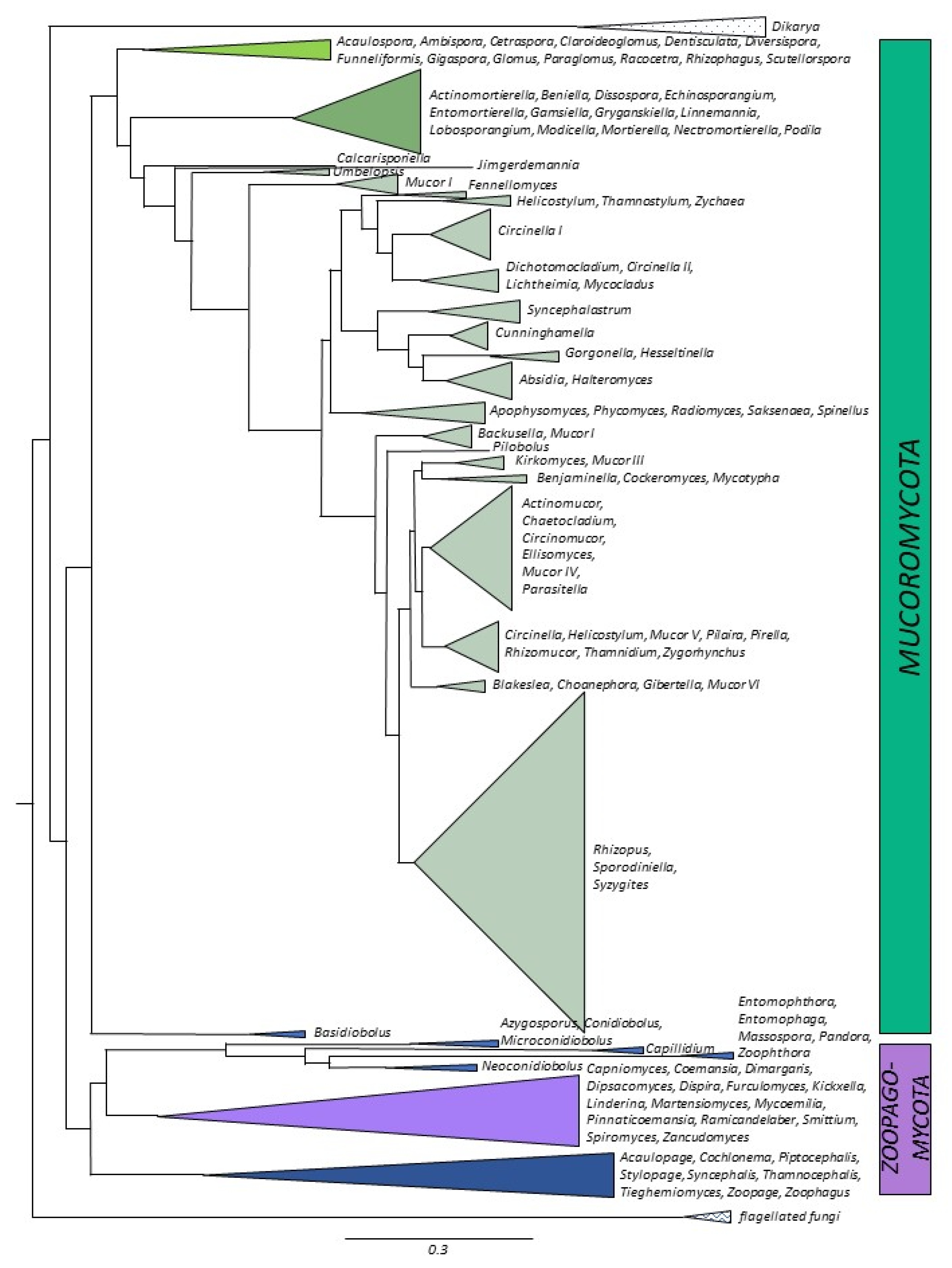
3. Genomes and the Evolution of the Zygomycetes
3.1. Mucoromycota
3.2. Mucoromycotina
3.3. Mortierellomycotina
3.4. Glomeromycotina
3.5. Zoopagomycota
3.6. Kickxellomycotina
3.7. Zoopagomycotina
3.8. Entomophthoromycotina
4. Insights from Genomics
4.1. Adaptation to Terrestrial Life Inferred from Genomes
4.2. Endobacteria
4.3. Secretomes, Gene Clusters, and Secondary Metabolites
4.4. Transposable Elements
5. Conclusions
Supplementary Materials
Funding
Conflicts of Interest
References
- Heaton, L.L.M.; Jones, N.S.; Fricker, M.D. A Mechanistic Explanation of the Transition to Simple Multicellularity in Fungi. Nat. Commun. 2020, 11, 2594. [Google Scholar] [CrossRef]
- Nagy, L.G.; Varga, T.; Csernetics, Á.; Virágh, M. Fungi Took a Unique Evolutionary Route to Multicellularity: Seven Key Challenges for Fungal Multicellular Life. Fungal Biol. Rev. 2020, 34, 151–169. [Google Scholar] [CrossRef]
- de Hoog, G.S.; Guarro, J.; Gené, J.; Figueras, M.J. Atlas of Clinical Fungi, 2nd ed.; Centraalbureau voor Schimmelcultures (CBS): Utrecht, The Netherlands, 2000; ISBN 90-70351-43-9. [Google Scholar]
- Benny, G.L.; Smith, M.E.; Kirk, P.M.; Tretter, E.D.; White, M.M. Challenges and Future Perspectives in the Systematics of Kickxellomycotina, Mortierellomycotina, Mucoromycotina, and Zoopagomycotina. In Biology of Microfungi; Li, D.-W., Ed.; Fungal Biology; Springer International Publishing: Cham, Switzerland, 2016; pp. 65–126. ISBN 978-3-319-29137-6. [Google Scholar]
- James, T.Y.; Letcher, P.M.; Longcore, J.E.; Mozley-Standridge, S.E.; Porter, D.; Powell, M.J.; Griffith, G.W.; Vilgalys, R. A Molecular Phylogeny of the Flagellated Fungi (Chytridiomycota) and Description of a New Phylum (Blastocladiomycota). Mycologia 2006, 98, 860–871. [Google Scholar] [CrossRef] [PubMed]
- Spatafora, J.W.; Chang, Y.; Benny, G.L.; Lazarus, K.; Smith, M.E.; Berbee, M.L.; Bonito, G.; Corradi, N.; Grigoriev, I.; Gryganskyi, A.; et al. A Phylum-Level Phylogenetic Classification of Zygomycete Fungi Based on Genome-Scale Data. Mycologia 2016, 108, 1028–1046. [Google Scholar] [CrossRef] [PubMed]
- Nagy, L.G.; Kovács, G.M.; Krizsán, K. Complex Multicellularity in Fungi: Evolutionary Convergence, Single Origin, or Both? Biol. Rev. Camb. Philos. Soc. 2018, 93, 1778–1794. [Google Scholar] [CrossRef]
- Kiss, E.; Hegedüs, B.; Virágh, M.; Varga, T.; Merényi, Z.; Kószó, T.; Bálint, B.; Prasanna, A.N.; Krizsán, K.; Kocsubé, S.; et al. Comparative Genomics Reveals the Origin of Fungal Hyphae and Multicellularity. Nat. Commun. 2019, 10, 4080. [Google Scholar] [CrossRef] [PubMed]
- Stajich, J.E. Fungal Genomes and Insights into the Evolution of the Kingdom. Microbiol. Spectr. 2017, 5, 1–25. [Google Scholar] [CrossRef]
- Berbee, M.L.; James, T.Y.; Strullu-Derrien, C. Early Diverging Fungi: Diversity and Impact at the Dawn of Terrestrial Life. Annu. Rev. Microbiol. 2017, 71, 41–60. [Google Scholar] [CrossRef]
- Ahrendt, S.R.; Quandt, C.A.; Ciobanu, D.; Clum, A.; Salamov, A.; Andreopoulos, B.; Cheng, J.-F.; Woyke, T.; Pelin, A.; Henrissat, B.; et al. Leveraging Single-Cell Genomics to Expand the Fungal Tree of Life. Nat. Microbiol. 2018, 3, 1417–1428. [Google Scholar] [CrossRef]
- Vandepol, N.; Liber, J.; Desirò, A.; Na, H.; Kennedy, M.; Barry, K.; Grigoriev, I.V.; Miller, A.N.; O’Donnell, K.; Stajich, J.E.; et al. Resolving the Mortierellaceae Phylogeny through Synthesis of Multi-Gene Phylogenetics and Phylogenomics. Fungal Divers. 2020, 104, 267–289. [Google Scholar] [CrossRef]
- Chang, M.C.; Hur, J.; Park, D. Interpreting the COVID-19 Test Results: A Guide for Physiatrists. Am. J. Phys. Med. Rehabil. 2020, 99, 583–585. [Google Scholar] [CrossRef] [PubMed]
- Spatafora, J.W.; Aime, M.C.; Grigoriev, I.V.; Stajich, J.E.; Blackwell, M. The Fungal Tree of Life: From Molecular Systematics to Genome-Scale Phylogenies. Microbiol. Spectrum. 2017, 5, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, N.K.; Stajich, J.E.; Benny, G.L.; Barry, K.; Mondo, S.; LaButti, K.; Lipzen, A.; Daum, C.; Grigoriev, I.V.; Ho, H.-M.; et al. Mycoparasites, Gut Dwellers, and Saprotrophs: Phylogenomic Reconstructions and Comparative Analyses of Kickxellomycotina Fungi. Genome Biol. Evol. 2023, 15, evac185. [Google Scholar] [CrossRef]
- Gryganskyi, A.P.; Muszewska, A. Whole Genome Sequencing and the Zygomycota. Fungal Genom. Biol. 2014, 4, e116. [Google Scholar] [CrossRef]
- Stephens, Z.D.; Lee, S.Y.; Faghri, F.; Campbell, R.H.; Zhai, C.; Efron, M.J.; Iyer, R.; Schatz, M.C.; Sinha, S.; Robinson, G.E. Big Data: Astronomical or Genomical? PLoS Biol. 2015, 13, e1002195. [Google Scholar] [CrossRef]
- Ma, L.-J.; Ibrahim, A.S.; Skory, C.; Grabherr, M.G.; Burger, G.; Butler, M.; Elias, M.; Idnurm, A.; Lang, B.F.; Sone, T.; et al. Genomic Analysis of the Basal Lineage Fungus Rhizopus oryzae Reveals a Whole-Genome Duplication. PLoS Genet. 2009, 5, e1000549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Yang, Y.; Castlebury, L.A.; Cerniglia, C.E. A Method for the Large Scale Isolation of High Transformation Efficiency Fungal Genomic DNA. FEMS Microbiol. Lett. 1996, 145, 261–265. [Google Scholar] [CrossRef]
- Davis, W.J.; Amses, K.R.; Benny, G.L.; Carter-House, D.; Chang, Y.; Grigoriev, I.; Smith, M.E.; Spatafora, J.W.; Stajich, J.E.; James, T.Y. Genome-Scale Phylogenetics Reveals a Monophyletic Zoopagales (Zoopagomycota, Fungi). Mol. Phylogenetics Evol. 2019, 133, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Montoliu-Nerin, M.; Sánchez-García, M.; Bergin, C.; Grabherr, M.; Ellis, B.; Kutschera, V.E.; Kierczak, M.; Johannesson, H.; Rosling, A. Building de Novo Reference Genome Assemblies of Complex Eukaryotic Microorganisms from Single Nuclei. Sci. Rep. 2020, 10, 1303. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Limpens, E.; Zhang, Z.; Ivanov, S.; Saunders, D.G.O.; Mu, D.; Pang, E.; Cao, H.; Cha, H.; Lin, T.; et al. Single Nucleus Genome Sequencing Reveals High Similarity among Nuclei of an Endomycorrhizal Fungus. PLoS Genet. 2014, 10, e1004078. [Google Scholar] [CrossRef]
- Małagocka, J.; Grell, M.N.; Lange, L.; Eilenberg, J.; Jensen, A.B. Transcriptome of an Entomophthoralean Fungus (Pandora formicae) Shows Molecular Machinery Adjusted for Successful Host Exploitation and Transmission. J. Invertebr. Pathol. 2015, 128, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Desirò, A.; Na, H.; Sandor, L.; Lipzen, A.; Clum, A.; Barry, K.; Grigoriev, I.V.; Martin, F.M.; Stajich, J.E.; et al. Phylogenomics of Endogonaceae and Evolution of Mycorrhizas within Mucoromycota. New Phytol. 2019, 222, 511–525. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, M.O.; Russ, C.; Ross, M.G.; Gabriel, S.B.; Nusbaum, C.; DePristo, M.A. Pacific Biosciences Sequencing Technology for Genotyping and Variation Discovery in Human Data. BMC Genom. 2012, 13, 375. [Google Scholar] [CrossRef]
- Maeda, T.; Kobayashi, Y.; Kameoka, H.; Okuma, N.; Takeda, N.; Yamaguchi, K.; Bino, T.; Shigenobu, S.; Kawaguchi, M. Evidence of Non-Tandemly Repeated RDNAs and Their Intragenomic Heterogeneity in Rhizophagus irregularis. Commun. Biol. 2018, 1, 87. [Google Scholar] [CrossRef]
- Segerman, B. The Most Frequently Used Sequencing Technologies and Assembly Methods in Different Time Segments of the Bacterial Surveillance and RefSeq Genome Databases. Front. Cell. Infect. Microbiol. 2020, 10, 527102. [Google Scholar] [CrossRef] [PubMed]
- El-Metwally, S.; Hamza, T.; Zakaria, M.; Helmy, M. Next-Generation Sequence Assembly: Four Stages of Data Processing and Computational Challenges. PLoS Comput. Biol. 2013, 9, e1003345. [Google Scholar] [CrossRef]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de Novo Short Read Assembly Using de Bruijn Graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef]
- Myers, J.M.; Bonds, A.E.; Clemons, R.A.; Thapa, N.A.; Simmons, D.R.; Carter-House, D.; Ortanez, J.; Liu, P.; Miralles-Durán, A.; Desirò, A.; et al. Survey of Early-Diverging Lineages of Fungi Reveals Abundant and Diverse Mycoviruses. mBio 2020, 11, e02027-20. [Google Scholar] [CrossRef]
- Chin, C.-S.; Alexander, D.H.; Marks, P.; Klammer, A.A.; Drake, J.; Heiner, C.; Clum, A.; Copeland, A.; Huddleston, J.; Eichler, E.E.; et al. Nonhybrid, Finished Microbial Genome Assemblies from Long-Read SMRT Sequencing Data. Nat. Methods 2013, 10, 563–569. [Google Scholar] [CrossRef]
- Shapiro, H. Outline of the Assembly Process: JAZZ, the JGI In-House Assembler. Available online: https://escholarship.org/uc/item/0g8792vf (accessed on 13 March 2023).
- Boisvert, S.; Raymond, F.; Godzaridis, É.; Laviolette, F.; Corbeil, J. Ray Meta: Scalable de Novo Metagenome Assembly and Profiling. Genome Biol. 2012, 13, R122. [Google Scholar] [CrossRef]
- Zimin, A.V.; Marçais, G.; Puiu, D.; Roberts, M.; Salzberg, S.L.; Yorke, J.A. The MaSuRCA Genome Assembler. Bioinformatics 2013, 29, 2669–2677. [Google Scholar] [CrossRef]
- Butler, J.; MacCallum, I.; Kleber, M.; Shlyakhter, I.A.; Belmonte, M.K.; Lander, E.S.; Nusbaum, C.; Jaffe, D.B. ALLPATHS: De Novo Assembly of Whole-Genome Shotgun Microreads. Genome Res. 2008, 18, 810–820. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. SOAPdenovo2: An Empirically Improved Memory-Efficient Short-Read de Novo Assembler. Gigascience 2012, 1, 18. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Miller, J.R.; Koren, S.; Sutton, G. Assembly Algorithms for Next-Generation Sequencing Data. Genomics 2010, 95, 315–327. [Google Scholar] [CrossRef]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and Accurate Long-Read Assembly via Adaptive k-Mer Weighting and Repeat Separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef]
- Lam, K.-K.; LaButti, K.; Khalak, A.; Tse, D. FinisherSC: A Repeat-Aware Tool for Upgrading de Novo Assembly Using Long Reads. Bioinformatics 2015, 31, 3207–3209. [Google Scholar] [CrossRef]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of Long, Error-Prone Reads Using Repeat Graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef]
- Li, D.; Liu, C.-M.; Luo, R.; Sadakane, K.; Lam, T.-W. MEGAHIT: An Ultra-Fast Single-Node Solution for Large and Complex Metagenomics Assembly via Succinct de Bruijn Graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef] [PubMed]
- Gregory, T.R. CHAPTER 1—Genome Size Evolution in Animals. In The Evolution of the Genome; Gregory, T.R., Ed.; Academic Press: Burlington, NJ, USA, 2005; pp. 3–87. ISBN 978-0-12-301463-4. [Google Scholar]
- Pellicer, J.; Hidalgo, O.; Dodsworth, S.; Leitch, I.J. Genome Size Diversity and Its Impact on the Evolution of Land Plants. Genes 2018, 9, 88. [Google Scholar] [CrossRef]
- Tavares, S.; Ramos, A.P.; Pires, A.S.; Azinheira, H.G.; Caldeirinha, P.; Link, T.; Abranches, R.; do Céu Silva, M.; Voegele, R.T.; Loureiro, J.; et al. Genome Size Analyses of Pucciniales Reveal the Largest Fungal Genomes. Front. Plant Sci. 2014, 5, 422. [Google Scholar] [CrossRef]
- Salazar, A.N.; Gorter de Vries, A.R.; van den Broek, M.; Brouwers, N.; de la Torre Cortès, P.; Kuijpers, N.G.A.; Daran, J.-M.G.; Abeel, T. Chromosome Level Assembly and Comparative Genome Analysis Confirm Lager-Brewing Yeasts Originated from a Single Hybridization. BMC Genom. 2019, 20, 916. [Google Scholar] [CrossRef]
- Wieloch, W. Chromosome Visualisation in Filamentous Fungi. J. Microbiol. Methods 2006, 67, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Boyce, G.R.; Gluck-Thaler, E.; Slot, J.C.; Stajich, J.E.; Davis, W.J.; James, T.Y.; Cooley, J.R.; Panaccione, D.G.; Eilenberg, J.; De Fine Licht, H.H.; et al. Psychoactive Plant- and Mushroom-Associated Alkaloids from Two Behavior Modifying Cicada Pathogens. Fungal Ecol. 2019, 41, 147–164. [Google Scholar] [CrossRef]
- Vinson, J.P.; Jaffe, D.B.; O’Neill, K.; Karlsson, E.K.; Stange-Thomann, N.; Anderson, S.; Mesirov, J.P.; Satoh, N.; Satou, Y.; Nusbaum, C.; et al. Assembly of Polymorphic Genomes: Algorithms and Application to Ciona savignyi. Genome Res. 2005, 15, 1127–1135. [Google Scholar] [CrossRef]
- Chibucos, M.C.; Soliman, S.; Gebremariam, T.; Lee, H.; Daugherty, S.; Orvis, J.; Shetty, A.C.; Crabtree, J.; Hazen, T.H.; Etienne, K.A.; et al. An Integrated Genomic and Transcriptomic Survey of Mucormycosis-Causing Fungi. Nat. Commun. 2016, 7, 12218. [Google Scholar] [CrossRef]
- Li, Y.; Steenwyk, J.L.; Chang, Y.; Wang, Y.; James, T.Y.; Stajich, J.E.; Spatafora, J.W.; Groenewald, M.; Dunn, C.W.; Hittinger, C.T.; et al. A Genome-Scale Phylogeny of the Kingdom Fungi. Curr. Biol. 2021, 31, 1653–1665.e5. [Google Scholar] [CrossRef]
- Strassert, J.F.H.; Monaghan, M.T. Phylogenomic Insights into the Early Diversification of Fungi. Curr. Biol. 2022, 32, 3628–3635.e3. [Google Scholar] [CrossRef]
- Galindo, L.J.; López-García, P.; Torruella, G.; Karpov, S.; Moreira, D. Phylogenomics of a New Fungal Phylum Reveals Multiple Waves of Reductive Evolution across Holomycota. Nat. Commun. 2021, 12, 4973. [Google Scholar] [CrossRef] [PubMed]
- Naranjo-Ortiz, M.A.; Gabaldón, T. Fungal Evolution: Diversity, Taxonomy and Phylogeny of the Fungi. Biol. Rev. Camb. Philos. Soc. 2019, 94, 2101–2137. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.A.; Ganley, A.R.D.; Gabaldón, T.; Cox, M.P. The Case of the Missing Ancient Fungal Polyploids. Am. Nat. 2016, 188, 602–614. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.N.; Salgado, J.F.M.; Natsopoulou, M.E.; Kristensen, T.; Stajich, J.E.; De Fine Licht, H.H. Diploidy within a Haploid Genus of Entomopathogenic Fungi. Genome Biol. Evol. 2021, 13, evab158. [Google Scholar] [CrossRef]
- Amses, K.R.; Simmons, D.R.; Longcore, J.E.; Mondo, S.J.; Seto, K.; Jerônimo, G.H.; Bonds, A.E.; Quandt, C.A.; Davis, W.J.; Chang, Y.; et al. Diploid-Dominant Life Cycles Characterize the Early Evolution of Fungi. Proc. Natl. Acad. Sci. USA 2022, 119, e2116841119. [Google Scholar] [CrossRef]
- Genre, A.; Lanfranco, L.; Perotto, S.; Bonfante, P. Unique and Common Traits in Mycorrhizal Symbioses. Nat. Rev. Microbiol. 2020, 18, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Hoysted, G.A.; Jacob, A.S.; Kowal, J.; Giesemann, P.; Bidartondo, M.I.; Duckett, J.G.; Gebauer, G.; Rimington, W.R.; Schornack, S.; Pressel, S.; et al. Mucoromycotina Fine Root Endophyte Fungi Form Nutritional Mutualisms with Vascular Plants. Plant Physiol. 2019, 181, 565–577. [Google Scholar] [CrossRef]
- Wani, Z.A.; Kumar, A.; Sultan, P.; Bindu, K.; Riyaz-Ul-Hassan, S.; Ashraf, N. Mortierella alpina CS10E4, an Oleaginous Fungal Endophyte of Crocus sativus L. Enhances Apocarotenoid Biosynthesis and Stress Tolerance in the Host Plant. Sci. Rep. 2017, 7, 8598. [Google Scholar] [CrossRef] [PubMed]
- Walsh, E.; Luo, J.; Khiste, S.; Scalera, A.; Sajjad, S.; Zhang, N. Pygmaeomycetaceae, a New Root-Associated Family in Mucoromycotina from the Pygmy Pine Plains. Mycologia 2021, 113, 134–145. [Google Scholar] [CrossRef]
- Liu, S.; Liu, M.; Liao, Q.-G.; Lü, F.-B.; Zhao, X.-L. Effects of Inoculated Mycorrhizal Fungi and Non-Mycorrhizal Beneficial Micro-Organisms on Plant Traits, Nutrient Uptake and Root-Associated Fungal Community Composition of the Cymbidium hybridum in Greenhouse. J. Appl. Microbiol. 2021, 131, 413–424. [Google Scholar] [CrossRef]
- Steinbrink, J.M.; Miceli, M.H. Clinical Review of Mucormycosis. Infect. Dis. Clin. N. Am. 2021, 35, 435–452. [Google Scholar] [CrossRef]
- Corrochano, L.M.; Kuo, A.; Marcet-Houben, M.; Polaino, S.; Salamov, A.; Villalobos-Escobedo, J.M.; Grimwood, J.; Álvarez, M.I.; Avalos, J.; Bauer, D.; et al. Expansion of Signal Transduction Pathways in Fungi by Extensive Genome Duplication. Curr. Biol. 2016, 26, 1577–1584. [Google Scholar] [CrossRef]
- Partida-Martinez, L.P.; Hertweck, C. Pathogenic Fungus Harbours Endosymbiotic Bacteria for Toxin Production. Nature 2005, 437, 884–888. [Google Scholar] [CrossRef] [PubMed]
- Torres-Cortés, G.; Ghignone, S.; Bonfante, P.; Schüßler, A. Mosaic Genome of Endobacteria in Arbuscular Mycorrhizal Fungi: Transkingdom Gene Transfer in an Ancient Mycoplasma-Fungus Association. Proc. Natl. Acad. Sci. USA 2015, 112, 7785–7790. [Google Scholar] [CrossRef]
- Mondo, S.J.; Salvioli, A.; Bonfante, P.; Morton, J.B.; Pawlowska, T.E. Nondegenerative Evolution in Ancient Heritable Bacterial Endosymbionts of Fungi. Mol. Biol. Evol. 2016, 33, 2216–2231. [Google Scholar] [CrossRef] [PubMed]
- Bonfante, P.; Desirò, A. Who Lives in a Fungus? The Diversity, Origins and Functions of Fungal Endobacteria Living in Mucoromycota. ISME J. 2017, 11, 1727–1735. [Google Scholar] [CrossRef] [PubMed]
- Uehling, J.; Gryganskyi, A.; Hameed, K.; Tschaplinski, T.; Misztal, P.K.; Wu, S.; Desirò, A.; Vande Pol, N.; Du, Z.; Zienkiewicz, A.; et al. Comparative Genomics of Mortierella elongata and Its Bacterial Endosymbiont Mycoavidus cysteinexigens. Environ. Microbiol. 2017, 19, 2964–2983. [Google Scholar] [CrossRef] [PubMed]
- Lastovetsky, O.A.; Ahn, E.; Mondo, S.J.; Toomer, K.H.; Zhang, A.; Johnson, L.M.; Pawlowska, T.E. Distribution and Population Structure of Endobacteria in Arbuscular Mycorrhizal Fungi at North Atlantic Dunes. ISME J. 2018, 12, 3001–3013. [Google Scholar] [CrossRef]
- Espino-Vázquez, A.N.; Bermúdez-Barrientos, J.R.; Cabrera-Rangel, J.F.; Córdova-López, G.; Cardoso-Martínez, F.; Martínez-Vázquez, A.; Camarena-Pozos, D.A.; Mondo, S.J.; Pawlowska, T.E.; Abreu-Goodger, C.; et al. Narnaviruses: Novel Players in Fungal-Bacterial Symbioses. ISME J. 2020, 14, 1743–1754. [Google Scholar] [CrossRef] [PubMed]
- Bever, J.D.; Kang, H.-J.; Kaonongbua, W.; Wang, M. Genomic Organization and Mechanisms of Inheritance in Arbuscular Mycorrhizal Fungi: Contrasting the Evidence and Implications of Current Theories. In Mycorrhiza; Varma, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 135–148. ISBN 978-3-540-78824-9. [Google Scholar]
- Martin, F.; Gianinazzi-Pearson, V.; Hijri, M.; Lammers, P.; Requena, N.; Sanders, I.R.; Shachar-Hill, Y.; Shapiro, H.; Tuskan, G.A.; Young, J.P.W. The Long Hard Road to a Completed Glomus intraradices Genome. New Phytol. 2008, 180, 747–750. [Google Scholar] [CrossRef]
- Lebreton, A.; Corre, E.; Jany, J.-L.; Brillet-Guéguen, L.; Pèrez-Arques, C.; Garre, V.; Monsoor, M.; Debuchy, R.; Le Meur, C.; Coton, E.; et al. Comparative Genomics Applied to Mucor Species with Different Lifestyles. BMC Genom. 2020, 21, 135. [Google Scholar] [CrossRef]
- Vellanki, S.; Navarro-Mendoza, M.I.; Garcia, A.E.; Murcia, L.; Perez-Arques, C.; Garre, V.; Nicolas, F.E.; Lee, S.C. Mucor circinelloides: Growth, Maintenance and Genetic Manipulation. Curr. Protoc. Microbiol. 2018, 49, e53. [Google Scholar] [CrossRef]
- Boon, E.; Zimmerman, E.; Lang, B.F.; Hijri, M. Intra-Isolate Genome Variation in Arbuscular Mycorrhizal Fungi Persists in the Transcriptome. J. Evol. Biol. 2010, 23, 1519–1527. [Google Scholar] [CrossRef] [PubMed]
- Muszewska, A.; Okrasińska, A.; Steczkiewicz, K.; Drgas, O.; Orłowska, M.; Perlińska-Lenart, U.; Aleksandrzak-Piekarczyk, T.; Szatraj, K.; Zielenkiewicz, U.; Piłsyk, S.; et al. Metabolic Potential, Ecology and Presence of Associated Bacteria Is Reflected in Genomic Diversity of Mucoromycotina. Front. Microbiol. 2021, 12, 636986. [Google Scholar] [CrossRef] [PubMed]
- Muszewska, A.; Steczkiewicz, K.; Ginalski, K. DIRS and Ngaro Retrotransposons in Fungi. PLoS ONE 2013, 8, e76319. [Google Scholar] [CrossRef] [PubMed]
- Muszewska, A.; Steczkiewicz, K.; Stepniewska-Dziubinska, M.; Ginalski, K. Cut-and-Paste Transposons in Fungi with Diverse Lifestyles. Genome Biol. Evol. 2017, 9, 3463–3477. [Google Scholar] [CrossRef]
- Mondo, S.J.; Dannebaum, R.O.; Kuo, R.C.; Louie, K.B.; Bewick, A.J.; LaButti, K.; Haridas, S.; Kuo, A.; Salamov, A.; Ahrendt, S.R.; et al. Widespread Adenine N6-Methylation of Active Genes in Fungi. Nat. Genet. 2017, 49, 964–968. [Google Scholar] [CrossRef] [PubMed]
- Corradi, N.; Sanders, I.R. Evolution of the P-Type II ATPase Gene Family in the Fungi and Presence of Structural Genomic Changes among Isolates of Glomus intraradices. BMC Evol. Biol. 2006, 6, 21. [Google Scholar] [CrossRef]
- Halary, S.; Malik, S.-B.; Lildhar, L.; Slamovits, C.H.; Hijri, M.; Corradi, N. Conserved Meiotic Machinery in Glomus spp., a Putatively Ancient Asexual Fungal Lineage. Genome Biol. Evol. 2011, 3, 950–958. [Google Scholar] [CrossRef]
- Halary, S.; Daubois, L.; Terrat, Y.; Ellenberger, S.; Wöstemeyer, J.; Hijri, M. Mating Type Gene Homologues and Putative Sex Pheromone-Sensing Pathway in Arbuscular Mycorrhizal Fungi, a Presumably Asexual Plant Root Symbiont. PLoS ONE 2013, 8, e80729. [Google Scholar] [CrossRef]
- Gryganskyi, A.P.; Golan, J.; Dolatabadi, S.; Mondo, S.; Robb, S.; Idnurm, A.; Muszewska, A.; Steczkiewicz, K.; Masonjones, S.; Liao, H.-L.; et al. Phylogenetic and Phylogenomic Definition of Rhizopus Species. G3 (Bethesda) 2018, 8, 2007–2018. [Google Scholar] [CrossRef]
- Gryganskyi, A.P.; Nie, Y.; Hajek, A.E.; Hodge, K.T.; Liu, X.-Y.; Aadland, K.; Voigt, K.; Anishchenko, I.M.; Kutovenko, V.B.; Kava, L.; et al. The Early Terrestrial Fungal Lineage of Conidiobolus-Transition from Saprotroph to Parasitic Lifestyle. J. Fungi 2022, 8, 789. [Google Scholar] [CrossRef]
- Tisserant, E.; Malbreil, M.; Kuo, A.; Kohler, A.; Symeonidi, A.; Balestrini, R.; Charron, P.; Duensing, N.; Frei dit Frey, N.; Gianinazzi-Pearson, V.; et al. Genome of an Arbuscular Mycorrhizal Fungus Provides Insight into the Oldest Plant Symbiosis. Proc. Natl. Acad. Sci. USA 2013, 110, 20117–20122. [Google Scholar] [CrossRef]
- Wang, Y.; Chang, Y.; Ortañez, J.; Peña, J.F.; Carter-House, D.; Reynolds, N.K.; Smith, M.E.; Benny, G.; Mondo, S.J.; Salamov, A.; et al. Divergent Evolution of Early Terrestrial Fungi Reveals the Evolution of Mucormycosis Pathogenicity Factors. Genome Biol. Evol. 2023, 15, evad046. [Google Scholar] [CrossRef]
- Desirò, A.; Rimington, W.R.; Jacob, A.; Pol, N.V.; Smith, M.E.; Trappe, J.M.; Bidartondo, M.I.; Bonito, G. Multigene Phylogeny of Endogonales, an Early Diverging Lineage of Fungi Associated with Plants. IMA Fungus 2017, 8, 245–257. [Google Scholar] [CrossRef]
- Ogura-Tsujita, Y.; Yamamoto, K.; Hirayama, Y.; Ebihara, A.; Morita, N.; Imaichi, R. Fern Gametophytes of Angiopteris lygodiifolia and Osmunda japonica Harbor Diverse Mucoromycotina Fungi. J. Plant Res. 2019, 132, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Rimington, W.R.; Pressel, S.; Duckett, J.G.; Field, K.J.; Bidartondo, M.I. Evolution and Networks in Ancient and Widespread Symbioses between Mucoromycotina and Liverworts. Mycorrhiza 2019, 29, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, S.; Polaino, S.; Shakya, V.P.S.; Idnurm, A. A New Genetic Linkage Map of the Zygomycete Fungus Phycomyces blakesleeanus. PLoS ONE 2013, 8, e58931. [Google Scholar] [CrossRef]
- Linde, J.; Schwartze, V.; Binder, U.; Lass-Flörl, C.; Voigt, K.; Horn, F. De Novo Whole-Genome Sequence and Genome Annotation of Lichtheimia ramosa. Genome Announc. 2014, 2, e00888-14. [Google Scholar] [CrossRef] [PubMed]
- Schwartze, V.U.; Winter, S.; Shelest, E.; Marcet-Houben, M.; Horn, F.; Wehner, S.; Linde, J.; Valiante, V.; Sammeth, M.; Riege, K.; et al. Gene Expansion Shapes Genome Architecture in the Human Pathogen Lichtheimia corymbifera: An Evolutionary Genomics Analysis in the Ancient Terrestrial Mucorales (Mucoromycotina). PLoS Genet. 2014, 10, e1004496. [Google Scholar] [CrossRef]
- Torres-Cruz, T.J.; Billingsley Tobias, T.L.; Almatruk, M.; Hesse, C.N.; Kuske, C.R.; Desirò, A.; Benucci, G.M.N.; Bonito, G.; Stajich, J.E.; Dunlap, C.; et al. Bifiguratus adelaidae, Gen. et Sp. Nov., a New Member of Mucoromycotina in Endophytic and Soil-Dwelling Habitats. Mycologia 2017, 109, 363–378. [Google Scholar] [CrossRef]
- Hirose, D.; Degawa, Y.; Inaba, S.; Tokumasu, S. The Anamorphic Genus Calcarisporiella Is a New Member of the Mucoromycotina. Mycoscience 2012, 53, 256–260. [Google Scholar] [CrossRef]
- Nagy, L.G.; Petkovits, T.; Kovács, G.M.; Voigt, K.; Vágvölgyi, C.; Papp, T. Where Is the Unseen Fungal Diversity Hidden? A Study of Mortierella Reveals a Large Contribution of Reference Collections to the Identification of Fungal Environmental Sequences. New Phytol. 2011, 191, 789–794. [Google Scholar] [CrossRef]
- Smith, M.E.; Gryganskyi, A.; Bonito, G.; Nouhra, E.; Moreno-Arroyo, B.; Benny, G. Phylogenetic Analysis of the Genus Modicella Reveals an Independent Evolutionary Origin of Sporocarp-Forming Fungi in the Mortierellales. Fungal Genet. Biol. 2013, 61, 61–68. [Google Scholar] [CrossRef]
- Macias, A.M.; Geiser, D.M.; Stajich, J.E.; Łukasik, P.; Veloso, C.; Bublitz, D.C.; Berger, M.C.; Boyce, G.R.; Hodge, K.; Kasson, M.T. Evolutionary Relationships among Massospora Spp. (Entomophthorales), Obligate Pathogens of Cicadas. Mycologia 2020, 112, 1060–1074. [Google Scholar] [CrossRef] [PubMed]
- Bonito, G.; Hameed, K.; Ventura, R.; Krishnan, J.; Schadt, C.W.; Vilgalys, R. Isolating a Functionally Relevant Guild of Fungi from the Root Microbiome of Populus. Fungal Ecol. 2016, 22, 35–42. [Google Scholar] [CrossRef]
- Vandepol, N.; Liber, J.; Yocca, A.; Matlock, J.; Edger, P.; Bonito, G. Linnemannia elongata (Mortierellaceae) Stimulates Arabidopsis thaliana Aerial Growth and Responses to Auxin, Ethylene, and Reactive Oxygen Species. PLoS ONE 2022, 17, e0261908. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Bonito, G.; Hsu, C.-M.; Hameed, K.; Vilgalys, R.; Liao, H.-L. Mortierella elongata Increases Plant Biomass among Non-Leguminous Crop Species. Agronomy 2020, 10, 754. [Google Scholar] [CrossRef]
- Wurlitzer, J.M.; Stanišić, A.; Wasmuth, I.; Jungmann, S.; Fischer, D.; Kries, H.; Gressler, M. Bacterial-like Nonribosomal Peptide Synthetases Produce Cyclopeptides in the Zygomycetous Fungus Mortierella alpina. Appl. Environ. Microbiol. 2021, 87, e02051-20. [Google Scholar] [CrossRef]
- Remy, W.; Taylor, T.N.; Hass, H.; Kerp, H. Four Hundred-Million-Year-Old Vesicular Arbuscular Mycorrhizae. Proc. Natl. Acad. Sci. USA 1994, 91, 11841–11843. [Google Scholar] [CrossRef]
- Smith, S.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Cambridge, MA, USA, 2008; ISBN 978-0-12-370526-6. [Google Scholar]
- Hijri, M.; Sanders, I.R. The Arbuscular Mycorrhizal Fungus Glomus intraradices Is Haploid and Has a Small Genome Size in the Lower Limit of Eukaryotes. Fungal Genet. Biol. 2004, 41, 253–261. [Google Scholar] [CrossRef]
- Sędzielewska, K.A.; Fuchs, J.; Temsch, E.M.; Baronian, K.; Watzke, R.; Kunze, G. Estimation of the Glomus intraradices Nuclear DNA Content. New Phytol. 2011, 192, 794–797. [Google Scholar] [CrossRef] [PubMed]
- Venice, F.; Ghignone, S.; Salvioli di Fossalunga, A.; Amselem, J.; Novero, M.; Xianan, X.; Sędzielewska Toro, K.; Morin, E.; Lipzen, A.; Grigoriev, I.V.; et al. At the Nexus of Three Kingdoms: The Genome of the Mycorrhizal Fungus Gigaspora margarita Provides Insights into Plant, Endobacterial and Fungal Interactions. Environ. Microbiol. 2020, 22, 122–141. [Google Scholar] [CrossRef]
- Ropars, J.; Toro, K.S.; Noel, J.; Pelin, A.; Charron, P.; Farinelli, L.; Marton, T.; Krüger, M.; Fuchs, J.; Brachmann, A.; et al. Evidence for the Sexual Origin of Heterokaryosis in Arbuscular Mycorrhizal Fungi. Nat. Microbiol. 2016, 1, 16033. [Google Scholar] [CrossRef]
- Idnurm, A.; Walton, F.J.; Floyd, A.; Heitman, J. Identification of the Sex Genes in an Early Diverged Fungus. Nature 2008, 451, 193–196. [Google Scholar] [CrossRef]
- Kuhn, G.; Hijri, M.; Sanders, I.R. Evidence for the Evolution of Multiple Genomes in Arbuscular Mycorrhizal Fungi. Nature 2001, 414, 745–748. [Google Scholar] [CrossRef] [PubMed]
- Pawlowska, T.E.; Taylor, J.W. Organization of Genetic Variation in Individuals of Arbuscular Mycorrhizal Fungi. Nature 2004, 427, 733–737. [Google Scholar] [CrossRef] [PubMed]
- Wewer, V.; Brands, M.; Dörmann, P. Fatty Acid Synthesis and Lipid Metabolism in the Obligate Biotrophic Fungus Rhizophagus irregularis during Mycorrhization of Lotus japonicus. Plant J. 2014, 79, 398–412. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; San Clemente, H.; Roy, S.; Bécard, G.; Zhao, B.; Roux, C. A Survey of the Gene Repertoire of Gigaspora rosea Unravels Conserved Features among Glomeromycota for Obligate Biotrophy. Front. Microbiol. 2016, 7, 233. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Maeda, T.; Yamaguchi, K.; Kameoka, H.; Tanaka, S.; Ezawa, T.; Shigenobu, S.; Kawaguchi, M. The Genome of Rhizophagus clarus HR1 Reveals a Common Genetic Basis for Auxotrophy among Arbuscular Mycorrhizal Fungi. BMC Genom. 2018, 19, 465. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; Brands, M.; Wewer, V.; Dörmann, P.; Harrison, M.J. Arbuscular Mycorrhiza-Specific Enzymes FatM and RAM2 Fine-Tune Lipid Biosynthesis to Promote Development of Arbuscular Mycorrhiza. New Phytol. 2017, 214, 1631–1645. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, W.; Xie, Q.; Liu, N.; Liu, L.; Wang, D.; Zhang, X.; Yang, C.; Chen, X.; Tang, D.; et al. Plants Transfer Lipids to Sustain Colonization by Mutualistic Mycorrhizal and Parasitic Fungi. Science 2017, 356, 1172–1175. [Google Scholar] [CrossRef] [PubMed]
- Keymer, A.; Pimprikar, P.; Wewer, V.; Huber, C.; Brands, M.; Bucerius, S.L.; Delaux, P.-M.; Klingl, V.; von Röpenack-Lahaye, E.; Wang, T.L.; et al. Lipid Transfer from Plants to Arbuscular Mycorrhiza Fungi. eLife 2017, 6, e29107. [Google Scholar] [CrossRef] [PubMed]
- Luginbuehl, L.H.; Menard, G.N.; Kurup, S.; Van Erp, H.; Radhakrishnan, G.V.; Breakspear, A.; Oldroyd, G.E.D.; Eastmond, P.J. Fatty Acids in Arbuscular Mycorrhizal Fungi Are Synthesized by the Host Plant. Science 2017, 356, 1175–1178. [Google Scholar] [CrossRef] [PubMed]
- Kameoka, H.; Tsutsui, I.; Saito, K.; Kikuchi, Y.; Handa, Y.; Ezawa, T.; Hayashi, H.; Kawaguchi, M.; Akiyama, K. Stimulation of Asymbiotic Sporulation in Arbuscular Mycorrhizal Fungi by Fatty Acids. Nat. Microbiol. 2019, 4, 1654–1660. [Google Scholar] [CrossRef]
- Sugiura, Y.; Akiyama, R.; Tanaka, S.; Yano, K.; Kameoka, H.; Marui, S.; Saito, M.; Kawaguchi, M.; Akiyama, K.; Saito, K. Myristate Can Be Used as a Carbon and Energy Source for the Asymbiotic Growth of Arbuscular Mycorrhizal Fungi. Proc. Natl. Acad. Sci. USA 2020, 117, 25779–25788. [Google Scholar] [CrossRef] [PubMed]
- Sperschneider, J.; Yildirir, G.; Rizzi, Y.; Malar, C.M.; Sorwar, E.; Chen, E.C.; Iwasaki, W.; Brauer, E.K.; Bosnich, W.; Gutjahr, C.; et al. Resolving the Haplotypes of Arbuscular Mycorrhizal Fungi Highlights the Role of Two Nuclear Populations in Host Interactions. bioRxiv 2023. [Google Scholar] [CrossRef]
- Hughes, D.P.; Araújo, J.P.M.; Loreto, R.G.; Quevillon, L.; de Bekker, C.; Evans, H.C. From So Simple a Beginning: The Evolution of Behavioral Manipulation by Fungi. Adv. Genet. 2016, 94, 437–469. [Google Scholar] [CrossRef] [PubMed]
- Vilela, R.; Mendoza, L. Human Pathogenic Entomophthorales. Clin. Microbiol. Rev. 2018, 31, e00014-18. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, K.L.; Benny, G.L.; Ho, H.-M.; Smith, M.E. Phylogenetic Systematics of Syncephalis (Zoopagales, Zoopagomycotina), a Genus of Ubiquitous Mycoparasites. Mycologia 2017, 109, 333–349. [Google Scholar] [CrossRef]
- Lovett, B.; Macias, A.; Stajich, J.E.; Cooley, J.; Eilenberg, J.; de Fine Licht, H.H.; Kasson, M.T. Behavioral Betrayal: How Select Fungal Parasites Enlist Living Insects to Do Their Bidding. PLoS Pathog. 2020, 16, e1008598. [Google Scholar] [CrossRef]
- Chang, Y.; Wang, S.; Sekimoto, S.; Aerts, A.L.; Choi, C.; Clum, A.; LaButti, K.M.; Lindquist, E.A.; Yee Ngan, C.; Ohm, R.A.; et al. Phylogenomic Analyses Indicate That Early Fungi Evolved Digesting Cell Walls of Algal Ancestors of Land Plants. Genome Biol. Evol. 2015, 7, 1590–1601. [Google Scholar] [CrossRef]
- Wang, Y.; White, M.M.; Kvist, S.; Moncalvo, J.-M. Genome-Wide Survey of Gut Fungi (Harpellales) Reveals the First Horizontally Transferred Ubiquitin Gene from a Mosquito Host. Mol. Biol. Evol. 2016, 33, 2544–2554. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Stata, M.; Wang, W.; Stajich, J.E.; White, M.M.; Moncalvo, J.-M. Comparative Genomics Reveals the Core Gene Toolbox for the Fungus-Insect Symbiosis. mBio 2018, 9, e00636-18. [Google Scholar] [CrossRef]
- Tabima, J.F.; Trautman, I.A.; Chang, Y.; Wang, Y.; Mondo, S.; Kuo, A.; Salamov, A.; Grigoriev, I.V.; Stajich, J.E.; Spatafora, J.W. Phylogenomic Analyses of Non-Dikarya Fungi Supports Horizontal Gene Transfer Driving Diversification of Secondary Metabolism in the Amphibian Gastrointestinal Symbiont, Basidiobolus. G3 (Bethesda) 2020, 10, 3417–3433. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.-C.; Ho, H.-M.; Reynolds, N.; Smith, M.E.; Benny, G.L.; Chien, C.-Y.; Tsai, J.-L. Preliminary Phylogeny of Coemansia (Kickxellales), with Descriptions of Four New Species from Taiwan. Mycologia 2017, 109, 815–831. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; White, M.M.; Moncalvo, J.-M. Draft Genome Sequence of Capniomyces stellatus, the Obligate Gut Fungal Symbiont of Stonefly. Genome Announc. 2016, 4, e00761-16. [Google Scholar] [CrossRef]
- Tretter, E.D.; Johnson, E.M.; Benny, G.L.; Lichtwardt, R.W.; Wang, Y.; Kandel, P.; Novak, S.J.; Smith, J.F.; White, M.M. An Eight-Gene Molecular Phylogeny of the Kickxellomycotina, Including the First Phylogenetic Placement of Asellariales. Mycologia 2014, 106, 912–935. [Google Scholar] [CrossRef] [PubMed]
- Lichtward, R.W. The Trichomycetes, Fungal Associates of Arthropods, 2nd ed.; Springer Science & Business Media: New York, NY, USA; Berlin/Heidelberg, Germany; Tokio, Japan, 2012. [Google Scholar]
- Wang, Y.; White, M.M.; Moncalvo, J.-M. Diversification of the Gut Fungi Smittium and Allies (Harpellales) Co-Occurred with the Origin of Complete Metamorphosis of Their Symbiotic Insect Hosts (Lower Diptera). Mol. Phylogenet. Evol. 2019, 139, 106550. [Google Scholar] [CrossRef]
- White, M.M.; Guàrdia Valle, L.; Lichtwardt, R.W.; Siri, A.; Strongman, D.B.; William, R.T.; Gause, W.J.; Tretter, E.D. New Species and Emendations of Orphella: Taxonomic and Phylogenetic Reassessment of the Genus to Establish the Orphellales, for Stonefly Gut Fungi with a Twist. Mycologia 2018, 110, 147–178. [Google Scholar] [CrossRef]
- Wang, Y.; Tretter, E.D.; Lichtwardt, R.W.; White, M.M. Overview of 75 Years of Smittium Research, Establishing a New Genus for Smittium culisetae, and Prospects for Future Revisions of the “Smittium” Clade. Mycologia 2013, 105, 90–111. [Google Scholar] [CrossRef]
- White, M.M.; Siri, A.; Lichtwardt, R.W. Trichomycete Insect Symbionts in Great Smoky Mountains National Park and Vicinity. Mycologia 2006, 98, 333–352. [Google Scholar] [CrossRef]
- Mariotti, M.; Salinas, G.; Gabaldón, T.; Gladyshev, V.N. Utilization of Selenocysteine in Early-Branching Fungal Phyla. Nat. Microbiol. 2019, 4, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Yu, D.-S.; Wang, C.-F.; Liu, X.-Y.; Huang, B. A Taxonomic Revision of the Genus Conidiobolus (Ancylistaceae, Entomophthorales): Four Clades Including Three New Genera. MycoKeys 2020, 66, 55–81. [Google Scholar] [CrossRef] [PubMed]
- Humber, R.A. Synopsis of a Revised Classification for the Entomophthorales (Zygomycotina). Mycotaxon 1989, 34, 441–460. [Google Scholar]
- Nagahama, T.; Sato, H.; Shimazu, M.; Sugiyama, J. Phylogenetic Divergence of the Entomophthoralean Fungi: Evidence from Nuclear 18S Ribosomal RNA Gene Sequences. Mycologia 1995, 87, 203–209. [Google Scholar] [CrossRef]
- Bergman, K.; Eslava, A.P.; Cerdá-Olmedo, E. Mutants of Phycomyces with Abnormal Phototropism. Mol. Gen. Genet. 1973, 123, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Idnurm, A.; Rodríguez-Romero, J.; Corrochano, L.M.; Sanz, C.; Iturriaga, E.A.; Eslava, A.P.; Heitman, J. The Phycomyces MadA Gene Encodes a Blue-Light Photoreceptor for Phototropism and Other Light Responses. Proc. Natl. Acad. Sci. USA 2006, 103, 4546–4551. [Google Scholar] [CrossRef]
- Liu, Y.; He, Q.; Cheng, P. Photoreception in Neurospora: A Tale of Two White Collar Proteins. Cell. Mol. Life Sci. 2003, 60, 2131–2138. [Google Scholar] [CrossRef]
- Sanz, C.; Rodríguez-Romero, J.; Idnurm, A.; Christie, J.M.; Heitman, J.; Corrochano, L.M.; Eslava, A.P. Phycomyces MadB Interacts with MadA to Form the Primary Photoreceptor Complex for Fungal Phototropism. Proc. Natl. Acad. Sci. USA 2009, 106, 7095–7100. [Google Scholar] [CrossRef]
- Silva, F.; Torres-Martínez, S.; Garre, V. Distinct White Collar-1 Genes Control Specific Light Responses in Mucor circinelloides. Mol. Microbiol. 2006, 61, 1023–1037. [Google Scholar] [CrossRef]
- Polaino, S.; Villalobos-Escobedo, J.M.; Shakya, V.P.S.; Miralles-Durán, A.; Chaudhary, S.; Sanz, C.; Shahriari, M.; Luque, E.M.; Eslava, A.P.; Corrochano, L.M.; et al. A Ras GTPase Associated Protein Is Involved in the Phototropic and Circadian Photobiology Responses in Fungi. Sci. Rep. 2017, 7, 44790. [Google Scholar] [CrossRef]
- Verma, S.; Idnurm, A. The Uve1 Endonuclease Is Regulated by the White Collar Complex to Protect Cryptococcus neoformans from UV Damage. PLoS Genet. 2013, 9, e1003769. [Google Scholar] [CrossRef]
- Schimek, C.; Eibel, P.; Horie, T.; Galland, P.; Ootaki, T. Protein Crystals in Phycomyces Sporangiophores Are Involved in Graviperception. Adv. Space Res. 1999, 24, 687–696. [Google Scholar] [CrossRef]
- Nguyen, T.A.; Greig, J.; Khan, A.; Goh, C.; Jedd, G. Evolutionary Novelty in Gravity Sensing through Horizontal Gene Transfer and High-Order Protein Assembly. PLoS Biol. 2018, 16, e2004920. [Google Scholar] [CrossRef]
- Dupont, S.; Lemetais, G.; Ferreira, T.; Cayot, P.; Gervais, P.; Beney, L. Ergosterol Biosynthesis: A Fungal Pathway for Life on Land? Evolution 2012, 66, 2961–2968. [Google Scholar] [CrossRef]
- Pawlowska, T.E.; Gaspar, M.L.; Lastovetsky, O.A.; Mondo, S.J.; Real-Ramirez, I.; Shakya, E.; Bonfante, P. Biology of Fungi and Their Bacterial Endosymbionts. Annu. Rev. Phytopathol. 2018, 56, 289–309. [Google Scholar] [CrossRef] [PubMed]
- Bonfante, P.; Venice, F.; Lanfranco, L. The Mycobiota: Fungi Take Their Place between Plants and Bacteria. Curr. Opin. Microbiol. 2019, 49, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Okrasińska, A.; Bokus, A.; Duk, K.; Gęsiorska, A.; Sokołowska, B.; Miłobędzka, A.; Wrzosek, M.; Pawłowska, J. New Endohyphal Relationships between Mucoromycota and Burkholderiaceae Representatives. Appl. Environ. Microbiol. 2021, 87, e02707-20. [Google Scholar] [CrossRef] [PubMed]
- Bianciotto, V.; Lumini, E.; Bonfante, P.; Vandamme, P. “Candidatus Glomeribacter Gigasporarum” Gen. Nov., Sp. Nov., an Endosymbiont of Arbuscular Mycorrhizal Fungi. Int. J. Syst. Evol. Microbiol. 2003, 53, 121–124. [Google Scholar] [CrossRef]
- Lumini, E.; Bianciotto, V.; Jargeat, P.; Novero, M.; Salvioli, A.; Faccio, A.; Bécard, G.; Bonfante, P. Presymbiotic Growth and Sporal Morphology Are Affected in the Arbuscular Mycorrhizal Fungus Gigaspora margarita Cured of Its Endobacteria. Cell. Microbiol. 2007, 9, 1716–1729. [Google Scholar] [CrossRef]
- Salvioli, A.; Ghignone, S.; Novero, M.; Navazio, L.; Venice, F.; Bagnaresi, P.; Bonfante, P. Symbiosis with an Endobacterium Increases the Fitness of a Mycorrhizal Fungus, Raising Its Bioenergetic Potential. ISME J. 2016, 10, 130–144. [Google Scholar] [CrossRef]
- Dearth, S.P.; Castro, H.F.; Venice, F.; Tague, E.D.; Novero, M.; Bonfante, P.; Campagna, S.R. Metabolome Changes Are Induced in the Arbuscular Mycorrhizal Fungus Gigaspora margarita by Germination and by Its Bacterial Endosymbiont. Mycorrhiza 2018, 28, 421–433. [Google Scholar] [CrossRef]
- Ohshima, S.; Sato, Y.; Fujimura, R.; Takashima, Y.; Hamada, M.; Nishizawa, T.; Narisawa, K.; Ohta, H. Mycoavidus cysteinexigens Gen. Nov., Sp. Nov., an Endohyphal Bacterium Isolated from a Soil Isolate of the Fungus Mortierella elongata. Int. J. Syst. Evol. Microbiol. 2016, 66, 2052–2057. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yao, Q.; Dearth, S.P.; Entler, M.R.; Castro Gonzalez, H.F.; Uehling, J.K.; Vilgalys, R.J.; Hurst, G.B.; Campagna, S.R.; Labbé, J.L.; et al. Integrated Proteomics and Metabolomics Suggests Symbiotic Metabolism and Multimodal Regulation in a Fungal-Endobacterial System. Environ. Microbiol. 2017, 19, 1041–1053. [Google Scholar] [CrossRef]
- Takashima, Y.; Seto, K.; Degawa, Y.; Guo, Y.; Nishizawa, T.; Ohta, H.; Narisawa, K. Prevalence and Intra-Family Phylogenetic Divergence of Burkholderiaceae-Related Endobacteria Associated with Species of Mortierella. Microbes Environ. 2018, 33, 417–427. [Google Scholar] [CrossRef]
- Büttner, H.; Niehs, S.P.; Vandelannoote, K.; Cseresnyés, Z.; Dose, B.; Richter, I.; Gerst, R.; Figge, M.T.; Stinear, T.P.; Pidot, S.J.; et al. Bacterial Endosymbionts Protect Beneficial Soil Fungus from Nematode Attack. Proc. Natl. Acad. Sci. USA 2021, 118, e2110669118. [Google Scholar] [CrossRef]
- Mondo, S.J.; Lastovetsky, O.A.; Gaspar, M.L.; Schwardt, N.H.; Barber, C.C.; Riley, R.; Sun, H.; Grigoriev, I.V.; Pawlowska, T.E. Bacterial Endosymbionts Influence Host Sexuality and Reveal Reproductive Genes of Early Divergent Fungi. Nat. Commun. 2017, 8, 1843. [Google Scholar] [CrossRef]
- Richter, I.; Radosa, S.; Cseresnyés, Z.; Ferling, I.; Büttner, H.; Niehs, S.P.; Gerst, R.; Scherlach, K.; Figge, M.T.; Hillmann, F.; et al. Toxin-Producing Endosymbionts Shield Pathogenic Fungus against Micropredators. mBio 2022, 13, e01440-22. [Google Scholar] [CrossRef] [PubMed]
- Itabangi, H.; Sephton-Clark, P.C.S.; Tamayo, D.P.; Zhou, X.; Starling, G.P.; Mahamoud, Z.; Insua, I.; Probert, M.; Correia, J.; Moynihan, P.J.; et al. A Bacterial Endosymbiont of the Fungus Rhizopus microsporus Drives Phagocyte Evasion and Opportunistic Virulence. Curr. Biol. 2022, 32, 1115–1130.e6. [Google Scholar] [CrossRef]
- Partida-Martinez, L.P.; Hertweck, C. A Gene Cluster Encoding Rhizoxin Biosynthesis in “Burkholderia rhizoxina”, the Bacterial Endosymbiont of the Fungus Rhizopus microsporus. Chembiochem 2007, 8, 41–45. [Google Scholar] [CrossRef]
- Estrada-de Los Santos, P.; Palmer, M.; Chávez-Ramírez, B.; Beukes, C.; Steenkamp, E.T.; Briscoe, L.; Khan, N.; Maluk, M.; Lafos, M.; Humm, E.; et al. Whole Genome Analyses Suggests That Burkholderia sensu lato Contains Two Additional Novel Genera (Mycetohabitans gen. nov., and Trinickia gen. nov.): Implications for the Evolution of Diazotrophy and Nodulation in the Burkholderiaceae. Genes 2018, 9, 389. [Google Scholar] [CrossRef] [PubMed]
- Lastovetsky, O.A.; Krasnovsky, L.D.; Qin, X.; Gaspar, M.L.; Gryganskyi, A.P.; Huntemann, M.; Clum, A.; Pillay, M.; Palaniappan, K.; Varghese, N.; et al. Molecular Dialogues between Early Divergent Fungi and Bacteria in an Antagonism versus a Mutualism. mBio 2020, 11, e02088-20. [Google Scholar] [CrossRef]
- Naumann, M.; Schüßler, A.; Bonfante, P. The Obligate Endobacteria of Arbuscular Mycorrhizal Fungi Are Ancient Heritable Components Related to the Mollicutes. ISME J. 2010, 4, 862–871. [Google Scholar] [CrossRef]
- Toomer, K.H.; Chen, X.; Naito, M.; Mondo, S.J.; den Bakker, H.C.; VanKuren, N.W.; Lekberg, Y.; Morton, J.B.; Pawlowska, T.E. Molecular Evolution Patterns Reveal Life History Features of Mycoplasma-Related Endobacteria Associated with Arbuscular Mycorrhizal Fungi. Mol. Ecol. 2015, 24, 3485–3500. [Google Scholar] [CrossRef]
- Desirò, A.; Hao, Z.; Liber, J.A.; Benucci, G.M.N.; Lowry, D.; Roberson, R.; Bonito, G. Mycoplasma-Related Endobacteria within Mortierellomycotina Fungi: Diversity, Distribution and Functional Insights into Their Lifestyle. ISME J. 2018, 12, 1743–1757. [Google Scholar] [CrossRef]
- Desirò, A.; Faccio, A.; Kaech, A.; Bidartondo, M.I.; Bonfante, P. Endogone, One of the Oldest Plant-Associated Fungi, Host Unique Mollicutes-Related Endobacteria. New Phytol. 2015, 205, 1464–1472. [Google Scholar] [CrossRef]
- Naito, M.; Desirò, A.; González, J.B.; Tao, G.; Morton, J.B.; Bonfante, P.; Pawlowska, T.E. “Candidatus Moeniiplasma Glomeromycotorum”, an Endobacterium of Arbuscular Mycorrhizal Fungi. Int. J. Syst. Evol. Microbiol. 2017, 67, 1177–1184. [Google Scholar] [CrossRef]
- Naito, M.; Morton, J.B.; Pawlowska, T.E. Minimal Genomes of Mycoplasma-Related Endobacteria Are Plastic and Contain Host-Derived Genes for Sustained Life within Glomeromycota. Proc. Natl. Acad. Sci. USA 2015, 112, 7791–7796. [Google Scholar] [CrossRef] [PubMed]
- Robinson, A.J.; House, G.L.; Morales, D.P.; Kelliher, J.M.; Gallegos-Graves, L.V.; LeBrun, E.S.; Davenport, K.W.; Palmieri, F.; Lohberger, A.; Bregnard, D.; et al. Widespread Bacterial Diversity within the Bacteriome of Fungi. Commun. Biol. 2021, 4, 1168. [Google Scholar] [CrossRef] [PubMed]
- Telagathoti, A.; Probst, M.; Peintner, U. Habitat, Snow-Cover and Soil PH, Affect the Distribution and Diversity of Mortierellaceae Species and Their Associations to Bacteria. Front. Microbiol. 2021, 12, 669784. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, C.L.; Lovett, B.; Kasson, M.T.; Stajich, J.E. Metagenome-Assembled Genomes of Bacteria Associated with Massospora cicadina Fungal Plugs from Infected Brood VIII Periodical Cicadas. Microbiol. Resour. Announc. 2022, 11, e00413-22. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Wang, Y.; Mondo, S.; Ahrendt, S.; Andreopoulos, W.; Barry, K.; Beard, J.; Benny, G.L.; Blankenship, S.; Bonito, G.; et al. Evolution of Zygomycete Secretomes and the Origins of Terrestrial Fungal Ecologies. iScience 2022, 25, 104840. [Google Scholar] [CrossRef] [PubMed]
- Shelest, E.; Voigt, K. 2 Genomics to Study Basal Lineage Fungal Biology: Phylogenomics Suggests a Common Origin. In Fungal Genomics; The Mycota; Springer: Berlin/Heidelberg, Germany, 2014; Volume XIII, pp. 31–60. ISBN 978-3-642-45217-8. [Google Scholar]
- Voigt, K.; Wolf, T.; Ochsenreiter, K.; Nagy, G.; Kaerger, K.; Shelest, E.; Papp, T. 15 Genetic and Metabolic Aspects of Primary and Secondary Metabolism of the Zygomycetes. In Biochemistry and Molecular Biology; The Mycota; Springer: Berlin/Heidelberg, Germany, 2016; Volume III, pp. 361–385. ISBN 978-3-319-27788-2. [Google Scholar]
- Koczyk, G.; Pawłowska, J.; Muszewska, A. Terpenoid Biosynthesis Dominates among Secondary Metabolite Clusters in MUcoromycotina Genomes. J. Fungi 2021, 7, 285. [Google Scholar] [CrossRef]
- Baldeweg, F.; Warncke, P.; Fischer, D.; Gressler, M. Fungal Biosurfactants from Mortierella alpina. Org. Lett. 2019, 21, 1444–1448. [Google Scholar] [CrossRef]
- Lebreton, A.; Meslet-Cladière, L.; Morin-Sardin, S.; Coton, E.; Jany, J.-L.; Barbier, G.; Corre, E. Comparative Analysis of Five Mucor Species Transcriptomes. Genomics 2019, 111, 1306–1314. [Google Scholar] [CrossRef] [PubMed]
- Winkelmann, G. A Search for Glomuferrin: A Potential Siderophore of Arbuscular Mycorrhizal Fungi of the Genus Glomus. Biometals 2017, 30, 559–564. [Google Scholar] [CrossRef]
- Prakash, H.; Rudramurthy, S.M.; Gandham, P.S.; Ghosh, A.K.; Kumar, M.M.; Badapanda, C.; Chakrabarti, A. Apophysomyces variabilis: Draft Genome Sequence and Comparison of Predictive Virulence Determinants with Other Medically Important Mucorales. BMC Genom. 2017, 18, 736. [Google Scholar] [CrossRef] [PubMed]
- Carroll, C.S.; Grieve, C.L.; Murugathasan, I.; Bennet, A.J.; Czekster, C.M.; Liu, H.; Naismith, J.; Moore, M.M. The Rhizoferrin Biosynthetic Gene in the Fungal Pathogen Rhizopus delemar Is a Novel Member of the NIS Gene Family. Int. J. Biochem. Cell Biol. 2017, 89, 136–146. [Google Scholar] [CrossRef]
- Škríba, A.; Patil, R.H.; Hubáček, P.; Dobiáš, R.; Palyzová, A.; Marešová, H.; Pluháček, T.; Havlíček, V. Rhizoferrin Glycosylation in Rhizopus microsporus. J. Fungi 2020, 6, 89. [Google Scholar] [CrossRef]
- Takeda, I.; Tamano, K.; Yamane, N.; Ishii, T.; Miura, A.; Umemura, M.; Terai, G.; Baker, S.E.; Koike, H.; Machida, M. Genome Sequence of the Mucoromycotina Fungus Umbelopsis isabellina, an Effective Producer of Lipids. Genome Announc. 2014, 2, e00071-14. [Google Scholar] [CrossRef]
- Gluck-Thaler, E.; Ralston, T.; Konkel, Z.; Ocampos, C.G.; Ganeshan, V.D.; Dorrance, A.E.; Niblack, T.L.; Wood, C.W.; Slot, J.C.; Lopez-Nicora, H.D.; et al. Giant Starship Elements Mobilize Accessory Genes in Fungal Genomes. Mol. Biol. Evol. 2022, 39, msac109. [Google Scholar] [CrossRef]
- Urquhart, A.S.; Chong, N.F.; Yang, Y.; Idnurm, A. A Large Transposable Element Mediates Metal Resistance in the Fungus Paecilomyces variotii. Curr. Biol. 2022, 32, 937–950.e5. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gryganskyi, A.P.; Golan, J.; Muszewska, A.; Idnurm, A.; Dolatabadi, S.; Mondo, S.J.; Kutovenko, V.B.; Kutovenko, V.O.; Gajdeczka, M.T.; Anishchenko, I.M.; et al. Sequencing the Genomes of the First Terrestrial Fungal Lineages: What Have We Learned? Microorganisms 2023, 11, 1830. https://doi.org/10.3390/microorganisms11071830
Gryganskyi AP, Golan J, Muszewska A, Idnurm A, Dolatabadi S, Mondo SJ, Kutovenko VB, Kutovenko VO, Gajdeczka MT, Anishchenko IM, et al. Sequencing the Genomes of the First Terrestrial Fungal Lineages: What Have We Learned? Microorganisms. 2023; 11(7):1830. https://doi.org/10.3390/microorganisms11071830
Chicago/Turabian StyleGryganskyi, Andrii P., Jacob Golan, Anna Muszewska, Alexander Idnurm, Somayeh Dolatabadi, Stephen J. Mondo, Vira B. Kutovenko, Volodymyr O. Kutovenko, Michael T. Gajdeczka, Iryna M. Anishchenko, and et al. 2023. "Sequencing the Genomes of the First Terrestrial Fungal Lineages: What Have We Learned?" Microorganisms 11, no. 7: 1830. https://doi.org/10.3390/microorganisms11071830
APA StyleGryganskyi, A. P., Golan, J., Muszewska, A., Idnurm, A., Dolatabadi, S., Mondo, S. J., Kutovenko, V. B., Kutovenko, V. O., Gajdeczka, M. T., Anishchenko, I. M., Pawlowska, J., Tran, N. V., Ebersberger, I., Voigt, K., Wang, Y., Chang, Y., Pawlowska, T. E., Heitman, J., Vilgalys, R., ... Stajich, J. E. (2023). Sequencing the Genomes of the First Terrestrial Fungal Lineages: What Have We Learned? Microorganisms, 11(7), 1830. https://doi.org/10.3390/microorganisms11071830