

Beneficial Effect of the New Leptodophora sp. Strain on Development of Blueberry Microclones in the Process of Their Adaptation

 , , , , and
, , , , and 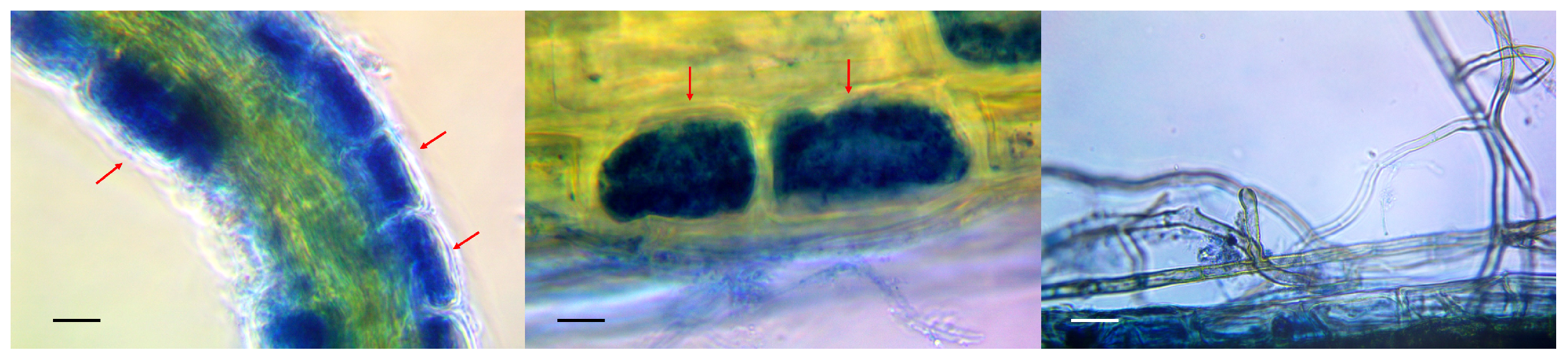{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
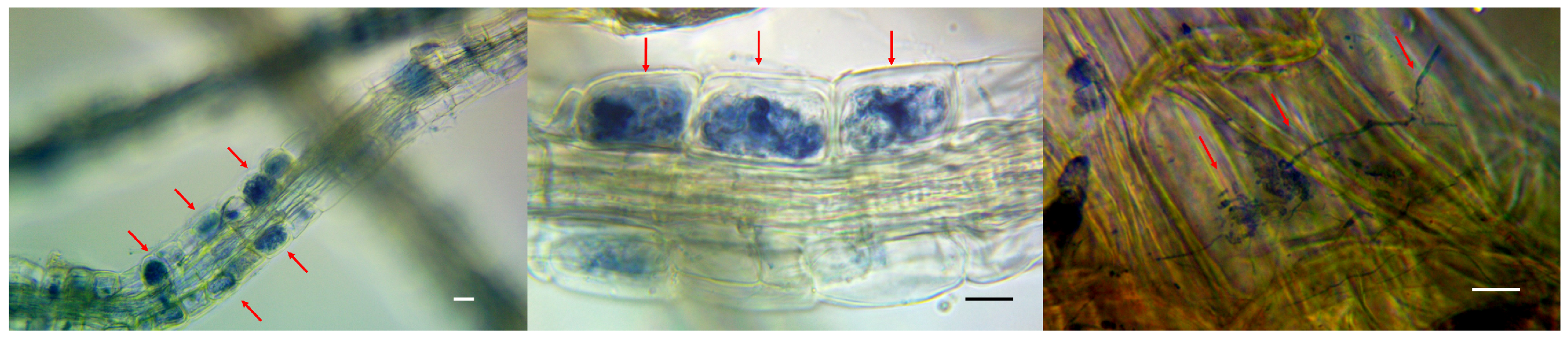
2.1. Collection of Primary Hosts and Identification of Mycorrhiza
2.2. Obtaining Pure Cultures of Micromycetes
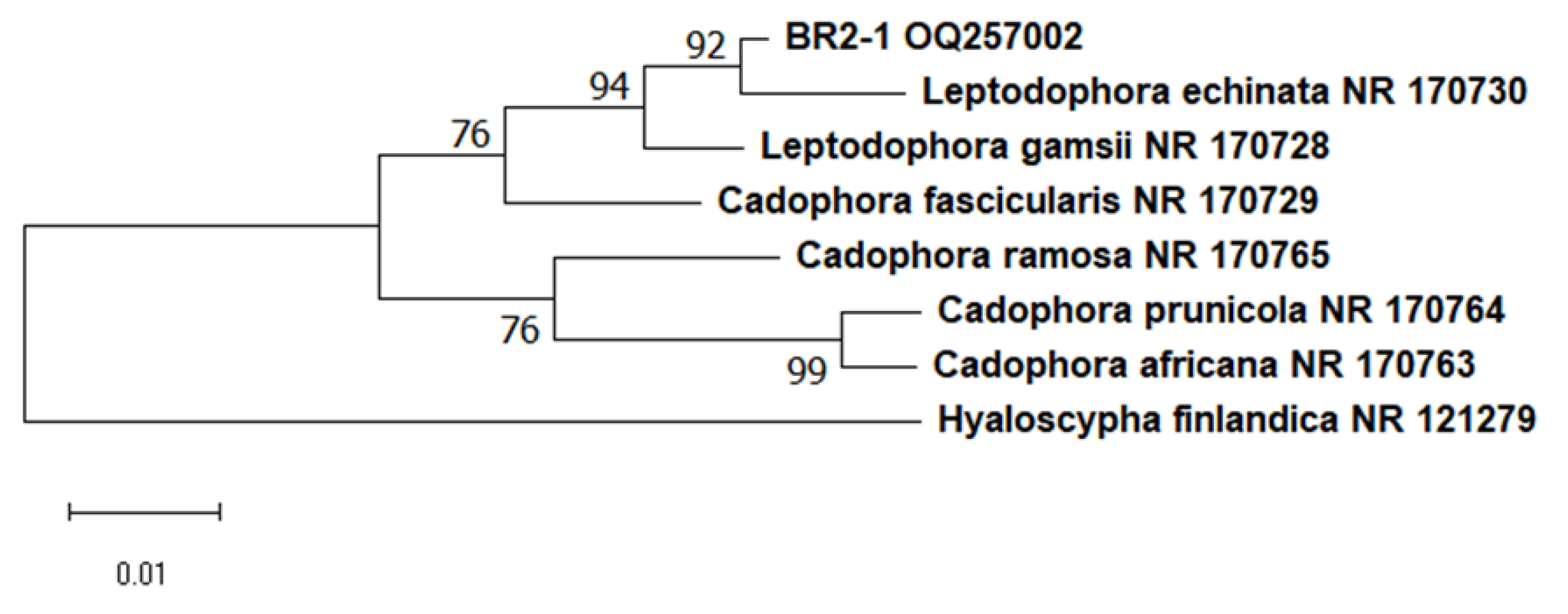
2.3. Molecular Identification
2.4. Cultivation on Different Substrates
2.5. Adaptation of Microclones of Sterile Host Plants
2.6. Collection and Analysis of Data on Vaccinium corymbosum L. Microclones
3. Results and Discussion
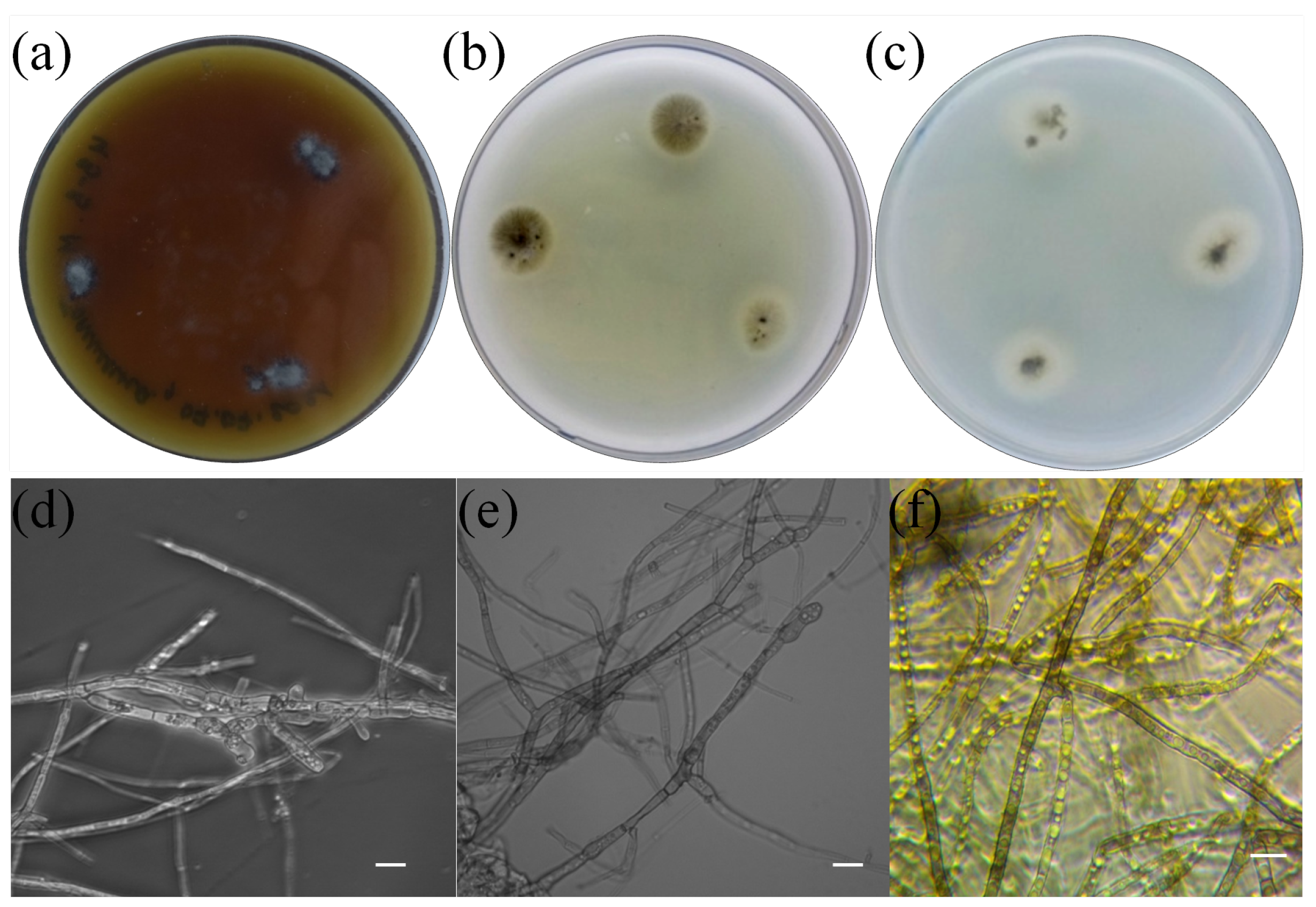
3.1. Isolation and Identification of Pure Culture
3.2. Commercial Cultivation
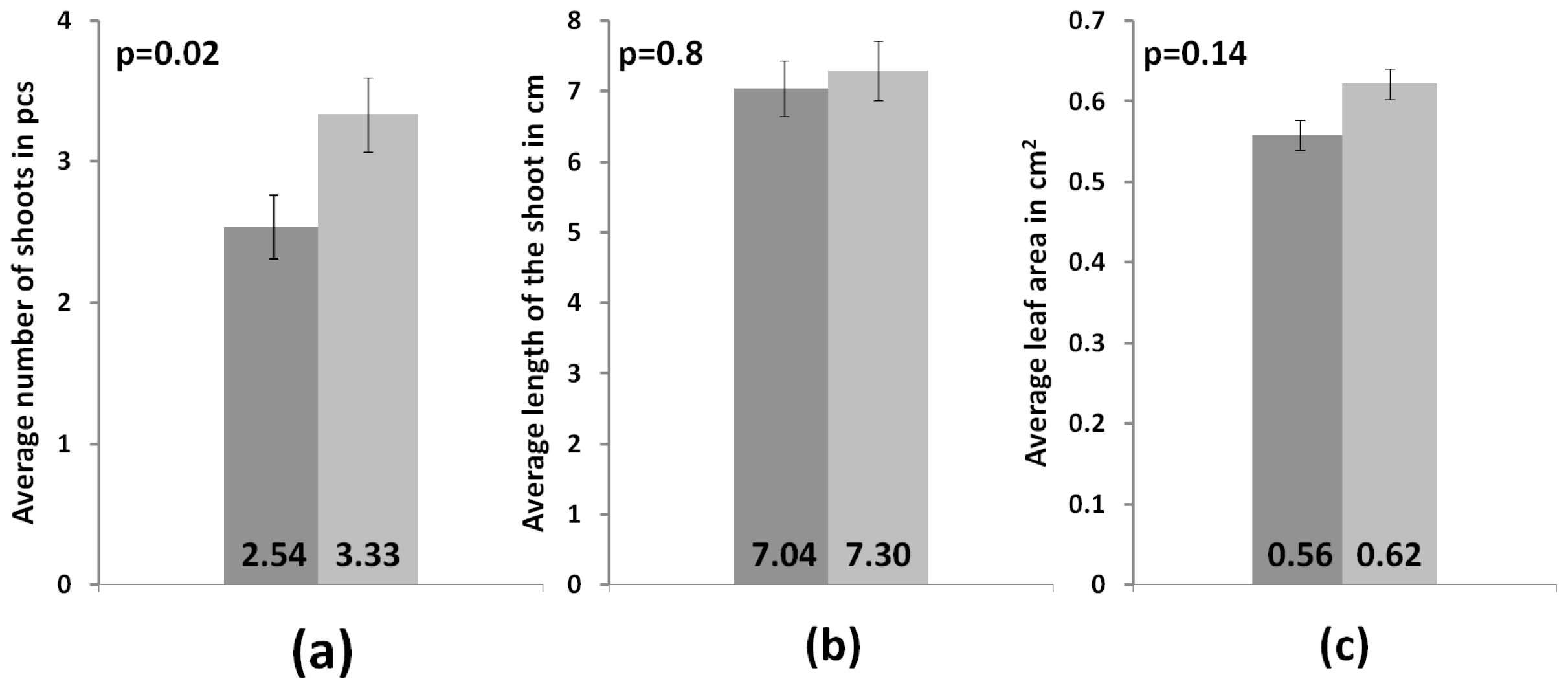
3.3. Development of Microclones during Adaptation
3.4. Effect of BR2-1 Isolate on Agricultural Crops
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ErM | Ericoid mycorrhizal |
| AM | Arbuscular mycorrhizal |
References
- Doronkin, V.M.; Kovtonyuk, N.K.A. (Eds.) Pyrolaceae—Lamiaceae (Labiatae). In Flora of Siberia; Nauka: Novosibirsk, Russia, 1997; p. 296. [Google Scholar]
- Nancy, C.J.; Catherine, A.G.; Cardo, n.Z.; Whitbeck, J.K. Mycorrhizas: Symbiotic Mediators of Rhizosphere and Ecosystem Processes. Rhizosphere 2007, 18, 73–100. [Google Scholar] [CrossRef]
- Mu, D.; Du, N.; Zwiazek, J.J. Inoculation with Ericoid Mycorrhizal Associations Alleviates Drought Stress in Lowland and Upland Velvetleaf Blueberry (Vaccinium myrtilloides) Seedlings. Plants 2021, 10, 2786. [Google Scholar] [CrossRef] [PubMed]
- Xiangying, W.; Jianjun, C.; Chunying, Z.; Hong, L. Ericoid mycorrhizal fungus enhances microcutting rooting of Rhododendron fortunei and subsequent growth. Hortic. Res. 2020, 7, 831–846. [Google Scholar] [CrossRef]
- Vohnik, M. Ericoid mycorrhizal symbiosis: Theoretical background and methods for its comprehensive investigation. Mycorrhiza 2020, 30, 671–695. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorhizal Symbiosis; Smith, S.E., Read, D.J., Eds.; Academic Press: New York, NY, USA, 2008; p. 1012. [Google Scholar] [CrossRef]
- Vralstad, T.; Myhre, E.; Schumacher, T. Molecular diversity and phylogenetic affinities of symbiotic root-associated ascomycetes of the Helotiales in burnt and metal polluted habitats. New Phytol. 2002, 155, 131–148. [Google Scholar] [CrossRef]
- Kohout, P. Biogeography, of Ericoid Mycorrhiza. In Biogeography of Mycorrhizal Symbiosis.; Tedersoo, L., Ed.; Springer: Cham, Switzerland, 2017; pp. 179–193. [Google Scholar] [CrossRef]
- Daghino, S.; Martino, E.; Perotto, S.V.S. Metabarcoding of fungal assemblages in Vaccinium myrtillus endosphere suggests colonization of above-ground organs by some ericoid mycorrhizal and DSE fungi. Sci. Rep. 2022, 12, 11013. [Google Scholar] [CrossRef]
- Hazard, C.; Gosling, P.; Mitchell, D.T.; Doohan, F.M.; Bending, G.D. Diversity of fungi associated with hair roots of ericaceous plants is affected by land use. FEMS iMcrobiol. Ecol. 2014, 87, 586–600. [Google Scholar] [CrossRef]
- Selosse, M.A.; Setaro, S.; Glatard, F.; Richard, F.; Urcelay, C.; Weiß, M. Sebacinales are common mycorrhizal associates of Ericaceae. New Phytol. 2007, 174, 864–878. [Google Scholar] [CrossRef]
- Allen, T.R.; Millar, T.; Berch, S.M.; Berbee, M.L. Culturing and direct DNA extraction find different fungi from the same ericoid mycorrhizal roots. New Phytol. 2003, 160, 255–272. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Zhuang, W.Y. Phylogenetic relationships of some members in the genus Hymenoscyphus (Ascomycetes, Helotiales). Nova Hedwig. 2004, 78, 475–484. [Google Scholar] [CrossRef]
- Walker, J.F.; Aldrich-Wolfe, L.; Riffel, A.; Barbare, H.; Simpson, N.B.; Trowbridge, J.; Jumpponen, A. Diverse Helotiales associated with the roots of three species of Arctic Ericaceae provide no evidence for host specificity. New Phytol. 2011, 191, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Fehrer, J.; Reblova, M.; Bambasova, V.; Vohnik, M. The root-symbiotic Rhizoscyphus ericae aggregate and Hyaloscypha (Leotiomycetes) are congeneric: Phylogenetic and experimental evidence. Stud. Mycol. 2019, 92, 195–225. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Partel, K.; Jairus, T.; Gates, G. Ascomycetes associated with ectomycorrhizas: Molecular diversity and ecology with particular reference to the Helotiales. Environ. Microbiol. 2009, 11, 3166–3178. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.K.; Dai, C.C.; Liu, X.Z. Mechanisms of fungal endophytes in plant protection against pathogens. Afr. J. Microbiol. Res. 2010, 4, 1346–1351. [Google Scholar]
- Domsch, K.H.; Gams, W.; Anderson, T.H. Compendium of Soil Fungi. Eur. J. Soil Sci. 2007, 59, 1007. [Google Scholar]
- Berch, S.M.; Allen, T.R.; Berbee, M.L. Molecular detection, community structure and phylogency of ericoid mycorrhizal fungi. Plant. Soil. 2002, 244, 55–66. [Google Scholar] [CrossRef]
- Koukol, O.; Macia-Vicente, J.G. Leptodophora gen. nov. (Helotiales, Leotiomycetes) proposed to accommodate selected root-associated members of the genus Cadophora. Czech Mycol. 2022, 74, 57–66. [Google Scholar] [CrossRef]
- Litwinczuk, W. Micropropagation of Vaccinium sp. by in vitro axillary shoot proliferation. Methods Mol. Biol. 2013, 994, 63–76. [Google Scholar]
- Chizhik, O.V.; Reshetnikov, V.N.; Filipenya, V.L. Influence of microorganisms on the adaptation of cloned planting material of tree and shrub species of the genus Vaccinium. Physiol. Biochem. Cultiv. Plants 2013, 45, 254–259. [Google Scholar]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and straining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Smith, S.E.; Dickson, S. VA Mycorrhizas: Basic Research Techniques, 1st ed.; Cooperative Research Centre for Soil and Land Management: Adelaide, Australia, 1997. [Google Scholar]
- Jansa, J.; Vosatka, M. In vitro and post vitro inoculation of micropropagated Rhododendrons with ericoid mycorrhizal fungi. Appl. Soil Ecol. 2000, 15, 125–136. [Google Scholar] [CrossRef]
- Makarov, S.S.; Kuznetsova, I.B.; Smirnov, V.S. Influence of sterilization methods and types of blue honeysuckle explants on their viability under in vitro conditions. Lesokhoz. Inform. Electron. Netw. Mag. 2018, 2, 96–101. Available online: http://lhi.vniilm.ru (accessed on 30 January 2023).
- Blagoveshchenskaya, E.Y. Mycological Research. Fundamentals of Laboratory Technology: A Tutorial, 3rd ed.; LENAND: Moscow, Russia, 2021. [Google Scholar]
- Sambrook, J.; Russel, D.W. Purification of nucleic acids by extraction with phenol: Chloroform. Cold Spring Harb. Protoc. 2001, 42, pdb.prot4455. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S. Nuclear Ribosomal Internal Transcribed Spacer (ITS) Region as a Universal DNA Barcode Marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, D.T.; Lee, B.S.; Taylor, W.J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Singha, I.M.; Kakoty, Y.; Unni, B.G.; Das, J.; Kalita, M.C. Identification and characterization of Fusarium sp. using ITS and RAPD causing fusarium wilt of tomato isolated from Assam, North East India. J. Genet. Eng. Biotechnol. 2016, 14, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russel, D.W. Agarose Gel Electrophoresis. Cold Spring Harb. Protoc. 2006, 2006, pdb.prot4020. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Thompson, J.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F. The CLUSTAL X Windows Interface: Flexible Strategies for Multiple Sequence Alignment Aided by Quality Analysis Tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Dorofeeva, M.M.; Bonetskaya, S.A. Comparative analysis of some classical and modern methods for determining the area of the leaf surface. Plant Resour. 2020, 56, 182–192. [Google Scholar] [CrossRef]
- Mann, H.B.; Whitney, D.R. On a test of whether one of two random variables is stochastically larger than the other. Ann. Math. Stat. 1947, 18, 50–60. [Google Scholar] [CrossRef]
- Scagel, C.F.; Yang, W.Q. Cultural Variation and Mycorrhizal Status of Blueberry Plants in NW Oregon Commercial Production Fields. Int. J. Fruit Sci. 2005, 5, 85–111. [Google Scholar] [CrossRef]
- Scagel, C.F. Mycorrhizal Status of Sand-Based Cranberry (Vaccinium macrocarpon) Bogs in Southern Oregon. Small Fruits Rev. 2003, 2, 31–41. [Google Scholar] [CrossRef]
- Johansson, M. The influence of ammonium nitrate on the root growth and ericoid mycorrhizal colonization of Calluna vulgaris (L.) Hull from a Danish heathland. Oecologia 2000, 123, 418–424. [Google Scholar] [CrossRef]
- Stalpers, J.A. Identification of wood-inhabiting Aphyllophorales in pure culture. Stud. Mycol. 1978, 16, 248. [Google Scholar]
- Macia-Vicente, J.G.; Piepenbring, M.; Koukol, O. Brassicaceous roots as an unexpected diversity hot-spot of helotialean endophytes. IMA Fungus 2020, 11, 16. [Google Scholar] [CrossRef]
- Glynou, K.; Ali, T.; Buch, A.K.; Haghi Kia, S.; Ploch, S.; Xia, X.; Celik, A.; Thines, M.; Macia-Vicente, J.G. Continental-scale distribution of root endophytes. Environ. Microbiol. 2016, 18, 2418–2434. [Google Scholar] [CrossRef]
- Bizabani, C.; Dames, J. Effects of inoculating Lachnum and Cadophora isolates on the growth of Vaccinium corymbosum. Microbiol. Res. 2015, 181, 68–74. [Google Scholar] [CrossRef]
- Golldack, J.; Schubert, P.; Tauschke, M.; Schwarzel, H.; Hofflich, G.; Lentzsch, P.; Munzenberger, B. Mycorrhization and plant growth of highbush blueberry (Vaccinium corymbosum L.) on arable land in Germany. In Proceedings of the 3rd International Conference on Mycorrhiza, Adelaide, Australia, 8–13 July 2001. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Topilina, Y.S.; Luk‘yanova, E.A.; Glukhova, L.B.; Shurupova, M.N.; Gerasimchuk, A.L.; Frank, Y.A.; Antsiferov, D.V. Beneficial Effect of the New Leptodophora sp. Strain on Development of Blueberry Microclones in the Process of Their Adaptation. Microorganisms 2023, 11, 1406. https://doi.org/10.3390/microorganisms11061406
Topilina YS, Luk‘yanova EA, Glukhova LB, Shurupova MN, Gerasimchuk AL, Frank YA, Antsiferov DV. Beneficial Effect of the New Leptodophora sp. Strain on Development of Blueberry Microclones in the Process of Their Adaptation. Microorganisms. 2023; 11(6):1406. https://doi.org/10.3390/microorganisms11061406
Chicago/Turabian StyleTopilina, Yulia S., Evgeniya A. Luk‘yanova, Lubov B. Glukhova, Margarita N. Shurupova, Anna L. Gerasimchuk, Yulia A. Frank, and Dmitry V. Antsiferov. 2023. "Beneficial Effect of the New Leptodophora sp. Strain on Development of Blueberry Microclones in the Process of Their Adaptation" Microorganisms 11, no. 6: 1406. https://doi.org/10.3390/microorganisms11061406
APA StyleTopilina, Y. S., Luk‘yanova, E. A., Glukhova, L. B., Shurupova, M. N., Gerasimchuk, A. L., Frank, Y. A., & Antsiferov, D. V. (2023). Beneficial Effect of the New Leptodophora sp. Strain on Development of Blueberry Microclones in the Process of Their Adaptation. Microorganisms, 11(6), 1406. https://doi.org/10.3390/microorganisms11061406