
Absence of Hepatitis E Virus (HEV) Circulation in the Most Widespread Wild Croatian Canine Species, the Red Fox (Vulpes vulpes) and Jackal (Canis aureus moreoticus)





Abstract
1. Introduction
2. Materials and Methods
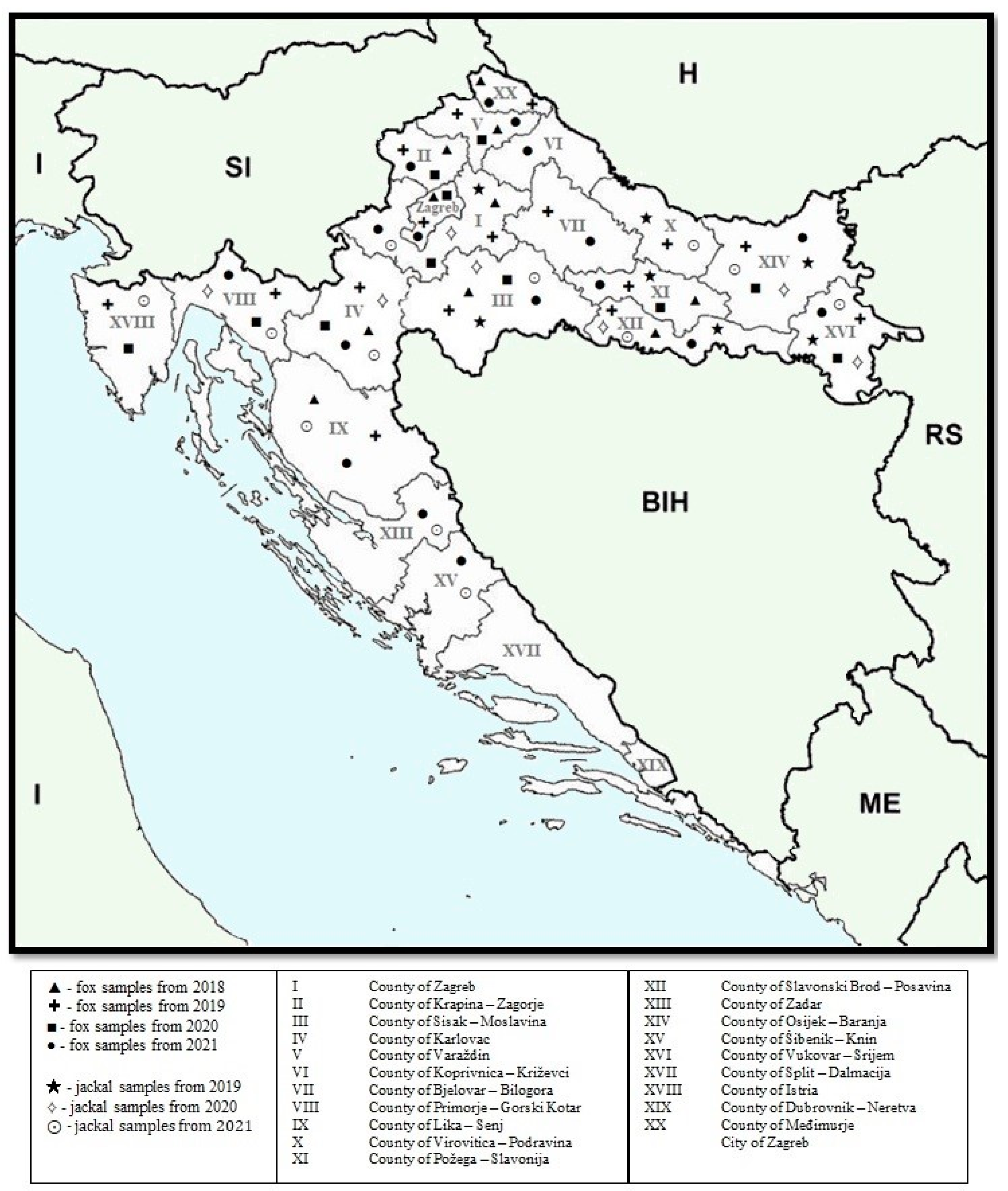
2.1. Sample Collection and Preparation
2.2. RNA Extraction and HEV Detection through Real-Time RT-PCR
2.3. Detection of Specific Anti-HEV Antibodies through Enzyme-Linked Immunosorbent Assay (ELISA)
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Hepatitis E. Available online: https://www.who.int/news-room/fact-sheets/detail/hepatitis-e (accessed on 26 January 2023).
- Khuroo, M.S. Discovery of hepatitis E: The epidemic non-A, non-B hepatitis 30 years down the memory lane. Virus Res. 2011, 161, 3–14. [Google Scholar]
- Emerson, S.U.; Purcell, R.H. Hepatitis E virus. Rev. Med. Virol. 2002, 13, 145–154. [Google Scholar] [CrossRef]
- Okamoto, H. Genetic variability and evolution of hepatitis E virus. Virus Res. 2007, 127, 216–228. [Google Scholar] [CrossRef]
- Tam, A.W.; Smith, M.M.; Guerra, M.E.; Huang, C.C.; Bradley, D.W.; Fry, K.E.; Reyes, G.R. Hepatitis E virus (HEV): Molecular cloning and sequencing of the full-length viral genome. Virology 1991, 185, 120–131. [Google Scholar] [CrossRef]
- LeDesma, R.; Nimgaonkar, I.; Ploss, A. Hepatitis E virus replication. Viruses 2019, 11, 719. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses: ICTV. Available online: https://ictv.global/taxonomy (accessed on 26 January 2023).
- Purdy, M.A.; Harrison, T.J.; Jameel, S.; Meng, X.-J.; Okamoto, H.; Van Der Poel, W.H.M.; Smith, D.B. ICTV Report Consortium ICTV Virus Taxonomy Profile: Hepeviridae. J. Gen. Virol. 2017, 98, 2645–2646. [Google Scholar] [CrossRef]
- Takova, K.; Koynarski, T.; Minkov, I.; Ivanova, Z.; Toneva, V.; Zahmanova, G. Increasing Hepatitis E Virus Seroprevalence in Domestic Pigs and Wild Boar in Bulgaria. Animals 2020, 10, 1521. [Google Scholar] [CrossRef]
- Spahr, C.; Knauf-Witzens, T.; Vahlenkamp, T.; Ulrich, R.G.; Johne, R. Hepatitis E virus and related viruses in wild, domestic and zoo animals: A review. Zoonoses Public Health 2017, 65, 11–29. [Google Scholar] [CrossRef]
- Ryll, R.; Heckel, G.; Corman, V.M.; Drexler, J.F.; Ulrich, R.G. Genomic and spatial variability of a European common vole hepevirus. Arch. Virol. 2019, 164, 2671–2682. [Google Scholar] [CrossRef]
- Kenney, S.P.; Meng, X.-J. Hepatitis E Virus Genome Structure and Replication Strategy. Cold Spring Harb. Perspect. Med. 2018, 9, a031724. [Google Scholar] [CrossRef]
- Van der Poel, W.H. Food and environmental routes of hepatitis E virus transmission. Curr. Opin. Virol. 2014, 4, 91–96. [Google Scholar] [CrossRef]
- Yugo, D.M.; Meng, X.-J. Hepatitis E virus: Foodborne, waterborne and zoonotic transmission. Int. J. Environ. Res. Public Health 2013, 10, 4507–4533. [Google Scholar] [CrossRef]
- Andraud, M.; Dumarest, M.; Cariolet, R.; Aylaj, B.; Barnaud, E.; Eono, F.; Pavio, N.; Rose, N. Direct contact and environmental contaminations are responsible for HEV transmission in pigs. Vet. Res. 2013, 44, 102. [Google Scholar] [CrossRef] [PubMed]
- Kasorndorkbua, C.; Guenette, D.K.; Huang, F.F.; Thomas, P.J.; Meng, X.-J.; Halbur, P.G. Routes of transmission of swine hepatitis E virus in pigs. J. Clin. Microbiol. 2004, 42, 5047–5052. [Google Scholar] [CrossRef]
- Said, B.; Ijaz, S.; Kafatos, G.; Booth, L.; Thomas, H.L.; Walsh, A.; Ramsay, M.; Morgan, D. Hepatitis E outbreak on cruise ship. Emerg. Infect. Dis. 2009, 15, 1738–1744. [Google Scholar] [CrossRef] [PubMed]
- Tamada, Y.; Yano, K.; Yatsuhashi, H.; Inoue, O.; Mawatari, F.; Ishibashi, H. Consumption of wild boar linked to cases of hepatitis E. J. Hepatol. 2004, 40, 869–870. [Google Scholar] [CrossRef]
- Tei, S.; Kitajima, N.; Takahashi, K.; Mishiro, S. Zoonotic transmission of hepatitis E virus from deer to human beings. Lancet 2003, 362, 371–373. [Google Scholar] [CrossRef]
- Yazaki, Y.; Mizuo, H.; Takahashi, M.; Nishizawa, T.; Sasaki, N.; Gotanda, Y.; Okamoto, H. Sporadic acute or fulminant hepatitis E in Hokkaido, Japan, may be food-borne, as suggested by the presence of hepatitis E virus in pig liver as food. J. Gen. Virol. 2003, 84, 2351–2357. [Google Scholar] [CrossRef]
- Krumbholz, A.; Mohn, U.; Lange, J.; Motz, M.; Wenzel, J.J.; Jilg, W.; Walther, M.; Straube, E.; Wutzler, P.; Zell, R. Prevalence of hepatitis E virus-specific antibodies in humans with occupational exposure to pigs. Med. Microbiol. Immunol. 2012, 201, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Raj, V.S.; Smits, S.L.; Pas, S.D.; Provacia, L.B.; Moorman-Roest, H.; Osterhaus, A.; Haagmans, B.L. Novel Hepatitis E Virus in Ferrets, the Netherlands. Emerg. Infect. Dis. 2012, 18, 1369–1370. [Google Scholar] [CrossRef]
- Krog, J.S.; Breum, S.Ø.; Jensen, T.H.; Larsen, L.E. Hepatitis E Virus Variant in Farmed Mink, Denmark. Emerg. Infect. Dis. 2013, 19, 2028–2030. [Google Scholar] [CrossRef]
- Johnson, N.; Freuling, C.; Vos, A.; Un, H.; Valtchovski, R.; Turcitu, M.; Dumistrescu, F.; Vuta, V.; Velic, R.; Sandrac, V.; et al. Epidemiology of rabies in Southeast Europe. Dev. Biol. 2008, 131, 189–198. [Google Scholar]
- Bodewes, R.; Ruiz-Gonzalez, A.; Schapendonk, C.M.; Brand, J.M.A.V.D.; Osterhaus, A.; Smits, S.L. Viral metagenomic analysis of feces of wild small carnivores. Virol. J. 2014, 11, 89. [Google Scholar] [CrossRef]
- Szewczyk, T.; Werszko, J.; Myczka, A.W.; Laskowski, Z.; Karbowiak, G. Molecular detection of Anaplasma phagocytophilum in wild carnivores in north-eastern Poland. Parasites Vectors 2019, 12, 465. [Google Scholar] [CrossRef]
- Takumi, K.; Sprong, H.; Hofmeester, T.R. Impact of vertebrate communities on Ixodes ricinus-borne disease risk in forest areas. Parasites Vectors 2019, 12, 434. [Google Scholar] [CrossRef]
- Bodewes, R.; Van Der Giessen, J.; Haagmans, B.L.; Osterhaus, A.; Smits, S.L. Identification of Multiple Novel Viruses, Including a Parvovirus and a Hepevirus, in Feces of Red Foxes. J. Virol. 2013, 87, 7758–7764. [Google Scholar] [CrossRef]
- Eiden, M.; Dähnert, L.; Spoerel, S.; Vina-Rodriguez, A.; Schröder, R.; Conraths, F.J.; Groschup, M.H. Spatial-Temporal Dynamics of Hepatitis E Virus Infection in Foxes (Vulpes vulpes) in Federal State of Brandenburg, Germany, 1993–2012. Front. Microbiol. 2020, 11, 115. [Google Scholar] [CrossRef]
- Lanszki, Z.; Kurucz, K.; Zeghbib, S.; Kemenesi, G.; Lanszki, J.; Jakab, F. Identification of Hepatitis E Virus in the Feces of Red Foxes (Vulpes vulpes). Animals 2020, 10, 1841. [Google Scholar] [CrossRef]
- Lanszki, J.; Heltai, M.; Kövér, G.; Zalewski, A. Non-linear relationship between body size of terrestrial carnivores and their trophic niche breadth and overlap. Basic Appl. Ecol. 2019, 38, 36–46. [Google Scholar] [CrossRef]
- Soe, E.; Davison, J.; Süld, K.; Valdmann, H.; Laurimaa, L.; Saarma, U. Europe-wide biogeographical patterns in the diet of an ecologically and epidemiologically important mesopredator, the red fox Vulpes vulpes: A quantitative review. Mammal Rev. 2017, 47, 198–211. [Google Scholar] [CrossRef]
- Johne, R.; Dremsek, P.; Reetz, J.; Heckel, G.; Hess, M.; Ulrich, R.G. Hepeviridae: An expanding family of vertebrate viruses. Infect. Genet. Evol. 2014, 27, 212–229. [Google Scholar] [CrossRef]
- Xia, J.; Zeng, H.; Liu, L.; Zhang, Y.; Liu, P.; Geng, J.; Wang, L.; Wang, L.; Zhuang, H. Swine and rabbits are the main reservoirs of hepatitis E virus in China: Detection of HEV RNA in feces of farmed and wild animals. Arch. Virol. 2015, 160, 2791–2798. [Google Scholar] [CrossRef]
- Jemeršić, L.; Roić, B.; Balatinec, J.; Keros, T. Hepatitis E—Are we threatened? Hepatitis E—Jesmo li ugroženi? Vet. Stanica 2010, 41, 383–397. [Google Scholar]
- Prpić, J.; Černi, S.; Škorić, D.; Keros, T.; Brnić, D.; Cvetnić, Ž.; Jemeršić, L. Distribution and molecular characterization of Hepatitis E virus in domestic animals and wildlife in Croatia. Food Environ. Virol. 2015, 7, 195–205. [Google Scholar] [CrossRef]
- Jemeršić, L.; Keros, T.; Maltar, L.; Barbić, L.; Vilibić-Cavlek, T.; Jeličić, P.; Ðaković-Rode, O.; Prpić, J. Differences in hepatitis E virus (HEV) presence in naturally infected seropositive domestic pigs and wild boars—An indication of wild boars having an important role in HEV epidemiology. Vet. Arh. 2017, 87, 651–663. [Google Scholar] [CrossRef]
- Jemeršić, L.; Prpić, J.; Brnić, D.; Keros, T.; Pandak, N.; Đaković-Rode, O. Genetic diversity of hepatitis E virus (HEV) strains derived from humans, swine and wild boars in Croatia from 2010 to 2017. BMC Infect. Dis. 2019, 19, 269. [Google Scholar] [CrossRef]
- Burri, C.; VIal, F.; Ryser-Degiorgis, M.P.; Schwermer, H.; Darling, K.; Reist, M.; Wu, N.; Beerli, O.; Schoning, J.; Cavassini, M.; et al. Seroprevalence of hepatitis E virus in domestic pigs and wild boars in Switzerland. Zoonoses Public Health 2014, 61, 537–544. [Google Scholar] [CrossRef]
- Anita, A.; Gorgan, L.; Anita, D.; Oslobanu, L.; Pavio, N.; Savuta, G. Evidence of hepatitis E infection in swine and humans in East Region of Romania. Int. J. Infect. Dis. 2014, 29, 232–237. [Google Scholar] [CrossRef]
- Montagnaro, S.; De Martinis, C.; Sasso, S.; Ciarci, R.; Damiano, S.; Auletta, L.; Iovane, V.; Zottola, T.; Pagnini, U. Viral and antibody prevalence of hepatitis E in European wild boars (Sus scrofa) and hunters at Zoonotic risk in the Latium Region. J. Comp. Pathol. 2015, 153, 1–8. [Google Scholar] [CrossRef]
- Thiry, D.; Mauroy, A.; Saegerman, C.; Licoppe, A.; Fett, T.; Thomas, I.; Brochier, B.; Thiry, E.; Linden, A. Belgian wildlife as potential zoonotic reservoir of hepatitis E. Trans. Emerg. Dis. 2015, 64, 764–773. [Google Scholar] [CrossRef]
- Mazzei, M.; Forzan, M.; Pizzurro, F.; Picciolli, F.; Bandecchi, P.; Poli, A. Detection of hepatitis E virus antibodies in domestic and wild animal species in central Italy. Clin. Microbiol. 2015, 4, 227. [Google Scholar] [CrossRef]
- Prpić, J.; Keros, T.; Vucelja, M.; Bjedov, L.; Ðaković-Rode, O.; Margaletić, J.; Habrun, B.; Jemeršić, L. First evidence of hepatitis E virus infection in a small mammal (yellow-necked mouse) from Croatia. PLoS ONE 2019, 14, e0225583. [Google Scholar] [CrossRef]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef]
- Kruse, H. Wildlife as source of zoonotic infections. Emerg. Infect. Dis. J. 2004, 10, 102–112. [Google Scholar] [CrossRef]
- World Health Organization. WHO Consultation on Public Health and Animal Transmissible Spongiform Encephalopathies: Epidemiology, Risk and Research Requirements, Geneva, Switzerland, 1–3 December 1999. Available online: https://apps.who.int/iris/handle/10665/66422 (accessed on 25 February 2023).
- Weiss, S.; Nowak, K.; Fahr, J.; Wibbelt, G.; Mombouli, J.V.; Parra, H.J.; Leendertz, F. Henipavirus-related sequences in fruit bat bushmeat, Republic of Congo. Emerg. Infect. Dis. 2012, 18, 1527–1536. [Google Scholar] [CrossRef]
- Pernet, O.; Schneider, B.S.; Beaty, S.M.; LeBreton, M.; Yun, T.E.; Park, A.; Zachariah, T.T.; Bowden, T.A.; Hitchens, P.; Ramirez, C.M.; et al. Evidence for henipavirus spillover into human populations in Africa. Nat. Commun. 2014, 5, 5342. [Google Scholar] [CrossRef]
- Taylor, L.H.; Latham, S.M.; Mark, E. Risk factors for human disease emergence. Philos. Trans. R. Soc. Lond. Ser. B. Biol. Sci. 2001, 356, 983–989. [Google Scholar] [CrossRef]
- Williams, E.; Yuill, T.; Artois, M.; Fischer, J.; Haigh, S. Emerging infectious diseases in wildlife. Rev. Sci. Tech. Off. Int. Epizoot. 2002, 21, 139–158. [Google Scholar] [CrossRef]
- Johnson, C.K.; Hitchens, P.L.; Pandit, P.S.; Rushmore, J.; Evans, T.; Smiley, Y.; Cristine, C.W.; Doyle, M.M. Global shifts in mammalian population trends reveal key predictors of virus spillover risk. Proc. R. Soc. B. 2020, 287, 20192736. [Google Scholar] [CrossRef]
- Wolfe, N.D.; Daszak, P.; Kilpatrick, A.M.; Burke, D.S. Bushmeat, hunting, deforestation, and prediction of zoonotic disease emergence. Emerg. Infect. Dis. 2005, 11, 1822–1827. [Google Scholar] [CrossRef]
- Johne, R.; Plenge-Bönig, A.; Hess, M.; Ulrich, R.G.; Reetz, J.; Schielke, A. Detection of a novel hepatitis E-like virus in faeces of wild rats using a nested broad-spectrum RT-PCR. J. Gen. Virol. 2009, 91, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Bedeković, T.; Lemo, N.; Lojkić, I.; Mihaljević, Z.; Jungić, A.; Cvetnić, Z.; Cač, Z.; Hostnik, P. Modification of the fluorescent antibody virus neutralisation test—Elimination of the cytotoxic effect for the detection of rabies virus neutralising antibodies. J. Virol. Methods 2013, 189, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Wen, G.P.; Tang, Z.M.; Yang, F.; Zhang, K.; Ji, W.F.; Cai, W.; Huang, S.J.; Wu, T.; Zhang, J.; Zheng, Z.Z.; et al. A valuable antigen detection method for diagnosis of acute hepatitis E. J. Clin. Microbiol. 2015, 53, 782–788. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Geng, Y.; Harrison, T.J.; Huang, W.; Song, A.; Wang, Y. Evaluation of an antigen-capture EIA for the diagnosis of hepatitis E virus infection. J. Viral Hepat. 2015, 22, 957–963. [Google Scholar] [CrossRef]
- Lu, J.; Huang, Y.; Wang, P.; Li, Q.; Li, Z.; Jiang, J.; Guo, Q.; Gui, H.; Xie, Q. Dynamics of Hepatitis E Virus (HEV) Antibodies and Development of a Multifactorial Model To Improve the Diagnosis of HEV Infection in Resource-Limited Settings. J. Clin. Microbiol. 2021, 21, 59. [Google Scholar] [CrossRef]
- Haydon, D.T.; Cleaveland, S.; Taylor, L.H.; Laurenson, M.K. Identifying reservoirs of infection: A conceptual and practical challenge. Emerg. Infect. Dis. 2002, 8, 1468–1473. [Google Scholar]
- Hassell, J.M.; Begon, M.; Ward, M.J.; Fèvre, E.M. Urbanization and disease emergence: Dynamics at the wildlife-livestock-human Interface. Trends Ecol. Evol. 2017, 32, 55–67. [Google Scholar] [CrossRef]
- Mackenstedt, U.; Jenkins, D.; Romig, T. The role of wildlife in the transmission of parasitic zoonoses in peri-urban and urban areas. Int. J. Parasitol. Parasites Wildl. 2015, 4, 71–79. [Google Scholar] [CrossRef]
- Najm, N.A.; Meyer-Kayser, E.; Hoffmann, L.; Herb, I.; Fensterer, V.; Pfister, K.; Silaghi, C. A molecular survey of Babesia spp. and Theileria spp. in red foxes (Vulpes vulpes) and their ticks from Thuringia, Germany. Ticks Tick Borne Dis. 2014, 5, 386–391. [Google Scholar] [CrossRef]
- Vuitton, D.A.; Demonmerot, F.; Knapp, J.; Richou, C.; Grenouillet, F.; Chauchet, A.; Vuitton, L.; Bresson-Hadni, S.; Millon, L. Clinical epidemiology of human AE in Europe. Vet. Parasitol. 2015, 213, 110–120. [Google Scholar] [CrossRef]
- Sridhar, S.; Yip, C.C.; Wu, S.; Cai, J.; Zhang, A.J.-X.; Leung, K.-H.; Chung, T.W.; Chan, J.F.; Chan, W.-M.; Teng, J.L.; et al. Rat Hepatitis E Virus as Cause of Persistent Hepatitis after Liver Transplant. Emerg. Infect. Dis. 2018, 24, 2241–2250. [Google Scholar] [CrossRef] [PubMed]
- Dremsek, P.; Wenzel, J.J.; Johne, R.; Ziller, M.; Hofmann, J.; Groschup, M.H.; Werdermann, S.; Mohn, U.; Dorn, S.; Motz, M.; et al. Seroprevalence study in forestry workers from eastern Germany using novel genotype 3- and rat hepatitis E virus-specific immunoglobulin G ELISAs. Med. Microbiol. Immunol. 2011, 201, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Hamaguchi, S.; Ngo, C.C.; Li, T.-C.; Ando, S.; Yoshimatsu, K.; Yasuda, S.P.; Koma, T.; Isozumi, R.; Tsuda, Y.; et al. Serological evidence of infection with rodent-borne hepatitis E virus HEV-C1 or antigenically related virus in humans. J. Vet. Med. Sci. 2016, 78, 1677–1681. [Google Scholar] [CrossRef] [PubMed]
- Andonov, A.; Robbins, M.; Borlang, J.; Cao, J.; Hatchette, T.; Stueck, A.; Deschambault, Y.; Murnaghan, K.; Varga, J.; Johnston, L. Rat Hepatitis E Virus Linked to Severe Acute Hepatitis in an Immunocompetent Patient. J. Infect. Dis. 2019, 220, 951–955. [Google Scholar] [CrossRef]
- Ryll, R.; Eiden, M.; Heuser, E.; Weinhardt, M.; Ziege, M.; Höper, H.; Groschup, M.H.; Heckel, G.; Johne, R.; Ulrich, R.G. Hepatitis E virus in feral rabbits along a rural-urban transect in Central Germany. Infect. Genet. Evol. 2018, 61, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Chaussade, H.; Rigaud, E.; Allix, A.; Carpentier, A.; Touzé, A.; Delzescaux, D.; Choutet, P.; Garcia-Bonnet, N.; Coursaget, P. Hepatitis E virus seroprevalence and risk factors for individuals in working contact with animals. J. Clin. Virol. 2013, 58, 504–508. [Google Scholar] [CrossRef]
- Arnold, J.; Humer, A.; Heltai, M.; Murariu, D.; Spassov, N.; Hackländer, K. Current status and distribution of golden jackals Canis aureus in Europe. Mammal Rev. 2012, 42, 1–11. [Google Scholar] [CrossRef]
- Duscher, G.G.; Kübber-Heiss, A.; Richter, B.; Suchentrunk, F. A golden jackal (Canis aureus) from Austria bearing Hepatozoon canis—Import due to immigration into a non-endemic area? Ticks Tick Borne Dis. 2013, 4, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Majláthová, V.; Hurníková, Z.; Majláth, I.; Petko, B. Hepatozoon canis infection in Slovakia: Imported or autochthonous? Vector Borne Zoonotic Dis. 2007, 7, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Shamir, M.; Yakobson, B.; Baneth, G.; King, R.; Dar-Verker, S.; Markovics, A.; Aroch, I. Antibodies to selected canine pathogens and infestation with intestinal helminths in golden jackals (Canis aureus) in Israel. Vet. J. 2001, 162, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Cirović, D.; Chochlakis, D.; Tomanović, S.; Sukara, R.; Penezić, A.; Tselentis, Y.; Psaroulaki, A. Presence of Leishmania and Brucella species in the golden jackal Canis aureus in Serbia. BioMed Res. Int. 2014, 2014, 728516. [Google Scholar] [CrossRef]
- Zhang, Y.; Gong, W.; Song, W.T.; Fu, H.; Wang, L.; Li, M.; Wang, L.; Zhuang, H. Different susceptibility and pathogenesis of rabbit genotype 3 hepatitis E virus (HEV-3) and human HEV-3 (JRC-HE3) in SPF rabbits. Vet. Microbiol. 2017, 207, 1–6. [Google Scholar] [CrossRef]
- Li, S.; He, Q.; Yan, L.; Li, M.; Liang, Z.; Shu, J.; Zhang, F.; Wang, L.; Wang, L. Infectivity and pathogenicity of different hepatitis E virus genotypes/subtypes in rabbit model. Emerg. Microbes Infect. 2020, 9, 2697–2705. [Google Scholar] [CrossRef] [PubMed]
- Davidson, W.R.; Appel, M.J.; Doster, G.L.; Baker, O.E.; Brown, J.F. Diseases and parasites of red foxes, gray foxes, and coyotes from commercial sources selling to fox-chasing enclosures. J. Wildl. Dis. 1992, 28, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Goller, K.V.; Fickel, J.; Hofer, H.; Beier, S.; East, M.L. Coronavirus genotype diversity and prevalence of infection in wild carnivores in the Serengeti National Park, Tanzania. Arch. Virol. 2013, 158, 729–734. [Google Scholar] [CrossRef] [PubMed]
- Lanszki, Z.; Horváth, G.F.; Bende, Z.; Lanszki, J. Differences in the diet and trophic niche of three sympatric carnivores in a marshland. Mammal Res. 2019, 65, 93–104. [Google Scholar] [CrossRef]
- Jemeršić, L.; Lojkić, I.; Krešić, N.; Keros, T.; Zelenika, T.A.; Jurinović, L.; Skok, D.; Bata, I.; Boras, J.; Habrun, B.; et al. Investigating the Presence of SARS CoV-2 in Free-Living and Captive Animals. Pathogens 2021, 10, 635. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
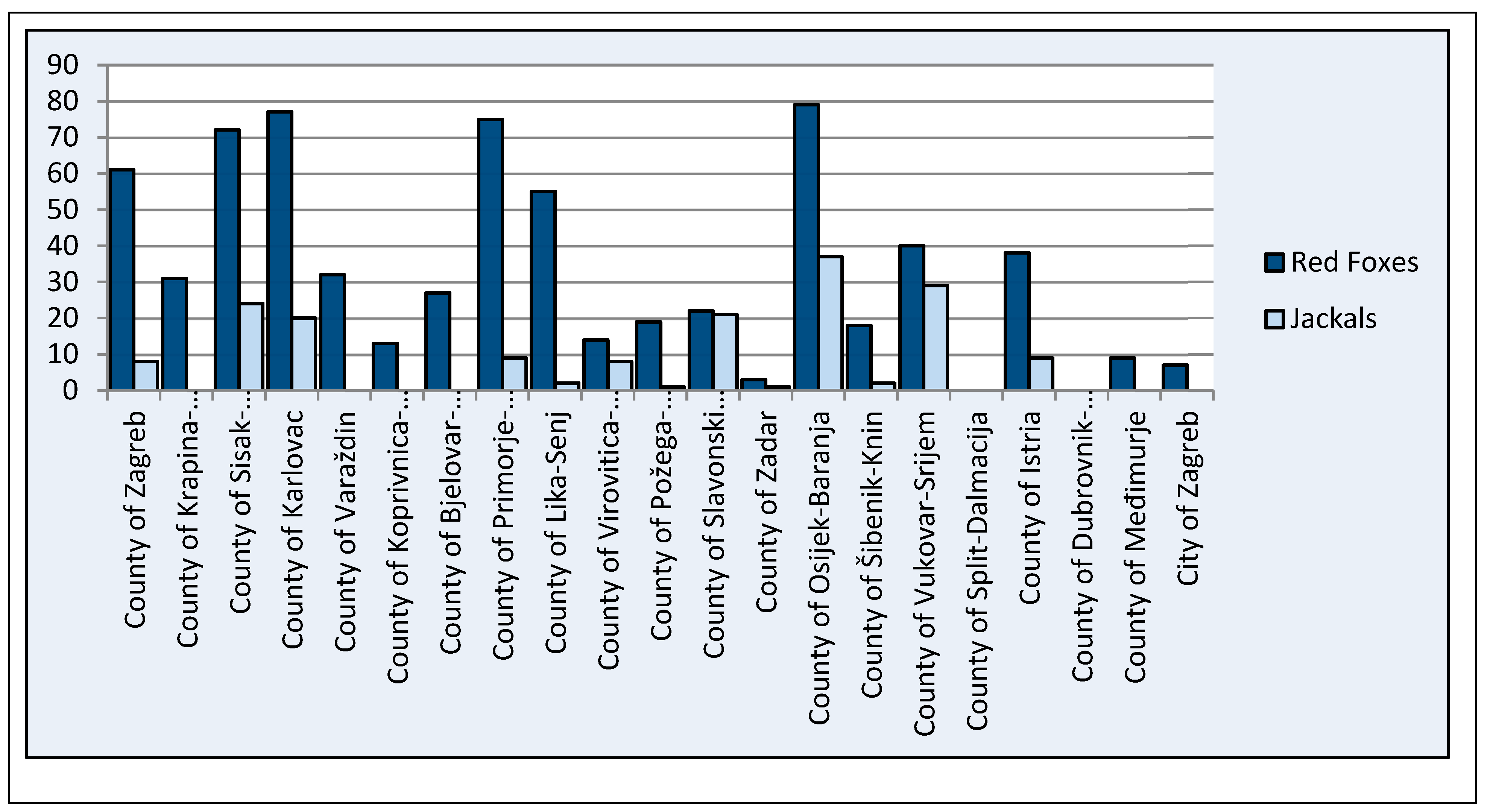
| County | Number of Feces Samples of Red Foxes | Number of Muscle Samples of Red Foxes | Number of Feces Samples of Jackals | Number of Muscle Samples of Jackals |
|---|---|---|---|---|
| County of Zagreb | 28 | 33 | 5 | 3 |
| County of Krapina-Zagorje | 16 | 15 | 0 | 0 |
| County of Sisak-Moslavina | 28 | 44 | 12 | 12 |
| County of Karlovac | 37 | 40 | 18 | 2 |
| County of Varaždin | 25 | 7 | 0 | 0 |
| County of Koprivnica-Križevci | 0 | 13 | 0 | 0 |
| County of Bjelovar-Bilogora | 2 | 25 | 0 | 0 |
| County of Primorje-Gorski Kotar | 27 | 48 | 9 | 0 |
| County of Lika-Senj | 6 | 49 | 0 | 2 |
| County of Virovitica-Podravina | 5 | 9 | 8 | 0 |
| County of Požega-Slavonija | 8 | 11 | 1 | 0 |
| County of Slavonski Brod-Posavina | 7 | 15 | 15 | 6 |
| County of Zadar | 0 | 3 | 0 | 1 |
| County of Osijek-Baranja | 51 | 28 | 27 | 10 |
| County of Šibenik-Knin | 0 | 18 | 0 | 2 |
| County of Vukovar-Srijem | 31 | 9 | 26 | 3 |
| County of Split-Dalmacija | 0 | 0 | 0 | 0 |
| County of Istria | 24 | 14 | 7 | 2 |
| County of Dubrovnik-Neretva | 0 | 0 | 0 | 0 |
| County of Međimurje | 7 | 2 | 0 | 0 |
| City of Zagreb | 7 | 0 | 0 | 0 |
| 309 | 383 | 128 | 43 | |
| Σ | 692 | 171 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prpić, J.; Kunić, A.; Keros, T.; Lojkić, I.; Brnić, D.; Jemeršić, L. Absence of Hepatitis E Virus (HEV) Circulation in the Most Widespread Wild Croatian Canine Species, the Red Fox (Vulpes vulpes) and Jackal (Canis aureus moreoticus). Microorganisms 2023, 11, 834. https://doi.org/10.3390/microorganisms11040834
Prpić J, Kunić A, Keros T, Lojkić I, Brnić D, Jemeršić L. Absence of Hepatitis E Virus (HEV) Circulation in the Most Widespread Wild Croatian Canine Species, the Red Fox (Vulpes vulpes) and Jackal (Canis aureus moreoticus). Microorganisms. 2023; 11(4):834. https://doi.org/10.3390/microorganisms11040834
Chicago/Turabian StylePrpić, Jelena, Ana Kunić, Tomislav Keros, Ivana Lojkić, Dragan Brnić, and Lorena Jemeršić. 2023. "Absence of Hepatitis E Virus (HEV) Circulation in the Most Widespread Wild Croatian Canine Species, the Red Fox (Vulpes vulpes) and Jackal (Canis aureus moreoticus)" Microorganisms 11, no. 4: 834. https://doi.org/10.3390/microorganisms11040834
APA StylePrpić, J., Kunić, A., Keros, T., Lojkić, I., Brnić, D., & Jemeršić, L. (2023). Absence of Hepatitis E Virus (HEV) Circulation in the Most Widespread Wild Croatian Canine Species, the Red Fox (Vulpes vulpes) and Jackal (Canis aureus moreoticus). Microorganisms, 11(4), 834. https://doi.org/10.3390/microorganisms11040834