Abstract
Pectinases are a series of enzymes that degrade pectin and have been used extensively in the food, feed, and textile industries. The ruminant animal microbiome is an excellent source for mining novel pectinases. Two polygalacturonase genes, IDSPga28-4 and IDSPga28-16, from rumen fluid cDNA, were cloned and heterologously expressed. Recombinant IDSPGA28-4 and IDSPGA28-16 were stable from pH 4.0 to 6.0, with activities of 31.2 ± 1.5 and 330.4 ± 12.4 U/mg, respectively, against polygalacturonic acid. Hydrolysis product analysis and molecular dynamics simulation revealed that IDSPGA28-4 was a typical processive exo-polygalacturonase and cleaved galacturonic acid monomers from polygalacturonic acid. IDSPGA28-16 cleaved galacturonic acid only from substrates with a degree of polymerization greater than two, suggesting a unique mode of action. IDSPGA28-4 increased the light transmittance of grape juice from 1.6 to 36.3%, and IDSPGA28-16 increased the light transmittance of apple juice from 1.9 to 60.6%, indicating potential application in the beverage industry, particularly for fruit juice clarification.
1. Introduction
The plant structural polymers cellulose, hemicellulose, pectin, and lignin are the most abundant renewable organic carbon source in nature. Pectin, the most complex of these polysaccharides, gives plants structural rigidity and resistance to attack by insects and pathogens [1,2]. Pectin mainly comprises homogalacturonan regions (also known as “smooth” regions) consisting of D-galacturonic acid units, linked by α-1,4 glycosidic bonds, and substituted homogalacturonan regions (also known as “hairy” regions) such as xylogalacturonan, rhamnogalacturonan I, and rhamnogalacturonan II, which account for 15–30% of the pectin in fruits and vegetables [3,4]. Complete hydrolytic degradation of pectin requires a series of functional pectic enzymes, because of its complexity and heterogeneity. These enzymes include polygalacturonase (three types), pectate lyases (EC 4.2.2.2 and EC 4.2.2.9), pectin methylesterase (EC 3.2.1.11), and rhamnogalacturonase (EC 3.2.1.171) [5,6]. Breakdown of homogalacturonan, the simplest structural region in pectin, is initiated by endo-polygalacturonase (EC 3.2.1.15), which attacks the α-1,4 glycosidic bonds between D-polygalacturonic acid residues and generates pectic oligogalacturonides. Exo-polygalacturonase (EC 3.2.1.67) or exo-poly-α-digalacturonosidase (EC 3.2.1.82) then cleave the first glycosidic bond from the non-reducing end of the oligogalacturonides to release monomers or dimers.
Pectinases have been used extensively in the food, feed, and textile industries, because of their effectiveness in degrading plant material, such as fruit pulp. According to the carbohydrate-active enzyme (CAZyme) database (http://www.cazy.org/, accessed on 10 March 2022) [7], these pectinolytic enzymes are mainly classified into the glycoside hydrolase (GH) family 28. Many GHs have been identified from bacteria, fungi, and higher plants and more than 12,000 GH28 gene sequences have been deposited in GenBank, but only about 100 of them have been functionally characterized. For instance, NfPG I, derived from Neosartorya fischeri P1, was reported as the most catalytically active endo-polygalacturonase with a specific activity of 40,123 U/mg [8]. To satisfy the demands of industrial bioprocessing, a highly active (25,900 U/mg) endo-polygalacturonase, TePG28b, with good thermostability (optimum temperature 70 °C), was isolated from a thermophilic fungus, Talaromyces leycettanus [4]. The hydrolytic behavior, catalytic mechanism, and molecular basis of endo-acting polygalacturonases have also been well-elucidated [9,10,11]. In contrast, to our knowledge, characterized exo-polygalacturonases and exo-poly-α-digalacturonosidases are much less catalytically efficient, with specific activities <300 U/mg [5,12,13,14].
Ruminants, such as sheep and cattle, digest nutrient-poor plant material, such as grass, very efficiently, because of the numerous cellulolytic bacteria and fungi in the rumen. A variety of CAZymes including glucanase [15,16], cellulase [17], and xylanase [18] have been mined using multi-omics sequencing and heterologous expression strategies. Apart from degradation by CAZymes, the metabolism of complex polysaccharides such as pectin is also orchestrated by gene clusters, termed polysaccharide utilization loci (PULs), found in gut microbes, particularly in the Bacteroides genus [19,20]. Thus, gastrointestinal microbes in ruminants are of tremendous interest for mining novel CAZymes for industrial application. In our previous study, ~15,000 unigenes including 4225 open reading frames (ORFs) were obtained from the sheep rumen microbiome using a metatranscriptomics strategy [17]. Of these, 173 ORFs were annotated to the GH28 family. In order to explore highly efficient pectinases, we randomly picked ten of the 173 ORFs for gene cloning and succeeded in obtaining two full-length GH28 family proteins. In this study, the two GH28 polygalacturonase genes, IDSPga28-4 (GenBank: OK149199) and IDSPga28-16 (GenBank: OK149200), were cloned from rumen fluid cDNA and heterologously expressed in Escherichia coli. The biochemical properties, hydrolysis products, and molecular basis for catalysis of the recombinant enzymes were investigated.
2. Materials and Methods
2.1. Materials
Sheep rumen cDNA was prepared as described previously [17], stored at −80 °C, and used within one month. The plasmid pET-30a (+) was from Invitrogen (Shanghai, China), and expression host E. coli BL21 (DE3) was from Tiangen (Beijing, China). Polygalacturonic acid (PGA) from citrus pectin and rhamnogalacturonan I from potato were from Megazyme (Wicklow, Ireland).
D-(+)-Galacturonic acid (GalA) monohydrate, digalacturonic acid ((GalA)2), trigalacturonic acid ((GalA)3), and pectin from citrus peel (~60% degree of esterification) were from Sigma-Aldrich (St. Louis, MO, USA). Isopropyl-thio-β-D-galactopyranoside (IPTG), kanamycin, and Luria–Bertani (LB) medium were from Sangon (Shanghai, China).
2.2. Gene Cloning, Expression, and Protein Purification
Two putative GH28 family genes, IDSPga28-4 and IDSPga28-16, were PCR-amplified (Table S1) from the cDNA of Hu sheep rumen fluid using Super pfx DNA polymerase (CWBIO, Beijing, China). The expected fragments were purified and cloned into the truncated pET-30a(+) vector by BamHI + XhoI via homologous recombination using the Trelief™ SoSoo Cloning kit (TsingKe Biotech, Beijing, China). The resulting ligation products were transformed into E. coli BL21 (DE3)-competent cells by heat shock transformation and streaked onto an LB agar plate (5 g/L yeast extract, 10 g/L tryptone, 10 g/L sodium chloride, 20 g/L agar) supplemented with 50 μg/mL of kanamycin, and then incubated at 37 °C for 16 h. Ten colonies of each gene construct were selected for colony-PCR validation. Positive recombinant plasmids were further confirmed by Sanger sequencing (Sangon, Shanghai, China). The recombinant hosts were designated as BL21/pET30a/IDSPga28-4 and BL21/pET30a/IDSPga28-16, respectively. Subsequently, IPTG-induced gene expression, cell pellet collection, and sonication were carried out as described previously [16], and then crude enzyme, resuspended in ice-cold phosphate-buffered saline (PBS, pH 7.4), was loaded onto a HisTrap™ 5 mL column (GE Healthcare BioSciences, Pittsburgh, PA, USA) fitted to an ÄKTA start protein purification system. The bound proteins were eluted with elution buffer, supplemented with a linear gradient of 20–250 mM imidazole. Eluted fractions were analyzed by 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and the purified protein fractions were used for enzymatic assays.
2.3. Sequence Analysis
Amino acid alignment was performed using ClustalO (https://www.ebi.ac.uk/Tools/msa/clustalo/, accessed on 10 March 2022) and BLASTp (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 10 March 2022). Signal peptides were predicted using SignalP 5.0 (https://services.healthtech.dtu.dk/, accessed on 10 March 2022). The isoelectric point and molecular weight were predicted by ProtParam (https://web.expasy.org/protparam/, accessed on 10 March 2022). Multiple sequence alignment was conducted by MUSCLE with MEGA software (v.7.0, Kumar Lab, Temple University, Philadelphia, PA, USA). Phylogenetic tree analysis was performed using the Neighbor joining (NJ) statistical method based on the Poisson correction model with 1000 bootstrap replications, and then the tree was imported into the online software iTOL v6.4 (https://itol.embl.de/, accessed on 12 September 2022) for further optimization. The three-dimensional (3-D) structures were modeled by SWISS-MODEL (https://www.swissmodel.expasy.org/, accessed on 21 October 2022) and visualized using PyMOL v2.4 (https://pymol.org/2/, accessed on 21 October 2022).
2.4. Enzyme Activity Assay
Polygalacturonase activity was determined using the 3,5-dinitrosalicylic acid (DNS) method [21] with GalA as the standard. Briefly, purified enzyme solution (15 μL, 0.43 μg protein) and PGA (60 μL, 0.25% w/v) were dissolved in 100 mM citric acid-Na2HPO4 buffer (pH 5.0), unless otherwise stated. After incubation at 40 °C for 10 min, DNS solution (75 μL) was added to terminate the reaction, followed by 10 min in a boiling water bath, and then the absorbance of the reaction mixture was measured at 540 nm, using a Spark multimode microplate reader (Tecan, Männedorf, Switzerland). One enzyme unit (U) was defined as 1 μmol of reducing sugar released per minute. Enzyme concentration was measured by the Bradford method [22] using bovine serum albumin as the standard. To investigate the substrate specificity, purified enzyme was reacted with 0.25% (w/v) PGA, rhamnogalacturonan I, or pectin.
2.5. Characterization of Purified Recombinant Enzymes
To determine the optimum pH, purified IDSPGA28-4, or IDSPGA28-16, was incubated with 0.25% (w/v) PGA over a pH range of 3.0–10.0 (pH 3.0–8.0, citric acid-Na2HPO4 buffer; pH 8.0–9.0, Tris-HCl buffer; pH 9.0–10.0, glycine-NaOH buffer) at 40 °C for 10 min, and then the reducing sugar produced was measured as described above. The optimum temperature of each enzyme was determined in buffer at the corresponding optimum pH and temperatures between 30 and 70 °C. The maximum enzyme activity was designated as 100%. All assays were performed in quadruplicate.
The pH stability was determined by measuring the residual activities of IDSPGA28-4 or IDSPGA28-16 under the optimum conditions (40 °C and pH 5.0) after preincubating the enzymes at 25 °C for 1 h in various buffers of pH 3.0–10.0. For the thermostability assay, IDSPGA28-4 or IDSPGA28-16 was pretreated at 30, 40, or 50 °C for 1 h. Aliquots were collected at different time intervals (10, 20, 30, and 60 min) and used to determine the residual activity. The initial enzyme activity before preincubation was designated as 100%. All assays were performed in quadruplicate.
2.6. Hydrolysis Product Analysis
Approximately 5 μg of purified enzyme was incubated with 0.25% polygalacturonic acid at pH 5.0 and 37 °C for 3 h. Aliquots sampled at different time intervals (30 min, 1 h, and 3 h) were boiled for 10 min, centrifuged at 25 °C and 10,000× g for 10 min, and then subjected to thin-layer chromatography (TLC) and high-performance anion exchange chromatography (HPAEC). The products formed by treating 0.25% (GalA)n (n = 1, 2, 3) with the purified enzymes for 24 h were analyzed by TLC and HPAEC.
TLC was conducted using a 10 × 10 cm silica gel 60 plate (Merck, Darmstadt, Germany) with the developing solvent, 1-butanol/acetic acid/water = 2:1:1 (v/v/v). The plate was taken out when the solvent front was about 1 cm from the top and, after the solvent had evaporated completely, the plate was sprayed with visualization reagent (sulfuric acid: ethanol = 5:95, v/v). The plate was air-dried, then heated at 105 °C for 10 min, until spots appeared.
HPAEC was performed on an ICS-3000 system (Thermo Scientific Dionex, Sunnyvale, CA, USA) equipped with a CarboPac PA-200 column (250 mm × 2 mm; Thermo Scientific Dionex). The mobile phase (flow rate, 0.25 mL/min, at 30 °C) was composed of: (A) 0.1 M sodium hydroxide and (B) 0.1 M sodium hydroxide + 1 M sodium acetate. After sample injection, the following linear gradient was applied: 0 min, 0% B; 10 min, 10% B; 25 min, 30% B; 30 min, 100% B; 40 min, 0% B; 40–49 min 0% B.
2.7. Molecular Docking and Molecular Dynamics (MD) Simulation
The 3-D structures of (GalA)2 and (GalA)3 were downloaded from PubChem (https://pubchem.ncbi.nlm.nih.gov/, accessed on 1 November 2022), and the 3-D structure of IDSPGA28-16 was modeled with SWISS-MODEL (https://www.swissmodel.expasy.org/, accessed on 21 October 2022), using the known structure of Thermotoga maritima exo-polygalacturonase (PDB: 3JUR), which has the highest sequence similarity (41.08%) as the template. Molecular docking between IDSPGA28-16 and (GalA)2, or (GalA)3, was performed by AutoDock Vina (v1.1, CCSB, Scripps Research Institute, California, USA) [23]. A 3-D box was defined to restrict the conformational sampling space, in which the center point of the catalytic-site Asp residues (D251, D272, and D273) was set as the box center, and the box size was 30 Å, 28 Å, and 26 Å in the x-, y-, and z-dimensions, respectively. Exo-polygalacturonase hydrolyzes the first glycosidic bond from the non-reducing end of PGA, whereas exo-poly-α-digalacturonosidase hydrolyzes the second α-1,4-glycosidic bond from the non-reducing end of the substrate, according to the CAZy database. Therefore, the conformation of the protein–substrate complex, with the nonreducing end inside the catalytic pocket and the highest score from AutoDock Vina, was subjected to molecular dynamics (MD) simulation.
The MD simulations of the complexes were carried out by Gromacs 2019.6 (https://doi.org/10.5281/zenodo.3685922, accessed on 5 November 2022) with the AMBER ff14SB force field for proteins and GAFF for the sugar molecules, using TIP3P as the water model [24,25,26]. A dodecahedral box was generated, spaced 1 nm away from the periphery of the enzyme–substrate complex, with 69,517 water molecules as the solvent to fill the box for MD simulation of the IDSPGA28-16/(GalA)2 complex, and 47,197 water molecules for IDSPGA28-16/(GalA)3. Because IDSPGA28-16 is negatively charged, 10 of the water molecules were replaced with Na+ ions to keep the system electrically neutral. For energy minimization, 50,000 steps of conjugate gradient minimization and a steepest descent energy minimization for every 1000 steps were conducted, with hydrogen bonds constrained by LINCS [27]. The V-rescale method was used for temperature control, the Parrinello–Rahman method for pressure coupling, and the PME method for the calculation of long-range electrostatic interactions. The van der Waals interactions were cut-off at 10 Å. The 100 ns simulation was performed in the isothermal–isobaric NPT ensemble (300 K, 1 atm) with a time step of 2 fs.
Water molecules were removed and the periodicity of the trajectory modified, using the trjconv module of Gromacs, and then the root-mean-square deviation (RMSD) for every 2 ps was calculated by the rms module. The noncovalent interactions (NCIs) of the two complexes were analyzed by Multiwfn 3.8 using the independent gradient model (IGM) and plotted by VMD v1.9.3 (http://www.ks.uiuc.edu/Research/vmd/, accessed on 14 December 2022) [28,29]. The detailed schematic diagrams of the intermolecular interactions, especially hydrogen bonds within 3.5 Å, were plotted by LigPlot+ (v2.2, EMBL-EBI, Wellcome Genome Campus, Cambridgeshire, UK).
2.8. Fruit Juice Clarification with Exo-Polygalacturonase
Fresh grape juice, orange juice, and apple juice were extracted from Kyoho grapes, Gannan Navel oranges, and Shaanxi Red Fuji apples (500 g each), respectively, then filtered through eight layers of gauze to remove pulp solids. Ascorbic acid (0.5% w/v) was added to the freshly extracted fruit juice to minimize oxidation. Purified IDSPGA28-4, or IDSPGA28-16, (~500 μg) was incubated with aliquots of juice (5 mL) at 37 °C, without stirring, for 1.5 h. Purified IDSPGA28-4, or IDSPGA28-16, was boiled for 10 min before addition to the juice, as controls. The light transmittance of the treated juice was measured with a UV/vis spectrophotometer (Phoenix, Shanghai, China) at 660 nm (%T660), with the light transmittance of distilled water as 100%. All reactions were performed in triplicate.
2.9. Statistical Analysis
All data were represented as the mean ± standard deviation (SD). Statistical analysis was performed using GraphPad Prism software (v.8.0.2, San Diego, CA, USA), via Student’s t-test (* p < 0.05; ** p < 0.01; *** p < 0.001).
3. Results and Discussion
3.1. Gene Cloning and Sequence Analysis
In the past decade, multi-omics approaches, such as metagenomics and metatranscriptomics, have been adopted widely to improve the understanding of the composition and functionality of the gastrointestinal microbiome in humans and animals [30]. There is great interest in mining novel CAZymes from the ruminant microbiome, because of their promising potential for processing of plant-derived foods and beverages. In this study, two exo-polygalacturonase genes, IDSPga28-4 and IDSPga28-16, were cloned from rumen fluid cDNA. The ORFs of IDSPga28-4 and IDSPga28-16 encoded 541 and 458 amino acids, respectively. Multiple sequence alignment revealed that IDSPGA28-4 and IDSPGA28-16 shared the highest similarity with two GH28 proteins from an unclassified Oscillospiraceae bacterium (96.02% identity with GenBank: MBP3209358) and a Firmicutes bacterium (69.39% identity with GenBank: MBR1735702). However, neither of the latter enzymes have been functionally characterized.
Homology modeling suggested that IDSPGA28-4 and IDSPGA28-16, which have 41.08 and 37.60% identities, respectively, with the exo-polygalacturonase from Thermotoga maritima (PDB: 3JUR), have secondary structures mainly composed of parallel β-sheets typical of the GH28 family. In addition, each enzyme has eight conserved amino acid residues (N290, D292, D313, D314, H347, G353, R378, and K380 for IDSPGA28-4; N249, D251, D272, D273, H306, G312, R337, and K339 for IDSPGA28-16), compared with 3JUR (Figure S1) [10,31]. The overall structures revealed that a large cleft was formed by four surrounding loops in IDAPGA28-4 (T167-S202, Q225-A239, S320-N330, and N415-R437) and IDSPGA28-16 (Y97-S110, Q158-L198, S279-G289, and D373-E386) (Figure S2), which are similar to those of the exo-poly-α-digalacturonosidase, YeGH28, from Yersinia enterocolitica [5]. Moreover, three aspartates (D251, D272, D273) located at the bottom of the cleft have been proposed as the key catalytic residues. Phylogenetic tree analysis was performed on IDSPGA28-4, IDSPGA28-16, and those of the GH28 polygalacturonases that had been previously characterized according to their mode of action and origin (Figure 1). IDSPGA28-4 and IDSPGA28-16 were clustered on the same branch as two exo-polygalacturonases from Bacteroides thetaiotaomicron (GenBank: AAO79228 and AAO79260) and one from T. maritima (GenBank: AAD35522), indicating that these enzymes have a similar mode of action.
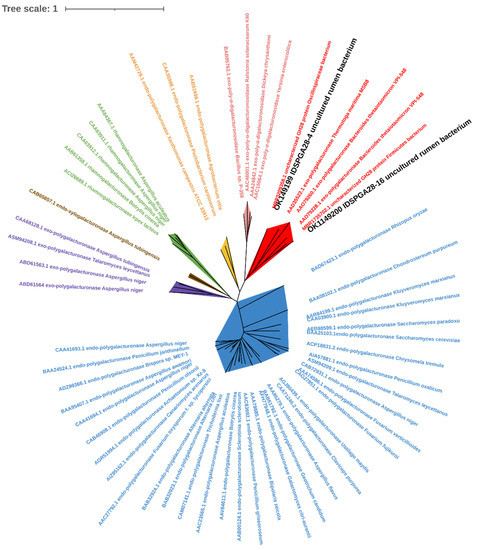
Figure 1.
Phylogenetic analysis of IDSPGA28-4 and IDSPGA28-16. Color-coding is as follows: cherry: bacterial exo-polygalacturonase (EC 3.2.1.67); plum: bacterial exo-poly-α-digalacturonosidase (EC 3.2.1.82); yellow: bacterial endo-polygalacturonase (EC 3.2.1.15); green: fungal rhamnogalacturonase (EC 3.2.1.171); brown: fungal endo-xylogalacturonase (EC 3.2.1.-); violet: fungal exo-polygalacturonase (EC 3.2.1.67); azure: fungal endo-polygalacturonase (EC 3.2.1.15).
3.2. Protein Expression and Enzymatic Properties
Recombinant IDSPGA28-4 and IDSPGA28-16 were successfully expressed in E. coli. Two electrophoresis bands of ~70 and ~60 kDa (Figure 2) were observed, consistent with the theoretical molecular masses of the two enzymes (IDSPGA28-4 ~60 kDa; IDSPGA28-16 ~51 kDa), added to that of the linked vector-peptide, respectively. Biochemical characterization revealed that both recombinant IDSPGA28-4 and IDSPGA28-16 were optimally active at 40 °C (Figure 3A,C); the enzymes were relatively stable below 40 °C but degraded rapidly above 50 °C (Figure 3B,D). Most CAZymes isolated from ruminants have temperature optima of 30–50 °C, including those from cattle [32], buffalo [33], sheep [15,16,17,18], and goat [34]. The mesophilic adaptation of IDSPGA28-4 and IDSPGA28-16 appears to result from accommodation to the ruminant gastrointestinal temperature (38–41 °C). IDSPGA28-4 and IDSPGA28-16 were optimally catalytically active at pH 5.0 and most stable from pH 4.0 to 6.0 (Figure 3E). After pretreatment for 1 h, both enzymes were relatively stable, retaining over 60% residual activity between pH 4.0 and 9.0 (Figure 3F).
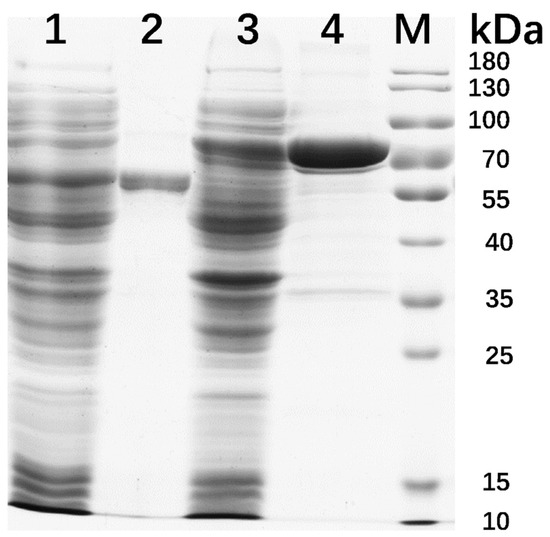
Figure 2.
SDS-PAGE analysis of recombinant IDSPGA28-4 and IDSPGA28-16. Lane 1, crude IDSPGA28-16; 2, purified IDSPGA28-16; 3, crude IDSPGA28-4; 4, purified IDSPGA28-4; M: standard protein marker.
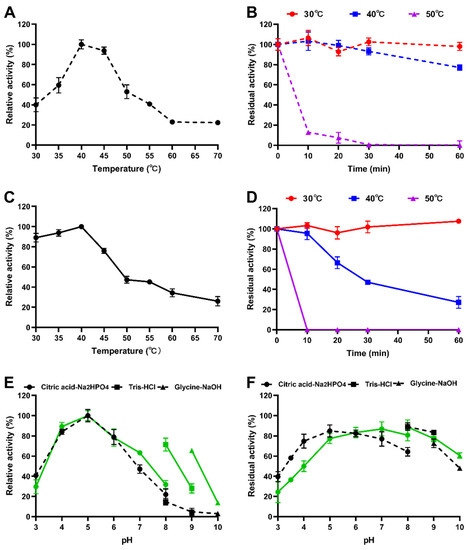
Figure 3.
Characterization of recombinant polygalacturonases. (A) Optimum temperature of IDSPGA28-4; (B) thermostability of IDSPGA28-4; (C) optimum temperature of IDSPGA28-16; (D) Thermostability of IDSPGA28-16; (E) optimum pH of IDSPGA28-4 (dotted line) and IDSPGA28-16 (solid line); (F) pH stability of IDSPGA28-4 (dotted line) and IDSPGA28-16 (solid line). Data represent the mean ± SD (n = 4).
Substrate selectivity determination indicated that both IDSPGA28-4 and IDSPGA28-16 were active against PGA, with specific activities of 31.2 ± 1.5 and 330.4 ± 12.4 U/mg (Table 1), respectively, but inactive toward pectin or rhamnogalacturonan I. Although some endo-polygalacturonases are highly active, exhibiting >20,000 U/mg against PGA [4,8], most exo-galacturonases have specific activities <300 U/mg [5,12,13,14]. Thus, IDSPGA28-16 was one of the most active exo-polygalacturonases found. The relatively high catalytic activity but poor thermostability of IDSPGA28-4 and IDSPGA28-16 suggested that protein engineering strategies such as directed evolution [35] and/or molecular cyclization [36] could improve their thermostability.

Table 1.
Specific activities of exo-polygalacturonases (EC 3.2.1.67) and exo-α-poly-digalacturonosidases (EC 3.2.1.82) against PGA.
3.3. Hydrolytic Products and Hydrolysis Pattern Analyses
To investigate the reaction modes of IDSPGA28-4 and IDSPGA28-16, the hydrolytic products of their action on PGA and (GalA)n (n = 1, 2, 3) were determined using TLC and HPAEC. IDSPGA28-4 exclusively cleaved GalA monomers from PGA (Figure 4A,B). Both IDSPGA28-4 and IDSPGA28-16 rapidly released products from PGA, and their concentrations increased to a plateau. After reaction for 3 h, IDSPGA28-4-catalyzed hydrolysis yielded 114.5 nmol of GalA (Figure 4A,B). Exo-polygalacturonase (EC 3.2.1.67) exclusively releases GalA monomers from substrates including digalacturonate [4,8,12,37]. To further analyze the mode of action of IDSPGA28-4 and IDSPGA28-16, mono- and oligogalacturonides were used as hydrolysis substrates. As expected, IDSPGA28-4 generated only GalA from both (GalA)2 and (GalA)3 (Figure 4D,E), indicating a conventional exo-acting mode for the enzyme.
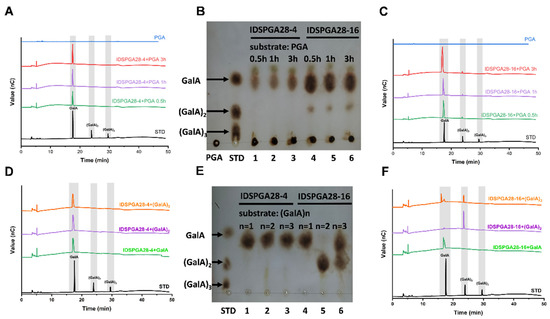
Figure 4.
Substrates hydrolysis by IDSPGA28-4 and IDSPGA28-16, analyzed by TLC and HPAEC. (A–C) Time-course of PGA hydrolysis by IDSPGA28-4, or IDSPGA28-16. Lanes 1-3 in (B): IDSPGA28-4+PGA reaction for 0.5, 1, or 3 h; Lanes 4-6 in (B): IDSPGA28-16+PGA reaction for 0.5, 1, or 3 h; (D–F) hydrolytic pattern of pectic oligosaccharides by IDSPGA28-4 or IDSPGA28-16. Lanes 1-3 in (E): reaction products of IDSPGA28-4 with GalA, (GalA)2, or (GalA)3 after 24 h; lanes 4-6 in (E): Reaction products of IDSPGA28-16 with GalA, (GalA)2, or (GalA)3 after 24 h. PGA, polygalacturonic acid; STD, standard oligosaccharides; GalA, galacturonic acid; (GalA)2, digalacturonic acid; (GalA)3, trigalacturonic acid.
Surprisingly, IDSPGA28-16 was capable of liberating both (GalA)2 and GalA from PGA, although GalA was the dominant product (Figure 4B,C). After reaction for 3 h, IDSPGA28-16 generated 338 nmol GalA and 16.3 nmol (4.6% of total reducing sugars) of (GalA)2. In addition, IDSPGA28-16 converted (GalA)3 into almost equal amounts of GalA and (GalA)2, but did not hydrolyze (GalA)2 (Figure 4E,F). The co-existence of GalA and (GalA)2 in IDSPGA28-16 hydrolysis products suggested that the enzyme could act in a combined exo/endo-mode, i.e., IDSPGA28-16 hydrolyzed PGA into oligogalacturonides with DP 3-10, and then subsequently converted them into the monomer and dimer.
Similar product profiles have been observed previously. Heterologous expression and characterization of five endo-polygalacturonases, BcPGs, from the plant pathogenic fungus, Botrytis cinerea, revealed that BcPG3 and BcPG6 mostly generated GalA and (GalA)2 from PGA, but trace amounts of (GalA)3 to (GalA)6 were also detected during the reaction [9]. The endopolygalacturonase PGD, derived from A. niger, mainly hydrolyzed PGA into GalA and (GalA)2 as final products, but (GalA)3 was also observed as an intermediate [38]. However, no intermediate oligogalacturonide products were observed during the time-course reaction of IDSPGA28-16.
Exo-poly-α-digalacturonosidase (EC 3.2.1.82) is an exo-acting hydrolase that cleaves the second α-1,4 glycosidic bond from the non-reducing end of PGA, exclusively producing (GalA)2 [5,14]. However, IDSPGA28-16 produced both GalA and (GalA)2 as hydrolytic products, suggesting that IDSPGA28-16 had a distinct mode of action from known endo-polygalacturonases (EC 3.2.1.15), exo-polygalacturonases (EC 3.2.1.67), and exo-poly-α-digalacturonosidases (EC 3.2.1.82). The product profiles (Figure 4) suggested that both IDSPGA28-4 and IDSPGA28-16 were processive exo-polygalacturonases that performed multiple glycosidic bond cleavages during each encounter with a PGA polymer chain [9,10,37,38].
3.4. Molecular Dynamics (MD) Simulation
To understand the structural basis of substrate binding and catalysis of IDSPGA28-16, molecular docking and MD simulation analyses were performed on the enzyme structural models. As molecular docking reflects the specific state at a certain moment, the most stable conformation of the complex was selected by molecular docking and then used for MD simulation (Figure 5). MD simulation of IDSPGA28-16 with (GalA)2 as a substrate reached equilibrium after 30 ns, with a root-mean-square deviation (RMSD) of 5.11 Å (Figure 5A). With (GalA)3 as the substrate, the enzyme–substrate complex reached equilibrium in ~20 ns, with a much smaller RMSD of 1.89 Å (Figure 5B). Generally, RMSD is used as a quantitative assessment of similarity between two protein structures or protein–substrate complexes; a relatively low RMSD is preferable for mechanistic simulation [39]. In this case, the conformation with the highest score obtained by molecular docking was used as the initial conformation for MD simulation. The high RMSD of the enzyme–(GalA)2 complex suggested that (GalA)2 could bind flexibly, in more than one conformation, whereas (GalA)3 was relatively tightly constrained.
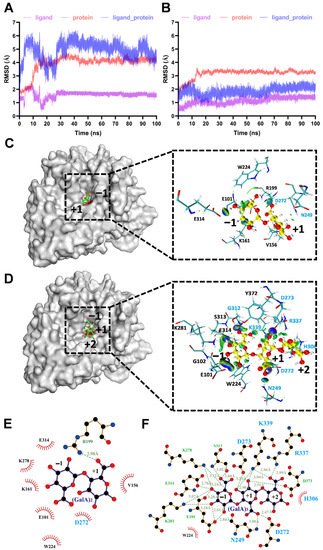
Figure 5.
MD simulation of IDSPGA28-16 enzyme–substrate complexes. (A,B) RMSD diagram for MD simulation of IDSPGA28-16 and (GalA)2 (A) or (GalA)3 (B); (C,D) analysis of MD simulation results of IDSPGA28-16 with (GalA)2 (C) or (GalA)3 (D); representative conformation diagram of the complex clustering for 70–100 ns and the visualized NCI iso-surface diagram between IDSPGA28-16 and (GalA)2 or (GalA)3. Blue indicates hydrogen bonds and green indicates van der Waals interactions; (E,F) detailed molecular interactions between IDSPGA28-16 and (GalA)2 (E) or (GalA)3 (F).
To further unravel the detailed molecular interactions between key active-site amino acid residues and substrates, the enzyme–(GalA)2 and enzyme–(GalA)3 representative structures obtained from MD simulations for 70–100 ns were used to visualize the non-covalent interaction (NCI) iso-surface diagram within 5 Å of the substrate (Figure 5C,D). The (GalA)2 or (GalA)3 was positioned at the bottom of the cleft formed by four surrounding loops in IDSPGA28-16 (Figure S2A,B). It is widely accepted that eight highly conserved residues, including three aspartates, participate in substrate binding and catalysis in endo- or exo-polygalacturonases [5,10,11,30]. Seven (N249, D272, D273, H306, G312, R337, and K339) of the eight key resides were located within 5 Å of (GalA)3 (Figure 5). More importantly, these residues had strong binding interactions with the substrate; two catalytic aspartates, D272 and D273, formed hydrogen bonds of 2.47 Å and 3.23 Å, respectively, with the +1 subsite of (GalA)3 (Figure 5F). Several basic amino acids, including K278, K287, R337, and K339, also interacted with the substrates, contributing to stabilization of the enzyme–substrate complex, in good agreement with previous reports [5,10]. Mutations of the conserved basic amino acids, i.e., arginine (R256N) and lysine (K258N), in the A. niger endopolygalacturonase II, dramatically reduced its catalytic activity, but decreased its Km 10-fold [11]. In the MD stimulation of the enzyme–(GalA)2 complex, only D272 of the eight key residues was within 5 Å of the substrate (Figure 5E), consistent with the inability of the enzyme to hydrolyze (GalA)2 (Figure 4).
Regarding the different acting modes of endo- and exo-polygalacturonases, the four conserved protein loops, particularly loop 1, appear to contribute strongly to substrate recognition and binding. Loop 1 in endo-polygalacturonases is involved in forming one “wall” of the “substrate path”, contributing to a tunnel-like active site, rather than orientating to the active center [4,5,10,11,40]. The substrate tends to lay along the tunnel-like active site in endo-polygalacturonase, which randomly cleaves α-1,4 glycosidic bonds via a single-attack manner, generating pectic oligogalacturonides and monomers [4,9,41,42]. In contrast, loop 1 in exo- polygalacturonase [19,30] and exo-poly-α-digalacturonosidase [5] formed the “back wall” of the substrate-binding cavity, forming to a pocket-like active site (Figure 5 and Figure S2). Interestingly, in addition to several basic amino acid residues, the acidic residue E101, positioned on loop 1, also formed three strong hydrogen bonds with the −1 subsite (Figure 5F). After PGA, or a pectic oligogalacturonide substrate was bound to the buried basic amino acid residues, its reducing end was blocked by loop 1. Consequently, exo-polygalacturonase could only access and cleave the first α-1,4 glycosidic bond from the reducing end and remove monomers progressively (Figure 4A,B) [12,19]. Unlike known exo-polygalacturonases, IDSPGA28-16 was incapable of cleaving (GalA)2 (Figure 4E,F), probably because of unproductive substrate binding preventing access of the catalytic residues to the α-1,4 glycosidic bond (Figure 5A,C,E). Taken together, the above stoichiometry and stereochemistry data suggested that IDSPGA28-16 was neither a typical exo-polygalacturonase nor an exo-poly-α-digalacturonosidase. It is an unconventional exo-polygalacturonase.
3.5. Effect of Recombinant Enzymes on Fruit Juice Clarification
Many fruits contain high concentrations of pectin, which is responsible for the high viscosity of fruit pulp and the turbidity and often poor yield of freshly pressed fruit juice. Therefore, the treatment of fruit juice, or pulp with pectinases, such as polygalacturonase, pectin lyase, and rhamnogalacturonase, to degrade pectin has been extensively used industrially to increase juice yield, reduce viscosity, and for clarification of the juice, for increased consumer acceptance [43,44]. Although IDSPGA28-4 and IDSPGA28-16 could not degrade the highly esterified (60% methylated) citrus pectin, a previous study [4] reported that exo-polygalacturonase can degrade lightly esterified pectin and clarify grape juice. The effectiveness of IDSPGA28-4 and IDSPGA28-16 for clarification of orange, grape, and apple juice was determined. Treatment with IDSPGA28-4 significantly increased the light transmittance (%T660) of grape juice (pH = 3.6) from 1.6% to 36.3% (p < 0.001) (Figure 6A), whereas IDSPGA28-16 dramatically increased the %T660 of apple juice (pH = 3.4) from 1.9% to 60.6% (p < 0.001) and grape juice from 1.1% to 43.8% (p < 0.001) (Figure 6B).
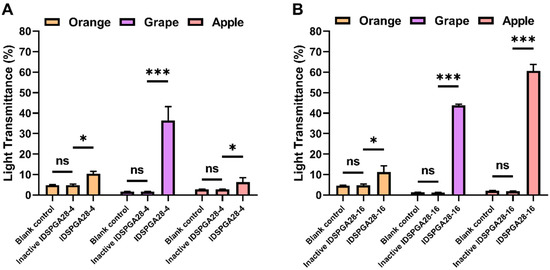
Figure 6.
Juice clarification using IDSPGA28-4 (A) or IDSPGA28-16 (B). ns, not significant; * p < 0.05; *** p < 0.001.
The acidic pH of most fruit juices, such as orange, grape, apple, lemon, and papaya, requires pectinases with acidic pH optima for industrial application [41]. After pre-incubation at pH 3.5 for 1 h, IDSPGA28-4 and IDSPGA28-16 retained ~50% of their original activities (Figure 3E,F), displaying resilience to acidic pH comparable to many previously reported pectinases [4,45,46]. IDSPGA28-4 and IDSPGA28-16 were less effective for clarifying orange juice (pH = 5.6) than grape and apple juice, even though they had high catalytic activity from pH 5.0 to 6.0 (Figure 3E), and the effectiveness in clarifying apple juice was different between two enzymes. The reason may be that the degrees of esterification of pectin contained in these fruits are different [47] and the two enzymes differ in their preference for the degree of pectin esterification. A cocktail of endo- and exo-acting hydrolyses, as well as pectin methylesterases, could further improve the extraction efficiency and clarification of fruit juice [4,8,41], which appears to be more attributable to the variable degree of esterification in pectin.
4. Conclusions
In this study, two exo-polygalacturonases from sheep rumen microbiota, IDSPGA28-4 and IDSPGA28-16, were found to hydrolyze PGA, and the latter was among the most catalytically active exo-polygalacturonases to date. Distinct from IDSPGA28-4 and previously reported exo-polygalacturonases, IDSPGA28-16 was capable of liberating galacturonic acid monomers from substrates only with DP > two, suggesting that it underwent a unique action mode. IDSPGA28-4 and IDSPGA28-16 were effective for fruit juice clarification, showing potential applications in the food and feed industries.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms11030760/s1, Figure S1: Amino acid alignment of bacterial exo-polygalacturonases. Three conserved catalytic residues (D251, D272, D273) are highlighted by triangles and five residues thought to participate in substrate-binding are indicated by arrows. Four key loops involved in substrate binding and catalysis are underlined in blue (IDSPGA28-16) or purple (IDSPGA28-4). Figure S2: Predicted three-dimensional structure and hydrophobic surface of IDSPGA28-4 (A-B) and IDSPGA28-16 (C-D). Exo-polygalacturonase from Thermotoga maritima; (PDB: 3JUR) served as the template for homology modeling. Eight conserved key residues are depicted as sticks. Four key loops involved in substrate binding and catalysis are highlighted in azure (IDSPGA28-16) or violet (IDSPGA28-4). The proposed catalytic center is indicated in red. Table S1: Oligonucleotides used in this study.
Author Contributions
Conceptualization, Q.W.; investigation, Q.D., X.S. and D.G.; software, Y.W. and M.L.; methodology, Y.L.; validation, N.L.; supervision, Q.W., J.W. and Z.W.; funding acquisition, Q.W. and J.W.; writing—original draft, Q.D.; writing—review and editing, Q.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Key R&D Program of Zhejiang Province (2022C02043) and the National Key Research and Development Program of China (2018YFD0501903).
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
The authors appreciate The Experimental Teaching Center, College of Animal Sciences, Zhejiang University, for facility support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef] [PubMed]
- Atmodjo, M.A.; Hao, Z.; Mohnen, D. Evolving views of pectin biosynthesis. Annu. Rev. Plant Biol. 2013, 64, 747–779. [Google Scholar] [CrossRef]
- Hamouda, H.I.; Ali, N.; Su, H.; Feng, J.; Lu, M.; Li, F.L. Exploration of two pectate lyases from Caldicellulosiruptor bescii reveals that the CBM66 module has a crucial role in pectic biomass degradation. Appl. Environ. Microbiol. 2020, 86, e00787-20. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Tu, T.; Zhang, D.; Ma, R.; You, S.; Wang, X.; Yao, B.; Luo, H.; Xu, B. Two acidic, thermophilic GH28 polygalacturonases from Talaromyces leycettanus JCM 12802 with application potentials for grape juice clarification. Food Chem. 2017, 237, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Abbott, D.W.; Boraston, A.B. The structural basis for exopolygalacturonase activity in a family 28 glycoside hydrolase. J. Mol. Biol. 2007, 368, 1215–1222. [Google Scholar] [CrossRef]
- Tu, T.; Meng, K.; Luo, H.; Turunen, O.; Zhang, L.; Cheng, Y.; Su, X.; Ma, R.; Shi, P.; Wang, Y.; et al. New insights into the role of T3 Loop in determining catalytic efficiency of GH28 endo-polygalacturonases. PLoS ONE 2015, 10, e0135413. [Google Scholar] [CrossRef] [PubMed]
- Drula, E.; Garron, M.-L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2022, 50, D571–D577. [Google Scholar] [CrossRef]
- Pan, X.; Li, K.; Ma, R.; Shi, P.; Huang, H.; Yang, P.; Meng, K.; Yao, B. Biochemical characterization of three distinct polygalacturonases from Neosartorya fischeri p1. Food Chem. 2015, 188, 569–575. [Google Scholar] [CrossRef]
- Kars, I.; Krooshof, G.H.; Wagemakers, L.; Joosten, R.; Benen, J.A.E.; Van Kan, J.A.L. Necrotizing activity of five Botrytis cinerea endopolygalacturonases produced in Pichia pastoris. Plant J. 2005, 43, 213–225. [Google Scholar] [CrossRef]
- Shimizu, T.; Nakatsu, T.; Miyairi, K.; Okuno, T.; Kato, H. Active-site architecture of endopolygalacturonase I from Stereum purpureum revealed by crystal structures in native and ligand-bound forms at atomic resolution. Biochemistry 2002, 41, 6651–6659. [Google Scholar] [CrossRef]
- Van Santen, Y.; Benen, J.A.E.; Schröter, K.-H.; Kalk, K.H.; Armand, S.; Visser, J.; Dijkstra, B.W. 1.68-Å crystal structure of endopolygalacturonase II from Aspergillus niger and identification of active site residues by site-directed mutagenesis. J. Biol. Chem. 1999, 274, 30474–30480. [Google Scholar] [CrossRef]
- Kluskens, L.D.; van Alebeek, G.-J.W.M.; Walther, J.; Voragen, A.G.J.; de Vos, W.M.; van der Oost, J. Characterization and mode of action of an exopolygalacturonase from the hyperthermophilic bacterium Thermotoga maritima. FEBS J. 2005, 272, 5464–5473. [Google Scholar] [CrossRef]
- Martens-Uzunova, E.S.; Zandleven, J.S.; Benen, J.A.E.; Awad, H.; Kools, H.J.; Beldman, G.; Voragen, A.G.J.; Van Den Berg, J.A.; Schaap, P.J. A new group of exo-acting family 28 glycoside hydrolases of Aspergillus niger that are involved in pectin degradation. Biochem. J. 2006, 400, 43–52. [Google Scholar] [CrossRef]
- Sawada, K.; Suzumatsu, A.; Kobayashi, T.; Ito, S. Molecular cloning and sequencing of the gene encoding an exopolygalacturonase of a Bacillus isolate and properties of its recombinant enzyme. Biochim. Biophys. Acta 2001, 1568, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Ariaeenejad, S.; Sheykh Abdollahzadeh Mamaghani, A.; Maleki, M.; Kavousi, K.; Foroozandeh Shahraki, M.; Hosseini Salekdeh, G. A novel high performance in-silico screened metagenome-derived alkali-thermostable endo-β-1,4-glucanase for lignocellulosic biomass hydrolysis in the harsh conditions. BMC Biotechnol. 2020, 20, 56. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.W.; Deng, Q.; Gao, D.Y.; He, B.; Yin, S.J.; Qian, L.C.; Wang, J.K.; Wang, Q. A novel bifunctional glucanase exhibiting high production of glucose and cellobiose from rumen bacterium. Int. J. Biol. Macromol. 2021, 173, 136–145. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Jin, S.W.; Cao, J.W.; Mi, L.; Wang, J.K. Metatranscriptomics of the Hu sheep rumen microbiome reveals novel cellulases. Biotechnol. Biofuels 2019, 12, 153. [Google Scholar] [CrossRef]
- Wang, Q.; Luo, Y.; He, B.; Jiang, L.S.; Liu, J.X.; Wang, J.K. Characterization of a novel xylanase gene from rumen content of Hu sheep. Appl. Biochem. Biotechnol. 2015, 177, 1424–1436. [Google Scholar] [CrossRef]
- Luis, A.S.; Briggs, J.; Zhang, X.; Farnell, B.; Ndeh, D.; Labourel, A.; Baslé, A.; Cartmell, A.; Terrapon, N.; Stott, K.; et al. Dietary pectic glycans are degraded by coordinated enzyme pathways in human colonic Bacteroides. Nat. Microbiol. 2018, 3, 210–219. [Google Scholar] [CrossRef]
- Ndeh, D.; Rogowski, A.; Cartmell, A.; Luis, A.S.; Baslé, A.; Gray, J.; Venditto, I.; Briggs, J.; Zhang, X.; Labourel, A.; et al. Complex pectin metabolism by gut bacteria reveals novel catalytic functions. Nature 2017, 544, 65–70. [Google Scholar] [CrossRef]
- Bailey, M.J.; Biely, P.; Poutanen, K. Interlaboratory testing of methods for assay of xylanase activity. J. Biotechnol. 1992, 23, 257–270. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: AutoDock vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. Ff14sb: Improving the accuracy of protein side chain and backbone parameters from ff99sb. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and testing of a general amber force field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: LINCS: A linear constraint solver for molecular simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Lu, T.; Chen, F. Multiwfn: A multifunctional wavefunction analyzer. J. Comput. Chem. 2012, 33, 580–592. [Google Scholar] [CrossRef]
- Lefebvre, C.; Rubez, G.; Khartabil, H.; Boisson, J.-C.; Contreras-García, J.; Hénon, E. Accurately extracting the signature of intermolecular interactions present in the NCI plot of the reduced density gradient versus electron density. Phys. Chem. Chem. Phys. 2017, 19, 17928–17936. [Google Scholar] [CrossRef]
- Hess, M.; Sczyrba, A.; Egan, R.; Kim, T.-W.; Chokhawala, H.; Schroth, G.; Luo, S.; Clark, D.S.; Chen, F.; Zhang, T.; et al. Metagenomic discovery of biomass-degrading genes and genomes from cow rumen. Science 2011, 331, 463–467. [Google Scholar] [CrossRef]
- Pijning, T.; van Pouderoyen, G.; Kluskens, L.; van der Oost, J.; Dijkstra, B.W. The crystal structure of a hyperthermoactive exopolygalacturonase from Thermotoga maritima reveals a unique tetramer. FEBS Lett. 2009, 583, 3665–3670. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.C.; Lee, J.H.; Han, Y.; Choi, J.H.; Song, J.J. A novel multifunctional cellulolytic enzyme screened from metagenomic resources representing ruminal bacteria. Biochem. Biophys. Res. Commun. 2013, 441, 567–572. [Google Scholar] [CrossRef]
- Dadheech, T.; Shah, R.; Pandit, R.; Hinsu, A.; Chauhan, P.S.; Jakhesara, S.; Kunjadiya, A.; Rank, D.; Joshi, C. Cloning, molecular modeling and characterization of acidic cellulase from buffalo rumen and its applicability in saccharification of lignocellulosic biomass. Int. J. Biol. Macromol. 2018, 113, 73–81. [Google Scholar] [CrossRef]
- Song, Y.H.; Lee, K.T.; Baek, J.Y.; Kim, M.J.; Kwon, M.R.; Kim, Y.J.; Park, M.R.; Ko, H.; Lee, J.S.; Kim, K.S. Isolation and characterization of a novel endo-β-1,4-glucanase from a metagenomic library of the black-goat rumen. Braz. J. Microbiol. 2017, 48, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Cecchini, D.A.; Pepe, O.; Pennacchio, A.; Fagnano, M.; Faraco, V. Directed evolution of the bacterial endo-β-1,4-glucanase from Streptomyces sp. G12 towards improved catalysts for lignocellulose conversion. AMB Express 2018, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.Y.; Sun, X.B.; Liu, M.Q.; Liu, Y.N.; Zhang, H.E.; Shi, X.L.; Li, Y.N.; Wang, J.K.; Yin, S.J.; Wang, Q. Characterization of thermostable and chimeric enzymes via isopeptide bond-mediated molecular cyclization. J. Agric. Food Chem. 2019, 67, 6837–6846. [Google Scholar] [CrossRef] [PubMed]
- Kester, H.C.M.; Someren, M.A.K.-V.; Müller, Y.; Visser, J. Primary structure and characterization of an exopolygalacturonase from Aspergillus tubingensis. Eur. J. Biochem. 1996, 240, 738–746. [Google Scholar] [CrossRef]
- Pařenicová, L.; Kester, H.C.M.; Benen, J.A.E.; Visser, J. Characterization of a novel endopolygalacturonase from Aspergillus niger with unique kinetic properties. FEBS Lett. 2000, 467, 333–336. [Google Scholar] [CrossRef]
- Maiorov, V.N.; Crippen, G.M. Significance of root-mean-square deviation in comparing three-dimensional structures of globular proteins. J. Mol. Biol. 1994, 235, 625–634. [Google Scholar] [CrossRef]
- Tu, T.; Meng, K.; Huang, H.; Luo, H.; Bai, Y.; Ma, R.; Su, X.; Shi, P.; Yang, P.; Wang, Y.; et al. Molecular Characterization of a thermophilic endo-polygalacturonase from Thielavia arenaria XZ7 with high catalytic efficiency and application potential in the food and feed industries. J. Agric. Food Chem. 2014, 62, 12686–12694. [Google Scholar] [CrossRef]
- Tu, T.; Meng, K.; Bai, Y.; Shi, P.; Luo, H.; Wang, Y.; Yang, P.; Zhang, Y.; Zhang, W.; Yao, B. High-yield production of a low-temperature-active polygalacturonase for papaya juice clarification. Food Chem. 2013, 141, 2974–2981. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Li, S.; Tan, H.; Li, K.; Chen, W.; Yin, H. A highly efficient biocatalytic conversion of renewable pectic polysaccharide biomass into bioactive oligogalacturonides. ACS Food Sci. Technol. 2021, 1, 338–346. [Google Scholar] [CrossRef]
- Sieiro, C.; García-Fraga, B.; López-Seijas, J.; da Silva, A.F.; Villa, T.G. Microbial Pectic Enzymes in the Food and Wine Industry; IntechOpen: London, UK, 2012; ISBN 978-953-307-905-9. [Google Scholar]
- Wang, J.; Zhang, Y.; Qin, X.; Gao, L.; Han, B.; Zhang, D.; Li, J.; Huang, H.; Zhang, W. Efficient expression of an acidic endo-polygalacturonase from Aspergillus niger and its application in juice production. J. Agric. Food Chem. 2017, 65, 2730–2736. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Huo, W.; Dai, X.; Dang, Y. Preparation of low-molecular-weight citrus pectin by recombinant Bacillus subtilis pectate lyase and promotion of growth of Bifidobacterium longum. Catal. Commun. 2018, 107, 39–42. [Google Scholar] [CrossRef]
- Sassi, A.H.; Tounsi, H.; Trigui-Lahiani, H.; Bouzouita, R.; Romdhane, Z.B.; Gargouri, A. A low-temperature polygalacturonase from P. occitanis: Characterization and application in juice clarification. Int. J. Biol. Macromol. 2016, 91, 158–164. [Google Scholar] [CrossRef]
- Leroux, J.; Langendorff, V.; Schick, G.; Vaishnav, V.; Mazoyer, J. Emulsion stabilizing properties of pectin. Food Hydrocoll. 2003, 17, 455–462. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).