
Comparison of Atmospheric and Lithospheric Culturable Bacterial Communities from Two Dissimilar Active Volcanic Sites, Surtsey Island and Fimmvörðuháls Mountain in Iceland
 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
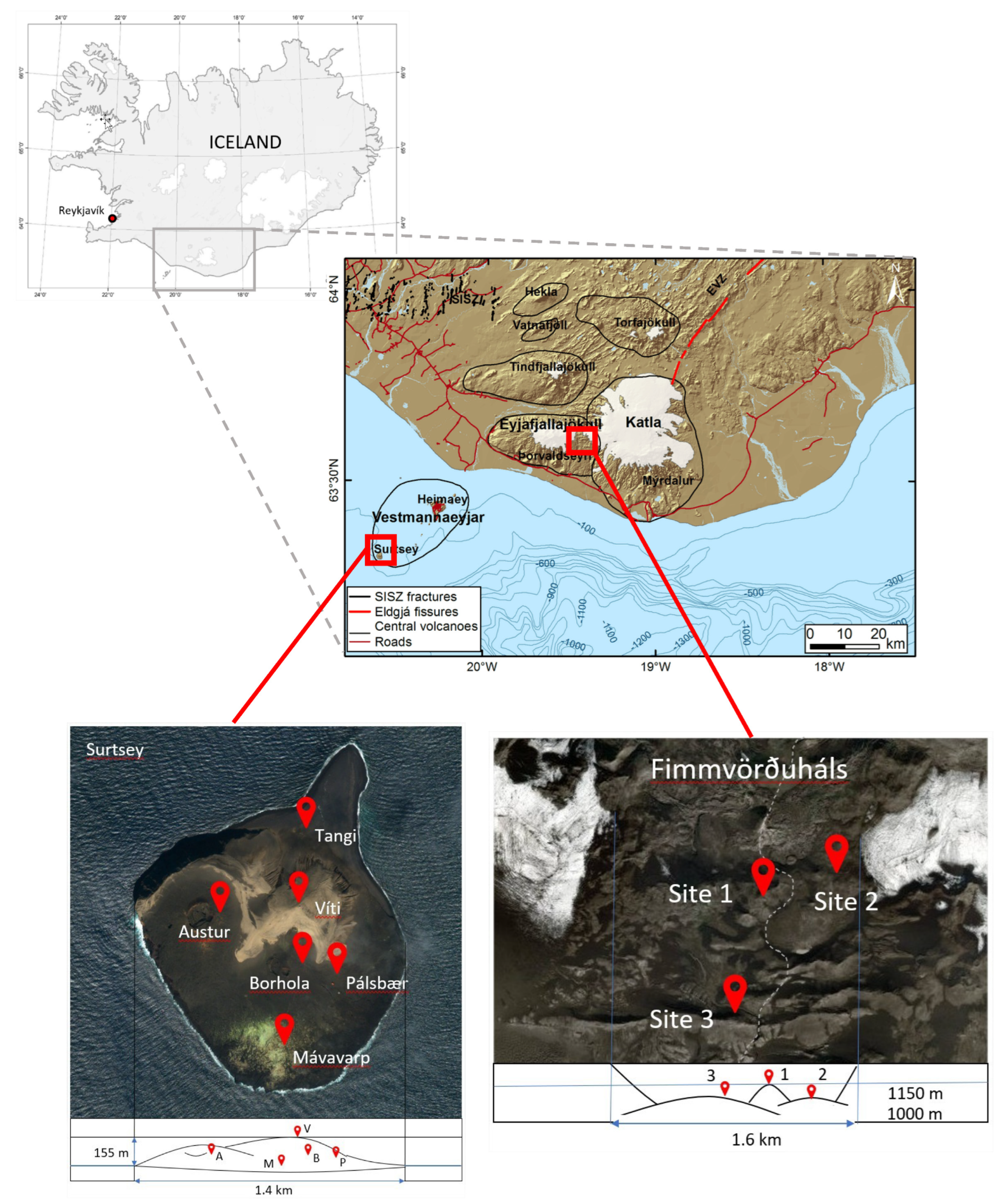
2.1. Site Description and Sample Collection
2.2. Isolation, Characterization and Grouping by MALDI-TOF Mass Spectrometry of the Microbial Strains
2.3. Identification of Group Representatives by 16S rRNA Gene Sequencing
2.4. Construction of Phylogenetic Trees
2.5. Comparison of the Microbial Communities Using Statistical Analyses
2.6. Predictive Sources of the Cultivated Isolates
2.7. Physiological Survival Properties of the Isolates
3. Results
3.1. Isolation, Characterization and Grouping by MALDI-TOF Mass Spectrometry of the Microbial Stains
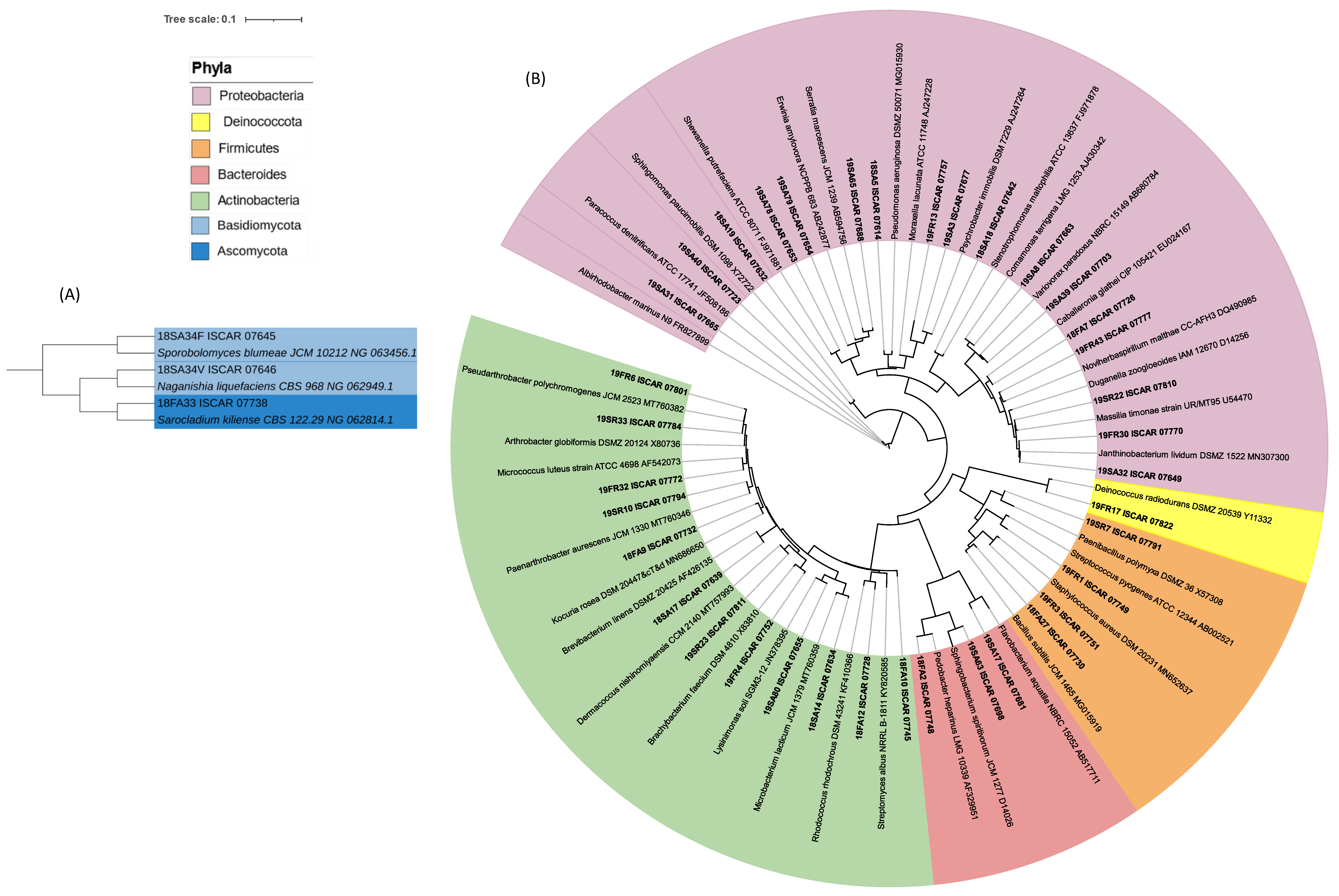
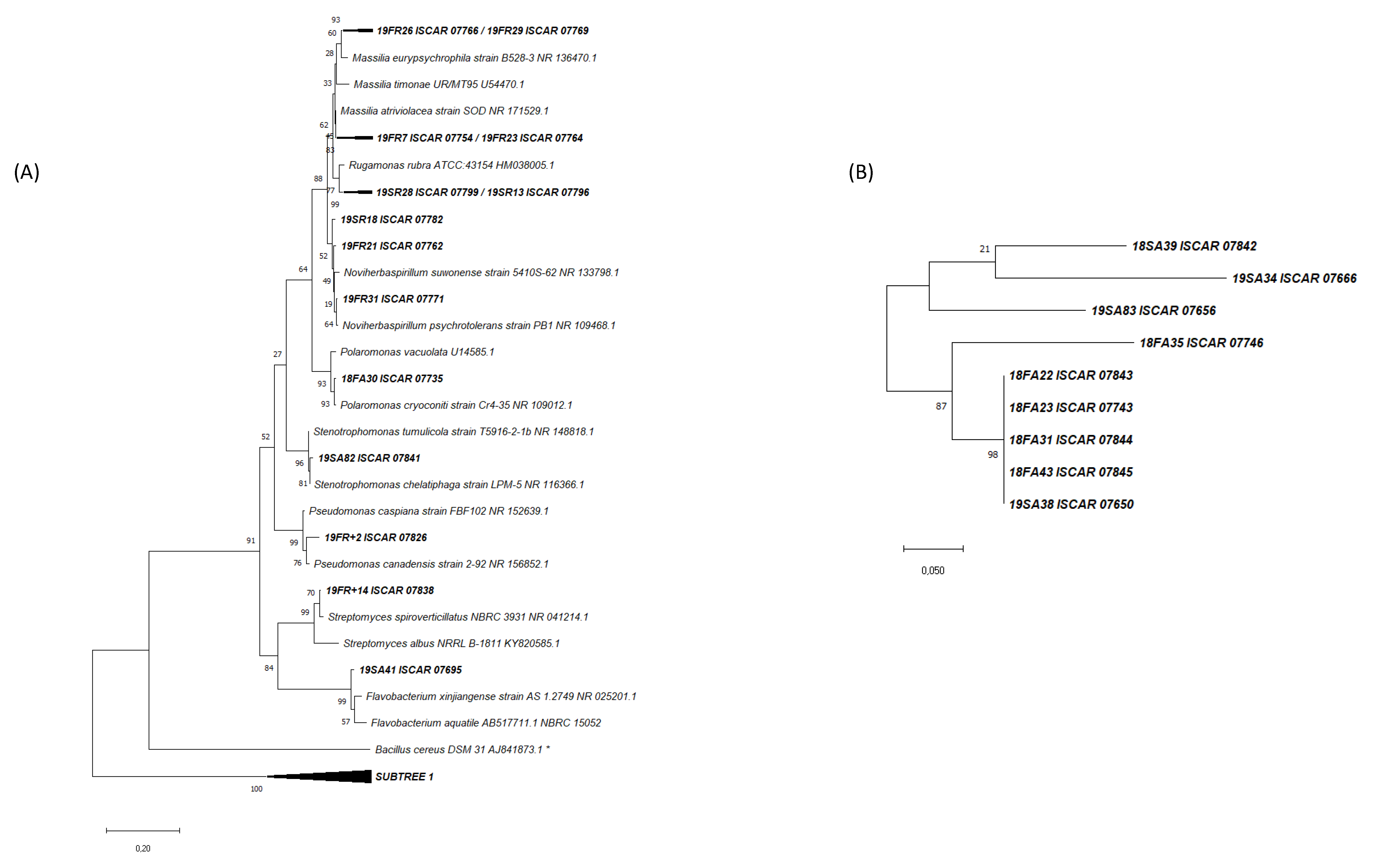
3.2. Phylogenetic Analysis of the Cultivated Microbial Isolates
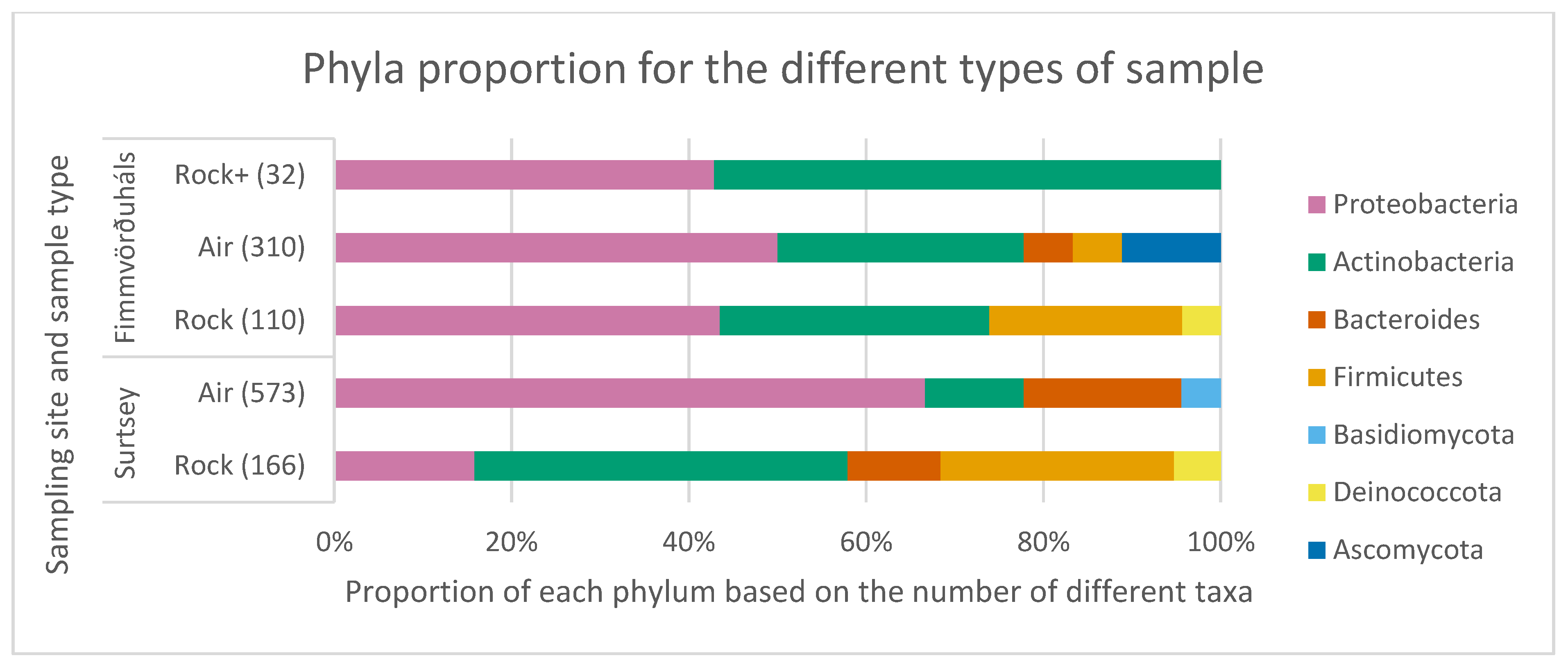
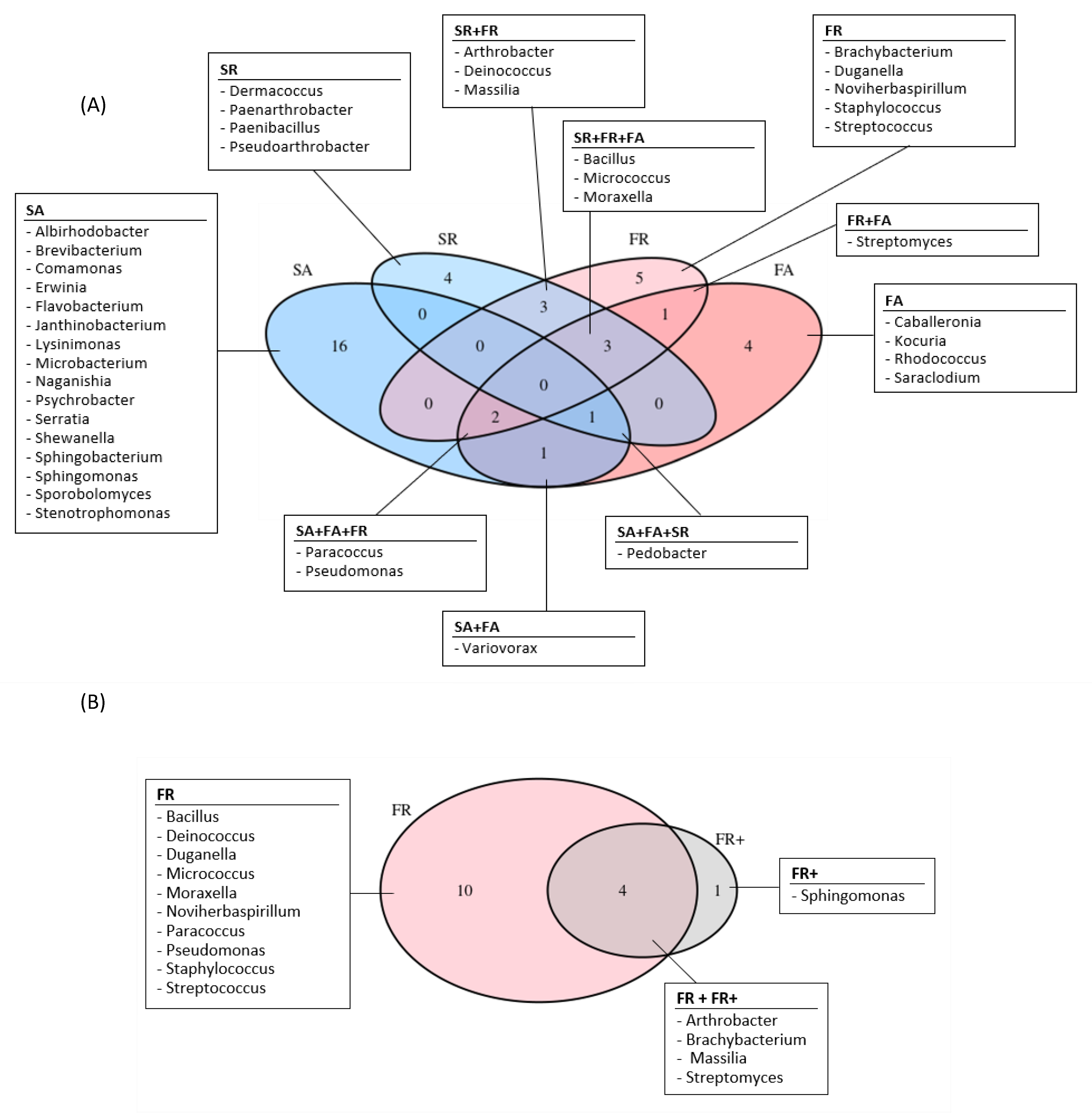
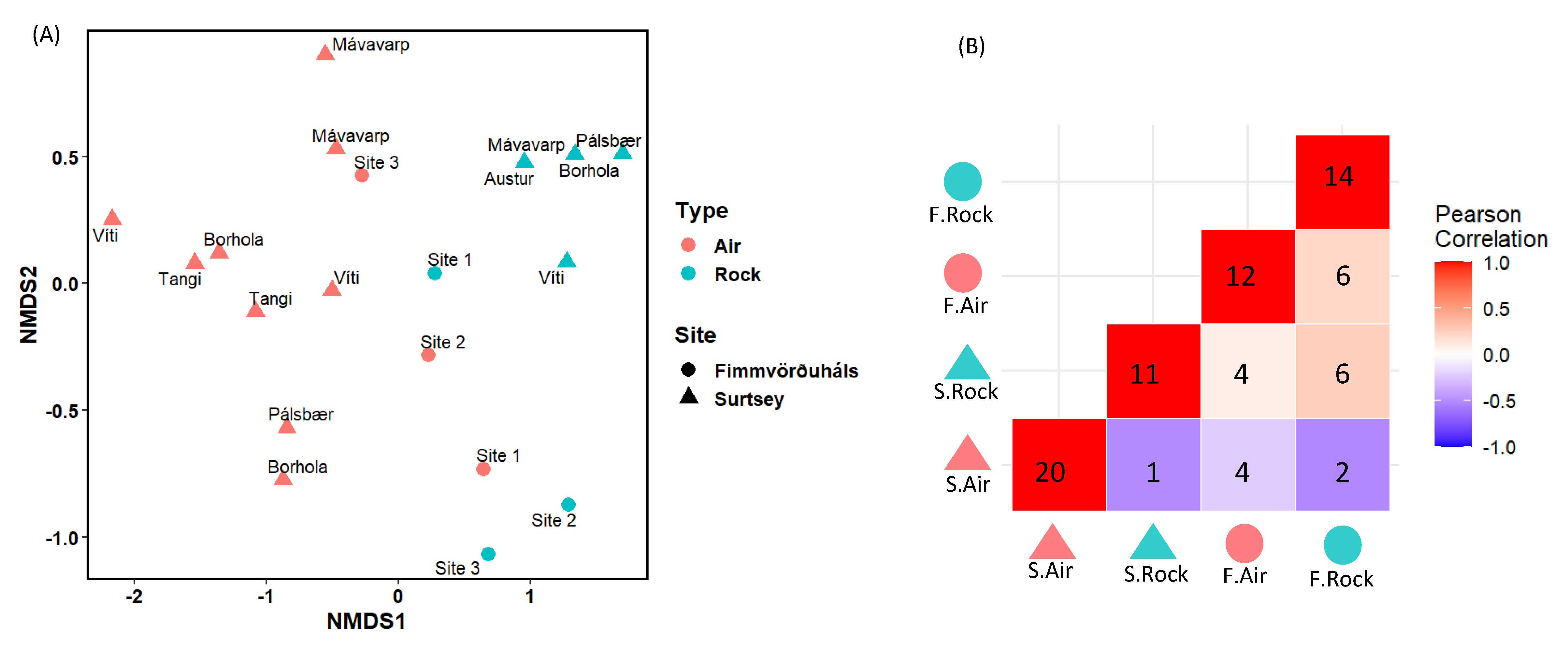
3.3. Comparison of the Microbial Communities Using Statistical Analyses
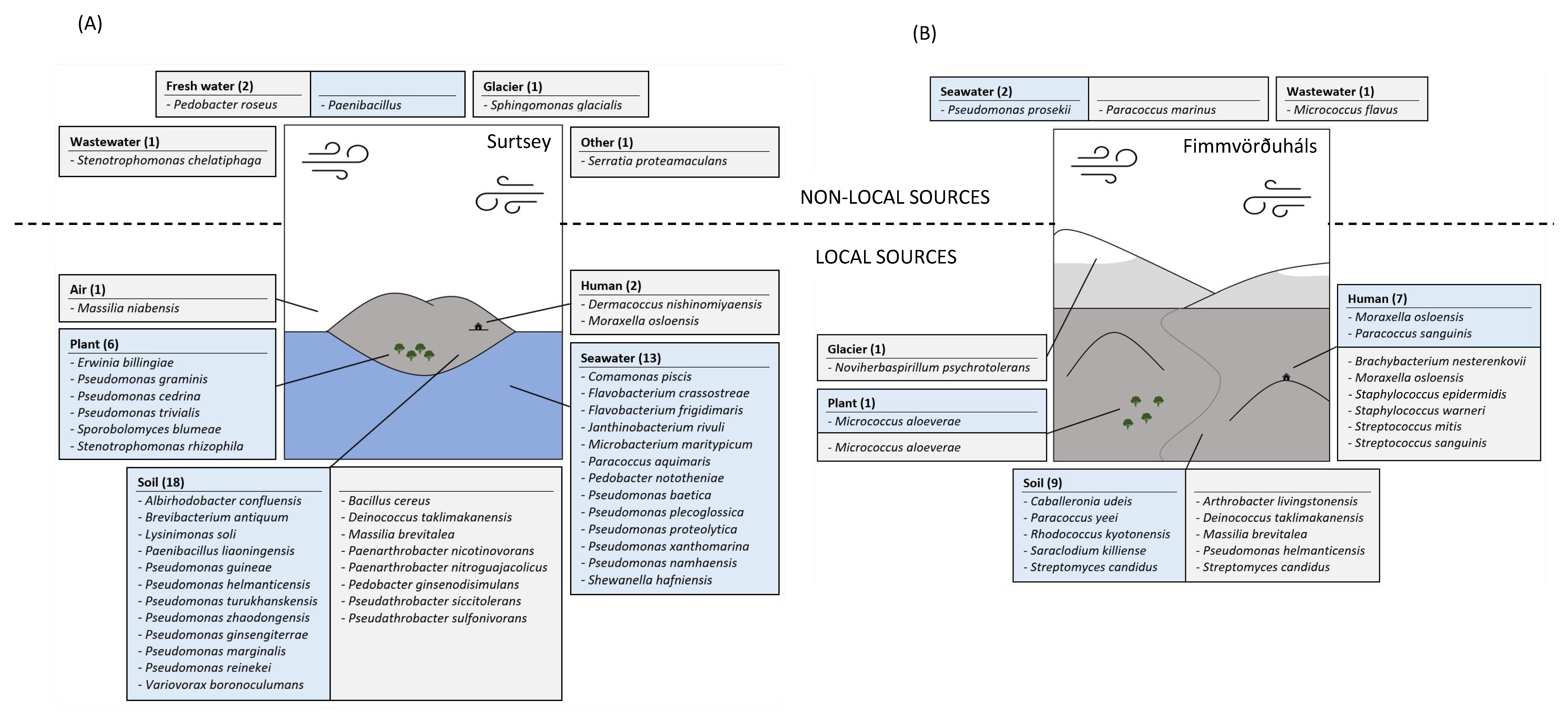
3.4. Predictive Sources of the Cultivated Isolates
3.5. Physiological Survival Properties of the Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xie, W.; Li, Y.; Bai, W.; Hou, J.; Ma, T.; Zeng, X.; Zhang, L.; An, T. The Source and Transport of Bioaerosols in the Air: A Review. Front. Environ. Sci. Eng. 2021, 15, 44. [Google Scholar] [CrossRef] [PubMed]
- Mayol, E.; Arrieta, J.M.; Jiménez, M.A.; Martínez-Asensio, A.; Garcias-Bonet, N.; Dachs, J.; González-Gaya, B.; Royer, S.-J.; Benítez-Barrios, V.M.; Fraile-Nuez, E.; et al. Long-Range Transport of Airborne Microbes over the Global Tropical and Subtropical Ocean. Nat. Commun. 2017, 8, 201. [Google Scholar] [CrossRef]
- Moore, R.A.; Bomar, C.; Kobziar, L.N.; Christner, B.C. Wildland Fire as an Atmospheric Source of Viable Microbial Aerosols and Biological Ice Nucleating Particles. ISME J. 2021, 15, 461–472. [Google Scholar] [CrossRef]
- González-Toril, E.; Osuna, S.; Viúdez-Moreiras, D.; Navarro-Cid, I.; del Toro, S.D.; Sor, S.; Bardera, R.; Puente-Sánchez, F.; de Diego-Castilla, G.; Aguilera, Á. Impacts of Saharan Dust Intrusions on Bacterial Communities of the Low Troposphere. Sci. Rep. 2020, 10, 6837. [Google Scholar] [CrossRef] [PubMed]
- Morales-Baquero, R.; Pérez-Martínez, C. Saharan versus Local Influence on Atmospheric Aerosol Deposition in the Southern Iberian Peninsula: Significance for N and P Inputs. Glob. Biogeochem. Cycles 2016, 30, 501–513. [Google Scholar] [CrossRef]
- Reche, I.; D’Orta, G.; Mladenov, N.; Winget, D.M.; Suttle, C.A. Deposition Rates of Viruses and Bacteria above the Atmospheric Boundary Layer. ISME J. 2018, 12, 1154–1162. [Google Scholar] [CrossRef]
- DasSarma, P.; DasSarma, S. Survival of Microbes in Earth’s Stratosphere. Curr. Opin. Microbiol. 2018, 43, 24–30. [Google Scholar] [CrossRef]
- Moeller, R.; Horneck, G.; Facius, R.; Stackebrandt, E. Role of Pigmentation in Protecting bacillus sp. Endospores against Environmental UV Radiation. FEMS Microbiol. Ecol. 2005, 51, 231–236. [Google Scholar] [CrossRef]
- Danevčič, T.; Borić Vezjak, M.; Tabor, M.; Zorec, M.; Stopar, D. Prodigiosin Induces Autolysins in Actively Grown Bacillus Subtilis Cells. Front. Microbiol. 2016, 7, 27. [Google Scholar] [CrossRef]
- Narsing Rao, M.P.; Xiao, M.; Li, W.-J. Fungal and Bacterial Pigments: Secondary Metabolites with Wide Applications. Front. Microbiol. 2017, 8, 1113. [Google Scholar] [CrossRef]
- Fröhlich-Nowoisky, J.; Kampf, C.J.; Weber, B.; Huffman, J.A.; Pöhlker, C.; Andreae, M.O.; Lang-Yona, N.; Burrows, S.M.; Gunthe, S.S.; Elbert, W.; et al. Bioaerosols in the Earth System: Climate, Health, and Ecosystem Interactions. Atmos. Res. 2016, 182, 346–376. [Google Scholar] [CrossRef]
- Hoose, C.; Kristjánsson, J.E.; Burrows, S.M. How Important Is Biological Ice Nucleation in Clouds on a Global Scale? Environ. Res. Lett. 2010, 5, 024009. [Google Scholar] [CrossRef]
- Sahyoun, M.; Wex, H.; Gosewinkel, U.; Šantl-Temkiv, T.; Nielsen, N.W.; Finster, K.; Sørensen, J.H.; Stratmann, F.; Korsholm, U.S. On the Usage of Classical Nucleation Theory in Quantification of the Impact of Bacterial INP on Weather and Climate. Atmos. Environ. 2016, 139, 230–240. [Google Scholar] [CrossRef]
- Hummel, M.; Hoose, C.; Gallagher, M.; Healy, D.A.; Huffman, J.A.; O’Connor, D.; Pöschl, U.; Pöhlker, C.; Robinson, N.H.; Schnaiter, M.; et al. Regional-Scale Simulations of Fungal Spore Aerosols Using an Emission Parameterization Adapted to Local Measurements of Fluorescent Biological Aerosol Particles. Atmos. Chem. Phys. 2015, 15, 6127–6146. [Google Scholar] [CrossRef]
- Woo, C.; Yamamoto, N. Falling Bacterial Communities from the Atmosphere. Environ. Microbiome 2020, 15, 22. [Google Scholar] [CrossRef]
- Azman, A.-S.; Mawang, C.-I.; Abubakar, S. Bacterial Pigments: The Bioactivities and as an Alternative for Therapeutic Applications. Nat. Prod. Commun. 2018, 13, 1747–1754. [Google Scholar] [CrossRef]
- Saviola, B. Pigments of Pathogenic Bacteria. J. Microbiol. Exp. 2018, 6, 114–115. [Google Scholar] [CrossRef]
- Liu, G.Y.; Nizet, V. Color Me Bad: Microbial Pigments as Virulence Factors. Trends Microbiol. 2009, 17, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Sigurdsson, A.R. The Bentlionic Coastal Fauna of Surtsey in 1968. In Surtsey Research Progress Report IV; The zoology of Iceland; Munksgaard, E., Ed.; University of California: Berkeley, CA, USA, 1970; Volume II-IV. [Google Scholar]
- Kelly, L.C.; Cockell, C.S.; Thorsteinsson, T.; Marteinsson, V.; Stevenson, J. Pioneer Microbial Communities of the Fimmvörðuháls Lava Flow, Eyjafjallajökull, Iceland. Microb. Ecol. 2014, 68, 504–518. [Google Scholar] [CrossRef] [PubMed]
- Bergsten, P.; Vannier, P.; Klonowski, A.M.; Knobloch, S.; Gudmundsson, M.T.; Jackson, M.D.; Marteinsson, V.T. Basalt-Hosted Microbial Communities in the Subsurface of the Young Volcanic Island of Surtsey, Iceland. Front. Microbiol. 2021, 12, 728977. [Google Scholar] [CrossRef]
- Šantl-Temkiv, T.; Amato, P.; Gosewinkel, U.; Thyrhaug, R.; Charton, A.; Chicot, B.; Finster, K.; Bratbak, G.; Löndahl, J. High-Flow-Rate Impinger for the Study of Concentration, Viability, Metabolic Activity, and Ice-Nucleation Activity of Airborne Bacteria. Environ. Sci. Technol. 2017, 51, 11224–11234. [Google Scholar] [CrossRef]
- Hyvärinen, A.M.; Martikainen, P.J.; Nevalainen, A.I. Suitability of Poor Medium in Counting Total Viable Airborne Bacteria. Grana 1991, 30, 414–417. [Google Scholar] [CrossRef]
- Chinivasagam, H.N.; Blackall, P.J. Investigation and Application of Methods for Enumerating Heterotrophs and Escherichia Coli in the Air within Piggery Sheds. J. Appl. Microbiol. 2005, 98, 1137–1145. [Google Scholar] [CrossRef]
- Marteinsson, V.; Klonowski, A.; Reynisson, E.; Vannier, P.; Sigurdsson, B.D.; Ólafsson, M. Microbial Colonization in Diverse Surface Soil Types in Surtsey and Diversity Analysis of Its Subsurface Microbiota. Biogeosciences 2015, 12, 1191–1203. [Google Scholar] [CrossRef]
- Sauer, S.; Freiwald, A.; Maier, T.; Kube, M.; Reinhardt, R.; Kostrzewa, M.; Geider, K. Classification and Identification of Bacteria by Mass Spectrometry and Computational Analysis. PLoS ONE 2008, 3, e2843. [Google Scholar] [CrossRef]
- Skirnisdottir, S.; Hreggvidsson, G.O.; Hjörleifsdottir, S.; Marteinsson, V.T.; Petursdottir, S.K.; Holst, O.; Kristjansson, J.K. Influence of Sulfide and Temperature on Species Composition and Community Structure of Hot Spring Microbial Mats. Appl. Environ. Microbiol. 2000, 66, 2835–2841. [Google Scholar] [CrossRef] [PubMed]
- Gorbushina, A.A. Life on the Rocks. Environ. Microbiol. 2007, 9, 1613–1631. [Google Scholar] [CrossRef]
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.C.; Levy, S.B.; Jackson, R.W. Pseudomonas Genomes: Diverse and Adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef]
- Wickham, H.; Bryan, J. Readxl: Read Excel Files R Package Version 1.3.1; R Package: Vienna, Austria, 2019. [Google Scholar]
- Oksanen, J. Vegan: Community Ecology Package. R package Version 2.5-3. 2018. Available online: https://CRAN.R-project.org/package=vegan (accessed on 10 February 2023).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Pedersen, T. Ggforce: Accelerating ‘Ggplot2’. 2022. Available online: https://github.com/thomasp85/ggforce (accessed on 10 February 2023).
- Slowikiwski, K. Ggrepel: Automatically Position Non-Overlapping Labels with “Ggplot2.” 2021. Available online: https://CRAN.R-project.org/package=ggrepel (accessed on 10 February 2023).
- Wickham, H. Reshaping Data with the Reshape Package. J. Stat. Softw. 2007, 21, 1–20. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V. R Package ‘Corrplot’: Visualization of a Correlation Matrix (Version 0.92). 2021. Available online: https://github.com/taiyun/corrplot (accessed on 10 February 2023).
- Stunder, B.J.B. Stunder, Global Data Assimilation System (GDAS) Archive Information; NOAA Air Resources Laboratory: Silver Spring, MD, USA, 2004. [Google Scholar]
- Stein, A.F.; Draxler, R.R.; Rolph, G.D.; Stunder, B.J.B.; Cohen, M.D.; Ngan, F. NOAA’s HYSPLIT Atmospheric Transport and Dispersion Modeling System. Bull. Am. Meteorol. Soc. 2015, 96, 2059–2077. [Google Scholar] [CrossRef]
- Seifried, J.S.; Wichels, A.; Gerdts, G. Spatial Distribution of Marine Airborne Bacterial Communities. Microbiologyopen 2015, 4, 475–490. [Google Scholar] [CrossRef]
- Jang, Y.-H.; Kim, S.-J.; Tamura, T.; Hamada, M.; Weon, H.-Y.; Suzuki, K.; Kwon, S.-W.; Kim, W.-G. Lysinimonas soli Gen. Nov., Sp. Nov., Isolated from Soil, and Reclassification of Leifsonia Kribbensis Dastager et al. 2009 as Lysinimonas kribbensis sp. Nov., Comb. Nov. Int. J. Syst. Evol. Microbiol. 2013, 63, 1403–1410. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Lee, S.-S. Paracoccus aquimaris sp. Nov., Isolated from Seawater. Antonie Van Leeuwenhoek 2015, 108, 871–877. [Google Scholar] [CrossRef]
- Nogi, Y.; Soda, K.; Oikawa, T. Flavobacterium frigidimaris sp. Nov., Isolated from Antarctic Seawater. Syst. Appl. Microbiol. 2005, 28, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-C.; Busse, H.-J.; Liu, H.-C.; Zhou, Y.-G.; Schinner, F.; Margesin, R. Sphingomonas glacialis sp. nov., a Psychrophilic Bacterium Isolated from Alpine Glacier Cryoconite. Int. J. Syst. Evol. Microbiol. 2011, 61, 587–591. [Google Scholar] [CrossRef]
- DSMZ. Leibniz Institut DSMZ-Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH; Curators of the DSMZ; DSM 937, Streptomyces Candidus DSM 40141 is a Mesophilic Bacterium that was Isolated from Soil. Available online: https://bacdive.dsmz.de/strain/15065 (accessed on 10 February 2023).
- Otto, M. Staphylococcus Epidermidis—The “accidental” Pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Bajerski, F.; Ganzert, L.; Mangelsdorf, K.; Lipski, A.; Busse, H.-J.; Padur, L.; Wagner, D. Herbaspirillum psychrotolerans sp. Nov., a Member of the Family Oxalobacteraceae from a Glacier Forefield. Int. J. Syst. Evol. Microbiol. 2013, 63, 3197–3203. [Google Scholar] [CrossRef]
- Khan, S.T.; Takaichi, S.; Harayama, S. Paracoccus marinus sp. Nov., an Adonixanthin Diglucoside-Producing Bacterium Isolated from Coastal Seawater in Tokyo Bay. Int. J. Syst. Evol. Microbiol. 2008, 58, 383–386. [Google Scholar] [CrossRef]
- Ganzert, L.; Bajerski, F.; Mangelsdorf, K.; Lipski, A.; Wagner, D. Arthrobacter livingstonensis sp. nov. and Arthrobacter cryotolerans sp. nov., Salt-Tolerant and Psychrotolerant Species from Antarctic Soil. Int. J. Syst. Evol. Microbiol. 2011, 61, 979–984. [Google Scholar] [CrossRef]
- Lee, J.-S.; Lee, K.C.; Pyun, Y.-R.; Bae, K.S. Arthrobacter koreensis sp. Nov., a Novel Alkalitolerant Bacterium from Soil. Int. J. Syst. Evol. Microbiol. 2003, 53, 1277–1280. [Google Scholar] [CrossRef]
- Brummaier, T.; Hinfothong, P.; Soe, N.L.; Tongmanakit, J.; Watthanaworawit, W.; Ling, C. Brachybacterium nesterenkovii Isolated from a Human Blood Culture—A First Report. New Microbes New Infect. 2020, 36, 100699. [Google Scholar] [CrossRef]
- Gavrish, E.I.; Krauzova, V.I.; Potekhina, N.V.; Karasev, S.G.; Plotnikova, E.G.; Altyntseva, O.V.; Korosteleva, L.A.; Evtushenko, L.I. Three New Species of Brevibacteria--Brevibacterium antiquum sp. Nov., Brevibacterium aurantiacum sp. nov. and Brevibacterium permense sp. Nov. Mikrobiologiia 2004, 73, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Soo Kim, P.; Hyun, D.-W.; Lee, J.-Y.; Sik Kim, H.; Joon Oh, S.; Shin, N.-R.; Bae, J.-W. Comamonas piscis sp. nov., Isolated from the Intestine of a Korean Rockfish, Sebastes Schlegelii. Int. J. Syst. Evol. Microbiol. 2016, 66, 780–785. [Google Scholar] [CrossRef]
- Liu, Z.; Kim, M.C.; Wang, L.; Zhu, G.; Zhang, Y.; Huang, Y.; Wei, Z.; Danzeng, W.; Peng, F. Deinococcus taklimakanensis sp. nov., Isolated from Desert Soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 4311–4316. [Google Scholar] [CrossRef]
- Humphry, D.R.; George, A.; Black, G.W.; Cummings, S.P. Flavobacterium frigidarium sp. Nov., an Aerobic, Psychrophilic, Xylanolytic and Laminarinolytic Bacterium from Antarctica. Int. J. Syst. Evol. Microbiol. 2001, 51, 1235–1243. [Google Scholar] [CrossRef]
- Lu, H.; Deng, T.; Cai, Z.; Liu, F.; Yang, X.; Wang, Y.; Xu, M. Janthinobacterium violaceinigrum sp. Nov., Janthinobacterium aquaticum sp. Nov. and Janthinobacterium rivuli sp. Nov., Isolated from a Subtropical Stream in China. Int. J. Syst. Evol. Microbiol. 2020, 70, 2719–2725. [Google Scholar] [CrossRef] [PubMed]
- Zul, D.; Wanner, G.; Overmann, J. Massilia brevitalea sp. Nov., a Novel Betaproteobacterium Isolated from Lysimeter Soil. Int. J. Syst. Evol. Microbiol. 2008, 58, 1245–1251. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Liu, Y.; Gu, Z.; Xu, B.; Wang, N.; Jiao, N.; Liu, H.; Zhou, Y. Massilia eurypsychrophila sp. Nov. a Facultatively Psychrophilic Bacteria Isolated from Ice Core. Int. J. Syst. Evol. Microbiol. 2015, 65, 2124–2129. [Google Scholar] [CrossRef]
- Guo, B.; Liu, Y.; Gu, Z.; Shen, L.; Liu, K.; Wang, N.; Xing, T.; Liu, H.; Zhou, Y.; Li, J. Massilia psychrophila sp. Nov., Isolated from an Ice Core. Int. J. Syst. Evol. Microbiol. 2016, 66, 4088–4093. [Google Scholar] [CrossRef]
- Weon, H.-Y.; Kim, B.-Y.; Hong, S.-B.; Jeon, Y.-A.; Koo, B.-S.; Kwon, S.-W.; Stackebrandt, E. Massilia niabensis sp. Nov. and Massilia niastensis sp. Nov., Isolated from Air Samples. Int. J. Syst. Evol. Microbiol. 2009, 59, 1656–1660. [Google Scholar] [CrossRef]
- Leibniz Institut DSMZ-Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH; Curators of the DSMZ; DSM 12512, Microbacterium Maritypicum 319 is a Mesophilic Bacterium that was Isolated from Sea Water and Marine Mud. Available online: https://bacdive.dsmz.de/strain/7409 (accessed on 10 February 2023).
- Prakash, O.; Nimonkar, Y.; Munot, H.; Sharma, A.; Vemuluri, V.R.; Chavadar, M.S.; Shouche, Y.S. Description of Micrococcus aloeverae sp. Nov., an Endophytic Actinobacterium Isolated from Aloe Vera. Int. J. Syst. Evol. Microbiol. 2014, 64, 3427–3433. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Wang, B.-J.; Jiang, C.-Y.; Liu, S.-J. Micrococcus flavus sp. Nov., Isolated from Activated Sludge in a Bioreactor. Int. J. Syst. Evol. Microbiol. 2007, 57, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Grewal, P.S. Endotoxin Activity of Moraxella Osloensis against the Grey Garden Slug, Deroceras reticulatum. Appl. Environ. Microbiol. 2002, 68, 3943–3947. [Google Scholar] [CrossRef]
- Lin, S.-Y.; Hameed, A.; Arun, A.B.; Liu, Y.-C.; Hsu, Y.-H.; Lai, W.-A.; Rekha, P.D.; Young, C.-C. Description of Noviherbaspirillum malthae Gen. Nov., sp. Nov., Isolated from an Oil-Contaminated Soil, and Proposal to Reclassify Herbaspirillum Soli, Herbaspirillum aurantiacum, Herbaspirillum canariense and Herbaspirillum psychrotolerans as Noviherbaspirillum Soli Comb. Nov., Noviherbaspirillum aurantiacum Comb. Nov., Noviherbaspirillum canariense Comb. Nov. and Noviherbaspirillum psychrotolerans Comb. Nov. Based on Polyphasic Analysis. Int. J. Syst. Evol. Microbiol. 2013, 63, 4100–4107. [Google Scholar] [CrossRef]
- Ai, H.-X.; Che, Y.-C.; Wang, L.; Zhang, L.; Gu, Y.; Tan, Y.-N.; Chang, A.K.; Liu, H.-S. Paenibacillus liaoningensis sp. Nov., Isolated from Soil. Int. J. Syst. Evol. Microbiol. 2016, 66, 3150–3156. [Google Scholar] [CrossRef][Green Version]
- Kämpfer, P.; Irgang, R.; Fernández-Negrete, G.; Busse, H.-J.; Poblete-Morales, M.; Fuentes-Messina, D.; Glaeser, S.P.; Avendaño-Herrera, R. Proposal of Pedobacter nototheniae sp. Nov., Isolated from the Spleen of a Black Rock Cod (Notothenia Coriiceps, Richardson 1844) from the Chilean Antarctica. Antonie Van Leeuwenhoek 2019, 112, 1465–1475. [Google Scholar] [CrossRef] [PubMed]
- Hoang, V.-A.; Kim, Y.-J.; Nguyen, N.-L.; Min, J.-W.; Yang, D.-C. Pedobacter ginsengiterrae sp. nov., Isolated from Soil of a Ginseng Field. Int. J. Syst. Evol. Microbiol. 2013, 63, 1273–1279. [Google Scholar] [CrossRef]
- Hwang, C.Y.; Choi, D.H.; Cho, B.C. Pedobacter roseus sp. Nov., Isolated from a Hypertrophic Pond, and Emended Description of the Genus Pedobacter. Int. J. Syst. Evol. Microbiol. 2006, 56, 1831–1836. [Google Scholar] [CrossRef] [PubMed]
- Leibniz Institut DSMZ-Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH; Curators of the DSMZ; DSM 28024 Pseudarthrobacter Siccitolerans 4J27 is an Aerobe, Mesophilic, Gram-Positive Bacterium that was Isolated from Nerium Oleander Rhizosphere Subjected to Seasonal Drought. Available online: https://bacdive.dsmz.de/strain/24196 (accessed on 10 February 2023).
- Bozal, N.; Montes, M.J.; Mercadé, E. Pseudomonas guineae sp. nov., a Novel Psychrotolerant Bacterium from an Antarctic Environment. Int. J. Syst. Evol. Microbiol. 2007, 57, 2609–2612. [Google Scholar] [CrossRef]
- Reddy, G.S.N.; Matsumoto, G.I.; Schumann, P.; Stackebrandt, E.; Shivaji, S. Psychrophilic Pseudomonads from Antarctica: Pseudomonas antarctica sp. nov., Pseudomonas meridiana sp. nov. and Pseudomonas proteolytica sp. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 713–719. [Google Scholar] [CrossRef]
- Ramírez-Bahena, M.-H.; Cuesta, M.J.; Flores-Félix, J.D.; Mulas, R.; Rivas, R.; Castro-Pinto, J.; Brañas, J.; Mulas, D.; González-Andrés, F.; Velázquez, E.; et al. Pseudomonas helmanticensis sp. Nov., Isolated from Forest Soil. Int. J. Syst. Evol. Microbiol. 2014, 64, 2338–2345. [Google Scholar] [CrossRef]
- Korshunova, T.Y.; Ramírez-Bahena, M.H.; Chetverikov, S.P.; Igual, J.M.; Peix, Á.; Loginov, O. Pseudomonas turukhanskensis sp. nov., isolated from oil-contaminated soils. Int. J. Syst. Evol. Microbiol. 2016, 66, 4657–4664. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Pan, Y.; Wang, K.; Zhang, X.; Zhang, C.; Zhang, S.; Fu, X.; Jiang, J. Pseudomonas zhaodongensis sp. Nov., Isolated from Saline and Alkaline Soils. Int. J. Syst. Evol. Microbiol. 2015, 65, 1022–1030. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, U.; Schumann, P.; Meyer, J.-M.; Ulrich, A. Pseudomonas cedrina subsp. fulgida subsp. Nov., a Fluorescent Bacterium Isolated from the Phyllosphere of Grasses; Emended Description of Pseudomonas Cedrina and Description of Pseudomonas cedrina Subsp. cedrina Subsp. Nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 1331–1335. [Google Scholar] [CrossRef][Green Version]
- Romanenko, L.A.; Uchino, M.; Falsen, E.; Lysenko, A.M.; Zhukova, N.V.; Mikhailov, V.V. Pseudomonas xanthomarina sp. nov., a Novel Bacterium Isolated from Marine Ascidian. J. Gen. Appl. Microbiol. 2005, 51, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Furihata, K.; Ding, L.-X.; Yokota, A. Rhodococcus kyotonensis sp. Nov., a Novel Actinomycete Isolated from Soil. Int. J. Syst. Evol. Microbiol. 2007, 57, 1956–1959. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fernández-Silva, F.; Capilla, J.; Mayayo, E.; Sutton, D.; Guarro, J. In Vitro Evaluation of Antifungal Drug Combinations against Sarocladium (Acremonium) Kiliense, an Opportunistic Emergent Fungus Resistant to Antifungal Therapies. Antimicrob. Agents Chemother. 2014, 58, 1259–1260. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, H.-C.; Zhang, J.-L.; Zhou, Y.-G.; Xin, Y.-H. Sphingomonas psychrolutea sp. nov., a Psychrotolerant Bacterium Isolated from Glacier Ice. Int. J. Syst. Evol. Microbiol. 2015, 65, 2955–2959. [Google Scholar] [CrossRef]
- Takashima, M.; Nakase, T. Four New Species of the Genus Sporobolomyces Isolated from Leaves in Thailand. Mycoscience 2000, 41, 357–369. [Google Scholar] [CrossRef]
- Wolf, A.; Fritze, A.; Hagemann, M.; Berg, G. Stenotrophomonas rhizophila sp. nov., a Novel Plant-Associated Bacterium with Antifungal Properties. Int. J. Syst. Evol. Microbiol. 2002, 52, 1937–1944. [Google Scholar] [CrossRef]
- Michel, K.H.; Shah, R.M.; Hamill, R.L. A 35512, a Complex of New Antibacterial Antibiotics Produced by Streptomyces Candidus. I. Isolation and Characterization. J. Antibiot. 1980, 33, 1397–1406. [Google Scholar] [CrossRef]
- Cockell, C.S.; Kelly, L.C.; Marteinsson, V. Actinobacteria —An Ancient Phylum Active in Volcanic Rock Weathering. Geomicrobiol. J. 2013, 30, 706–720. [Google Scholar] [CrossRef]
- Kelly, L.C.; Cockell, C.S.; Herrera-Belaroussi, A.; Piceno, Y.; Andersen, G.; DeSantis, T.; Brodie, E.; Thorsteinsson, T.; Marteinsson, V.; Poly, F.; et al. Bacterial Diversity of Terrestrial Crystalline Volcanic Rocks, Iceland. Microb. Ecol. 2011, 62, 69–79. [Google Scholar] [CrossRef]
- Lysnes, K.; Thorseth, I.H.; Steinsbu, B.O.; Øvreås, L.; Torsvik, T.; Pedersen, R.B. Microbial Community Diversity in Seafloor Basalt from the Arctic Spreading Ridges. FEMS Microbiol. Ecol. 2004, 50, 213–230. [Google Scholar] [CrossRef] [PubMed]
- Bottos, E.M.; Woo, A.C.; Zawar-Reza, P.; Pointing, S.B.; Cary, S.C. Airborne Bacterial Populations Above Desert Soils of the McMurdo Dry Valleys, Antarctica. Microb. Ecol. 2014, 67, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Fahlgren, C.; Hagström, A.; Nilsson, D.; Zweifel, U.L. Annual Variations in the Diversity, Viability, and Origin of Airborne Bacteria. Appl. Environ. Microbiol. 2010, 76, 3015–3025. [Google Scholar] [CrossRef] [PubMed]
- Vaïtilingom, M.; Attard, E.; Gaiani, N.; Sancelme, M.; Deguillaume, L.; Flossmann, A.I.; Amato, P.; Delort, A.-M. Long-Term Features of Cloud Microbiology at the Puy de Dôme (France). Atmos. Environ. 2012, 56, 88–100. [Google Scholar] [CrossRef]
- Hirsch, P.; Gallikowski, C.A.; Siebert, J.; Peissl, K.; Kroppenstedt, R.; Schumann, P.; Stackebrandt, E.; Anderson, R. Deinococcus frigens sp. nov., Deinococcus saxicola sp. nov., and Deinococcus marmoris sp. Nov., Low Temperature and Draught-Tolerating, UV-Resistant Bacteria from Continental Antarctica. Syst. Appl. Microbiol. 2004, 27, 636–645. [Google Scholar] [CrossRef]
- Yoo, S.-H.; Weon, H.-Y.; Kim, S.-J.; Kim, Y.-S.; Kim, B.-Y.; Kwon, S.-W. Deinococcus aerolatus sp. nov. and Deinococcus aerophilus sp. nov., Isolated from Air Samples. Int. J. Syst. Evol. Microbiol. 2010, 60, 1191–1195. [Google Scholar] [CrossRef]
- Gerber, E.; Bernard, R.; Castang, S.; Chabot, N.; Coze, F.; Dreux-Zigha, A.; Hauser, E.; Hivin, P.; Joseph, P.; Lazarelli, C.; et al. Deinococcus as New Chassis for Industrial Biotechnology: Biology, Physiology and Tools. J. Appl. Microbiol. 2015, 119, 1–10. [Google Scholar] [CrossRef]
- Gajewska, J.; Floryszak-Wieczorek, J.; Sobieszczuk-Nowicka, E.; Mattoo, A.; Arasimowicz-Jelonek, M. Fungal and Oomycete Pathogens and Heavy Metals: An Inglorious Couple in the Environment. IMA Fungus 2022, 13, 6. [Google Scholar] [CrossRef] [PubMed]
- Rosa, L.H.; Pinto, O.H.B.; Šantl-Temkiv, T.; Convey, P.; Carvalho-Silva, M.; Rosa, C.A.; Câmara, P.E.A.S. DNA Metabarcoding of Fungal Diversity in Air and Snow of Livingston Island, South Shetland Islands, Antarctica. Sci. Rep. 2020, 10, 21793. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich-Nowoisky, J.; Pickersgill, D.A.; Després, V.R.; Pöschl, U. High Diversity of Fungi in Air Particulate Matter. Proc. Natl. Acad. Sci. USA 2009, 106, 12814–12819. [Google Scholar]
- Uetake, J.; Tobo, Y.; Uji, Y.; Hill, T.C.J.; DeMott, P.J.; Kreidenweis, S.M.; Misumi, R. Seasonal Changes of Airborne Bacterial Communities Over Tokyo and Influence of Local Meteorology. Front. Microbiol. 2019, 10, 1572. [Google Scholar] [CrossRef] [PubMed]
- Choe, Y.-H.; Kim, M.; Woo, J.; Lee, M.J.; Lee, J.I.; Lee, E.J.; Lee, Y.K. Comparing Rock-Inhabiting Microbial Communities in Different Rock Types from a High Arctic Polar Desert. FEMS Microbiol. Ecol. 2018, 94, fiy070. [Google Scholar] [CrossRef]
- Kosina, M.; Barták, M.; Mašlaňová, I.; Pascutti, A.V.; Šedo, O.; Lexa, M.; Sedláček, I. Pseudomonas prosekii sp. Nov., a Novel Psychrotrophic Bacterium from Antarctica. Curr. Microbiol. 2013, 67, 637–646. [Google Scholar] [CrossRef]
- Tan, Y.-S.; Zhang, R.-K.; Liu, Z.-H.; Li, B.-Z.; Yuan, Y.-J. Microbial Adaptation to Enhance Stress Tolerance. Front. Microbiol. 2022, 13, 888746. [Google Scholar] [CrossRef]
- Amato, P.; Besaury, L.; Joly, M.; Penaud, B.; Deguillaume, L.; Delort, A.-M. Metatranscriptomic Exploration of Microbial Functioning in Clouds. Sci. Rep. 2019, 9, 4383. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Station-Abbreviation | Coordinates | Elevation | Characteristics | Year | Air Samples | Rock Samples | |
|---|---|---|---|---|---|---|---|---|
| 1-Year-Old | 9-Year-Old | |||||||
| Surtsey | Mávavarp-M | 63°17.880′ N 20°36.092′ W | 50 m | Gull nesting area-vegetation | 2018 | 2 | 0 | 0 |
| 2019 | 2 | 2 | 0 | |||||
| Borhola-B | 63°18.097′ N 20°36.010′ W | 66 m | Boreholes-Human activity | 2018 | 2 | 0 | 0 | |
| 2019 | 2 | 2 | 0 | |||||
| Viti-V | 63°18.244′ N 20°36.062′ W | 166 m | Lighthouse–Highest point | 2018 | 2 | 0 | 0 | |
| 2019 | 2 | 2 | 0 | |||||
| Tangi-T | 63°18.455′ N 20°36.011′ W | 11 m | Peninsula–Rocks | 2018 | 2 | 0 | 0 | |
| 2019 | 2 | 0 | 0 | |||||
| Pálsbær-P | 63°18.047′ N 20°35.828′ W | 38 m | Human hut | 2018 | 0 | 0 | 0 | |
| 2019 | 2 | 2 | 0 | |||||
| Austur-A | 63°18.153′ N 20°36.609′ W | 108 m | Crater | 2018 | 0 | 0 | 0 | |
| 2019 | 0 | 2 | 0 | |||||
| Fimmvörðuháls | Top Eyjafja-1 | 63°37.985′ N 19°26.466′ W | 1036 m | Crater | 2018 | 2 | 0 | 0 |
| 2019 | 0 | 2 | 1 | |||||
| Lava Eyja-2 | 63°38.182′ N 19°25.991′ W | 1030 m | Lava field | 2018 | 2 | 0 | 0 | |
| 2019 | 0 | 2 | 0 | |||||
| Hut-3 | 63°36.654′ N 19°26.489′ W | 875 m | Human hut | 2018 | 2 | 0 | 0 | |
| 2019 | 0 | 2 | 1 | |||||
| Strain Name | Closest Match on BLAST | Survival Properties | ||||
|---|---|---|---|---|---|---|
| Genus | Species | Pigment Production | Cold-Adaptation | Other Properties | Reference | |
| 19SA31 | Albirhodobacter | confluentis | - | - | - | - |
| 19FR6 | Arthrobacter | livingstonensis | - | + | - | [48] |
| 19FR + 13 | Arthrobacter | koreensis | +, yellow | - | - | [49] |
| 18FA27 | Bacillus | cereus | - | - | - | - |
| 19FR + 12 | Brachybacterium | rhamnosum | - | - | - | - |
| 19FR4 | Brachybacterium | nesterenkovii | +, yellow | - | - | [50] |
| 18SA17 | Brevibacterium | antiquum | +, orange | - | - | [51] |
| 18FA7 | Caballeronia | udeis | - | - | - | - |
| 19SA8 | Comamonas | piscis | +, yellow | - | - | [52] |
| 19FR17 | Deinococcus | taklimakanensis | - | - | Gamma UV tolerant | [53] |
| 19SR23 | Dermacoccus | nishinomiyaensis | - | - | - | - |
| 19SA79 | Erwinia | billingiae | - | - | - | - |
| 19SA17 | Flavobacterium | frigidimaris | +, yellow | + | - | [54] |
| 19SA32 | Janthinobacterium | rivuli | +, purple | - | - | [55] |
| 19SA80 | Lysinimonas | soli | - | - | - | - |
| 19FR27 | Massilia | glaciei | - | - | - | - |
| 19SR22 | Massilia | brevitalea | +, yellow | - | - | [56] |
| 19FR22 | Massilia | eurypsychrophila | - | + | - | [57] |
| 19FR + 1 | Massilia | psychrophila | +, yellow | + | - | [58] |
| 19SR39 | Massilia | niabensis | +, yellow | - | - | [59] |
| 18SA14 | Microbacterium | maritypicum | +, yellow | - | - | [60] |
| 19FR32 | Micrococcus | aloeverae | +, yellow | - | - | [61] |
| 19FR42 | Micrococcus | flavus | +, yellow | - | - | [62] |
| 19FR13 | Moraxella | osloensis | +, purple | - | - | [63] |
| 19FR43 | Noviherbaspirilum | psychrotolerans | - | + | - | [64] |
| 19SR16 | Paenarthrobacter | nicotinovorans | - | - | - | - |
| 19SR10 | Paenarthrobacter | nitroguajacolicus | - | - | - | - |
| 19SR6 | Paenibacillus | liaoningensis | - | - | Endospore-forming | [65] |
| 19SR5 | Paenibacillus | algorifonticola | - | - | - | - |
| 19SA40 | Paracoccus | aquimaris | - | - | - | - |
| 18FA17 | Paracoccus | Sanguinis * | - | - | - | - |
| 18FA24 | Paracoccus | Yeei * | - | - | - | - |
| 19FR5 | Paracoccus | marinus | +, orange | - | - | [47] |
| 19SR31 | Pedobacter | ginsenosidimutans | - | - | - | - |
| 18SA31 | Pedobacter | nototheniae | +, pink | - | - | [66] |
| 19SA47 | Pedobacter | ginsengiterrae | +, pink | - | - | [67] |
| 18SA30 | Pedobacter | roseus | +, pink | - | - | [68] |
| 19SR20 | Pseudarthrobacter | sulfonivorans | - | - | - | - |
| 19SR33 | Pseudarthrobacter | siccitolerans | +, beige | - | xeroprotectant | [69] |
| 19SA35 | Pseudomonas | baetica | - | - | - | - |
| 19SA54 | Pseudomonas | guineae | - | + | - | [70] |
| 18SA13 | Pseudomonas | graminis | - | - | - | - |
| 18SA5 | Pseudomonas | marginalis | - | - | - | - |
| 19SA55 | Pseudomonas | plecoglossicida | - | - | - | - |
| 19SA52 | Pseudomonas | proteolytica | - | + | - | [71] |
| 19SA49 | Pseudomonas | trivialis | - | - | - | - |
| 18FA37 | Pseudomonas | caspiana | - | - | - | - |
| 19SA57 | Pseudomonas | helmanticensis | +, beige | - | - | [72] |
| 18SA9 | Pseudomonas | turukhanskensis | +, beige | - | - | [73] |
| 19SA5 | Pseudomonas | zhaodongensis | +, orange | - | - | [74] |
| 19SA71 | Pseudomonas | cedrina | +, yellow | - | - | [75] |
| 19SA12 | Pseudomonas | xanthomarina | +, yellow | - | - | [76] |
| 19SA2 | Psychrobacter | namhaensis | - | - | - | - |
| 18FA12 | Rhodococcus | kyotonensis | +, orange | - | - | [77] |
| 18FA15 | Saraclodium | kiliense | - | - | resistant to anti-fungal | [78] |
| 19SA65 | Serratia | proteamaculans | - | - | - | - |
| 19SA78 | Shewanella | hafniensis | - | - | - | - |
| 18SA19 | Sphingomonas | glacialis | +, yellow | + | - | [43] |
| 19FR + 6 | Sphingomonas | psychrolutea | +, orange | + | - | [79] |
| 18SA34F | Sporobolomyces | blumeae | +, orange | - | freeze–thaw tolerant | [80] |
| 19FR11 | Staphylococcus | Epidermidis * | - | - | - | - |
| 19FR30 | Staphylococcus | Warneri * | - | - | - | - |
| 19SA19 | Stenotrophomonas | rhizophila | - | - | Osmoprotective substances production and antifungal activity | [81] |
| 18SA18 | Stenotrophomonas | chelatiphaga | - | - | - | - |
| 19FR2 | Streptococcus | Mitis * | - | - | - | - |
| 19FR67 | Streptococcus | Sanguinis * | - | - | - | - |
| 18FA10 | Streptomyces | candidus | - | - | Produce antibiotics | [82] |
| 19SA39 | Variovorax | boronicumulans | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daussin, A.; Vannier, P.; Ménager, M.; Daboussy, L.; Šantl-Temkiv, T.; Cockell, C.; Marteinsson, V.Þ. Comparison of Atmospheric and Lithospheric Culturable Bacterial Communities from Two Dissimilar Active Volcanic Sites, Surtsey Island and Fimmvörðuháls Mountain in Iceland. Microorganisms 2023, 11, 665. https://doi.org/10.3390/microorganisms11030665
Daussin A, Vannier P, Ménager M, Daboussy L, Šantl-Temkiv T, Cockell C, Marteinsson VÞ. Comparison of Atmospheric and Lithospheric Culturable Bacterial Communities from Two Dissimilar Active Volcanic Sites, Surtsey Island and Fimmvörðuháls Mountain in Iceland. Microorganisms. 2023; 11(3):665. https://doi.org/10.3390/microorganisms11030665
Chicago/Turabian StyleDaussin, Aurélien, Pauline Vannier, Marine Ménager, Lola Daboussy, Tina Šantl-Temkiv, Charles Cockell, and Viggó Þór Marteinsson. 2023. "Comparison of Atmospheric and Lithospheric Culturable Bacterial Communities from Two Dissimilar Active Volcanic Sites, Surtsey Island and Fimmvörðuháls Mountain in Iceland" Microorganisms 11, no. 3: 665. https://doi.org/10.3390/microorganisms11030665
APA StyleDaussin, A., Vannier, P., Ménager, M., Daboussy, L., Šantl-Temkiv, T., Cockell, C., & Marteinsson, V. Þ. (2023). Comparison of Atmospheric and Lithospheric Culturable Bacterial Communities from Two Dissimilar Active Volcanic Sites, Surtsey Island and Fimmvörðuháls Mountain in Iceland. Microorganisms, 11(3), 665. https://doi.org/10.3390/microorganisms11030665