


Partial Substitution of Urea with Biochar Induced Improvements in Soil Enzymes Activity, Ammonia-Nitrite Oxidizers, and Nitrogen Uptake in the Double-Cropping Rice System
 ,
,  , ,
, ,  ,
,  , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site and Crop Management
2.2. Soil and Plant Sampling
2.3. Determination of Soil Chemical Properties and Plant N Concentrations
2.4. Soil Enzyme Measurements
2.5. DNA Extraction and Quantitative PCR Analysis
2.6. Measurement of Dry Matter Accumulation and Grain Yield
2.7. Statistical Analysis
3. Results
3.1. Soil Chemical Properties
3.2. Soil Enzymes Activities
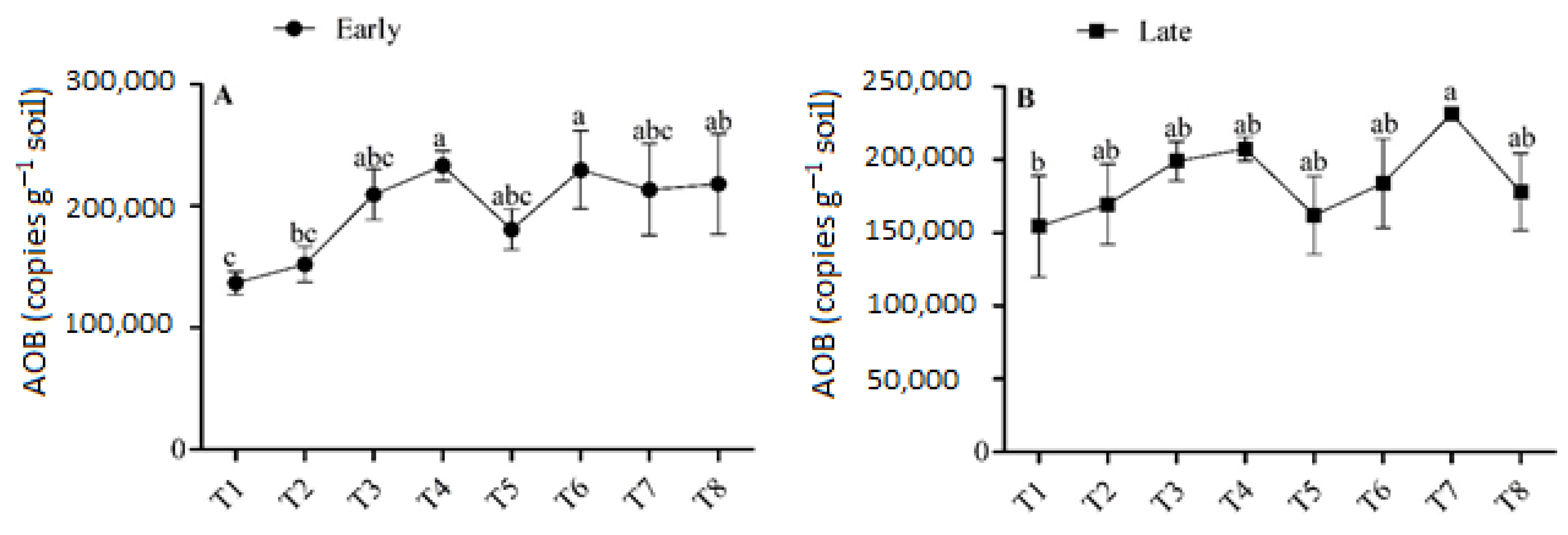
3.3. Abundance of AOB
3.4. Abundance of AOA
3.5. Abundance of NOB
3.6. N Concentration in Stems, Leaves, and Panicles
3.7. DM Production and Grain Yield
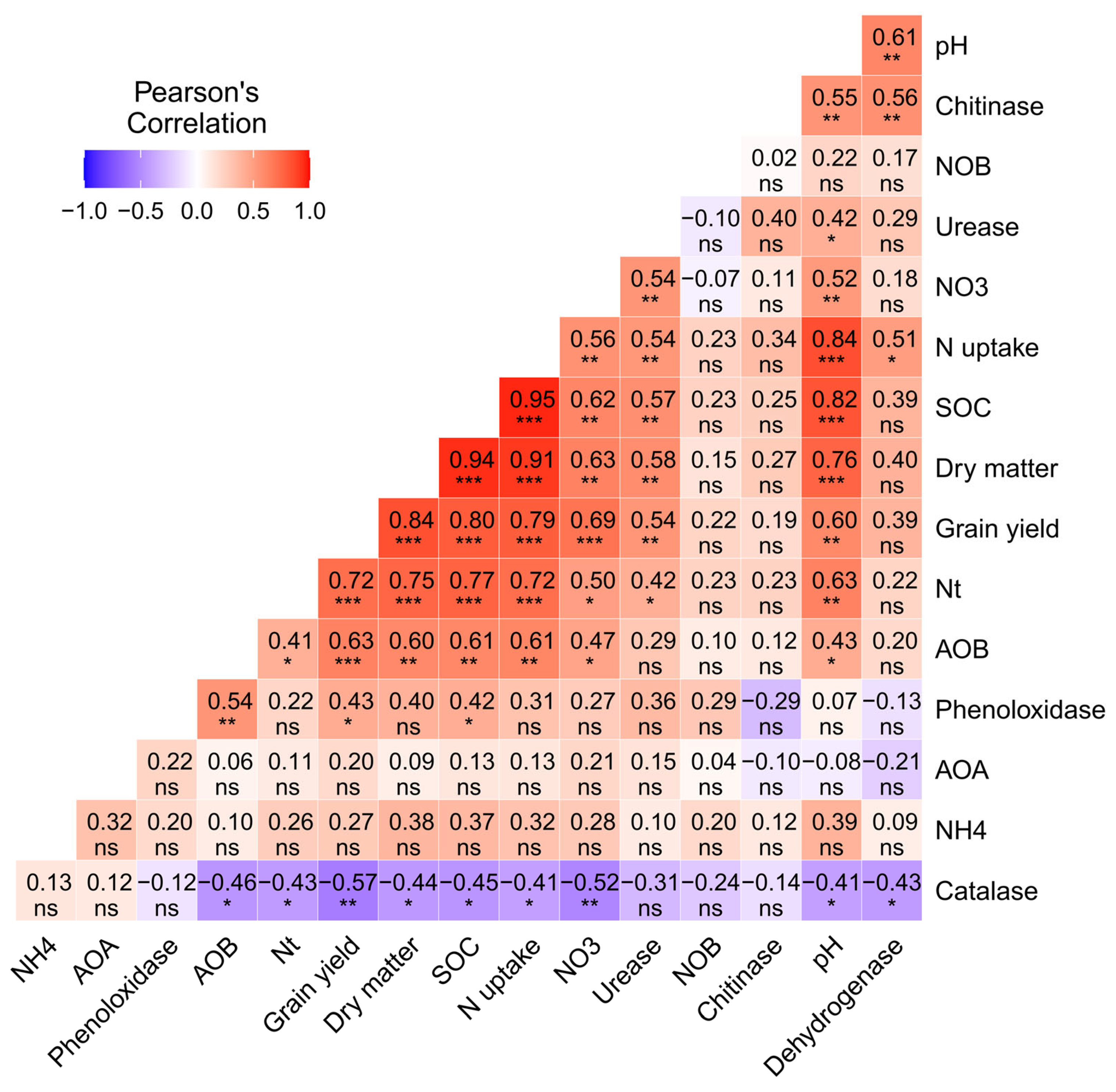
3.8. Correlation Heat Map Analysis
4. Discussion
4.1. Effect of Urea and Biochar Application on Soil Chemical Properties
4.2. Effect of Urea and Biochar Application on Soil Enzyme Activities
4.3. Effect of Urea and Biochar Application on the Abundances of AOB, AOB, and NOB
4.4. Effect of Urea and Biochar Application on Plant N Uptake, DM, and Grain Yield
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization of the United NationsStatistic Division. 2021. Available online: https://reliefweb.int/report/world/fao-statistical-yearbook-2021-world-food-and-agriculture (accessed on 8 November 2021).
- Ullah, S.; Ali, I.; Liang, H.; Zhao, Q.; Wei, S.; Muhammad, I.; Huang, M.; Ali, N.; Jiang, L.J.G.B. An approach to sustainable agriculture by untangling the fate of contrasting nitrogen sources in double-season rice grown with and without biochar. GCB Bioenergy 2021, 13, 382–392. [Google Scholar] [CrossRef]
- Nie, L.; Peng, S. Rice production in China. In Rice Production Worldwide; Springer: Cham, Switzerland, 2017; pp. 33–52. [Google Scholar]
- Frolking, S.; Qiu, J.; Boles, S.; Xiao, X.; Liu, J.; Zhuang, Y.; Li, C.; Qin, X.J.G.B.C. Combining remote sensing and ground census data to develop new maps of the distribution of rice agriculture in China. Glob. Biogeochem. Cycles 2002, 16, 38-1–38-10. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, J.; Zhang, W.; Cui, Z.; Ma, W.; Chen, X.; Jiang, R.J.A.P.S. Nutrient use efficiencies of major cereal crops in China and measures for improvement. Acta Pedol. Sin. 2008, 45, 915–924. [Google Scholar]
- Ladha, J.K.; Pathak, H.; Krupnik, T.J.; Six, J.; van Kessel, C. Efficiency of Fertilizer Nitrogen in Cereal Production: Retrospects and Prospects. Adv. Agron. 2005, 87, 85–156. [Google Scholar] [CrossRef]
- Chien, S.; Prochnow, L.; Cantarella, H. Chapter 8 Recent Developments of Fertilizer Production and Use to Improve Nutrient Efficiency and Minimize Environmental Impacts. Adv. Agron. 2009, 102, 267–322. [Google Scholar] [CrossRef]
- Ameloot, N.; De Neve, S.; Jegajeevagan, K.; Yildiz, G.; Buchan, D.; Funkuin, Y.N.; Prins, W.; Bouckaert, L.; Sleutel, S. Short-term CO2 and N2O emissions and microbial properties of biochar amended sandy loam soils. Soil Biol. Biochem. 2013, 57, 401–410. [Google Scholar] [CrossRef]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D.J.S.B. Biochar effects on soil biota—A review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Sun, H.; Yi, Z.; Jeyakumar, P.; Xia, C.; Feng, Y.; Lam, S.S.; Shi, W.; Environmental Pollution. Citric acid modified biochar ap-plication at a low dosage can synchronically mitigate the nitrogenous gas pollutants emission from rice paddy soils. Environ. Pollut. 2022, 312, 120068. [Google Scholar] [CrossRef]
- Atkinson, C.J.; Fitzgerald, J.D.; Hipps, N.A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: A review. Plant Soil 2010, 337, 1–18. [Google Scholar] [CrossRef]
- Xu, H.-J.; Wang, X.-H.; Li, H.; Yao, H.-Y.; Su, J.-Q.; Zhu, Y.-G. Biochar Impacts Soil Microbial Community Composition and Nitrogen Cycling in an Acidic Soil Planted with Rape. Environ. Sci. Technol. 2014, 48, 9391–9399. [Google Scholar] [CrossRef]
- Pan, F.; Chapman, S.J.; Li, Y.; Yao, H. Straw amendment to paddy soil stimulates denitrification but biochar amendment promotes anaerobic ammonia oxidation. J. Soils Sediments 2017, 17, 2428–2437. [Google Scholar] [CrossRef]
- Chen, S.; Li, D.; He, H.; Zhang, Q.; Lu, H.; Xue, L.; Feng, Y.; Sun, H. Substituting urea with biogas slurry and hydrothermal carbonization aqueous product could decrease NH3 volatilization and increase soil DOM in wheat growth cycle. Environ. Res. 2022, 214, 113997. [Google Scholar] [CrossRef]
- Velmourougane, K.; Venugopalan, M.; Bhattacharyya, T.; Sarkar, D.; Pal, D.; Sahu, A.; Ray, S.; Nair, K.; Prasad, J.; Singh, R. Soil dehydrogenase activity in agro-ecological sub regions of black soil regions in India. Geoderma 2013, 197, 186–192. [Google Scholar] [CrossRef]
- García-Gil, J.; Plaza, C.; Soler-Rovira, P.; Polo, A. Long-term effects of municipal solid waste compost application on soil enzyme activities and microbial biomass. Soil Biol. Biochem. 2000, 32, 1907–1913. [Google Scholar] [CrossRef]
- Henry, H.A.J.S.B. Reprint of “Soil extracellular enzyme dynamics in a changing climate”. Biochemistry 2013, 56, 53–59. [Google Scholar] [CrossRef]
- Sardar, K.; Qing, C.; Hesham, A.E.-L.; Yue, X.; He, J. Soil enzymatic activities and microbial community structure with different application rates of Cd and Pb. J. Environ. Sci. 2007, 19, 834–840. [Google Scholar]
- Zhu, X.; Chen, B.; Zhu, L.; Xing, B. Effects and mechanisms of biochar-microbe interactions in soil improvement and pollution remediation: A review. Environ. Pollut. 2017, 227, 98–115. [Google Scholar] [CrossRef]
- Kowalchuk, G.A.; Stephen, J.R.J.A.r.o.m. Ammonia-oxidizing bacteria: A model for molecular microbial ecology. Annu. Rev. Microbiol. 2001, 55, 485. [Google Scholar] [CrossRef]
- Ward, B.; Arp, D.; Klotz, M.J.D. Nitrification; American Society for Microbiology: Washington, D.C., USA, 2011. [Google Scholar]
- Li, K.; Qian, J.; Wang, P.; Wang, C.; Lu, B.; Tian, X.; Jin, W.; He, X.; Chen, H.; Zhang, Y. Differential responses of encoding-amoA nitrifiers and nir denitrifiers in activated sludge to anatase and rutile TiO2 nanoparticles: What is active functional guild in rate limiting step of nitrogen cycle? J. Hazard. Mater. 2020, 384, 121388. [Google Scholar] [CrossRef]
- Xu, N.; Tan, G.; Wang, H.; Gai, X. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, H.; Zhou, S.; Bai, N.; Zheng, X.; Li, S.; Zhang, J.; Lv, W. Effect of Straw and Straw Biochar on the Community Structure and Diversity of Ammonia-oxidizing Bacteria and Archaea in Rice-wheat Rotation Ecosystems. Sci. Rep. 2019, 9, 9367. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agricultural Chemical Analysis; China Agricultural Press: Beijing, China, 2000; pp. 265–267. [Google Scholar]
- Maynard, D.G.; Kalra, Y.P.; Crumbaugh, J.A. Nitrate and exchangeable ammonium nitrogen. In Soil Sampling and Methods of Analysis, 2nd ed.; Carter, M.R., Gregorich, E.G., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 71–80. [Google Scholar]
- Chen, Y.; Liu, J.; Liu, S.J.P. Effect of long-term mineral fertilizer application on soil enzyme activities and bacterial community composition. Plant Soil Environ. 2018, 64, 571–577. [Google Scholar] [CrossRef]
- Guan, S.Y.; Zhang, D.; Zhang, Z. Soil Enzyme and Its Research Methods; Chinese Agricultural Press: Beijing, China, 1986; pp. 274–297. [Google Scholar]
- Wang, Y.; Cheng, S.; Fang, H.; Yu, G.; Yang, X.; Xu, M.; Dang, X.; Li, L.; Wang, L. Relationships between ammonia-oxidizing communities, soil methane uptake and nitrous oxide fluxes in a subtropical plantation soil with nitrogen enrichment. Eur. J. Soil Biol. 2016, 73, 84–92. [Google Scholar] [CrossRef]
- Akhtar, K.; Wang, W.; Khan, A.; Ren, G.; Afridi, M.Z.; Feng, Y.; Yang, G. Wheat straw mulching offset soil moisture deficient for improving physiological and growth performance of summer sown soybean. Agric. Water Manag. 2018, 211, 16–25. [Google Scholar] [CrossRef]
- Reis, S.; Silva, M.; Castro, N.; Correia, V.; Gutierrez, J.; Lasheras, A.; Lanceros-Mendez, S.; Martins, P.J.S.M. Optimized anisotropic magnetoelectric response of Fe61. 6Co16. 4Si10. 8B11. 2/PVDF/Fe61. 6Co16. 4Si10. 8B11. 2 laminates for AC/DC magnetic field sensing. Smart Mater. Struct. 2016, 25, 055050. [Google Scholar] [CrossRef]
- Jaafar, N.M.; Clode, P.L.; Abbott, L.K. Soil Microbial Responses to Biochars Varying in Particle Size, Surface and Pore Properties. Pedosphere 2015, 25, 770–780. [Google Scholar] [CrossRef]
- Ali, I.; He, L.; Ullah, S.; Quan, Z.; Wei, S.; Iqbal, A.; Munsif, F.; Shah, T.; Xuan, Y.; Luo, Y.; et al. Biochar addition coupled with nitrogen fertilization impacts on soil quality, crop productivity, and nitrogen uptake under double-cropping system. Food Energy Secur. 2020, 9, e208. [Google Scholar] [CrossRef]
- Ullah, S.; Zhao, Q.; Wu, K.; Ali, I.; Liang, H.; Iqbal, A.; Wei, S.; Cheng, F.; Ahmad, S.; Jiang, L.; et al. Biochar application to rice with 15N-labelled fertilizers, enhanced leaf nitrogen concentration and assimilation by improving morpho-physiological traits and soil quality. Saudi J. Biol. Sci. 2021, 28, 3399–3413. [Google Scholar] [CrossRef]
- Biederman, L.A.; Harpole, W.S. Biochar and its effects on plant productivity and nutrient cycling: A meta-analysis. GCB Bioenergy 2013, 5, 202–214. [Google Scholar] [CrossRef]
- Ju, X.-T.; Xing, G.-X.; Chen, X.-P.; Zhang, S.-L.; Zhang, L.-J.; Liu, X.-J.; Cui, Z.-L.; Yin, B.; Christie, P.; Zhu, Z.-L.; et al. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 2009, 106, 3041–3046. [Google Scholar] [CrossRef]
- Rashid, M.; Hussain, Q.; Khan, K.S.; Al-Wabel, M.I.; Afeng, Z.; Akmal, M.; Ijaz, S.S.; Aziz, R.; Shah, G.A.; Mehdi, S.M. Prospects of biochar in alkaline soils to mitigate climate change. In Environment, Climate, Plant and Vegetation Growth; Springer: Berlin/Heidelberg, Germany, 2020; pp. 133–149. [Google Scholar]
- Nanda, S.; Dalai, A.K.; Berruti, F.; Kozinski, J.A. Biochar as an Exceptional Bioresource for Energy, Agronomy, Carbon Sequestration, Activated Carbon and Specialty Materials. Waste Biomass-Valorization 2015, 7, 201–235. [Google Scholar] [CrossRef]
- Lima, J.R.D.S.; Silva, W.D.M.; de Medeiros, E.V.; Duda, G.P.; Corrêa, M.M.; Filho, A.P.M.; Clermont-Dauphin, C.; Antonino, A.C.D.; Hammecker, C. Effect of biochar on physicochemical properties of a sandy soil and maize growth in a greenhouse experiment. Geoderma 2018, 319, 14–23. [Google Scholar] [CrossRef]
- Jian, H.; Qing-Zhong, Z.; Zhang-Liu, D.; Yi-Ding, W. Research advances in effects of biochar application on agroecosystem. Chin. J. Agrometeorol. 2012, 33, 232. [Google Scholar]
- Wang, X.; Song, D.; Liang, G.; Zhang, Q.; Ai, C.; Zhou, W. Maize biochar addition rate influences soil enzyme activity and microbial community composition in a fluvo-aquic soil. Appl. Soil Ecol. 2015, 96, 265–272. [Google Scholar] [CrossRef]
- Bailey, V.L.; Fansler, S.J.; Smith, J.L.; Bolton, H., Jr. Reconciling apparent variability in effects of biochar amendment on soil enzyme activities by assay optimization. Soil Biol. Biochem. 2011, 43, 296–301. [Google Scholar] [CrossRef]
- Zhang, X.; Dong, W.; Dai, X.; Schaeffer, S.; Yang, F.; Radosevich, M.; Xu, L.; Liu, X.; Sun, X. Responses of absolute and specific soil enzyme activities to long term additions of organic and mineral fertilizer. Sci. Total Environ. 2015, 536, 59–67. [Google Scholar] [CrossRef]
- Zhao, Q.; Tang, J.; Li, Z.; Yang, W.; Duan, Y. The Influence of Soil Physico-Chemical Properties and Enzyme Activities on Soil Quality of Saline-Alkali Agroecosystems in Western Jilin Province, China. Sustainability 2018, 10, 1529. [Google Scholar] [CrossRef]
- Foster, E.J.; Hansen, N.; Wallenstein, M.; Cotrufo, M.F. Biochar and manure amendments impact soil nutrients and microbial enzymatic activities in a semi-arid irrigated maize cropping system. Agric. Ecosyst. Environ. 2016, 233, 404–414. [Google Scholar] [CrossRef]
- Graber, E.R.; Harel, Y.M.; Kolton, M.; Cytryn, E.; Silber, A.; David, D.R.; Tsechansky, L.; Borenshtein, M.; Elad, Y. Biochar impact on development and productivity of pepper and tomato grown in fertigated soilless media. Plant Soil 2010, 337, 481–496. [Google Scholar] [CrossRef]
- Buchkowski, R.W.; Shaw, A.N.; Sihi, D.; Smith, G.R.; Keiser, A.D. Constraining Carbon and Nutrient Flows in Soil with Ecological Stoichiometry. Front. Ecol. Evol. 2019, 7, 382. [Google Scholar] [CrossRef]
- Jia, Z.; Conrad, R. Bacteria rather than Archaea dominate microbial ammonia oxidation in an agricultural soil. Environ. Microbiol. 2009, 11, 1658–1671. [Google Scholar] [CrossRef]
- Shen, J.-P.; Zhang, L.-M.; Zhu, Y.-G.; Zhang, J.-B.; He, J.-Z. Abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea communities of an alkaline sandy loam. Environ. Microbiol. 2008, 10, 1601–1611. [Google Scholar] [CrossRef]
- Prommer, J.; Wanek, W.; Hofhansl, F.; Trojan, D.; Offre, P.; Urich, T.; Schleper, C.; Sassmann, S.; Kitzler, B.; Soja, G.; et al. Biochar Decelerates Soil Organic Nitrogen Cycling but Stimulates Soil Nitrification in a Temperate Arable Field Trial. PLoS ONE 2014, 9, e86388. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, X.; Ma, B.; Chang, S.X.; Gong, J. Biochar addition affected the dynamics of ammonia oxidizers and nitrification in microcosms of a coastal alkaline soil. Biol. Fertil. Soils 2013, 50, 321–332. [Google Scholar] [CrossRef]
- Steinbeiss, S.; Gleixner, G.; Antonietti, M. Effect of biochar amendment on soil carbon balance and soil microbial activity. Soil Biol. Biochem. 2009, 41, 1301–1310. [Google Scholar] [CrossRef]
- Ball, P.N.; MacKenzie, M.D.; DeLuca, T.H.; Montana, W.E.H. Wildfire and Charcoal Enhance Nitrification and Ammonium-Oxidizing Bacterial Abundance in Dry Montane Forest Soils. J. Environ. Qual. 2010, 39, 1243–1253. [Google Scholar] [CrossRef]
- DeLuca, T.H.; MacKenzie, M.D.; Gundale, M.J.; Holben, W.E. Wildfire-Produced Charcoal Directly Influences Nitrogen Cycling in Ponderosa Pine Forests. Soil Sci. Soc. Am. J. 2006, 70, 448–453. [Google Scholar] [CrossRef]
- Hou, Q.; Zuo, T.; Wang, J.; Huang, S.; Wang, X.; Yao, L.; Ni, W. Responses of nitrification and bacterial community in three size aggregates of paddy soil to both of initial fertility and biochar addition. Appl. Soil Ecol. 2021, 166, 104004. [Google Scholar] [CrossRef]
- Spokas, K.A.; Reicosky, D.C. Impacts of Sixteen Different Biochars on Soil Greenhouse Gas Production. 2009. Available online: https://pubag.nal.usda.gov/catalog/47667 (accessed on 22 July 2022).
- Berglund, L.; DeLuca, T.; Zackrisson, O. Activated carbon amendments to soil alters nitrification rates in Scots pine forests. Soil Biol. Biochem. 2004, 36, 2067–2073. [Google Scholar] [CrossRef]
- Fan, F.; Yang, Q.; Li, Z.; Wei, D.; Cui, X.; Liang, Y. Impacts of Organic and Inorganic Fertilizers on Nitrification in a Cold Climate Soil are Linked to the Bacterial Ammonia Oxidizer Community. Microb. Ecol. 2011, 62, 982–990. [Google Scholar] [CrossRef]
- Xia, W.; Zhang, C.; Zeng, X.; Feng, Y.; Weng, J.; Lin, X.; Zhu, J.; Xiong, Z.; Xu, J.; Cai, Z.; et al. Autotrophic growth of nitrifying community in an agricultural soil. ISME J. 2011, 5, 1226–1236. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, C.; Lin, C.; Zhang, Y.; Chen, X.; Tang, L.; Liu, C.; Chen, Q.; Onwuka, M.I.; Song, T. Methane and Nitrous Oxide Flux after Biochar Application in Subtropical Acidic Paddy Soils under Tobacco-Rice Rotation. Sci. Rep. 2019, 9, 17277. [Google Scholar] [CrossRef]
- Bruun, S.; EL-Zehery, T.J. Biochar effect on the mineralization of soil organic matter. Pesqui. Agropecuária Bras. 2012, 47, 665–671. [Google Scholar] [CrossRef]
- Ali, I.; Ullah, S.; He, L.; Zhao, Q.; Iqbal, A.; Wei, S.; Shah, T.; Ali, N.; Bo, Y.; Adnan, M.; et al. Combined application of biochar and nitrogen fertilizer improves rice yield, microbial activity and N-metabolism in a pot experiment. Peerj 2020, 8, e10311. [Google Scholar] [CrossRef]
- Huang, M.; Yang, L.; Qin, H.; Jiang, L.; Zou, Y. Quantifying the effect of biochar amendment on soil quality and crop productivity in Chinese rice paddies. Field Crop. Res. 2013, 154, 172–177. [Google Scholar] [CrossRef]
- Ali, I.; Zhao, Q.; Wu, K.; Ullah, S.; Iqbal, A.; Liang, H.; Zhang, J.; Muhammad, I.; Amanullah; Khan, A.; et al. Biochar in Combination with Nitrogen Fertilizer is a Technique: To Enhance Physiological and Morphological Traits of Rice (Oryza sativa L.) by Improving Soil Physio-biochemical Properties. J. Plant Growth Regul. 2021, 41, 2406–2420. [Google Scholar] [CrossRef]
- Purakayastha, T.; Bera, T.; Bhaduri, D.; Sarkar, B.; Mandal, S.; Wade, P.; Kumari, S.; Biswas, S.; Menon, M.; Pathak, H.; et al. A review on biochar modulated soil condition improvements and nutrient dynamics concerning crop yields: Pathways to climate change mitigation and global food security. Chemosphere 2019, 227, 345–365. [Google Scholar] [CrossRef]
- Zahoor; Sun, D.; Li, Y.; Wang, J.; Tu, Y.; Wang, Y.; Hu, Z.; Zhou, S.; Wang, L.; Xie, G.; et al. Biomass saccharification is largely enhanced by altering wall polymer features and reducing silicon accumulation in rice cultivars harvested from nitrogen fertilizer supply. Bioresour. Technol. 2017, 243, 957–965. [Google Scholar] [CrossRef]
- Iqbal, A.; He, L.; Khan, A.; Wei, S.; Akhtar, K.; Ali, I.; Ullah, S.; Munsif, F.; Zhao, Q.; Jiang, L. Organic Manure Coupled with Inorganic Fertilizer: An Approach for the Sustainable Production of Rice by Improving Soil Properties and Nitrogen Use Efficiency. Agronomy 2019, 9, 651. [Google Scholar] [CrossRef]
- He, L.; Bi, Y.; Zhao, J.; Pittelkow, C.; Zhao, X.; Wang, S.; Xing, G. Population and community structure shifts of ammonia oxidizers after four-year successive biochar application to agricultural acidic and alkaline soils. Sci. Total Environ. 2018, 619, 1105–1115. [Google Scholar] [CrossRef]
- Plaimart, J.; Acharya, K.; Mrozik, W.; Davenport, R.J.; Vinitnantharat, S.; Werner, D. Coconut husk biochar amendment enhances nutrient retention by suppressing nitrification in agricultural soil following anaerobic digestate application. Environ. Pollut. 2020, 268, 115684. [Google Scholar] [CrossRef]
- Rotthauwe, J.H.; Witzel, K.P.; Liesack, W. The ammonia monooxygenase structural gene amoA as a functional marker: Molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl. Environ. Microbiol. 1997, 63, 4704–4712. [Google Scholar] [CrossRef]
- Nicol, G.W.; Leininger, S.; Schleper, C.; Prosser, J.I. The influence of soil pH on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea and bacteria. Environ. Microbiol. 2008, 10, 2966–2978. [Google Scholar] [CrossRef]
- Pester, M.; Maixner, F.; Berry, D.; Rattei, T.; Koch, H.; Lücker, S.; Nowka, B.; Richter, A.; Spieck, E.; Lebedeva, E.; et al. NxrB encoding the beta subunit of nitrite oxidoreductase as functional and phylogenetic marker for nitrite-oxidizing Nitrospira. Environ. Microbiol. 2014, 16, 3055–3071. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Urea (kg ha−1) | Biochar (t ha−1) | Biochar (kg plot−1) | Urea Basal (g plot−1) | Urea Tillering (g plot −1) | Urea Panicle Initiation (g plot−1) |
|---|---|---|---|---|---|---|
| (T1) N1B0 | 135 | 0 | 0 | 343.36 | 206.02 | 137.34 |
| (T2) N1B1 | 135 | 10 | 23.4 | 343.36 | 206.02 | 137.34 |
| (T3) N1B2 | 135 | 20 | 46.8 | 343.36 | 206.02 | 137.34 |
| (T4) N1B3 | 135 | 30 | 70.2 | 343.36 | 206.02 | 137.34 |
| (T5) N2B0 | 180 | 0 | 0 | 457.82 | 274.69 | 183.12 |
| (T6) N2B1 | 180 | 10 | 23.4 | 457.82 | 274.69 | 183.12 |
| (T7) N2B2 | 180 | 20 | 46.8 | 457.82 | 274.69 | 183.12 |
| (T8) N2B3 | 180 | 30 | 70.2 | 457.82 | 274.69 | 183.12 |
| Treatments | pH | Water % | SOC (g kg−1) | TN (g kg−1) | NH4+-N (mg kg−1) | NO3–-N (mg kg−1) |
|---|---|---|---|---|---|---|
| Early season | ||||||
| T1 | 6.02 ± 0.04 d | 32.17 ± 2.34 a | 9.90 ± 0.07 e | 1.19 ± 0.04 e | 25.95 ± 0.89 ab | 8.42 ± 0.12 d |
| T2 | 6.54 ± 0.06 a | 31.01 ± 1.67 a | 10.98 ± 0.05 d | 1.29 ± 0.05 cde | 29.63 ± 1.05 ab | 9.80 ± 0.65 abc |
| T3 | 6.45 ± 0.06 ab | 31.99 ± 3.12 a | 11.91 ± 0.05 c | 1.34 ± 0.06 cd | 28.22 ± 2.56 ab | 9.94 ± 0.55 abc |
| T4 | 6.48 ± 0.03 ab | 30.46 ± 2.27 a | 12.39 ± 0.03 b | 1.38 ± 0.04 bcd | 27.18 ± 1.52 ab | 9.92 ± 0.57 abc |
| T5 | 5.75 ± 0.04 e | 31.54 ± 1.89 a | 9.96 ± 0.04 e | 1.25 ± 0.05 de | 25.33 ± 0.78 b | 8.81 ± 0.27 cd |
| T6 | 6.26 ± 0.06 c | 29.72 ± 2.78 a | 11.91 ± 0.05 c | 1.43 ± 0.05 abc | 25.97 ± 1.38 ab | 9.11 ± 0.23 bcd |
| T7 | 6.34 ± 0.06 bc | 29.58 ± 3.12 a | 12.61 ± 0.05 ab | 1.49 ± 0.05 ab | 29.94 ± 3.21 ab | 10.24 ± 0.17 ab |
| T8 | 6.41 ± 0.02 abc | 31.05 ± 2.12 a | 12.95 ± 0.06 a | 1.53 ± 0.04 a | 32.30 ± 2.32 a | 10.41 ± 0.16 a |
| Late season | ||||||
| T1 | 5.84 ± 0.06 c | 30.27 ± 2.89 b | 9.57 ± 0.03 f | 1.22 ± 0.04 c | 24.60 ± 1.95 a | 9.48 ± 0.08 b |
| T2 | 6.26 ± 0.07 b | 30.96 ± 3.72 b | 12.55 ± 0.05 e | 1.39 ± 0.04 ab | 28.07 ± 3.21 a | 9.91 ± 0.25 b |
| T3 | 6.35 ± 0.06 b | 31.27 ± 1.98 ab | 13.41 ± 0.07 d | 1.41 ± 0.5 ab | 26.41 ± 1.25 a | 10.26 ± 0.64 ab |
| T4 | 6.68 ± 0.06 a | 31.39 2.89 ab | 13.89 ± 0.04 bc | 1.43 ± 0.05 a | 26.28 ± 1.06 a | 10.46 ± 0.52 ab |
| T5 | 5.74 ± 0.06 c | 30.58 ± 2.67 b | 9.71 ± 0.11 f | 1.28 ± 0.04 bc | 25.16 ± 2.32 a | 9.97 ± 0.77 ab |
| T6 | 6.26 ± 0.05 b | 31.58 ± 3.12 ab | 13.81 ± 0.07 c | 1.38 ± 0.04 ab | 26.28 ± 1.54 a | 10.91 ± 1.08 ab |
| T7 | 6.29 ± 0.06 b | 32.63 ± 2.18 a | 14.11 ± 0.06 b | 1.46 ± 0.04 a | 29.96 ± 1.78 a | 11.73 ± 0.42 a |
| T8 | 6.34 ± 0.07 b | 31.21 ± 1.89 ab | 14.46 ± 0.08a | 1.48 ± 0.02 a | 26.06 ± 1.48 a | 10.23 ± 0.38 ab |
| Treatments | Stem g N kg−1 | Leaves g N kg−1 | Panicles g N kg−1 |
|---|---|---|---|
| Early season | |||
| T1 | 3.79 ± 0.06 c | 11.90 ± 0.34 d | 9.48 ± 0.36 c |
| T2 | 4.91 ± 0.17 a | 13.82 ± 0.17 c | 10.37 ± 0.09 b |
| T3 | 4.88 ± 0.12 a | 15.47 ± 0.29 a | 11.60 ± 0.32 a |
| T4 | 4.87 ± 0.17 a | 14.83 ± 0.20 ab | 11.77 ± 0.27 a |
| T5 | 3.92 ± 0.09 bc | 12.41 ± 0.27 d | 9.85 ± 0.30 bc |
| T6 | 4.76 ± 0.21 a | 14.38 ± 0.07 bc | 11.72 ± 0.31 a |
| T7 | 4.43 ± 0.19 ab | 14.80 0.17 ab | 11.78 ± 0.32 a |
| T8 | 4.47 ± 0.30 a | 14.32 ± 0.32 bc | 11.97 ± 0.20 a |
| Late season | |||
| T1 | 4.64 ± 0.20 c | 12.40 ± 0.28 c | 9.61 ± 0.36 c |
| T2 | 6.09 ± 0.08 a | 14.53 ± 0.30 b | 10.50 ± 0.09 b |
| T3 | 6.01 ± 0.06 a | 15.44 ± 0.21 a | 11.73 ± 0.32 a |
| T4 | 6.17 ± 0.17 a | 15.64 ± 0.12 a | 11.90 ± 0.27 a |
| T5 | 5.05 ± 0.09 b | 12.81 ± 0.27 c | 10.03 ± 0.25 bc |
| T6 | 6.17 ± 0.11 a | 15.25 ± 0.11 a | 11.96 ± 0.37 a |
| T7 | 6.03 ± 0.05 a | 15.20 ± 0.17 ab | 12.00 ± 0.23 a |
| T8 | 6.16 ± 0.10 a | 15.31 ± 0.26 a | 11.90 ± 0.06 a |
| Treatments | Dry Matter kg h−1 | Grain Yield kg h−1 |
|---|---|---|
| Early season | ||
| T1 | 10,764.72 ± 177.14 c | 6621.32 ± 144.21 c |
| T2 | 12,340.67 ± 139.90 b | 7678.06 ± 164.28 b |
| T3 | 12,434.23 ± 160.66 ab | 8032.76 ± 43.46 ab |
| T4 | 12,400.67 ± 164.39 ab | 8418.80 ± 177.92 a |
| T5 | 11,067.33 ± 121.21 c | 7678.06 ± 239.63 b |
| T6 | 12,694.24 ± 55.80 ab | 7799.15 ± 140.11 b |
| T7 | 12,640.67± 57.95 ab | 8425.93 ± 126.01 a |
| T8 | 12,834.67 ± 159.68 a | 8447.29 ± 135.88 a |
| Late season | ||
| T1 | 10,018.21 ± 149.72 d | 6605.84 ± 78.01 d |
| T2 | 11,560.00 ± 147.35 b | 7910.11 ± 164.28 c |
| T3 | 11,402.33 ± 152.77 bc | 8264.81 ± 97.23 abc |
| T4 | 12,072.67 ± 130.67 a | 8365.95 ± 124.18 ab |
| T5 | 11,041.13 ± 178.63 c | 7910.11 ± 139.63 c |
| T6 | 11,525.23 ± 113.80 b | 8031.20 ± 140.11 bc |
| T7 | 12,208.67 ± 88.51 a | 8510.40 ± 90.62 a |
| T8 | 12,115.67 ± 131.68 a | 8533.19 ± 124.41 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullah, S.; Ali, I.; Yang, M.; Zhao, Q.; Iqbal, A.; Wu, X.; Ahmad, S.; Muhammad, I.; Khan, A.; Adnan, M.; et al. Partial Substitution of Urea with Biochar Induced Improvements in Soil Enzymes Activity, Ammonia-Nitrite Oxidizers, and Nitrogen Uptake in the Double-Cropping Rice System. Microorganisms 2023, 11, 527. https://doi.org/10.3390/microorganisms11020527
Ullah S, Ali I, Yang M, Zhao Q, Iqbal A, Wu X, Ahmad S, Muhammad I, Khan A, Adnan M, et al. Partial Substitution of Urea with Biochar Induced Improvements in Soil Enzymes Activity, Ammonia-Nitrite Oxidizers, and Nitrogen Uptake in the Double-Cropping Rice System. Microorganisms. 2023; 11(2):527. https://doi.org/10.3390/microorganisms11020527
Chicago/Turabian StyleUllah, Saif, Izhar Ali, Mei Yang, Quan Zhao, Anas Iqbal, Xiaoyan Wu, Shakeel Ahmad, Ihsan Muhammad, Abdullah Khan, Muhammad Adnan, and et al. 2023. "Partial Substitution of Urea with Biochar Induced Improvements in Soil Enzymes Activity, Ammonia-Nitrite Oxidizers, and Nitrogen Uptake in the Double-Cropping Rice System" Microorganisms 11, no. 2: 527. https://doi.org/10.3390/microorganisms11020527
APA StyleUllah, S., Ali, I., Yang, M., Zhao, Q., Iqbal, A., Wu, X., Ahmad, S., Muhammad, I., Khan, A., Adnan, M., Yuan, P., & Jiang, L. (2023). Partial Substitution of Urea with Biochar Induced Improvements in Soil Enzymes Activity, Ammonia-Nitrite Oxidizers, and Nitrogen Uptake in the Double-Cropping Rice System. Microorganisms, 11(2), 527. https://doi.org/10.3390/microorganisms11020527