



Current Metabolic Engineering Strategies for Photosynthetic Bioproduction in Cyanobacteria


Abstract
1. Introduction
2. Model Organisms and Emerging Species

{kind=link}
{kind=link}
| Organism | Genome Size (Mbp) | Plasmids | Doubling Time (hours) * | Growth Temperature * | CO2 Partial Pressure (%) * | Light Intensity (µmole photons m−2 s−1) * | Transformation Methods | Metabolism | Features | References |
|---|---|---|---|---|---|---|---|---|---|---|
| Syn6803 | 3.5 | 4 | 5.1 | 35 °C | 5 | 790 | NT ****, Conjugation, Electroporation | Mixotrophic, Autotrophic | Model organism, best studied cyanobacterium, numerous synthetic biology tools available | [22,24,88] |
| Syn7942 | 2.69 | 1 | 4.1 | 38 °C | 3 | 300 | NT ****, Conjugation, Electroporation | Autotrophic | Model organism, especially suited for circadian clock studies | [26,28,35] |
| Syn7002 | 3.0 | 7 | 2.5 2.5 ** | 30 °C 3 °C ** | 2 1 * | 760 250 ** | NT ****, Conjugation, Electroporation | Mixotrophic, Autotrophic | Model organisms, high cell doubling rate; resistant to salt and high light conditions | [39,89] |
| Ana7120 | 7.2 | 6 | 18–24 | 23–30 °C | 2 | 160 | Conjugation | Mixotrophic, Autotrophic | Model organism, nitrogen-fixing strain | [90,91,92] |
| Syn2973 | 2.69 | 2 | 2.1 | 41 °C | 3 | 500 | Conjugation, Electroporation | Autotrophic | Highest growth rate, numerous synthetic biology tools available | [51] |
| Syn11801 | 2.7 | / | 2.3 | 41 °C | 0.4 | 1000 | NT ****, Conjugation, Electroporation | Autotrophic | Fast-growing strain, thermophil | [58,59] |
| Syn11802 | 2.7 | / | 2.8 | 38 °C | 1 | 1000 | NT ****, Conjugation, Electroporation | Autotrophic | Fast-growing strain, thermophil | [60] |
| Syn11901 | 3.0 | 1 | 2.1 | 38 °C | 1 | 660 | NT ****, Conjugation | Mixotrophic, Autotrophic | Fast-growing strain, resistant to high-stress conditions | [61,93] |
| A. platensis C1 | 6.62 *** | / | ~60 | 30 °C | 0.3 | 108 | NT ****, Electroporation | Mixotrophic, Autotrophic | GRAS organism, production of proteins and pigments | [66,67,68,94,95] |
| F. diplosiphon | 9.9 *** | 14 | ~20 | 28–30 °C | N/A | 30 | Conjugation, Electroporation | Autotrophic | Suited for studying light absorption and photomorphogenesis | [96,97,98,99,100] |
| P. marinus | 1.6 | / | ~24 | 20 °C | N/A | 100 | / | Autotrophic | Suited as biomarker for ocean metabolism | [77,101,102] |
| G. violaceus 7421 | 4.6 | / | ~73 | 25 °C | N/A | 5 | Conjugation | Autotrophic | Suited for studying anoxygenic photosynthesis | [103,104,105,106] |
| T. elongatus BP-1 | 2.6 | / | 6 | 55 °C | 1.5 | 200 | Electroporation | Autotrophic | Thermophilic cyanobacterium, ideal for studying cyanobacterial photosystem | [83,107,108] |
3. Current Methods for DNA Transfer into Cyanobacteria
3.1. Genomic Integration and Extrachromosomal DNA Insertion
3.2. Transformation Methods
3.2.1. Conjugation
3.2.2. Electroporation
3.2.3. Natural Transformation
4. Tools for Manipulating Gene Expression in Cyanobacteria
4.1. Increasing Gene Expression by Promoter Engineering
4.1.1. Strong Native Promoters
4.1.2. Synthetic Inducible Promoters
4.1.3. Chemically Inducible Promoters
4.1.4. Environmentally Inducible Promoters
4.2. Designing Ribosomal Binding Sites for Improving Gene Expression
4.2.1. Ribosome Binding Site (RBS)
4.2.2. RiboJ
4.2.3. Bicistronic Design (BCD)
4.3. Using CRISPR for Gene Expression Regulation and Markerless Genome Editing
4.4. RNA-based Regulatory System
4.4.1. Riboswitches
4.4.2. Riboregulators
4.4.3. Toehold Switches and Small Transcription Activating RNAs (STARs)
5. Metabolic Engineering Approaches for Increasing Cyanobacterial Production
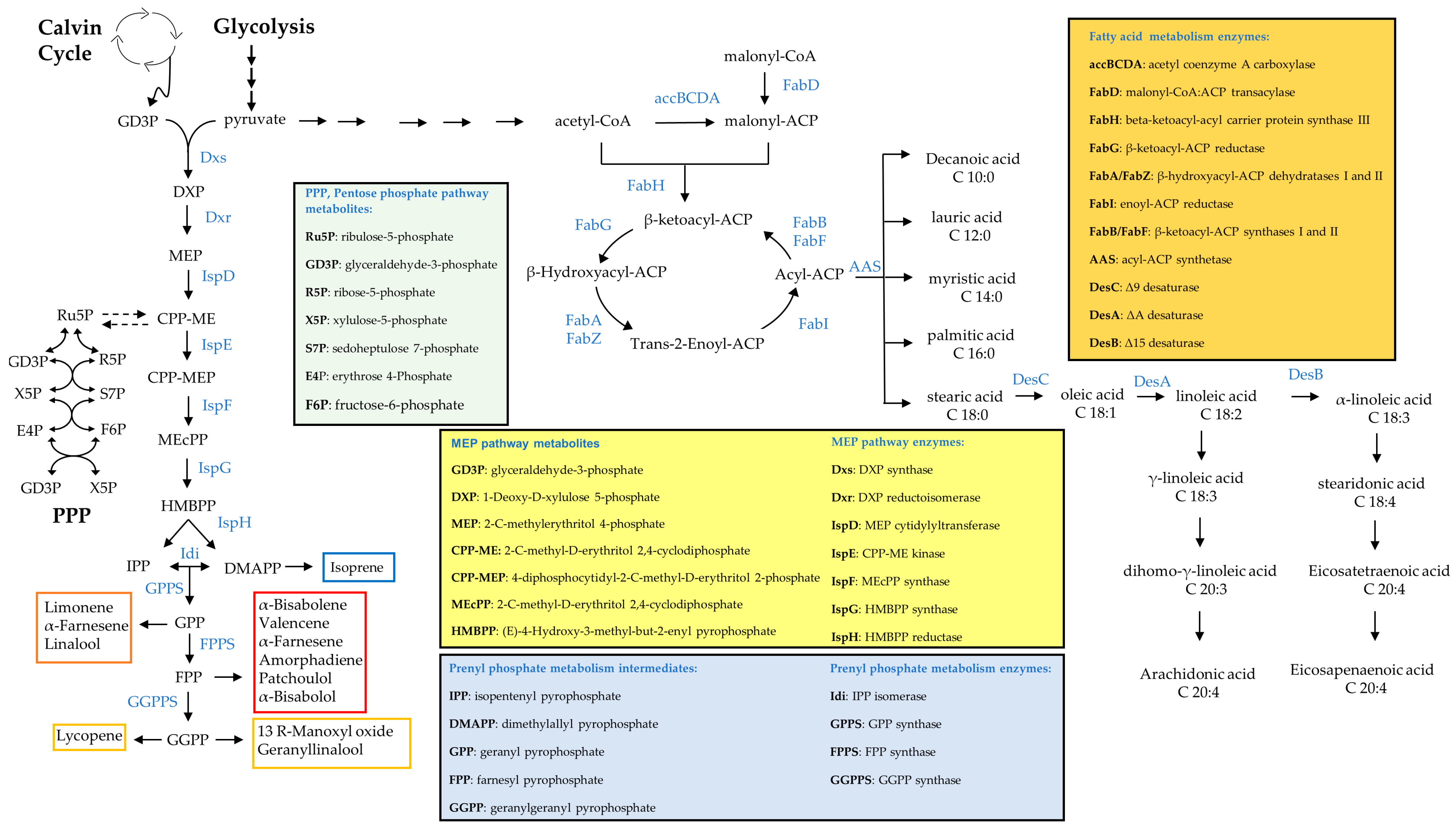
5.1. Fatty Acid Production Engineering
5.2. Carbohydrate Production Engineering
| Compounds | Organism | Titer | Reference |
|---|---|---|---|
| Free Fatty acids (secreted) | Syn6803 Syn7942 | 197 mg L−1 0.64 g L−1 | [232] [233] |
| Free Fatty acids | Syn11901 | 1.5 g L−1 | [61] |
| Fatty alcohol | Syn6803 Syn6803 | 9.7 μg OD730−1 L−1 2.8 mg gCDW−1 | [148] [234] |
| Alpha linoleic acid (C18:3) | Syn7942 | 22% of total FA | [235] |
| Alpha linoleic acid (C18:3), Stearidonic acid (C18:4), Eicosatetraenoic acid (C20:4) | Lep0902, Syn7002 | 40% of total FA | [236] |
| Total fatty acids | Syn7942 + R. glutinis | 39 mg L−1 | [242] |
| Sucrose | Syn7942 Syn2973 Syn2973 | 36.1 mg L−1 h−1 35.5 mg L−1 h−1 79 mg L−1 h−1 | [237] [55] [238] |
| Trehalose | Syn7942 | 5.7 g L−1 | [239] |
| Extracellular polysaccharide ‘cyanoflan’ | Cyanothece sp. CCY 0110 | 1.8 g L−1 | [240] |
| Isoprene (C5) | Syn7942 Syn6803 | 1.26 g L−1 12.3 mg gCDW−1 | [30] [243] |
| Limonene (C10) | Syn2973 Syn6803 Ana7120 Syn7002 | 16.4 mg L−1 6.7 mg L−1 3.6 μg L−1 4 mg L−1 | [244] [245] [246] [247] |
| (S)-linalool | Syn6803 | 11.4 mg L−1 | [248] |
| β-phellandrene | Syn6803 | 10 mg gCDW−1 | [249] |
| (E)-α-bisabolene (C15) | Syn6803 Syn7002 | 180 mg L−1 0.6 mg L−1 | [250] [247] |
| α-Farnesene | Ana7120 Syn7942 | 305.4 μg L−1 12.99 mg L−1 | [251] [252] |
| Amorphadiene | Syn7942 | 19.8 mg L−1 | [253] |
| Valencene | Syn6803 | 9.6 mg L−1 | [248] |
| β-Caryophyllene | Syn6803 | N.R.* | [254] |
| Patchoulol | Syn6803 | 17.3 mg L−1 | [255] |
| α-bisabolol | Syn6803 | 96.3 mg L−1 | [255] |
| 13R-manoyl oxide (C20) | Syn6803 | 2 mg L−1 | [256] |
| Geranyllinalool (Floating and intracellular) | Syn6803 | 390 μg gCDW−1 | [257] |
| Squalene (C30) | Syn7942 Syn6803 | 9.5 mg L−1 5.1 mg L−1 | [258] [259] |
| Lycopene (C40) | Syn6803 | 1.5 mg gCDW−1 | [260] |
5.3. Terpenoid Production Engineering
5.4. Nanocompartments Engineering
5.5. An Ecosystem Approach for a More Sustainable Production
6. Conclusions
Funding
Conflicts of Interest
References
- Schopf, J.W.; Packer, B.M. Early Archean (3.3-billion to 3.5-billion-year-old) microfossils from Warrawoona Group, Australia. Science 1987, 237, 70–73. [Google Scholar] [CrossRef]
- Wagner, S.C. Biological Nitrogen Fixation. Nat. Educ. Knowl. 2011, 3, 15. [Google Scholar]
- Tandeau de Marsac, N. Adaptation of cyanobacteria to environmental stimuli: New steps towards molecular mechanisms. FEMS Microbiol. Lett. 1993, 104, 119–189. [Google Scholar] [CrossRef]
- Jensen, P.E.; Leister, D. Chloroplast evolution, structure and functions. F1000prime Rep. 2014, 6, 40. [Google Scholar] [CrossRef]
- Nelson, N. Photosystems and global effects of oxygenic photosynthesis. Biochim. Biophys. Acta 2011, 1807, 856–863. [Google Scholar] [CrossRef]
- Buschke, N.; Schafer, R.; Becker, J.; Wittmann, C. Metabolic engineering of industrial platform microorganisms for biorefinery applications--optimization of substrate spectrum and process robustness by rational and evolutive strategies. Bioresour. Technol. 2013, 135, 544–554. [Google Scholar] [CrossRef]
- Hirokawa, Y.; Suzuki, I.; Hanai, T. Optimization of isopropanol production by engineered cyanobacteria with a synthetic metabolic pathway. J. Biosci. Bioeng. 2015, 119, 585–590. [Google Scholar] [CrossRef]
- Dismukes, G.C.; Carrieri, D.; Bennette, N.; Ananyev, G.M.; Posewitz, M.C. Aquatic phototrophs: Efficient alternatives to land-based crops for biofuels. Curr. Opin. Biotechnol. 2008, 19, 235–240. [Google Scholar] [CrossRef]
- Lau, N.S.; Matsui, M.; Abdullah, A.A. Cyanobacteria: Photoautotrophic Microbial Factories for the Sustainable Synthesis of Industrial Products. Biomed. Res. Int. 2015, 2015, 754934. [Google Scholar] [CrossRef]
- Vavitsas, K.; Kugler, A.; Satta, A.; Hatzinikolaou, D.G.; Lindblad, P.; Fewer, D.P.; Lindberg, P.; Toivari, M.; Stensjo, K. Doing synthetic biology with photosynthetic microorganisms. Physiol. Plant. 2021, 173, 624–638. [Google Scholar] [CrossRef]
- Vranova, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef]
- Tzin, V.; Galili, G. New insights into the shikimate and aromatic amino acids biosynthesis pathways in plants. Mol. Plant 2010, 3, 956–972. [Google Scholar] [CrossRef]
- Shabestary, K.; Hudson, E.P. Computational metabolic engineering strategies for growth-coupled biofuel production by Synechocystis. Metab. Eng. Commun. 2016, 3, 216–226. [Google Scholar] [CrossRef]
- Santos-Merino, M.; Singh, A.K.; Ducat, D.C. New Applications of Synthetic Biology Tools for Cyanobacterial Metabolic Engineering. Front. Bioeng. Biotechnol. 2019, 7, 33. [Google Scholar] [CrossRef]
- Hitchcock, A.; Hunter, C.N.; Canniffe, D.P. Progress and challenges in engineering cyanobacteria as chassis for light-driven biotechnology. Microb. Biotechnol. 2020, 13, 363–367. [Google Scholar] [CrossRef]
- Mukherjee, B.; Madhu, S.; Wangikar, P.P. The role of systems biology in developing non-model cyanobacteria as hosts for chemical production. Curr. Opin. Biotechnol. 2020, 64, 62–69. [Google Scholar] [CrossRef]
- NIH GenBank Sequence Database. Available online: https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 20 November 2022).
- Gale, G.A.R.; Schiavon Osorio, A.A.; Mills, L.A.; Wang, B.; Lea-Smith, D.J.; McCormick, A.J. Emerging Species and Genome Editing Tools: Future Prospects in Cyanobacterial Synthetic Biology. Microorganisms 2019, 7, 409. [Google Scholar] [CrossRef]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef]
- Los, D.A.; Zorina, A.; Sinetova, M.; Kryazhov, S.; Mironov, K.; Zinchenko, V.V. Stress sensors and signal transducers in cyanobacteria. Sensors 2010, 10, 2386–2415. [Google Scholar] [CrossRef]
- Google Scholar. Available online: https://scholar.google.com (accessed on 20 November 2022).
- Kaneko, T.; Sato, S.; Kotani, H.; Tanaka, A.; Asamizu, E.; Nakamura, Y.; Miyajima, N.; Hirosawa, M.; Sugiura, M.; Sasamoto, S.; et al. Sequence analysis of the genome of the unicellular cyanobacterium Synechocystis sp. strain PCC6803. II. Sequence determination of the entire genome and assignment of potential protein-coding regions. DNA Res. 1996, 3, 109–136. [Google Scholar] [CrossRef]
- Kaneko, T.; Tanaka, A.; Sato, S.; Kotani, H.; Sazuka, T.; Miyajima, N.; Sugiura, M.; Tabata, S. Sequence analysis of the genome of the unicellular cyanobacterium Synechocystis sp. strain PCC6803. I. Sequence features in the 1 Mb region from map positions 64% to 92% of the genome. DNA Res. 1995, 2, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; You, L.; Liu, D.; Hollinshead, W.; Tang, Y.J.; Zhang, F. Development of Synechocystis sp. PCC 6803 as a phototrophic cell factory. Mar. Drugs. 2013, 11, 2894–2916. [Google Scholar] [CrossRef] [PubMed]
- Golden, S.S. The international journeys and aliases of Synechococcus elongatus. N. Z. J. Bot. 2019, 57, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Shestakov, S.V.; Khyen, N.T. Evidence for genetic transformation in blue-green alga Anacystis nidulans. Mol. Gen. Genet. 1970, 107, 372–375. [Google Scholar] [CrossRef] [PubMed]
- Latifi, A.; Ruiz, M.; Zhang, C.C. Oxidative stress in cyanobacteria. FEMS Microbiol. Rev. 2009, 33, 258–278. [Google Scholar] [CrossRef] [PubMed]
- Golden, S.S.; Johnson, C.H.; Kondo, T. The cyanobacterial circadian system: A clock apart. Curr. Opin. Microbiol. 1998, 1, 669–673. [Google Scholar] [CrossRef]
- Clerico, E.M.; Ditty, J.L.; Golden, S.S. Specialized Techniques for Site-Directed Mutagenesis in Cyanobacteria. In Circadian Rhythms: Methods and Protocols; Rosato, E., Ed.; Humana Press: Totowa, NJ, USA, 2007; pp. 155–171. [Google Scholar]
- Gao, X.; Gao, F.; Liu, D.; Zhang, H.; Nie, X.; Yang, C. Engineering the methylerythritol phosphate pathway in cyanobacteria for photosynthetic isoprene production from CO2. Energy Environ. Sci. 2016, 9, 1400–1411. [Google Scholar] [CrossRef]
- Lan, E.I.; Chuang, D.S.; Shen, C.R.; Lee, A.M.; Ro, S.Y.; Liao, J.C. Metabolic engineering of cyanobacteria for photosynthetic 3-hydroxypropionic acid production from CO2 using Synechococcus elongatus PCC 7942. Metab. Eng. 2015, 31, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Hirokawa, Y.; Dempo, Y.; Fukusaki, E.; Hanai, T. Metabolic engineering for isopropanol production by an engineered cyanobacterium, Synechococcus elongatus PCC 7942, under photosynthetic conditions. J. Biosci. Bioeng. 2017, 123, 39–45. [Google Scholar] [CrossRef]
- Sarnaik, A.; Abernathy, M.H.; Han, X.; Ouyang, Y.; Xia, K.; Chen, Y.; Cress, B.; Zhang, F.; Lali, A.; Pandit, R.; et al. Metabolic engineering of cyanobacteria for photoautotrophic production of heparosan, a pharmaceutical precursor of heparin. Algal Res. 2019, 37, 57–63. [Google Scholar] [CrossRef]
- Hirokawa, Y.; Matsuo, S.; Hamada, H.; Matsuda, F.; Hanai, T. Metabolic engineering of Synechococcus elongatus PCC 7942 for improvement of 1,3-propanediol and glycerol production based on in silico simulation of metabolic flux distribution. Microb. Cell Fact. 2017, 16, 212. [Google Scholar] [CrossRef] [PubMed]
- Holtman, C.K.; Chen, Y.; Sandoval, P.; Gonzales, A.; Nalty, M.S.; Thomas, T.L.; Youderian, P.; Golden, S.S. High-throughput functional analysis of the Synechococcus elongatus PCC 7942 genome. DNA Res. 2005, 12, 103–115. [Google Scholar] [CrossRef]
- Broddrick, J.T.; Rubin, B.E.; Welkie, D.G.; Du, N.; Mih, N.; Diamond, S.; Lee, J.J.; Golden, S.S.; Palsson, B.O. Unique attributes of cyanobacterial metabolism revealed by improved genome-scale metabolic modeling and essential gene analysis. Proc. Natl. Acad. Sci. USA 2016, 113, E8344–E8353. [Google Scholar] [CrossRef]
- Van Baalen, C. Studies on Marine Blue-Green Algae. Bot. Mar. 1962, 4, 129–139. [Google Scholar] [CrossRef]
- Nomura, C.T.; Sakamoto, T.; Bryant, D.A. Roles for heme-copper oxidases in extreme high-light and oxidative stress response in the cyanobacterium Synechococcus sp. PCC 7002. Arch. Microbiol. 2006, 185, 471–479. [Google Scholar] [CrossRef]
- Bernstein, H.C.; McClure, R.S.; Hill, E.A.; Markillie, L.M.; Chrisler, W.B.; Romine, M.F.; McDermott, J.E.; Posewitz, M.C.; Bryant, D.A.; Konopka, A.E.; et al. Unlocking the Constraints of Cyanobacterial Productivity: Acclimations Enabling Ultrafast Growth. mBio 2016, 7, e00949-16. [Google Scholar] [CrossRef] [PubMed]
- Ruffing, A.M.; Jensen, T.J.; Strickland, L.M. Genetic tools for advancement of Synechococcus sp. PCC 7002 as a cyanobacterial chassis. Microb. Cell Fact. 2016, 15, 190. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.T.; Hill, E.A.; Kucek, L.A.; Konopka, A.E.; Beliaev, A.S.; Reed, J.L. Computational evaluation of Synechococcus sp. PCC 7002 metabolism for chemical production. Biotechnol. J. 2013, 8, 619–630. [Google Scholar] [CrossRef]
- Kumar, K.; Mella-Herrera, R.A.; Golden, J.W. Cyanobacterial heterocysts. Cold Spring Harb Perspect. Biol. 2010, 2, a000315. [Google Scholar] [CrossRef]
- Muro-Pastor, A.M.; Hess, W.R. Heterocyst differentiation: From single mutants to global approaches. Trends Microbiol. 2012, 20, 548–557. [Google Scholar] [CrossRef]
- Adolph, K.W.; Haselkorn, R. Isolation and characterization of a virus infecting the blue-green alga Nostoc muscorum. Virology 1971, 46, 200–208. [Google Scholar] [CrossRef]
- Flores, E.; Herrero, A. Compartmentalized function through cell differentiation in filamentous cyanobacteria. Nat. Rev. Microbiol. 2010, 8, 39–50. [Google Scholar] [CrossRef]
- Kultschar, B.; Llewellyn, C. Secondary Metabolites in Cyanobacteria; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Ungerer, J.; Pakrasi, H.B. Cpf1 Is A Versatile Tool for CRISPR Genome Editing Across Diverse Species of Cyanobacteria. Sci. Rep. 2016, 6, 39681. [Google Scholar] [CrossRef]
- Higo, A.; Isu, A.; Fukaya, Y.; Ehira, S.; Hisabori, T. Application of CRISPR Interference for Metabolic Engineering of the Heterocyst-Forming Multicellular Cyanobacterium Anabaena sp. PCC 7120. Plant Cell Physiol. 2018, 59, 119–127. [Google Scholar] [CrossRef]
- Norena-Caro, D.A.; Zuniga, C.; Pete, A.J.; Saemundsson, S.A.; Donaldson, M.R.; Adams, A.J.; Dooley, K.M.; Zengler, K.; Benton, M.G. Analysis of the cyanobacterial amino acid metabolism with a precise genome-scale metabolic reconstruction of Anabaena sp. UTEX 2576. Biochem. Eng. J. 2021, 171, 108008. [Google Scholar] [CrossRef]
- Cumino, A.C.; Marcozzi, C.; Barreiro, R.; Salerno, G.L. Carbon cycling in Anabaena sp. PCC 7120. Sucrose synthesis in the heterocysts and possible role in nitrogen fixation. Plant Physiol. 2007, 143, 1385–1397. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Liberton, M.; Cliften, P.F.; Head, R.D.; Jacobs, J.M.; Smith, R.D.; Koppenaal, D.W.; Brand, J.J.; Pakrasi, H.B. Synechococcus elongatus UTEX 2973, a fast growing cyanobacterial chassis for biosynthesis using light and CO(2). Sci. Rep. 2015, 5, 8132. [Google Scholar] [CrossRef]
- Tan, X.; Hou, S.; Song, K.; Georg, J.; Klahn, S.; Lu, X.; Hess, W.R. The primary transcriptome of the fast-growing cyanobacterium Synechococcus elongatus UTEX 2973. Biotechnol. Biofuels 2018, 11, 218. [Google Scholar] [CrossRef]
- Cui, J.; Sun, T.; Li, S.; Xie, Y.; Song, X.; Wang, F.; Chen, L.; Zhang, W. Improved Salt Tolerance and Metabolomics Analysis of Synechococcus elongatus UTEX 2973 by Overexpressing Mrp Antiporters. Front. Bioeng. Biotechnol. 2020, 8, 500. [Google Scholar] [CrossRef]
- Nagarajan, A.; Zhou, M.; Nguyen, A.Y.; Liberton, M.; Kedia, K.; Shi, T.; Piehowski, P.; Shukla, A.; Fillmore, T.L.; Nicora, C.; et al. Proteomic Insights into Phycobilisome Degradation, A Selective and Tightly Controlled Process in The Fast-Growing Cyanobacterium Synechococcus elongatus UTEX 2973. Biomolecules 2019, 9, 374. [Google Scholar] [CrossRef]
- Song, K.; Tan, X.; Liang, Y.; Lu, X. The potential of Synechococcus elongatus UTEX 2973 for sugar feedstock production. Appl. Microbiol. Biotechnol. 2016, 100, 7865–7875. [Google Scholar] [CrossRef] [PubMed]
- Mollers, K.B.; Cannella, D.; Jorgensen, H.; Frigaard, N.U. Cyanobacterial biomass as carbohydrate and nutrient feedstock for bioethanol production by yeast fermentation. Biotechnol. Biofuels 2014, 7, 64. [Google Scholar] [CrossRef]
- Wendt, K.E.; Ungerer, J.; Cobb, R.E.; Zhao, H.; Pakrasi, H.B. CRISPR/Cas9 mediated targeted mutagenesis of the fast growing cyanobacterium Synechococcus elongatus UTEX 2973. Microb. Cell Fact. 2016, 15, 115. [Google Scholar] [CrossRef]
- Jaiswal, D.; Sengupta, A.; Sohoni, S.; Sengupta, S.; Phadnavis, A.G.; Pakrasi, H.B.; Wangikar, P.P. Genome Features and Biochemical Characteristics of a Robust, Fast Growing and Naturally Transformable Cyanobacterium Synechococcus elongatus PCC 11801 Isolated from India. Sci. Rep. 2018, 8, 16632. [Google Scholar] [CrossRef]
- Sengupta, S.; Jaiswal, D.; Sengupta, A.; Shah, S.; Gadagkar, S.; Wangikar, P.P. Metabolic engineering of a fast-growing cyanobacterium Synechococcus elongatus PCC 11801 for photoautotrophic production of succinic acid. Biotechnol. Biofuels 2020, 13, 89. [Google Scholar] [CrossRef]
- Jaiswal, D.; Sengupta, A.; Sengupta, S.; Madhu, S.; Pakrasi, H.B.; Wangikar, P.P. A Novel Cyanobacterium Synechococcus elongatus PCC 11802 has Distinct Genomic and Metabolomic Characteristics Compared to its Neighbor PCC 11801. Sci. Rep. 2020, 10, 191. [Google Scholar] [CrossRef] [PubMed]
- Wlodarczyk, A.; Selao, T.T.; Norling, B.; Nixon, P.J. Newly discovered Synechococcus sp. PCC 11901 is a robust cyanobacterial strain for high biomass production. Commun. Biol. 2020, 3, 215. [Google Scholar] [CrossRef] [PubMed]
- Thuan, N.H.; An, T.T.; Shrestha, A.; Canh, N.X.; Sohng, J.K.; Dhakal, D. Recent Advances in Exploration and Biotechnological Production of Bioactive Compounds in Three Cyanobacterial Genera: Nostoc, Lyngbya and Microcystis. Front. Chem. 2019, 7, 604. [Google Scholar] [CrossRef]
- Karkos, P.D.; Leong, S.C.; Karkos, C.D.; Sivaji, N.; Assimakopoulos, D.A. Spirulina in clinical practice: Evidence-based human applications. Evid. Based Complement. Altern. Med. 2011, 2011, 531053. [Google Scholar] [CrossRef]
- Wan, D.; Wu, Q.; Kuča, K. Spirulina. In Nutraceuticals; Gupta, R.C., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 569–583. [Google Scholar]
- Food and Drug Administration. Spirulina, the Dried Biomass of Arthrospira Platensis. Agency Response Letter GRAS Notice No. GRN000127. 10 April 2003. Available online: https://www.accessdata.fda.gov/scripts/fdcc/index.cfm?set=GRASNotices&id=127 (accessed on 4 February 2023).
- Jeamton, W.; Dulsawat, S.; Tanticharoen, M.; Vonshak, A.; Cheevadhanarak, S. Overcoming Intrinsic Restriction Enzyme Barriers Enhances Transformation Efficiency in Arthrospira platensis C1. Plant Cell Physiol. 2017, 58, 822–830. [Google Scholar] [CrossRef]
- Jester, B.W.; Zhao, H.; Gewe, M.; Adame, T.; Perruzza, L.; Bolick, D.T.; Agosti, J.; Khuong, N.; Kuestner, R.; Gamble, C.; et al. Development of spirulina for the manufacture and oral delivery of protein therapeutics. Nat. Biotechnol. 2022, 40, 956–964. [Google Scholar] [CrossRef]
- Hilzinger, J.M.; Friedline, S.; Sivanandan, D.; Cheng, Y.-F.; Yamazaki, S.; Clark, D.S.; Skerker, J.M.; Arkin, A.P. Acetaminophen production in the edible, filamentous cyanobacterium Arthrospira platensis. bioRxiv 2022, 2022.06.2030.498297. [Google Scholar] [CrossRef]
- Kumar, V.; Maurya, P.K.; Mondal, S.; Sinha, R.P.; Singh, S.P. Photomorphogenesis in the Cyanobacterium Fremyella diplosiphon Improves Photosynthetic Efficiency. In Cyanobacteria; Mishra, A.K., Tiwari, D.N., Rai, A.N., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 131–143. [Google Scholar]
- Haney, A.M.; Kehoe, D.M. Fremyella diplosiphon. Trends Microbiol. 2019, 27, 562–563. [Google Scholar] [CrossRef] [PubMed]
- Biller, S.J.; Berube, P.M.; Lindell, D.; Chisholm, S.W. Prochlorococcus: The structure and function of collective diversity. Nat. Rev. Microbiol. 2015, 13, 13–27. [Google Scholar] [CrossRef]
- Palenik, B.; Haselkorn, R. Multiple evolutionary origins of prochlorophytes, the chlorophyll b-containing prokaryotes. Nature 1992, 355, 265–267. [Google Scholar] [CrossRef] [PubMed]
- Partensky, F.; Hess, W.R.; Vaulot, D. Prochlorococcus, a marine photosynthetic prokaryote of global significance. Microbiol. Mol. Biol. Rev. 1999, 63, 106–127. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.L.; Chisholm, S.W. Code and context: Prochlorococcus as a model for cross-scale biology. Trends Microbiol. 2007, 15, 398–407. [Google Scholar] [CrossRef]
- Shaw, S.L.; Chisholm, S.W.; Prinn, R.G. Isoprene production by Prochlorococcus, a marine cyanobacterium and other phytoplankton. Mar. Chem. 2003, 80, 227–245. [Google Scholar] [CrossRef]
- Bourgade, B.; Stensjo, K. Synthetic biology in marine cyanobacteria: Advances and challenges. Front. Microbiol. 2022, 13, 994365. [Google Scholar] [CrossRef]
- Laurenceau, R.; Bliem, C.; Osburne, M.S.; Becker, J.W.; Biller, S.J.; Cubillos-Ruiz, A.; Chisholm, S.W. Toward a genetic system in the marine cyanobacterium Prochlorococcus. Access. Microbiol. 2020, 2, acmi000107. [Google Scholar] [CrossRef]
- Rippka, R.; Waterbury, J.; Cohen-Bazire, G. A cyanobacterium which lacks thylakoids. Arch. Microbiol. 1974, 100, 419–436. [Google Scholar] [CrossRef]
- Bernat, G.; Schreiber, U.; Sendtko, E.; Stadnichuk, I.N.; Rexroth, S.; Rogner, M.; Koenig, F. Unique properties vs. common themes: The atypical cyanobacterium Gloeobacter violaceus PCC 7421 is capable of state transitions and blue-light-induced fluorescence quenching. Plant Cell Physiol. 2012, 53, 528–542. [Google Scholar] [CrossRef] [PubMed]
- Montejano, G.; Becerra-Absalón, I.; Gold-Morgan, M.; Osorio-Santos, K. Gloeobacter violaceus: Primitive reproductive scheme and its significance. Plant Syst. Evol. 2018, 304, 1221–1229. [Google Scholar] [CrossRef]
- Yamaoka, T.; Satoh, K.; Katoh, S. Photosynthetic activities of a thermophilic blue-green alga. Plant Cell Physiol. 1978, 19, 943–954. [Google Scholar] [CrossRef]
- Onai, K.; Morishita, M.; Kaneko, T.; Tabata, S.; Ishiura, M. Natural transformation of the thermophilic cyanobacterium Thermosynechococcus elongatus BP-1: A simple and efficient method for gene transfer. Mol. Genet. Genom. 2004, 271, 50–59. [Google Scholar] [CrossRef]
- Nakamura, Y.; Kaneko, T.; Sato, S.; Ikeuchi, M.; Katoh, H.; Sasamoto, S.; Watanabe, A.; Iriguchi, M.; Kawashima, K.; Kimura, T.; et al. Complete genome structure of the thermophilic cyanobacterium Thermosynechococcus elongatus BP-1. DNA Res. 2002, 9, 123–130. [Google Scholar] [CrossRef]
- Elli, A.F.; Jelezko, F.; Tietz, C.; Studier, H.; Brecht, M.; Bittl, R.; Wrachtrup, J. Red pool chlorophylls of photosystem I of the cyanobacterium Thermosynechococcus elongatus: A single-molecule study. Biochemistry 2006, 45, 1454–1458. [Google Scholar] [CrossRef]
- Falke, S.; Feiler, C.; Chapman, H.; Sarrou, I. Crystal structures of native cytochrome c(6) from Thermosynechococcus elongatus in two different space groups and implications for its oligomerization. Acta Cryst. F Struct. Biol. Commun. 2020, 76, 444–452. [Google Scholar] [CrossRef]
- Kern, J.; Loll, B.; Luneberg, C.; DiFiore, D.; Biesiadka, J.; Irrgang, K.D.; Zouni, A. Purification, characterisation and crystallisation of photosystem II from Thermosynechococcus elongatus cultivated in a new type of photobioreactor. Biochim. Biophys. Acta 2005, 1706, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Kupitz, C.; Grotjohann, I.; Conrad, C.E.; Roy-Chowdhury, S.; Fromme, R.; Fromme, P. Microcrystallization techniques for serial femtosecond crystallography using photosystem II from Thermosynechococcus elongatus as a model system. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130316. [Google Scholar] [CrossRef]
- Zavřel, T.; Sinetova, M.A.; Búzová, D.; Literáková, P.; Červený, J. Characterization of a model cyanobacteriumSynechocystissp. PCC 6803 autotrophic growth in a flat-panel photobioreactor. Eng. Life Sci. 2015, 15, 122–132. [Google Scholar] [CrossRef]
- Ludwig, M.; Bryant, D.A. Synechococcus sp. Strain PCC 7002 Transcriptome: Acclimation to Temperature, Salinity, Oxidative Stress and Mixotrophic Growth Conditions. Front. Microbiol. 2012, 3, 354. [Google Scholar] [CrossRef] [PubMed]
- Videau, P.; Cozy, L.M. Anabaena sp. strain PCC 7120: Laboratory Maintenance, Cultivation and Heterocyst Induction. Curr. Protoc. Microbiol. 2019, 52, e71. [Google Scholar] [CrossRef]
- Prasanna, R.; Kumar, R.; Sood, A.; Prasanna, B.M.; Singh, P.K. Morphological, physiochemical and molecular characterization of Anabaena strains. Microbiol. Res. 2006, 161, 187–202. [Google Scholar] [CrossRef]
- Berberoğlu, H.; Jay, J.; Pilon, L. Effect of nutrient media on photobiological hydrogen production by Anabaena variabilis ATCC 29413. Int. J. Hydrog. Energy 2008, 33, 1172–1184. [Google Scholar] [CrossRef]
- Selão, T.T.; Włodarczyk, A.; Nixon, P.J.; Norling, B. Growth and selection of the cyanobacterium Synechococcus sp. PCC 7002 using alternative nitrogen and phosphorus sources. Metab. Eng. 2019, 54, 255–263. [Google Scholar] [CrossRef] [PubMed]
- El Baky, H.H.A.; El Baroty, G.S.; Mostafa, E.M. Optimization Growth of Spirulina (Arthrospira) Platensis in Photobioreactor Under Varied Nitrogen Concentration for Maximized Biomass, Carotenoids and Lipid Contents. Recent Pat. Food Nutr. Agric. 2020, 11, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Qin, S.; Hu, Y.; Song, Z.; Ying, J.; Li, P.; Dong, W.; Zhao, F.; Yang, H.; Bao, Q. Whole genomic DNA sequencing and comparative genomic analysis of Arthrospira platensis: High genome plasticity and genetic diversity. DNA Res. 2016, 23, 325–338. [Google Scholar] [CrossRef]
- Agostoni, M.; Lucker, B.F.; Smith, M.A.Y.; Kanazawa, A.; Blanchard, G.J.; Kramer, D.M.; Montgomery, B.L. Competition-based phenotyping reveals a fitness cost for maintaining phycobilisomes under fluctuating light in the cyanobacterium Fremyella diplosiphon. Algal Res. 2016, 15, 110–119. [Google Scholar] [CrossRef]
- Quest, B.; Hubschmann, T.; Sharda, S.; Tandeau de Marsac, N.; Gartner, W. Homologous expression of a bacterial phytochrome. The cyanobacterium Fremyella diplosiphon incorporates biliverdin as a genuine, functional chromophore. FEBS J. 2007, 274, 2088–2098. [Google Scholar] [CrossRef]
- Tabatabai, B.; Fathabad, S.G.; Bonyi, E.; Rajini, S.; Aslan, K.; Sitther, V. Nanoparticle-mediated Impact on Growth and Fatty Acid Methyl Ester Composition in the Cyanobacterium Fremyella diplosiphon. Bioenergy Res. 2019, 12, 409–418. [Google Scholar] [CrossRef]
- Tabatabai, B.; Adusei, A.; Shrivastava, A.K.; Singh, P.K.; Sitther, V. Nitrogen Deprivation in Fremyella diplosiphon Augments Lipid Production without Affecting Growth. Energies 2020, 13, 5769. [Google Scholar] [CrossRef]
- Tabatabai, B.; Arumanayagam, A.S.; Enitan, O.; Mani, A.; Natarajan, S.S.; Sitther, V. Overexpression of hlyB and mdh genes confers halotolerance in Fremyella diplosiphon, a freshwater cyanobacterium. Enzym. Microb. Technol. 2017, 103, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Tetu, S.G.; Sarker, I.; Schrameyer, V.; Pickford, R.; Elbourne, L.D.H.; Moore, L.R.; Paulsen, I.T. Plastic leachates impair growth and oxygen production in Prochlorococcus, the ocean’s most abundant photosynthetic bacteria. Commun. Biol. 2019, 2, 184. [Google Scholar] [CrossRef] [PubMed]
- Dufresne, A.; Salanoubat, M.; Partensky, F.; Artiguenave, F.; Axmann, I.M.; Barbe, V.; Duprat, S.; Galperin, M.Y.; Koonin, E.V.; Le Gall, F.; et al. Genome sequence of the cyanobacterium Prochlorococcus marinus SS120, a nearly minimal oxyphototrophic genome. Proc. Natl. Acad. Sci. USA 2003, 100, 10020–10025. [Google Scholar] [CrossRef]
- Guo, H.; Xu, X. Broad host range plasmid-based gene transfer system in the cyanobacterium Gloeobacter violaceus which lacks thylakoids *. Prog. Nat. Sci. 2004, 14, 31–35. [Google Scholar] [CrossRef]
- Koyama, K.; Suzuki, H.; Noguchi, T.; Akimoto, S.; Tsuchiya, T.; Mimuro, M. Oxygen evolution in the thylakoid-lacking cyanobacterium Gloeobacter violaceus PCC 7421. Biochim. Biophys. Acta 2008, 1777, 369–378. [Google Scholar] [CrossRef]
- Kula-Maximenko, M.; Zielinski, K.J.; Slesak, I. The Role of Selected Wavelengths of Light in the Activity of Photosystem II in Gloeobacter violaceus. Int. J. Mol. Sci. 2021, 22, 4021. [Google Scholar] [CrossRef]
- Nakamura, Y.; Kaneko, T.; Sato, S.; Mimuro, M.; Miyashita, H.; Tsuchiya, T.; Sasamoto, S.; Watanabe, A.; Kawashima, K.; Kishida, Y.; et al. Complete genome structure of Gloeobacter violaceus PCC 7421, a cyanobacterium that lacks thylakoids. DNA Res. 2003, 10, 137–145. [Google Scholar] [CrossRef]
- Anne Knight, R.; Brand, J.J.; Alper, H.S. Coordinated Response and Regulation of Carotenogenesis in Thermosynechococcus elongatus (BP-1): Implications for Commercial Application. Ph.D. Thesis, The University of Texas at Austin, Austin, TX, USA, 2014. [Google Scholar]
- Sacko, O.; Barnes, C.L.; Greene, L.H.; Lee, J.W. Survivability of Wild-Type and Genetically Engineered Thermosynechococcus elongatus BP1 with Different Temperature Conditions. Appl. Biosaf. 2020, 25, 104–117. [Google Scholar] [CrossRef]
- Elhai, J.; Wolk, C.P. Conjugal transfer of DNA to cyanobacteria. Methods Enzym. 1988, 167, 747–754. [Google Scholar] [CrossRef]
- Arora, N.; Jaiswal, D.; Sengupta, S.; Wangikar, P.P. Metabolic engineering of cyanobacteria for production of platform chemicals: A synthetic biology approach. In Handbook of Algal Science, Technology and Medicine; Konur, O., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 127–145. [Google Scholar]
- Ng, A.H.; Berla, B.M.; Pakrasi, H.B. Fine-Tuning of Photoautotrophic Protein Production by Combining Promoters and Neutral Sites in the Cyanobacterium Synechocystis sp. Strain PCC 6803. Appl. Environ. Microbiol. 2015, 81, 6857–6863. [Google Scholar] [CrossRef] [PubMed]
- Pinto, F.; Pacheco, C.C.; Oliveira, P.; Montagud, A.; Landels, A.; Couto, N.; Wright, P.C.; Urchueguía, J.F.; Tamagnini, P. Improving a Synechocystis-based photoautotrophic chassis through systematic genome mapping and validation of neutral sites. DNA Res. 2015, 22, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Zerulla, K.; Ludt, K.; Soppa, J. The ploidy level of Synechocystis sp. PCC 6803 is highly variable and is influenced by growth phase and by chemical and physical external parameters. Microbiology 2016, 162, 730–739. [Google Scholar] [CrossRef] [PubMed]
- Lea-Smith, D.J.; Vasudevan, R.; Howe, C.J. Generation of Marked and Markerless Mutants in Model Cyanobacterial Species. J. Vis. Exp. 2016, 111, 54001. [Google Scholar] [CrossRef]
- Pope, M.A.; Hodge, J.A.; Nixon, P.J. An Improved Natural Transformation Protocol for the Cyanobacterium Synechocystis sp. PCC 6803. Front. Plant Sci. 2020, 11, 372. [Google Scholar] [CrossRef]
- Riaz, S.; Jiang, Y.; Xiao, M.; You, D.; Klepacz-Smółka, A.; Rasul, F.; Daroch, M. Generation of miniploid cells and improved natural transformation procedure for a model cyanobacterium Synechococcus elongatus PCC 7942. Front. Microbiol. 2022, 13, 959043. [Google Scholar] [CrossRef]
- Taton, A.; Unglaub, F.; Wright, N.E.; Zeng, W.Y.; Paz-Yepes, J.; Brahamsha, B.; Palenik, B.; Peterson, T.C.; Haerizadeh, F.; Golden, S.S.; et al. Broad-host-range vector system for synthetic biology and biotechnology in cyanobacteria. Nucleic Acids Res. 2014, 42, e136. [Google Scholar] [CrossRef]
- Chen, Y.; Taton, A.; Go, M.; London, R.E.; Pieper, L.M.; Golden, S.S.; Golden, J.W. Self-replicating shuttle vectors based on pANS, a small endogenous plasmid of the unicellular cyanobacterium Synechococcus elongatus PCC 7942. Microbiology (Reading) 2016, 162, 2029–2041. [Google Scholar] [CrossRef]
- Jin, H.; Wang, Y.; Idoine, A.; Bhaya, D. Construction of a Shuttle Vector Using an Endogenous Plasmid From the Cyanobacterium Synechocystis sp. PCC6803. Front. Microbiol. 2018, 9, 1662. [Google Scholar] [CrossRef]
- Bishe, B.; Taton, A.; Golden, J.W. Modification of RSF1010-Based Broad-Host-Range Plasmids for Improved Conjugation and Cyanobacterial Bioprospecting. iScience 2019, 20, 216–228. [Google Scholar] [CrossRef]
- Ferreira, E.A.; Pacheco, C.C.; Pinto, F.; Pereira, J.; Lamosa, P.; Oliveira, P.; Kirov, B.; Jaramillo, A.; Tamagnini, P. Expanding the toolbox for Synechocystis sp. PCC 6803: Validation of replicative vectors and characterization of a novel set of promoters. Synth. Biol. 2018, 3, ysy014. [Google Scholar] [CrossRef]
- Behle, A.; Axmann, I.M. pSHDY: A New Tool for Genetic Engineering of Cyanobacteria. In Plant Synthetic Biology: Methods and Protocols; Zurbriggen, M.D., Ed.; Springer US: New York, NY, USA, 2022; pp. 67–79. [Google Scholar]
- Kaltenbrunner, A.; Reimann, V.; Hoffmann, U.A.; Aoyagi, T.; Sakata, M.; Nimura-Matsune, K.; Watanabe, S.; Steglich, C.; Wilde, A.; Hess, W.R. Regulation of pSYSA defense plasmid copy number in Synechocystis through RNase E and a highly transcribed asRNA. bioRxiv 2022, 2022.11.30.518505. [Google Scholar] [CrossRef]
- Vioque, A. Transformation of cyanobacteria. Adv. Exp. Med. Biol. 2007, 616, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Koksharova, O.A.; Wolk, C.P. Genetic tools for cyanobacteria. Appl. Microbiol. Biotechnol. 2002, 58, 123–137. [Google Scholar] [CrossRef]
- Eaton-Rye, J.J. The Construction of Gene Knockouts in the Cyanobacterium Synechocystis sp. PCC 6803. In Photosynthesis Research Protocols; Carpentier, R., Ed.; Humana Press: Totowa, NJ, USA, 2004; pp. 309–324. [Google Scholar]
- Al-Haj, L.; Lui, Y.T.; Abed, R.M.; Gomaa, M.A.; Purton, S. Cyanobacteria as Chassis for Industrial Biotechnology: Progress and Prospects. Life 2016, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Elhai, J.; Vepritskiy, A.; Muro-Pastor, A.M.; Flores, E.; Wolk, C.P. Reduction of conjugal transfer efficiency by three restriction activities of Anabaena sp. strain PCC 7120. J. Bacteriol. 1997, 179, 1998–2005. [Google Scholar] [CrossRef]
- Wendt, K.E.; Pakrasi, H.B. Genomics Approaches to Deciphering Natural Transformation in Cyanobacteria. Front. Microbiol. 2019, 10, 1259. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.V.; Martens, S.B.B.; Lanes, C.F.C.; Marins, L.F. Improved genetic transformation of Synechococcus elongatus PCC 7942 using linear DNA fragments in association with a DNase inhibitor. Biotechnol. Res. Innov. 2017, 1, 123–128. [Google Scholar] [CrossRef]
- Kufryk, G.I.; Sachet, M.; Schmetterer, G.; Vermaas, W.F. Transformation of the cyanobacterium Synechocystis sp. PCC 6803 as a tool for genetic mapping: Optimization of efficiency. FEMS Microbiol. Lett. 2002, 206, 215–219. [Google Scholar] [CrossRef]
- Nagarajan, A.; Winter, R.; Eaton-Rye, J.; Burnap, R. A synthetic DNA and fusion PCR approach to the ectopic expression of high levels of the D1 protein of photosystem II in Synechocystis sp. PCC 6803. J. Photochem. Photobiol. B 2011, 104, 212–219. [Google Scholar] [CrossRef]
- Stucken, K.; Koch, R.; Dagan, T. Cyanobacterial defense mechanisms against foreign DNA transfer and their impact on genetic engineering. Biol. Res. 2013, 46, 373–382. [Google Scholar] [CrossRef]
- Hall, R.J.; Whelan, F.J.; McInerney, J.O.; Ou, Y.; Domingo-Sananes, M.R. Horizontal Gene Transfer as a Source of Conflict and Cooperation in Prokaryotes. Front. Microbiol. 2020, 11, 1569. [Google Scholar] [CrossRef]
- Stevens, S.E.; Porter, R.D. Transformation in Agmenellum quadruplicatum. Proc. Natl. Acad. Sci. USA 1980, 77, 6052–6056. [Google Scholar] [CrossRef]
- Grigorieva, G.; Shestakov, S. Transformation in the cyanobacterium Synechocystis sp. 6803. FEMS Microbiol. Lett. 1982, 13, 367–370. [Google Scholar] [CrossRef]
- Dittmann, E.; Neilan, B.A.; Erhard, M.; von Döhren, H.; Börner, T. Insertional mutagenesis of a peptide synthetase gene that is responsible for hepatotoxin production in the cyanobacterium Microcystis aeruginosa PCC 7806. Mol. Microbiol. 1997, 26, 779–787. [Google Scholar] [CrossRef]
- Nies, F.; Mielke, M.; Pochert, J.; Lamparter, T. Natural transformation of the filamentous cyanobacterium Phormidium lacuna. PLoS ONE 2020, 15, e0234440. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Sun, T.; Xu, C.; Chen, L.; Zhang, W. Development and optimization of genetic toolboxes for a fast-growing cyanobacterium Synechococcus elongatus UTEX 2973. Metab Eng. 2018, 48, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Schirmacher, A.M.; Hanamghar, S.S.; Zedler, J.A.Z. Function and Benefits of Natural Competence in Cyanobacteria: From Ecology to Targeted Manipulation. Life 2020, 10, 249. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Pakrasi, H.B. Exploring native genetic elements as plug-in tools for synthetic biology in the cyanobacterium Synechocystis sp. PCC 6803. Microb. Cell Fact. 2018, 17, 48. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.; Jansson, C. Influence of light on accumulation of photosynthesis-specific transcripts in the cyanobacterium Synechocystis 6803. Plant Mol. Biol. 1989, 13, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, P.; Park, S.; Melis, A. Engineering a platform for photosynthetic isoprene production in cyanobacteria, using Synechocystis as the model organism. Metab. Eng. 2010, 12, 70–79. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, H.; Zhang, Y.; Li, Y.; Ma, Y. Designing and creating a modularized synthetic pathway in cyanobacterium Synechocystis enables production of acetone from carbon dioxide. Metab. Eng. 2012, 14, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.H.; Camsund, D.; Lindblad, P.; Heidorn, T. Design and characterization of molecular tools for a Synthetic Biology approach towards developing cyanobacterial biotechnology. Nucleic Acids Res. 2010, 38, 2577–2593. [Google Scholar] [CrossRef]
- Deng, M.D.; Coleman, J.R. Ethanol synthesis by genetic engineering in cyanobacteria. Appl. Environ. Microbiol. 1999, 65, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Piechura, J.R.; Amarnath, K.; O’Shea, E.K. Natural changes in light interact with circadian regulation at promoters to control gene expression in cyanobacteria. Elife 2017, 6, e32032. [Google Scholar] [CrossRef]
- Tan, X.; Yao, L.; Gao, Q.; Wang, W.; Qi, F.; Lu, X. Photosynthesis driven conversion of carbon dioxide to fatty alcohols and hydrocarbons in cyanobacteria. Metab. Eng. 2011, 13, 169–176. [Google Scholar] [CrossRef]
- Gao, Z.; Zhao, H.; Li, Z.; Tan, X.; Lu, X. Photosynthetic production of ethanol from carbon dioxide in genetically engineered cyanobacteria. Energy Environ. Sci. 2012, 5, 9857–9865. [Google Scholar] [CrossRef]
- Wegelius, A.; Li, X.; Turco, F.; Stensjo, K. Design and characterization of a synthetic minimal promoter for heterocyst-specific expression in filamentous cyanobacteria. PLoS ONE 2018, 13, e0203898. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, H.; Meng, H.; Zhu, Y.; Bao, G.; Zhang, Y.; Li, Y.; Ma, Y. Discovery of a super-strong promoter enables efficient production of heterologous proteins in cyanobacteria. Sci. Rep. 2014, 4, 4500. [Google Scholar] [CrossRef]
- Englund, E.; Liang, F.; Lindberg, P. Evaluation of promoters and ribosome binding sites for biotechnological applications in the unicellular cyanobacterium Synechocystis sp. PCC 6803. Sci. Rep. 2016, 6, 36640. [Google Scholar] [CrossRef]
- Camsund, D.; Lindblad, P. Engineered transcriptional systems for cyanobacterial biotechnology. Front. Bioeng. Biotechnol. 2014, 2, 40. [Google Scholar] [CrossRef]
- Markley, A.L.; Begemann, M.B.; Clarke, R.E.; Gordon, G.C.; Pfleger, B.F. Synthetic biology toolbox for controlling gene expression in the cyanobacterium Synechococcus sp. strain PCC 7002. ACS Synth. Biol. 2015, 4, 595–603. [Google Scholar] [CrossRef]
- Li, H.; Shen, C.R.; Huang, C.H.; Sung, L.Y.; Wu, M.Y.; Hu, Y.C. CRISPR-Cas9 for the genome engineering of cyanobacteria and succinate production. Metab Eng. 2016, 38, 293–302. [Google Scholar] [CrossRef]
- Sengupta, A.; Madhu, S.; Wangikar, P.P. A Library of Tunable, Portable and Inducer-Free Promoters Derived from Cyanobacteria. ACS Synth. Biol. 2020, 9, 1790–1801. [Google Scholar] [CrossRef]
- Gilbert, W.; Muller-Hill, B. Isolation of the lac repressor. Proc. Natl. Acad. Sci. USA 1966, 56, 1891–1898. [Google Scholar] [CrossRef]
- Niederholtmeyer, H.; Wolfstadter, B.T.; Savage, D.F.; Silver, P.A.; Way, J.C. Engineering cyanobacteria to synthesize and export hydrophilic products. Appl. Environ. Microbiol. 2010, 76, 3462–3466. [Google Scholar] [CrossRef]
- Albers, S.C.; Gallegos, V.A.; Peebles, C.A. Engineering of genetic control tools in Synechocystis sp. PCC 6803 using rational design techniques. J. Biotechnol. 2015, 216, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Peters, G.; De Paepe, B.; De Wannemaeker, L.; Duchi, D.; Maertens, J.; Lammertyn, J.; De Mey, M. Development of N-acetylneuraminic acid responsive biosensors based on the transcriptional regulator NanR. Biotechnol. Bioeng. 2018, 115, 1855–1865. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, S.; Zhang, F.; Sun, T.; Chen, L.; Zhang, W. Development of a N-Acetylneuraminic Acid-Based Sensing and Responding Switch for Orthogonal Gene Regulation in Cyanobacterial Synechococcus Strains. ACS Synth. Biol. 2021, 10, 1920–1930. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.H.; Lindblad, P. Wide-dynamic-range promoters engineered for cyanobacteria. J. Biol. Eng. 2013, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Cavet, J.S.; Borrelly, G.P.; Robinson, N.J. Zn, Cu and Co in cyanobacteria: Selective control of metal availability. FEMS Microbiol. Rev. 2003, 27, 165–181. [Google Scholar] [CrossRef]
- Peca, L.; Kós, P.B.; Vass, I. Characterization of the activity of heavy metal-responsive promoters in the cyanobacterium Synechocystis PCC 6803. Acta Biol. Hung. 2007, 58, 11–22. [Google Scholar] [CrossRef]
- Roussou, S.; Albergati, A.; Liang, F.; Lindblad, P. Engineered cyanobacteria with additional overexpression of selected Calvin-Benson-Bassham enzymes show further increased ethanol production. Metab. Eng. Commun. 2021, 12, e00161. [Google Scholar] [CrossRef] [PubMed]
- Loeschcke, A.; Dienst, D.; Wewer, V.; Hage-Hulsmann, J.; Dietsch, M.; Kranz-Finger, S.; Huren, V.; Metzger, S.; Urlacher, V.B.; Gigolashvili, T.; et al. The photosynthetic bacteria Rhodobacter capsulatus and Synechocystis sp. PCC 6803 as new hosts for cyclic plant triterpene biosynthesis. PLoS ONE 2017, 12, e0189816. [Google Scholar] [CrossRef]
- Oliver, J.W.; Atsumi, S. Metabolic design for cyanobacterial chemical synthesis. Photosynth. Res. 2014, 120, 249–261. [Google Scholar] [CrossRef]
- Cao, Y.Q.; Li, Q.; Xia, P.F.; Wei, L.J.; Guo, N.; Li, J.W.; Wang, S.G. AraBAD Based Toolkit for Gene Expression and Metabolic Robustness Improvement in Synechococcus elongatus. Sci. Rep. 2017, 7, 18059. [Google Scholar] [CrossRef]
- Kelly, C.L.; Taylor, G.M.; Hitchcock, A.; Torres-Mendez, A.; Heap, J.T. A Rhamnose-Inducible System for Precise and Temporal Control of Gene Expression in Cyanobacteria. ACS Synth. Biol. 2018, 7, 1056–1066. [Google Scholar] [CrossRef]
- Behle, A.; Saake, P.; Germann, A.T.; Dienst, D.; Axmann, I.M. Comparative Dose-Response Analysis of Inducible Promoters in Cyanobacteria. ACS Synth. Biol. 2020, 9, 843–855. [Google Scholar] [CrossRef]
- Abe, K.; Miyake, K.; Nakamura, M.; Kojima, K.; Ferri, S.; Ikebukuro, K.; Sode, K. Engineering of a green-light inducible gene expression system in Synechocystis sp. PCC6803. Microb. Biotechnol. 2014, 7, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Shono, C.; Ariyanti, D.; Abe, K.; Sakai, Y.; Sakamoto, I.; Tsukakoshi, K.; Sode, K.; Ikebukuro, K. A Green Light-Regulated T7 RNA Polymerase Gene Expression System for Cyanobacteria. Mar. Biotechnol. 2021, 23, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Immethun, C.M.; Ng, K.M.; DeLorenzo, D.M.; Waldron-Feinstein, B.; Lee, Y.C.; Moon, T.S. Oxygen-responsive genetic circuits constructed in Synechocystis sp. PCC 6803. Biotechnol. Bioeng. 2016, 113, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Davy, A.M.; Kildegaard, H.F.; Andersen, M.R. Cell Factory Engineering. Cell Syst. 2017, 4, 262–275. [Google Scholar] [CrossRef] [PubMed]
- Immethun, C.M.; DeLorenzo, D.M.; Focht, C.M.; Gupta, D.; Johnson, C.B.; Moon, T.S. Physical, chemical and metabolic state sensors expand the synthetic biology toolbox for Synechocystis sp. PCC 6803. Biotechnol. Bioeng. 2017, 114, 1561–1569. [Google Scholar] [CrossRef]
- Hirose, Y.; Shimada, T.; Narikawa, R.; Katayama, M.; Ikeuchi, M. Cyanobacteriochrome CcaS is the green light receptor that induces the expression of phycobilisome linker protein. Proc. Natl. Acad. Sci. USA 2008, 105, 9528–9533. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.M.; Korosh, T.C.; Nielsen, D.R.; Pfleger, B.F. Optimization of a T7-RNA polymerase system in Synechococcus sp. PCC 7002 mirrors the protein overproduction phenotype from E. coli BL21(DE3). Appl. Microbiol. Biotechnol. 2021, 105, 1147–1158. [Google Scholar] [CrossRef]
- Azevedo, R.; Lopes, J.L.; de Souza, M.M.; Quirino, B.F.; Cançado, L.J.; Marins, L.F. Synechococcus elongatus as a model of photosynthetic bioreactor for expression of recombinant β-glucosidases. Biotechnol. Biofuels 2019, 12, 174. [Google Scholar] [CrossRef]
- Kang, Y.; Weber, K.D.; Qiu, Y.; Kiley, P.J.; Blattner, F.R. Genome-wide expression analysis indicates that FNR of Escherichia coli K-12 regulates a large number of genes of unknown function. J. Bacteriol. 2005, 187, 1135–1160. [Google Scholar] [CrossRef]
- Moon, T.S.; Lou, C.; Tamsir, A.; Stanton, B.C.; Voigt, C.A. Genetic programs constructed from layered logic gates in single cells. Nature 2012, 491, 249–253. [Google Scholar] [CrossRef]
- Heidorn, T.; Camsund, D.; Huang, H.-H.; Lindberg, P.; Oliveira, P.; Stensjö, K.; Lindblad, P. Chapter Twenty-Four—Synthetic Biology in Cyanobacteria: Engineering and Analyzing Novel Functions. In Methods in Enzymology; Voigt, C., Ed.; Academic Press: Cambridge, MA, USA, 2011; Volume 497, pp. 539–579. [Google Scholar]
- Ma, J.; Campbell, A.; Karlin, S. Correlations between Shine-Dalgarno sequences and gene features such as predicted expression levels and operon structures. J. Bacteriol. 2002, 184, 5733–5745. [Google Scholar] [CrossRef]
- Salis, H.M.; Mirsky, E.A.; Voigt, C.A. Automated design of synthetic ribosome binding sites to control protein expression. Nat. Biotechnol. 2009, 27, 946–950. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Morgan, J.A.; Ungerer, J.; Wang, B.; Maness, P.-C.; Yu, J. The plasticity of cyanobacterial metabolism supports direct CO2 conversion to ethylene. Nat. Plants 2015, 1, 15053. [Google Scholar] [CrossRef]
- Wang, B.; Eckert, C.; Maness, P.C.; Yu, J. A Genetic Toolbox for Modulating the Expression of Heterologous Genes in the Cyanobacterium Synechocystis sp. PCC 6803. ACS Synth. Biol. 2018, 7, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Kosuri, S.; Goodman, D.B.; Cambray, G.; Mutalik, V.K.; Gao, Y.; Arkin, A.P.; Endy, D.; Church, G.M. Composability of regulatory sequences controlling transcription and translation in Escherichia coli. Proc. Natl. Acad. Sci. USA 2013, 110, 14024–14029. [Google Scholar] [CrossRef] [PubMed]
- Clifton, K.P.; Jones, E.M.; Paudel, S.; Marken, J.P.; Monette, C.E.; Halleran, A.D.; Epp, L.; Saha, M.S. The genetic insulator RiboJ increases expression of insulated genes. J. Biol. Eng. 2018, 12, 23. [Google Scholar] [CrossRef]
- Vlkova, M.; Morampalli, B.R.; Silander, O.K. Efficiency of the synthetic self-splicing RiboJ ribozyme is robust to cis- and trans-changes in genetic background. Microbiologyopen 2021, 10, e1232. [Google Scholar] [CrossRef]
- Buzayan, J.M.; Gerlach, W.L.; Bruening, G. Satellite tobacco ringspot virus RNA: A subset of the RNA sequence is sufficient for autolytic processing. Proc. Natl. Acad. Sci. USA 1986, 83, 8859–8862. [Google Scholar] [CrossRef]
- Lou, C.; Stanton, B.; Chen, Y.J.; Munsky, B.; Voigt, C.A. Ribozyme-based insulator parts buffer synthetic circuits from genetic context. Nat. Biotechnol. 2012, 30, 1137–1142. [Google Scholar] [CrossRef]
- Yu, H.; Wang, Z.; Xu, H.; Guo, J.; Ma, Q.; Mu, X.; Luo, Y. A method for Absolute Protein Expression Quantity Measurement Employing Insulator RiboJ. Engineering 2018, 4, 881–887. [Google Scholar] [CrossRef]
- Liu, X.; Miao, R.; Lindberg, P.; Lindblad, P. Modular engineering for efficient photosynthetic biosynthesis of 1-butanol from CO2 in cyanobacteria. Energy Environ. Sci. 2019, 12, 2765–2777. [Google Scholar] [CrossRef]
- Mutalik, V.K.; Guimaraes, J.C.; Cambray, G.; Lam, C.; Christoffersen, M.J.; Mai, Q.A.; Tran, A.B.; Paull, M.; Keasling, J.D.; Arkin, A.P.; et al. Precise and reliable gene expression via standard transcription and translation initiation elements. Nat. Methods 2013, 10, 354–360. [Google Scholar] [CrossRef]
- Englund, E.; Shabestary, K.; Hudson, E.P.; Lindberg, P. Systematic overexpression study to find target enzymes enhancing production of terpenes in Synechocystis PCC 6803, using isoprene as a model compound. Metab. Eng. 2018, 49, 164–177. [Google Scholar] [CrossRef]
- Wegelius, A.; Khanna, N.; Esmieu, C.; Barone, G.D.; Pinto, F.; Tamagnini, P.; Berggren, G.; Lindblad, P. Generation of a functional, semisynthetic [FeFe]-hydrogenase in a photosynthetic microorganism. Energy Environ. Sci. 2018, 11, 3163–3167. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Cengic, I.; Cañadas, I.C.; Minton, N.P.; Hudson, E.P. Inducible CRISPR/Cas9 Allows for Multiplexed and Rapidly Segregated Single-Target Genome Editing in Synechocystis Sp. PCC 6803. ACS Synth. Biol. 2022, 11, 3100–3113. [Google Scholar] [CrossRef]
- Wang, S.-Y.; Li, X.; Wang, S.-G.; Xia, P.-F. Base editing for reprogramming cyanobacterium Synechococcus elongatus. Metab. Eng. 2023, 75, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Gordon, G.C.; Korosh, T.C.; Cameron, J.C.; Markley, A.L.; Begemann, M.B.; Pfleger, B.F. CRISPR interference as a titratable, trans-acting regulatory tool for metabolic engineering in the cyanobacterium Synechococcus sp. strain PCC 7002. Metab. Eng. 2016, 38, 170–179. [Google Scholar] [CrossRef]
- Yao, L.; Shabestary, K.; Bjork, S.M.; Asplund-Samuelsson, J.; Joensson, H.N.; Jahn, M.; Hudson, E.P. Pooled CRISPRi screening of the cyanobacterium Synechocystis sp PCC 6803 for enhanced industrial phenotypes. Nat. Commun. 2020, 11, 1666. [Google Scholar] [CrossRef]
- Lai, M.J.; Tsai, J.C.; Lan, E.I. CRISPRi-enhanced direct photosynthetic conversion of carbon dioxide to succinic acid by metabolically engineered cyanobacteria. Bioresour. Technol. 2022, 366, 128131. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarzyk, D.; Cengic, I.; Yao, L.; Hudson, E.P. Diversion of the long-chain acyl-ACP pool in Synechocystis to fatty alcohols through CRISPRi repression of the essential phosphate acyltransferase PlsX. Metab. Eng. 2018, 45, 59–66. [Google Scholar] [CrossRef]
- Knoot, C.J.; Biswas, S.; Pakrasi, H.B. Tunable Repression of Key Photosynthetic Processes Using Cas12a CRISPR Interference in the Fast-Growing Cyanobacterium Synechococcus sp. UTEX 2973. ACS Synth. Biol. 2020, 9, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Woo, H.M. A Logic NAND Gate for Controlling Gene Expression in a Circadian Rhythm in Cyanobacteria. ACS Synth. Biol. 2020, 9, 3210–3216. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.E.; Golden, S.S. Circadian Rhythms in Cyanobacteria. Microbiol. Mol. Biol. Rev. 2015, 79, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Roth, A.; Breaker, R.R. The structural and functional diversity of metabolite-binding riboswitches. Annu. Rev. Biochem. 2009, 78, 305–334. [Google Scholar] [CrossRef]
- Apura, P.; Domingues, S.; Viegas, S.C.; Arraiano, C.M. Reprogramming bacteria with RNA regulators. Biochem. Soc. Trans. 2019, 47, 1279–1289. [Google Scholar] [CrossRef]
- Winkler, W.; Nahvi, A.; Breaker, R.R. Thiamine derivatives bind messenger RNAs directly to regulate bacterial gene expression. Nature 2002, 419, 952–956. [Google Scholar] [CrossRef]
- Winkler, W.C.; Cohen-Chalamish, S.; Breaker, R.R. An mRNA structure that controls gene expression by binding FMN. Proc. Natl. Acad. Sci. USA 2002, 99, 15908–15913. [Google Scholar] [CrossRef]
- Garst, A.D.; Edwards, A.L.; Batey, R.T. Riboswitches: Structures and mechanisms. Cold Spring Harb. Perspect. Biol. 2011, 3, a003533. [Google Scholar] [CrossRef]
- Serganov, A.; Nudler, E. A decade of riboswitches. Cell 2013, 152, 17–24. [Google Scholar] [CrossRef]
- Nakahira, Y.; Ogawa, A.; Asano, H.; Oyama, T.; Tozawa, Y. Theophylline-dependent riboswitch as a novel genetic tool for strict regulation of protein expression in Cyanobacterium Synechococcus elongatus PCC 7942. Plant Cell Physiol. 2013, 54, 1724–1735. [Google Scholar] [CrossRef]
- Ma, A.T.; Schmidt, C.M.; Golden, J.W. Regulation of gene expression in diverse cyanobacterial species by using theophylline-responsive riboswitches. Appl. Environ. Microbiol. 2014, 80, 6704–6713. [Google Scholar] [CrossRef] [PubMed]
- Ohbayashi, R.; Akai, H.; Yoshikawa, H.; Hess, W.R.; Watanabe, S. A tightly inducible riboswitch system in Synechocystis sp. PCC 6803. J. Gen. Appl. Microbiol. 2016, 62, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Higo, A.; Isu, A.; Fukaya, Y.; Hisabori, T. Spatio-Temporal Gene Induction Systems in the Heterocyst-Forming Multicellular Cyanobacterium Anabaena sp. PCC 7120. Plant Cell Physiol. 2018, 59, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Perez, A.A.; Liu, Z.; Rodionov, D.A.; Li, Z.; Bryant, D.A. Complementation of Cobalamin Auxotrophy in Synechococcus sp. Strain PCC 7002 and Validation of a Putative Cobalamin Riboswitch In Vivo. J. Bacteriol. 2016, 198, 2743–2752. [Google Scholar] [CrossRef]
- Klahn, S.; Bolay, P.; Wright, P.R.; Atilho, R.M.; Brewer, K.I.; Hagemann, M.; Breaker, R.R.; Hess, W.R. A glutamine riboswitch is a key element for the regulation of glutamine synthetase in cyanobacteria. Nucleic Acids Res. 2018, 46, 10082–10094. [Google Scholar] [CrossRef]
- Abe, K.; Sakai, Y.; Nakashima, S.; Araki, M.; Yoshida, W.; Sode, K.; Ikebukuro, K. Design of riboregulators for control of cyanobacterial (Synechocystis) protein expression. Biotechnol. Lett. 2014, 36, 287–294. [Google Scholar] [CrossRef]
- Sakamoto, I.; Abe, K.; Kawai, S.; Tsukakoshi, K.; Sakai, Y.; Sode, K.; Ikebukuro, K. Improving the induction fold of riboregulators for cyanobacteria. RNA Biol. 2018, 15, 353–358. [Google Scholar] [CrossRef]
- Sakai, Y.; Abe, K.; Nakashima, S.; Ellinger, J.J.; Ferri, S.; Sode, K.; Ikebukuro, K. Scaffold-fused riboregulators for enhanced gene activation in Synechocystis sp. PCC 6803. Microbiologyopen 2015, 4, 533–540. [Google Scholar] [CrossRef]
- Ueno, K.; Sakai, Y.; Shono, C.; Sakamoto, I.; Tsukakoshi, K.; Hihara, Y.; Sode, K.; Ikebukuro, K. Applying a riboregulator as a new chromosomal gene regulation tool for higher glycogen production in Synechocystis sp. PCC 6803. Appl. Microbiol. Biotechnol. 2017, 101, 8465–8474. [Google Scholar] [CrossRef]
- To, A.C.; Chu, D.H.; Wang, A.R.; Li, F.C.; Chiu, A.W.; Gao, D.Y.; Choi, C.H.J.; Kong, S.K.; Chan, T.F.; Chan, K.M.; et al. A comprehensive web tool for toehold switch design. Bioinformatics 2018, 34, 2862–2864. [Google Scholar] [CrossRef]
- Chappell, J.; Westbrook, A.; Verosloff, M.; Lucks, J.B. Computational design of small transcription activating RNAs for versatile and dynamic gene regulation. Nat. Commun. 2017, 8, 1051. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.R.; Prasad, S. Metabolic engineering of bacteria. Indian J. Microbiol. 2011, 51, 403–409. [Google Scholar] [CrossRef]
- Mund, N.K.; Liu, Y.; Chen, S. Advances in metabolic engineering of cyanobacteria for production of biofuels. Fuel 2022, 322, 124117. [Google Scholar] [CrossRef]
- Aikawa, S.; Joseph, A.; Yamada, R.; Izumi, Y.; Yamagishi, T.; Matsuda, F.; Kawai, H.; Chang, J.-S.; Hasunuma, T.; Kondo, A. Direct conversion of Spirulina to ethanol without pretreatment or enzymatic hydrolysis processes. Energy Environ. Sci. 2013, 6, 1844–1849. [Google Scholar] [CrossRef]
- Harun, R.; Jason, W.S.Y.; Cherrington, T.; Danquah, M.K. Exploring alkaline pre-treatment of microalgal biomass for bioethanol production. Appl. Energy 2011, 88, 3464–3467. [Google Scholar] [CrossRef]
- Vavitsas, K.; Fabris, M.; Vickers, C.E. Terpenoid Metabolic Engineering in Photosynthetic Microorganisms. Genes 2018, 9, 520. [Google Scholar] [CrossRef]
- Satta, A.; Esquirol, L.; Ebert, B.E.; Newman, J.; Peat, T.S.; Plan, M.; Schenk, G.; Vickers, C.E. Molecular characterization of cyanobacterial short-chain prenyltransferases and discovery of a novel GGPP phosphatase. FEBS J. 2022, 289, 6672–6693. [Google Scholar] [CrossRef]
- Krishnan, A.; McNeil, B.A.; Stuart, D.T. Biosynthesis of Fatty Alcohols in Engineered Microbial Cell Factories: Advances and Limitations. Front. Bioeng. Biotechnol. 2020, 8, 610936. [Google Scholar] [CrossRef] [PubMed]
- Slocombe, S.P.; Zhang, Q.; Ross, M.; Anderson, A.; Thomas, N.J.; Lapresa, A.; Rad-Menendez, C.; Campbell, C.N.; Black, K.D.; Stanley, M.S.; et al. Unlocking nature’s treasure-chest: Screening for oleaginous algae. Sci. Rep. 2015, 5, 9844. [Google Scholar] [CrossRef]
- Liu, X.; Sheng, J.; Curtiss, R., 3rd. Fatty acid production in genetically modified cyanobacteria. Proc. Natl. Acad. Sci. USA 2011, 108, 6899–6904. [Google Scholar] [CrossRef]
- Kato, A.; Takatani, N.; Ikeda, K.; Maeda, S.I.; Omata, T. Removal of the product from the culture medium strongly enhances free fatty acid production by genetically engineered Synechococcus elongatus. Biotechnol. Biofuels 2017, 10, 141. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Qi, F.; Tan, X.; Lu, X. Improved production of fatty alcohols in cyanobacteria by metabolic engineering. Biotechnol. Biofuels 2014, 7, 94. [Google Scholar] [CrossRef] [PubMed]
- Santos-Merino, M.; Garcillan-Barcia, M.P.; de la Cruz, F. Engineering the fatty acid synthesis pathway in Synechococcus elongatus PCC 7942 improves omega-3 fatty acid production. Biotechnol. Biofuels 2018, 11, 239. [Google Scholar] [CrossRef] [PubMed]
- Poole, L.B.; Parsonage, D.; Sergeant, S.; Miller, L.R.; Lee, J.; Furdui, C.M.; Chilton, F.H. Acyl-lipid desaturases and Vipp1 cooperate in cyanobacteria to produce novel omega-3 PUFA-containing glycolipids. Biotechnol. Biofuels 2020, 13, 83. [Google Scholar] [CrossRef] [PubMed]
- Ducat, D.C.; Avelar-Rivas, J.A.; Way, J.C.; Silver, P.A. Rerouting carbon flux to enhance photosynthetic productivity. Appl. Environ. Microbiol. 2012, 78, 2660–2668. [Google Scholar] [CrossRef]
- Lin, P.C.; Zhang, F.; Pakrasi, H.B. Enhanced production of sucrose in the fast-growing cyanobacterium Synechococcus elongatus UTEX 2973. Sci. Rep. 2020, 10, 390. [Google Scholar] [CrossRef]
- Qiao, Y.; Wang, W.; Lu, X. Engineering cyanobacteria as cell factories for direct trehalose production from CO(2). Metab. Eng. 2020, 62, 161–171. [Google Scholar] [CrossRef]
- Mota, R.; Guimaraes, R.; Buttel, Z.; Rossi, F.; Colica, G.; Silva, C.J.; Santos, C.; Gales, L.; Zille, A.; De Philippis, R.; et al. Production and characterization of extracellular carbohydrate polymer from Cyanothece sp. CCY 0110. Carbohydr. Polym. 2013, 92, 1408–1415. [Google Scholar] [CrossRef]
- Mota, R.; Vidal, R.; Pandeirada, C.; Flores, C.; Adessi, A.; De Philippis, R.; Nunes, C.; Coimbra, M.A.; Tamagnini, P. Cyanoflan: A cyanobacterial sulfated carbohydrate polymer with emulsifying properties. Carbohydr. Polym. 2020, 229, 115525. [Google Scholar] [CrossRef]
- Li, T.; Li, C.T.; Butler, K.; Hays, S.G.; Guarnieri, M.T.; Oyler, G.A.; Betenbaugh, M.J. Mimicking lichens: Incorporation of yeast strains together with sucrose-secreting cyanobacteria improves survival, growth, ROS removal and lipid production in a stable mutualistic co-culture production platform. Biotechnol. Biofuels 2017, 10, 55. [Google Scholar] [CrossRef]
- Chaves, J.E.; Melis, A. Biotechnology of cyanobacterial isoprene production. Appl. Microbiol. Biotechnol. 2018, 102, 6451–6458. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.C.; Zhang, F.; Pakrasi, H.B. Enhanced limonene production in a fast-growing cyanobacterium through combinatorial metabolic engineering. Metab Eng. Commun. 2021, 12, e00164. [Google Scholar] [CrossRef]
- Lin, P.C.; Saha, R.; Zhang, F.; Pakrasi, H.B. Metabolic engineering of the pentose phosphate pathway for enhanced limonene production in the cyanobacterium Synechocysti s sp. PCC 6803. Sci. Rep. 2017, 7, 17503. [Google Scholar] [CrossRef] [PubMed]
- Halfmann, C.; Gu, L.; Zhou, R. Engineering cyanobacteria for the production of a cyclic hydrocarbon fuel from CO2 and H2O. Green Chem. 2014, 16, 3175–3185. [Google Scholar] [CrossRef]
- Davies, F.K.; Work, V.H.; Beliaev, A.S.; Posewitz, M.C. Engineering Limonene and Bisabolene Production in Wild Type and a Glycogen-Deficient Mutant of Synechococcus sp. PCC 7002. Front. Bioeng. Biotechnol. 2014, 2, 21. [Google Scholar] [CrossRef] [PubMed]
- Matsudaira, A.; Hoshino, Y.; Uesaka, K.; Takatani, N.; Omata, T.; Usuda, Y. Production of glutamate and stereospecific flavors, (S)-linalool and (+)-valencene, by Synechocystis sp. PCC6803. J. Biosci. Bioeng. 2020, 130, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Formighieri, C.; Melis, A. Sustainable heterologous production of terpene hydrocarbons in cyanobacteria. Photosynth. Res. 2016, 130, 123–135. [Google Scholar] [CrossRef]
- Rodrigues, J.S.; Lindberg, P. Metabolic engineering of Synechocystis sp. PCC 6803 for improved bisabolene production. Metab Eng. Commun. 2021, 12, e00159. [Google Scholar] [CrossRef]
- Halfmann, C.; Gu, L.; Gibbons, W.; Zhou, R. Genetically engineering cyanobacteria to convert CO(2), water and light into the long-chain hydrocarbon farnesene. Appl. Microbiol. Biotechnol. 2014, 98, 9869–9877. [Google Scholar] [CrossRef]
- Pattharaprachayakul, N.; Lee, H.J.; Incharoensakdi, A.; Woo, H.M. Evolutionary Engineering of Cyanobacteria to Enhance the Production of alpha-Farnesene from CO(2). J. Agric. Food Chem. 2019, 67, 13658–13664. [Google Scholar] [CrossRef]
- Choi, S.Y.; Lee, H.J.; Choi, J.; Kim, J.; Sim, S.J.; Um, Y.; Kim, Y.; Lee, T.S.; Keasling, J.D.; Woo, H.M. Photosynthetic conversion of CO2 to farnesyl diphosphate-derived phytochemicals (amorpha-4,11-diene and squalene) by engineered cyanobacteria. Biotechnol. Biofuels 2016, 9, 202. [Google Scholar] [CrossRef] [PubMed]
- Reinsvold, R.E.; Jinkerson, R.E.; Radakovits, R.; Posewitz, M.C.; Basu, C. The production of the sesquiterpene beta-caryophyllene in a transgenic strain of the cyanobacterium Synechocystis. J. Plant Physiol. 2011, 168, 848–852. [Google Scholar] [CrossRef] [PubMed]
- Dienst, D.; Wichmann, J.; Mantovani, O.; Rodrigues, J.S.; Lindberg, P. High density cultivation for efficient sesquiterpenoid biosynthesis in Synechocystis sp. PCC 6803. Sci. Rep. 2020, 10, 5932. [Google Scholar] [CrossRef]
- Vavitsas, K.; Rue, E.O.; Stefansdottir, L.K.; Gnanasekaran, T.; Blennow, A.; Crocoll, C.; Gudmundsson, S.; Jensen, P.E. Responses of Synechocystis sp. PCC 6803 to heterologous biosynthetic pathways. Microb. Cell Fact. 2017, 16, 140. [Google Scholar] [CrossRef]
- Formighieri, C.; Melis, A. Heterologous synthesis of geranyllinalool, a diterpenol plant product, in the cyanobacterium Synechocystis. Appl. Microbiol. Biotechnol. 2017, 101, 2791–2800. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Woo, H.M. CRISPRi-dCas12a: A dCas12a-Mediated CRISPR Interference for Repression of Multiple Genes and Metabolic Engineering in Cyanobacteria. ACS Synth. Biol. 2020, 9, 2351–2361. [Google Scholar] [CrossRef] [PubMed]
- Pattanaik, B.; Englund, E.; Nolte, N.; Lindberg, P. Introduction of a green algal squalene synthase enhances squalene accumulation in a strain of Synechocystis sp. PCC 6803. Metab Eng. Commun. 2020, 10, e00125. [Google Scholar] [CrossRef]
- Taylor, G.M.; Hitchcock, A.; Heap, J.T. Combinatorial assembly platform enabling engineering of genetically stable metabolic pathways in cyanobacteria. Nucleic Acids Res. 2021, 49, e123. [Google Scholar] [CrossRef]
- Englund, E.; Pattanaik, B.; Ubhayasekera, S.J.; Stensjo, K.; Bergquist, J.; Lindberg, P. Production of squalene in Synechocystis sp. PCC 6803. PLoS ONE 2014, 9, e90270. [Google Scholar] [CrossRef]
- Ershov, Y.V.; Gantt, R.R.; Cunningham, F.X., Jr.; Gantt, E. Isoprenoid biosynthesis in Synechocystis sp. strain PCC6803 is stimulated by compounds of the pentose phosphate cycle but not by pyruvate or deoxyxylulose-5-phosphate. J. Bacteriol. 2002, 184, 5045–5051. [Google Scholar] [CrossRef]
- Poliquin, K.; Ershov, Y.V.; Cunningham, F.X., Jr.; Woreta, T.T.; Gantt, R.R.; Gantt, E. Inactivation of sll1556 in Synechocystis strain PCC 6803 impairs isoprenoid biosynthesis from pentose phosphate cycle substrates in vitro. J. Bacteriol. 2004, 186, 4685–4693. [Google Scholar] [CrossRef]
- Bentley, F.K.; Zurbriggen, A.; Melis, A. Heterologous expression of the mevalonic acid pathway in cyanobacteria enhances endogenous carbon partitioning to isoprene. Mol. Plant 2014, 7, 71–86. [Google Scholar] [CrossRef]
- Kerfeld, C.A.; Aussignargues, C.; Zarzycki, J.; Cai, F.; Sutter, M. Bacterial microcompartments. Nat. Rev. Microbiol. 2018, 16, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Murmu, G.; Mukherjee, K.; Saha, S.; Maity, D. A comprehensive overview of nanotechnology in sustainable agriculture. J. Biotechnol. 2022, 355, 21–41. [Google Scholar] [CrossRef]
- Jiang, M.; Song, Y.; Kanwar, M.K.; Ahammed, G.J.; Shao, S.; Zhou, J. Phytonanotechnology applications in modern agriculture. J. Nanobiotechnol. 2021, 19, 430. [Google Scholar] [CrossRef] [PubMed]
- Niklowitz, W.; Drews, G. Beiträge zur Cytologie der Blaualgen. Arch. Für Mikrobiol. 1957, 27, 150–165. [Google Scholar] [CrossRef]
- Occhialini, A.; Lin, M.T.; Andralojc, P.J.; Hanson, M.R.; Parry, M.A. Transgenic tobacco plants with improved cyanobacterial Rubisco expression but no extra assembly factors grow at near wild-type rates if provided with elevated CO2. Plant J. 2016, 85, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.R.; Lin, M.T.; Carmo-Silva, A.E.; Parry, M.A. Towards engineering carboxysomes into C3 plants. Plant J. 2016, 87, 38–50. [Google Scholar] [CrossRef]
- Fang, Y.; Huang, F.; Faulkner, M.; Jiang, Q.; Dykes, G.F.; Yang, M.; Liu, L.N. Engineering and Modulating Functional Cyanobacterial CO(2)-Fixing Organelles. Front. Plant Sci. 2018, 9, 739. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yan, X.; Aigner, H.; Bracher, A.; Nguyen, N.D.; Hee, W.Y.; Long, B.M.; Price, G.D.; Hartl, F.U.; Hayer-Hartl, M. Rubisco condensate formation by CcmM in beta-carboxysome biogenesis. Nature 2019, 566, 131–135. [Google Scholar] [CrossRef]
- Sutter, M.; Boehringer, D.; Gutmann, S.; Gunther, S.; Prangishvili, D.; Loessner, M.J.; Stetter, K.O.; Weber-Ban, E.; Ban, N. Structural basis of enzyme encapsulation into a bacterial nanocompartment. Nat. Struct. Mol. Biol. 2008, 15, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Tracey, J.C.; Coronado, M.; Giessen, T.W.; Lau, M.C.Y.; Silver, P.A.; Ward, B.B. The Discovery of Twenty-Eight New Encapsulin Sequences, Including Three in Anammox Bacteria. Sci. Rep. 2019, 9, 20122. [Google Scholar] [CrossRef] [PubMed]
- Nichols, R.J.; LaFrance, B.; Phillips, N.R.; Radford, D.R.; Oltrogge, L.M.; Valentin-Alvarado, L.E.; Bischoff, A.J.; Nogales, E.; Savage, D.F. Discovery and characterization of a novel family of prokaryotic nanocompartments involved in sulfur metabolism. Elife 2021, 10, e59288. [Google Scholar] [CrossRef] [PubMed]
- Hays, S.G.; Yan, L.L.W.; Silver, P.A.; Ducat, D.C. Synthetic photosynthetic consortia define interactions leading to robustness and photoproduction. J. Biol. Eng. 2017, 11, 4. [Google Scholar] [CrossRef]
- Arias, D.M.; Garcia, J.; Uggetti, E. Production of polymers by cyanobacteria grown in wastewater: Current status, challenges and future perspectives. N. Biotechnol. 2020, 55, 46–57. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Satta, A.; Esquirol, L.; Ebert, B.E. Current Metabolic Engineering Strategies for Photosynthetic Bioproduction in Cyanobacteria. Microorganisms 2023, 11, 455. https://doi.org/10.3390/microorganisms11020455
Satta A, Esquirol L, Ebert BE. Current Metabolic Engineering Strategies for Photosynthetic Bioproduction in Cyanobacteria. Microorganisms. 2023; 11(2):455. https://doi.org/10.3390/microorganisms11020455
Chicago/Turabian StyleSatta, Alessandro, Lygie Esquirol, and Birgitta E. Ebert. 2023. "Current Metabolic Engineering Strategies for Photosynthetic Bioproduction in Cyanobacteria" Microorganisms 11, no. 2: 455. https://doi.org/10.3390/microorganisms11020455
APA StyleSatta, A., Esquirol, L., & Ebert, B. E. (2023). Current Metabolic Engineering Strategies for Photosynthetic Bioproduction in Cyanobacteria. Microorganisms, 11(2), 455. https://doi.org/10.3390/microorganisms11020455