Abstract
Tick-borne relapsing fever group (RFG) borreliosis remains neglected as a human disease and little is known on its maintenance in ticks and vertebrates, especially in South America. Therefore, this study investigated borrelial infection in Ornithodoros ticks collected in rodent-inhabited rock formations in the Brazilian semiarid region, within the Caatinga biome. Collected ticks (Ornithodoros rietcorreai and Ornithodoros cf. tabajara) were allowed to feed under laboratory conditions on guinea pigs, which had blood samples examined daily by dark-field microscopy. No spirochetes were visualized in the blood of any of four O. rietcorreai-infested guinea pigs. Contrastingly, spirochetes were visualized between 9 and 39 days after tick feeding in the blood of three guinea pigs, each infested with O. cf. tabajara ticks from a different locality. Guinea pig infection was confirmed by passages into experimental animals and by generating DNA sequences of Borrelia spp. from the blood of spirochetemic guinea pigs. Three O. cf. tabajara populations were infected by the same borrelial organism, which was characterized as a novel RFG agent (named as ‘Candidatus Borrelia caatinga’) based on 10 Borrelia loci (rrs, flaB, glpQ, gyrB, clpX, pepX, pyrG, recG, rplB and uvrA). We demonstrated that O. cf. tabajara is a competent vector of the novel Borrelia sp. isolates, although none of the infected rodents developed clinical illness.
1. Introduction
The spirochete genus Borrelia constitute bacteria that infect vertebrates, to whom they are transmitted by hematophagous vectors. With the exception of Borrelia recurrentis, transmitted by the human clothing louse (Pediculus humanus humanus), all Borrelia species are primarily transmitted by ticks [1,2]. Species of the genus Borrelia are known to infect a variety of vertebrate hosts (mammals, birds, reptiles) and have been divided into three main groups: (i) the Lyme group (LG), represented by Borrelia burgdorferi sensu lato genospecies that are associated with hard ticks (Ixodidae family); (ii) the Reptile-Echidna group (REPG), represented by organisms associated with hard ticks; and (iii) the Relapsing Fever group (RFG), which contains numerous organisms mainly associated with soft ticks (Argasidae family) and a few ones associated with hard ticks, in addition to the louse-borne B. recurrentis [1,2]. Although there has been a recent proposal to split borrelial species into two genera (i.e., Borrelia for RFG species, and Borreliella for LG species) [3,4], this proposal is still controversial; hence, herein we opted to consider all borrelial species as belonging to the genus Borrelia, as recently discussed [5,6].
Most RFG borreliae are primarily associated with soft ticks of the genus Ornithodoros, in which the bacteria perpetuate through transstadial and transovarial passages [7]. Given the great capacity of Ornithodoros ticks to survive for several years without feeding, these arthropods are also pointed out as main reservoirs of RFG borreliae [7]. Once transmitted to vertebrates upon tick feeding, RFG borreliae replicate in the blood of competent hosts, which might suffer clinical illness. In humans, this condition is reported as ‘tick-borne relapsing fever’ [2,7]. RFG borreliae are maintained in enzootic cycles, mostly between soft ticks (Ornithodoros spp.) and rodents [7,8]. Once infected, rodents develop an initial peak of spirochetemia that lasts for a few days, followed by new spirochete relapses interspaced by a few days during an infection course of usually one month, when signs of illness might or might not be present [9,10].
Among more than 20 species of RFG borreliae described in different continents of the world, most are pathogenic for humans and associated with Ornithodoros ticks [8]. Although tick-borne relapsing fever was first reported during the 19th century, it remains neglected as a human disease and little is known on its maintenance in ticks and vertebrates [7,8]. In South America, only two Ornithodoros-associated Borrelia species have been described, Borrelia brasiliensis and Borrelia venezuelensis; the latter was associated with clinical cases of human relapsing fever in Colombia and Venezuela during the first half of the 20th century [11].
The first report of RFG Borrelia associated to Ornithodoros ticks from Brazil was performed by Davis [12], who observed spirochetes in the blood of mice that were infested with Ornithodoros brasiliensis ticks from Rio Grande do Sul state. Although the spirochetes were named as B. brasiliensis, the isolate was lost and never reported again. During this century, Muñoz-Leal et al. [13] isolated B. venezuelensis by feeding Ornithodoros rudis from Maranhão state on Vesper mice (Calomys callosus). Subsequently, this isolate (designated as B. venezuelensis RMA01) was cultured in vitro and its genome sequenced [14]. Borrelia venezuelensis RMA01 constitutes to date the sole isolate of a RFG borreliae transmitted by an Ornithodoros tick in South America [11]. In a recent study, Muñoz-Leal et al. [15] reported by molecular methods four novel RFG Borrelia genotypes in human-biting Ornithodoros ticks from Brazil: Borrelia sp. Omi2MT and Omi3MT in Ornithodoros mimon from Mato Grosso state, Borrelia sp. JericoCE in Ornithodoros hasei, Borrelia sp. OrietCE in Ornithodoros rietcorreai, and Borrelia sp. TabajaraCE in Ornithodoros tabajara; the latter three tick species were collected in Ceará state, within the Caatinga semiarid biome of Brazil. Despite these findings, human infection by RFG borreliae in Brazil remains unreported [16].
Based on the recent advances on the occurrence of RFG borreliae associated to Ornithodoros ticks in Brazil, especially in the Caatinga biome, the present study aimed to isolate borrelial organisms from Ornithodoros ticks collected in additional areas of this ecosystem. For this purpose, collected Ornithodoros ticks were allowed to feed on guinea pigs, which were monitored for successful borrelial isolation, and subsequent molecular characterization.
2. Materials and Methods
2.1. Study Sites and Collection of Ornithodoros Ticks
During a field expedition in October 2019, ticks were collected from the environment in four localities in the state of Pernambuco, northeastern Brazil: (i) Catimbau National Park, Serra das Torres, Buíque municipality (08°34′0.3″ S, 37°14′27.8″ W; elevation 777 m); (ii) Malhada Vermelha, Floresta municipality (08°36′44.2″ S, 38°32′29.9″ W; 377 m); (iii) Negreiros National Forest, Serrita municipality (07°59′22.0″ S, 39°24′46.1″ W; 475 m); and (iv) Capim District, Petrolina municipality (09°09′40.3″ S, 40°26′16.2″ W; 454 m) (Figure 1). The four localities are located within the Caatinga biome, which is characterized by a semiarid climate (temperatures averaging 27 °C throughout the year, mean annual rainfall typically <500 mm), and deciduous vegetation composed typically of xeric shrub land and thorn forest that consist primarily of small, thorny trees that shed their leaves seasonally [17]. The prospected environments included rock formations with vestiges (e.g., feces) of wild rodents, including visualizations of the rock cavy Kerodon rupestris (Rodentia: Caviidae). Ornithodoros ticks were collected with tweezers from under the rocks and stored in punctured plastic vials, which were kept in an improvised environmental chamber (1.5 L-plastic bottle with a piece of humid cotton on the bottom) until their arrival at the laboratory, where ticks were placed in an incubator at 26 °C and 80% relative humidity. Under this condition, field-collected engorged females oviposited fertile eggs that resulted in hatched larvae.
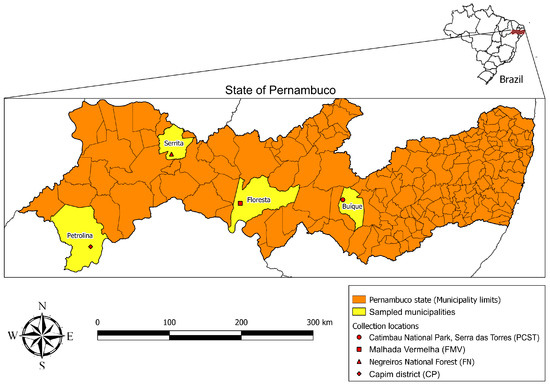
Figure 1.
Locations in the state of Pernambuco, northeastern Brazil, where Ornithodoros ticks were collected.
2.2. Taxonomic Identification of Ornithodoros Ticks
Unfed larvae born in the laboratory were killed in hot water, clarified with 25% KOH, and mounted on slides using Hoyer’s medium to observe morphological characters by optical microscopy (Olympus BX40 optical microscope, Olympus Optical Co., Ltd., Tokyo, Japan). Living adults and nymphs were visualized and counted under a stereomicroscope (Zeiss Stemi SV 11, Zeiss, Münich, Germany). Species were determined according to taxonomic keys [18] and original descriptions [19,20].
Identification of the ticks was complemented by molecular analysis. For this purpose, two or three adult specimens of each species were individually submitted to DNA extraction by the guanidine isothiocyanate and phenol/chloroform technique [21]. A PCR protocol targeting a ≈460 bp fragment of the tick mitochondrial 16S rRNA gene was performed as described [22]. A second PCR protocol targeting a ≈270 bp fragment of the nuclear Histone 3 (H3) gene was performed as described [23]. Amplicons of the expected size were prepared for sequencing using Big Dye Terminator Cycle Sequencing kit (Applied Biosystems, Foster City, CA, USA), and sequenced in an ABI automated sequencer (Applied Biosystems/Thermo Fisher Scientific, model ABI 3500 Genetic Analyzer, Foster City, CA, USA) with the same primers used for PCR. Obtained sequences were assembled, and primer-trimmed with Geneious R9 [24], and submitted to a BLAST analysis (www.ncbi.nlm.nih.gov/blast, accessed on 1 September 2022) to infer closest identities with congeneric ticks [25].
2.3. Isolation of Spirochetes from Ornithodoros Ticks
Attempts to isolate viable spirochetes were performed using field-collected ticks, which were separated in groups according to locations and species. Seven guinea pigs were infested, each one with ticks of one species from a single location. For this purpose, unengorged ticks were released inside a plastic feeding chamber (6 cm diameter) previously glued with a skin compatible-adhesive (Kamar Products, Zionsville, IN, USA) on the shaved dorsum of the guinea pig. Two hours after being released in the feeding chambers, engorged ticks were recovered and placed in the incubator for further studies. A drop of blood (≈2.5µL) was daily obtained from each of the seven guinea pigs by ear vein-puncture, expressed onto glass slides, and observed by dark-field microscopy to detect the presence of motile spirochetes. The mean number of spirochetes per field was calculated by counting the total number of motile spirochetes in 50 microscope fields at 200x magnification, dividing it by 50; results as decimal numbers were always rounded up. Experimental animals not presenting motile spirochetes during the first 21 days were considered negative and were not bled anymore. If a guinea pig showed motile spirochetes during the first 21 days, daily examinations were extended until 52 days after tick infestations.
Spirochetemic guinea pigs were anesthetized (xylazine 5 mg/kg + ketamine 35 mg/kg) and 2 mL of blood was collected by intracardiac puncture at the 18th day after tick infestation. Part of the blood was submitted to DNA extraction (see below) and the other part was intraperitoneally inoculated into five new experimental animals (three newborn guinea pigs, one mouse and one hamster, each one receiving ≈0.300 mL of blood) to perform the first passage of spirochetes into experimental animals in the laboratory. These inoculated animals were also evaluated daily through dark-field microscopy of blood samples, as described above. Two (newborn guinea pigs) of the five new rodents, when showing >10 spirochetes/field, were anesthetized (xylazine + ketamine) and euthanized by exsanguination via the intracardiac route. In this case, the collected blood was immediately put in heparin tubes, centrifuged, and the plasma was aliquoted into 2 mL-cryotubes, which were stored at −80 °C, and then at liquid nitrogen for cryopreservation of the isolated spirochetes. Rectal temperature of all animals was measured daily throughout the study with a digital clinical thermometer.
2.4. Molecular Analyses
DNA extraction of guinea pig blood samples was performed using the DNeasy Blood and Tissue Kit (Qiagen, Valencia, CA, USA), and tested by conventional PCR protocols for amplification of fragments of the borrelial genes 16S ribosomal RNA (rrs), flagellin (flaB), glycerophosphodiester phosphodiesterase (glpQ), and DNA gyrase subunit B (gyrB) (Table 1). In addition, we performed a multilocus sequencing typing (MLST) scheme according to Margos et al. [26] for amplification of the borrelial genes clpA, clpX, pepX, pyrG, recG, rplB, nifS and uvrA (Table S1—Supplementary Materials). DNA of Borrelia anserina strain PB [27] was used as positive control in all reactions. Obtained amplicons were visualized with UV light through 1.5% agarose gels stained with SYBR Safe (Thermo Fisher Scientifific, Waltham, MA, USA). Products containing a single expected size fragment were treated with ExoSAP-IT (USB Corporation, Cleveland, OH, USA) and prepared for sequencing with the BigDye kit (Applied Biosystems, Foster City, CA, USA). An ABI PRISM 3500 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA) was employed for sequencing using the same primers to perform PCRs. Obtained sequences were assembled, trimmed, and translated to amino acid (if applicable) with Geneious R9 [24]. Generated DNA sequences were submitted to BLAST analysis (www.ncbi.nlm.nih.gov/blast, accessed on 1 November 2022) to infer closest identities with other spirochetes [25].

Table 1.
List of the primer pairs used in the study for amplification of four Borrelia genes by conventional PCR assays.
2.5. Phylogenetic Analyses
Obtained consensus sequences and orthologous sequences retrieved from GenBank were aligned with MAFFT using default parameters [31]. Phylogenetics trees were inferred using Maximum likelihood (ML) and Bayesian inference (BI) methods in IQ-TREE v 1.6.12 [32] and MrBayes v 3.2.6 [33], respectively. Protein-coding gene present distinct nucleotide exchange rates (heterogeneity) at the first, second, and third codon positions so datasets were partitioned into three codon positions (position-1, position-2, and position-3) [33,34]. For ML analyses, the best-fit models for non-coding and protein-coding genes datasets were calculated with the ModelFinder commands “TESTNEWONLY -mrate G” and “TESTNEWONLYMERGE -mrate G”, respectively [35]. Trees were run with rapid hill-climbing approach and stochastic disturbance applying 1,000 ultrafast bootstrapsing pseudo-replicates (UFB) to evaluate tree robustness. UFB values < 70%, between 70–94%, and ≥95% were considered non-significant, moderate, and high statistical support, respectively [36].
BI phylogenies were constructed implementing the MrBayes commands “lset nst = mixed rates = gamma” and “lset = mixed rates = invgamma” for non-coding and protein encoding datasets, respectively [33,37,38]. BI analyses were run with two independent tests of 20 × 106 generations, each with four simultaneous Monte Carlo Markov chains (MCMC), sampling trees every 1000 generations, removing the first 25% as burn-in. Tracer software was used to confirm the MCMCs correlation as well as reached stationarity and effective sample size (ESS) [39]. All best-fit substitution models and partitions schemes were selected according to the Bayesian Information Criterion (BIC) [40]. Nodes with Bayesian posterior probabilities (BPP) values ≥ 0.70 were considered of high statistical support [41].
All trees were visualized and edited using FigTree v 1.4.1 (http://tree.bio.ed.ac.uk/software/figtree/, accessed on 1 September 2022) and Inkscape v 1.1 (https://inkscape.org/es/, accessed on 1 September 2022). Congruent topologies between ML and BI analyses were used to produce strict consensus trees in Geneious Prime with the Consensus Tree Builder tool, implementing a support threshold of 100% (www.geneious.com, accessed on 1 September 2022). The consensus phylogram included all monophyletic clades after comparing ML and BI topologies for each dataset.
2.6. Ethics Statement
Field collections of ticks were authorized by Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio permit Sisbio 65137-1). Animal experimentation was approved by the Ethic Committee on Animal Use of the Faculty of Veterinary Medicine of the University of São Paulo (projects number 4425171018 and 2655061218).
3. Results
3.1. Collected Ornithodoros Ticks
A total of 1505 tick specimens were collected in four localities. Morphological analyses resulted in the identification of two Ornithodoros species: 932 specimens of O. rietcorreai from Buíque, Floresta, Serrita and Petrolina, and 573 specimens of O. tabajara from the former three localities. Microscopical analyses of laboratory-reared unfed larvae in mounted slides showed morphological characters corresponding to the species of their respective parental specimens.
Partial sequences of the mitochondrial 16S rRNA gene were generated for two O. rietcorreai specimens; one from Buíque, which was 99% (422/427 bp) identical to O. rietcorreai from Ceará state, Brazil (GenBank MT021433), and the other from Petrolina, which was 99% (425/426 bp) identical to O. rietcorreai from Paraíba state, Brazil (GenBank KX130781). The two O. rietcorreai haplotypes from this study were 96% identical to each other.
Partial sequences of the mitochondrial 16S rRNA gene were generated for three O. tabajara adult specimens, which were 92% (396/429 bp) identical to O. tabajara from Ceará state, Brazil (GenBank MT021434). The sequences of two specimens (one from Buíque and another from Floresta) were 100% identical to each other, whereas the third specimen (from Serrita) generated a second haplotype that differed by a single nucleotide polymorphism (99%; 428/429 bp) from the other two specimens. Although the external morphology of the ticks from Buíque, Floresta and Serrita was compatible with O. tabajara, we are provisionally treating them as Ornithodoros cf. tabajara due to the relatively high polymorphism (8% difference) of their 16S rDNA partial sequences with the type sequence of O. tabajara (MT021434) reported by Muñoz-Leal et al. (2021b). Partial sequences (216 bp) of the nuclear H3 gene of the three specimens of O. cf. tabajara were identical to each other, and by BLAST analysis, they were 99% (174/175 bp) identical to O. tabajara from Ceará (OK247605). Ongoing studies are in progress to elucidate the taxonomic status of the O. cf. tabajara ticks collected in the present study.
3.2. Isolation of Spirochetes from Ornithodoros Ticks
Four guinea pigs (numbers 1, 3, 5, 7) were infested with 925 O. rietcorreai ticks from four localities (25 to 445 ticks per guinea pig), and another three guinea pigs (numbers 2, 4, 6) were infested with 568 O. cf. tabajara ticks from three localities (136 to 289 ticks per guinea pig) (Table 2). Although we did not count the exact number of ticks that were fully engorged two hours after been released in the feeding chambers, nearly all of them became at least partially engorged. No spirochetes were visualized by dark-field microscopy in the blood of any of the four O. rietcorreai-infested guinea pigs (1, 3, 5, 7) during 21 consecutive days after infestation. Spirochetes were visualized in the three guinea pigs (2, 4, 6) that were infested with O. cf. tabajara ticks from three localities (Figure 2) (Video S1—Supplementary Materials).
Table 2.
Results of infestation trials for isolation of spirochetes from Ornithodoros ticks that were collected in four localities of the state of Pernambuco, Caatinga biome, Brazil, during 2019.
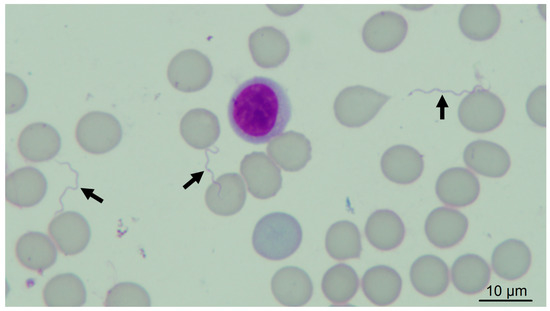
Figure 2.
Giemsa-stained blood smear of guinea pig, showing spirochetes (arrows); original magnification: 1000×.
During the 52-day course of dark-field microscopy monitoring of guinea pig 2 (infested with O. cf. tabajara from Buíque), motile spirochetes were visualized in blood at 9 to 11, 14 to 25, 27 to 30, and 39 days after tick infestation; a mean of ≤1 spirochete/microscope field was visualized generally, although the maximum mean count was five spirochetes/field at day 17 (Figure 3). In guinea pig 4 (infested with O. cf. tabajara from Floresta), motile spirochetes were visualized in blood from 13 to 30 days after tick infestation; while a mean of ≤1 spirochete/field was visualized at most of the times, maximum mean values were 10 spirochetes/field at days 15 and 22 (Figure 3). In guinea pig 6 (infested with O. cf. tabajara from Serrita), motile spirochetes were visualized in blood at 11, 14 to 28, and 30 to 31 days after tick infestation; similarly, a mean of ≤1 spirochete/field was visualized at most of the times, yet a maximum mean value of 30 spirochetes/field was observed at day 23 (Figure 3). The three isolates of spirochetes were recovered from guinea pigs numbers 2, 4, and 6 infested with O. cf. tabajara ticks, and were named Buíque-PCST, Floresta-FMV, and Serrita-FN, respectively.
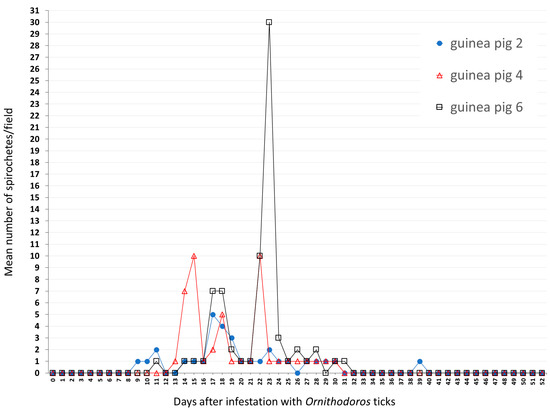
Figure 3.
Results of dark-field examination of blood samples of guinea pigs according to the number of days after infestation with Ornithodoros cf. tabajara from Buíque (guinea pig 2), Floresta (guinea pig 4) and Serrita (guinea pig 6). Values presented as the mean number of motile spirochetes per microscope field at 200× magnification in each sampled day.
Guinea pigs 2, 4, and 6 were bled at the 18th day after tick infestation (when the mean numbers of spirochetes/field were 4, 5, and 7, respectively), and their blood samples were inoculated into other animals to perform the first experimental animal passage of the spirochetes. In this case, the blood of guinea pig 2 (isolate Buíque-PCST) was inoculated into a newborn guinea pig and a mouse (Table 2). Blood samples of this newborn guinea pig showed motile spirochetes (mean: ≤1 to 15 spirochetes/field) from the 4th to the 10th day after inoculation (Figure 4), when it was euthanized by exsanguination and its plasma cryopreserved. Dark-field microscopy of the inoculated mouse revealed spirochetes only at the 7th, 8th, and 23th days after inoculation (mean ≤1 spirochete/field), despite of this experimental animal being daily examined until the 41st day (Figure 4). The blood of guinea pig 4 (isolate Floresta-FMV) was inoculated into a newborn guinea pig and a hamster (Table 2). Blood samples of this newborn guinea pig showed motile spirochetes (mean: ≤1 to 25 spirochetes/field) from the 3rd to the 5th day after inoculation (Figure 4), when it was euthanized by exsanguination and its plasma cryopreserved. Dark-field microscopy of the inoculated hamster revealed spirochetes only at the 1st, 2nd, and 9th days after inoculation (mean ≤1 or 7 spirochetes/field), despite of this experimental animal being daily examined until the 38th day (Figure 4). Finally, the blood of guinea pig 6 (isolate Serrita-FN) was inoculated into a newborn guinea pig, which showed motile spirochetes in blood at 2 to 4, 7 to 16, and 26 days after inoculation, with mean numbers of spirochetes/field varying from ≤1 to 15 (Figure 4). No experimental animal developed fever or clinical abnormalities during the present study.
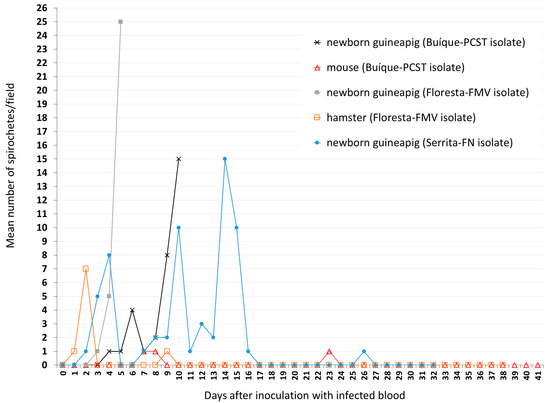
Figure 4.
Results of dark-field examination of blood samples of newborn guinea pigs, hamster, and mouse according to the number of days after inoculation with blood samples that were collected from guinea pigs showing spirochetemia due to Borrelia sp. isolate Buíque-PCST or Floresta-FMV or Serrita-FN. Values presented as the mean number of motile spirochetes per microscope field at 200x magnification in each sampled day.
3.3. Molecular Characterization of Spirochetes
PCR assays resulted in the successful amplification of fragments of four borrelial genes (rrs, flaB, glpQ, gyrB) from blood samples that were collected from spirochetemic guinea pigs 2, 4, and 6. For each borrelial gene, sequences were identical among the three guinea pigs, indicating that isolates Buíque-PCST, Floresta-FMV, and Serrita-FN represented the same Borrelia species. Results of BLAST analyses with 100% query cover showed that a 1410 bp-fragment of the rrs gene was >99.2% identical to the sequences of Borrelia hispanica (DQ057988, GU350705), Borrelia duttonii (CP000976, GU350711), and Borrelia crocidurae (CP003426, DQ057990); a 614 bp-fragment of the flaB gene was most identical (99.3%) to Borrelia sp. clone TabajaraCE from O. tabajara, Brazil (MT076263); a 450 bp-fragment of the glpQ gene was most identical (89.3%) to B. crocidurae (CP003426); and a 417 bp-fragment of the gyrB gene was most identical (89.9%) to B. crocidurae (CP004267) and B. duttonii (CP000976).
Phylogenetic analyses inferred from partial sequences of each of four genes (rrs, flaB, glpQ, gyrB) showed that in all cases, the sequences generated for Borrelia sp. (isolates Buíque-PCST, Floresta-FMV, and Serrita-FN) grouped within a clade composed by B. recurrentis and tick-borne relapsing fever borreliae of the Old World, such as B. hispanica, B. duttonii, B. crocidurae, and Borrelia persica (Figure 5 and Figure 6). For the borrelial genes rrs, flaB and glpQ, this clade also included genotypes of unnamed Borrelia spp. recently reported in Ornithodoros ticks from Brazil [15], such as Borrelia sp. Omi2MT and Borrelia sp. Omi3MT from O. mimon, Borrelia sp. TabajaraCE from O. tabajara, and Borrelia sp. OrietCE from O. rietcorreai (this latter one only for the rrs gene). Finally, in the rrs, flaB and glpQ phylogenetic trees, this large clade was sister to another large clade that contained Borrelia species associated to Ornithodoros ticks from the New World (such as the North American agents Borrelia turicatae, Borrelia parkeri and Borrelia johnsonii), and two agents from Brazil (B. venezuelensis from O. rudis, and Borrelia sp. JericoCE from O. hasei).
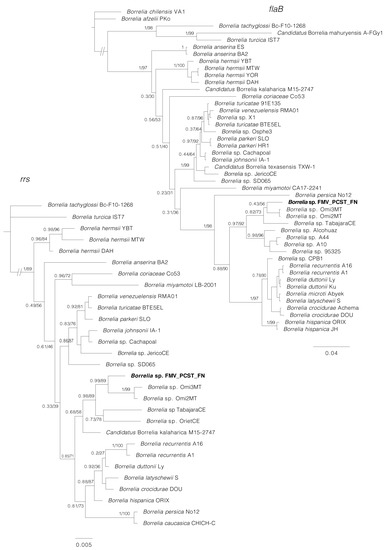
Figure 5.
Phylogenetic analyses of relapsing fever group (RFG) Borrelia spp. inferred for rrs and flaB partial sequences. The rrs tree is based on 29 sequences and an alignment of 1343 base pairs; best-fit evolutionary models calculated for ML and BI methods were TPM3u + F + G4; and M90, M15, M183, M177, M152, M85, respectively. The flaB tree is based on 46 sequences and an alignment of 638 base pairs; best-fit evolutionary models calculated for ML and BI methods were TVM + F + G4 (position-1), TPM2u + F + G4 (position-2), HKY + F + G4 (position-3); and M95, M27 (position-1); M34, M123, M129, M127 (position-2); M50, M152, M15, M90, M147, M157 (position-3), respectively. The sequence in bold (Borrelia sp. FMV_PCST_FN) represents the consensus of isolates Buíque-PCST, Floresta-FMV, and Serrita-FN from this study. Trees are drawn to scale. Numbers above or below tree branches represent Bayesian posterior probabilities/ML bootstrap values. Scale bar indicates nucleotide substitutions per site. GenBank accession numbers of the public sequences used for Borrelia phylogenies reconstruction based on rrs and flaB genes are shown in Table S2.
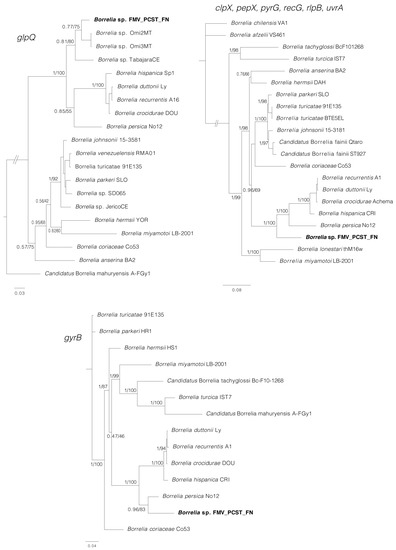
Figure 6.
Phylogenetic analyses of relapsing fever group (RFG) Borrelia spp. inferred for glpQ, gyrB, and concatenated clpX, pepX, pyrG, recG, rlpB, uvrA (MLST). The glpQ tree is based on 20 sequences and an alignment of 535 base pairs; best-fit evolutionary models calculated for ML and BI methods were TIM + F+I + G4 (position-1 and position-2), K3Pu + F+I + G4 (position-3); and M134, M200, M189, M198, M166, M203(position-1, 2, and 3) respectively. The gyrB tree is based on 14 sequences and an alignment of 1917 base pairs; best-fit evolutionary models calculated for ML and BI methods were K3Pu + F+I + G4 (position-1), TIM3 + F+I + G4 (position-2), TVM + F+I + G4 (position-3); and M125, M50, M189, M193, M157, M147 (position-1); M125, M134, M60, M189 (position-2); M147, M195, M189, M157, M203 (position-3), respectively. The MLST tree is based on 21 sequences and an alignment of 4,788 base pairs; best-fit evolutionary models calculated for ML and BI methods were GTR + F+G4 (position-1), GTR + F+G4 (position-2), GTR + F+I + G4 (position-3); and M202, M175, M193, M203 (position-1); M200, M203, M198, M134, M190, M189, M160 (position-2); M195, M157, M147 (position-3), respectively. The sequence in bold (Borrelia sp. FMV_PCST_FN) represents the consensus of isolates Buíque-PCST, Floresta-FMV, and Serrita-FN from this study. Trees are drawn to scale. Numbers above or below tree branches represent Bayesian posterior probabilities/ML bootstrap values. Scale bar indicates nucleotide substitutions per site. GenBank accession numbers of the public sequences used for Borrelia phylogenies reconstruction based on glpQ and gyrB genes are shown in Table S2. Sequence Type (ST) numbers of the sequences used for Borrelia phylogenies reconstruction based on MLST genes are shown in Table S3.
PCR amplification and DNA sequences were obtained for six of the eight MLST loci (clpX, pepX, pyrG, recG, rplB, and uvrA) from guinea pig blood samples. Pairwise comparisons proved that the Borrelia sequences from guinea pigs 2, 4, and 6 were identical with each other. The phylogenetic analysis of concatenated MLST sequences (Figure 6) corroborates the previous trees, indicating that Borrelia sp. (isolates Buíque-PCST, Floresta-FMV, and Serrita-FN) belongs to the RFG borreliae, in which it grouped within a clade that included B. recurrentis and tick-borne relapsing fever borreliae of the Old World (B. hispanica, B. duttonii, B. crocidurae and B. persica). This clade was sister to a large clade composed mostly by North American agents (B. turicatae, B. parkeri and B. johnsonii) associated with Ornithodoros spp.
4. Discussion
In this study we obtained three primary isolates (Buíque-PCST, Floresta-FMV, and Serrita-FN) of a novel RFG Borrelia species through the feeding of O. cf. tabajara ticks upon guinea pigs, which showed spirochetemia between 9 and 39 days after tick feeding. Guinea pig infection was confirmed by passage into experimental animal, based on the inoculation of guinea pig infected blood in newborn guinea pigs, mouse, and hamster. Although the three borrelial isolates were from three geographically distinct populations of O. cf. tabajara ticks, molecular analyses indicated that the three populations were infected by the same borrelial organism, as they showed identical DNA partial sequences of 10 Borrelia genes, rrs, flaB, glpQ, gyrB, clpX, pepX, pyrG, recG, rplB, and uvrA. Phylogenetic analyses based on these partial sequences indicated that isolates Buíque-PCST, Floresta-FMV, and Serrita-FN represent a distinct taxon that is more closely related to Old World Ornithodoros-associated Borrelia species than to New World borreliae (including B. venezuelensis, which was recently isolated from O. rudis from Brazil [13]).
In the phylogenetic analyses inferred from partial sequences of the borrelial genes rrs, flaB and/or glpQ, the isolates Buíque-PCST, Floresta-FMV, and Serrita-FN formed a monophyletic group with borrelial agents recently reported by molecular methods in Ornithodoros ticks from Brazil [15]. However, the phylogenetic distances between the herein characterized agent and these previous borrelial haplotypes were higher than the distances between several RFG Borrelia valid species (Figure 5 and Figure 6), reinforcing that isolates Buíque-PCST, Floresta-FMV and Serrita-FN represent a new RFG taxon. Interestingly, the insertion of this Brazilian monophyletic group within a large clade composed by Ornithodoros-associated RFG borreliae from the Old World (B. hispanica, B. duttonii, B. crocidurae, and B. persica) refute the classical paradigm that Ornithodoros-associated RFG borreliae are divided into two clades, one composed by Old World species, and another by New World species [8]. Undoubtedly, the historical support of this hypothesis was related to the little exploration of the diversity of RFG borreliae in many parts of the world, especially in South America. The improvement in phylogenetic tools adopted in recent studies have pointed out this division as rather artificial [7]. For instance, at least two novel RFG borrelial agents from Africa (an unnamed Borrelia sp. and ‘Candidatus Borrelia fainii’) were shown by phylogenetic analyses to belong to the classical New World clade of RFG Borrelia spp. [42,43].
The procedure of feeding field-collected ticks on laboratory animals (i.e., xenodiagnosis) has been used to recover spirochetes before cultivation in axenic media [44]. When this procedure is successful, it also demonstrates vector competence. Hence, herein we demonstrated that O. cf. tabajara is a competent vector of Borrelia sp. isolates Buíque-PCST, Floresta-FMV, and Serrita-FN. Previous studies demonstrated that the characterization of RFG borreliae through experimental infection of rodents resulted in variable degrees of clinical signs and borrelial pathogenicity [8]. In the present study, none of the infected rodents developed fever or any clinical sign of illness during the evaluated period, including during the highest peaks of spirochetemia. This finding is similar to studies with B. hermsii, which induced no signs of illness in chipmunks (Eutamias amoenus) and meadow voles (Microtus pennsylvanicus) during spirochetemia [9], and with Borrelia crocidurae, which also did not induce clinical signs in the multimammate rat during spirochetemia (Mastomys natalensis) [10]. In contrast, B. hermsii induced clinical illness coincidently with spirochetemia in pine squirrels (Tamiasciurus hudsonicus richardsoni) [9]. Since there is no correlation between the pathogenicity of a borrelial agent to humans and to laboratory animals, it is not known if Borrelia sp. isolates Buíque-PCST, Floresta-FMV, and Serrita-FN are capable of causing relapsing fever in humans.
Even though we used only one laboratory mouse and one hamster for experimental infections, guinea pigs were clearly more susceptible than the former two rodents to the infection by Borrelia sp. isolates Buíque-PCST, Floresta-FMV, and Serrita-FN; i.e., during spirochetemia peaks, mean number of spirochetes/microscopy field varied from 10 to 30 in all but one guinea pigs, contrasting to maximal peaks of ≤1 in a mouse and 7 spirochetes/microscopy field in a hamster (Figure 3 and Figure 4). Spirochetemia in guinea pigs were observed up to ≈30 days, similarly to previous studies using different rodent species infected with B. hermsii or B. crocidurae [9,10,45]. The spirochetemic period in guinea pigs was characterized by two or three peaks interspersed by few days with no or very low spirochetemia (≤1 spirochete/microscopy field). This pattern has been reported for RFG borreliae and is related to antigenic variation of bacterial major surface immunogenic proteins [variable major proteins (Vmps)] during the infection period [8].
Most of the RFG borreliae are maintained in enzootic cycles between Ornithodoros ticks and rodents [7,8]. The two tick species of the present study, O. rietcorreai and O. cf. tabajara, were collected from rocky formations inhabited by rodents, including the Caviidae rock cavy. In fact, this rodent species is regarded as one of main hosts for O. rietcorreai [19] and possibly also for O. tabajara [20]. This condition motivated us to test a laboratory Caviidae species (guinea pig, Cavia porcellus) for isolation of borreliae, given the phylogenetic close relatedness between rocky cavy and guinea pig [46]. Indeed, our results of spirochetemic guinea pigs claim for additional field studies to explore the occurrence of natural borrelial infection in rocky cavy and other Caviidae species living in rocky formations with the presence of O. cf. tabajara, and the likely participation of these rodents in the enzootic cycle of Borrelia sp. isolates Buíque-PCST, Floresta-FMV, and Serrita-FN.
Although O. cf. tabajara was collected in sympatry with O. rietcorreai in the prospected environments of the present study, borreliae were not observed in the guinea pigs infested with the latter tick species. This condition suggests a specificity affinity of isolates Buíque-PCST, Floresta-FMV, and Serrita-FN to O. cf. tabajara. In fact, a strict host specificity between borreliae and Ornithodoros ticks has been reported for most of the RFG agents [7,8]. On the other hand, we cannot exclude the possibility that O. rietcorreai specimens were infected by any of the current isolates, since we did not evaluate ticks by direct methods such as PCR targeting borrelial genes. In addition, it is also possible that those O. rietcorreai specimens were carrying another borrelial agent not infective for guinea pigs. For instance, Muñoz-Leal et al. [15] reported molecular detection of another RFG agent (Borrelia sp. OrietCE) in O. rietcorreai from Ceará state (Caatinga biome), suggesting that this tick species might also be associated with a specific borrelial agent.
Although the O. cf. tabajara ticks presented morphological features compatible with the original description of O. tabajara from Ceará state, their 16S rDNA sequences were 8% different, indicating that they could represent different species or two different lineages of O. tabajara. Interestingly, the original O. tabajara ticks from Ceará were found infected by another borrelial agent, Borrelia sp. TabajaraCE [15]. In our phylogenetic analyses inferred from partial sequences of the rrs, flaB and glpQ genes (Figure 5 and Figure 6), Borrelia sp. TabajaraCE was shown to be distinct but closely related to the O. cf. tabajara isolates (Buíque-PCST, Floresta-FMV, and Serrita-FN). Indeed, these findings are coherent with the paradigm of strict host specificity between borreliae and Ornithodoros ticks [7,8].
Based on the unique genetic profile of isolates Buíque-PCST, Floresta-FMV, and Serrita-FN, we propose to name them as ‘Candidatus Borrelia caatinga’, in allusion to their geographical origin, the Brazilian Caatinga biome. However, we are aware that a formal description and validation of the taxon ‘Ca. B. caatinga’ needs to be performed in a near future after its establishment in axenic media and determination of its entire genome.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms11020370/s1, Table S1: List of primers used in the multilocus sequencing typing (MLST) scheme according to Margos et al. [26] for amplification of eight genes of Borrelia spp. of the relapsing fever group; Table S2: GenBank accession numbers of the public sequences used for Borrelia phylogenies reconstruction based on rrs, flaB, glpQ and gyrB genes; Table S3: Sequence Type (ST) numbers of the sequences used for Borrelia phylogenies reconstruction based on MLST genes; Video S1: Motile spirochetes in the blood of guinea pig 6, observed by dark-field microscopy (original magnification: 200×).
Author Contributions
Conceptualization, G.M.B.d.O., S.M.-L. and M.B.L.; methodology, G.M.B.d.O., S.M.-L., A.S., B.C.W., Á.A.F.-M., M.C.H. and M.B.L.; software, S.M.-L. and A.S.; validation, G.M.B.d.O., S.M.-L. and M.B.L.; formal analysis, G.M.B.d.O., S.M.-L. Á.A.F.-M. and M.B.L.; investigation, G.M.B.d.O., S.M.-L., B.C.W., M.C.H. and M.B.L.; resources, G.M.B.d.O., S.M.-L., M.C.H. and M.B.L.; data curation, G.M.B.d.O., S.M.-L. and M.B.L.; writing—original draft preparation, G.M.B.d.O., S.M.-L. and M.B.L.; writing—review and editing, G.M.B.d.O., S.M.-L., A.S., B.C.W., Á.A.F.-M., M.C.H. and M.B.L.; supervision, M.B.L.; project administration, G.M.B.d.O. and M.B.L.; funding acquisition, G.M.B.d.O., S.M.-L. and M.B.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES Finance Code 001), Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP Grant # 2018/02521-1 to S.M.-L.), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq Grant # 409262/2018-8 to M.B.L., Grant # 314019/2021-9 to M.C.H, and scholarship to G.M.B.d.O.). S.M.-L. was also funded by Fondecyt Iniciación 11220177 (ANID, Chile).
Data Availability Statement
Partial sequences of ticks and borrelial genes generated in this study are available in GenBank under the accession numbers OP940118-OP940122 and OP937326-OP937328 for tick 16SrRNA and H3 genes, respectively; OP941213 for borrelial rrs gene, and OP952107-OP952115 for borrelial flaB, glpQ, gyrB, clpX, pepX, pyrG, recG, rplB, and uvrA protein genes. GenBank accession numbers of the public sequences used for Borrelia phylogenies reconstruction based on rrs, glpQ, gyrB and flaB genes are shown in Table S2. ST numbers of the sequences used for Borrelia phylogenies reconstruction based on MLST genes are shown in Table S3.
Acknowledgments
We are grateful to Eline A. R. Souza for valuable help during the fieldwork.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Trevisan, G.; Cinco, M.; Trevisini, S.; di Meo, N.; Chersi, K.; Ruscio, M.; Forgione, P.; Bonin, S. Borreliae part 1: Borrelia lyme group and echidna-reptile group. Biology 2021, 10, 1036. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, G.; Cinco, M.; Trevisini, S.; di Meo, N.; Ruscio, M.; Forgione, P.; Bonin, S. Borreliae part 2: Borrelia relapsing fever group and unclassified Borrelia. Biology 2021, 10, 1117. [Google Scholar] [CrossRef] [PubMed]
- Adeolu, M.; Gupta, R.S. A phylogenomic and molecular marker based proposal for the division of the genus Borrelia into two genera: The emended genus Borrelia containing only the members of the relapsing fever Borrelia, and the genus Borreliella gen. nov. containing the members of the lyme disease Borrelia (Borrelia burgdorferi sensu lato complex). Antonie Van Leeuwenhoek 2014, 105, 1049–1072. [Google Scholar] [CrossRef] [PubMed]
- Barbour, A.G.; Gupta, R.S. The family Borreliaceae (Spirochaetales), a diverse group in two genera of tick-borne spirochetes of mammals, birds, and reptiles. J. Med. Entomol. 2021, 58, 1513–1524. [Google Scholar] [CrossRef] [PubMed]
- Margos, G.; Gofton, A.; Wibberg, D.; Dangel, A.; Marosevic, D.; Loh, S.-M.; Oskam, C.; Fingerle, V. The genus Borrelia reloaded. PLoS ONE 2018, 13, e0208432. [Google Scholar] [CrossRef] [PubMed]
- Margos, G.; Wormser, G.P.; Schwartz, I.; Markowicz, M.; Henningsson, A.J.; Lienhard, R.; Stevenson, B.; Estrada-Peña, A.; Sing, A.; Fingerle, V.; et al. Evidence of taxonomic bias in public databases: The example of the genus Borrelia. Ticks Tick Borne Dis. 2022, 13, 101994. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.J. Relapsing fever borreliae: A global review. Clin. Lab. Med. 2015, 35, 847–865. [Google Scholar] [CrossRef]
- Talagrand-Reboul, E.; Boyer, P.H.; Bergström, S.; Vial, L.; Boulanger, N. Relapsing fevers: Neglected tick-borne diseases. Front. Cell. Infect. Microbiol. 2018, 8, 98. [Google Scholar] [CrossRef]
- Burgdorfer, W.; Mavros, A.J. Susceptibility of various species of rodents to the relapsing fever spirochete, Borrelia hermsii. Infect. Immun. 1970, 2, 256–259. [Google Scholar] [CrossRef]
- Boardman, K.; Rosenke, K.; Safronetz, D.; Feldmann, H.; Schwan, T.G. Host competency of the multimammate rat mastomys natalensis demonstrated by prolonged spirochetemias with the African relapsing fever spirochete borrelia crocidurae. Am. J. Trop. Med. Hyg. 2019, 101, 1272–1275. [Google Scholar] [CrossRef]
- Faccini-Martínez, Á.A.; Silva-Ramos, C.R.; Santodomingo, A.M.; Ramírez-Hernández, A.; Costa, F.B.; Labruna, M.B.; Muñoz-Leal, S. Historical overview and update on relapsing fever group Borrelia in Latin America. Parasit. Vectors 2022, 15, 196. [Google Scholar] [CrossRef] [PubMed]
- Davis, G.E. Observations on the biology of the argasid tick, Ornithodoros brasiliensis Aragão, 1923; with the recovery of a spirochete Borrelia brasiliensis, n. sp. J. Parasitol. 1952, 5, 473–476. [Google Scholar] [CrossRef]
- Muñoz-Leal, S.; Faccini-Martínez, Á.A.; Costa, F.B.; Marcili, A.; Mesquita, E.T.K.C.; Marques, E.P., Jr.; Labruna, M.B. Isolation and molecular characterization of a relapsing fever Borrelia recovered from Ornithodoros rudis in Brazil. Ticks Tick Borne Dis. 2018, 9, 864–871. [Google Scholar] [CrossRef]
- Kneubehl, A.R.; Krishnavajhala, A.; Leal, S.M.; Replogle, A.J.; Kingry, L.C.; Bermúdez, S.E.; Labruna, M.B.; Lopez, J.E. Comparative genomics of the Western Hemisphere soft tick-borne relapsing fever borreliae highlights extensive plasmid diversity. BMC Genomics. 2022, 23, 410. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Leal, S.; Faccini-Martínez, Á.A.; Teixeira, B.M.; Martins, M.M.; Serpa, M.C.A.; Oliveira, G.M.B.; Jorge, F.R.; Pacheco, R.C.; Costa, F.B.; Luz, H.R.; et al. Relapsing fever group borreliae in human-biting soft ticks, Brazil. Emerg. Infect. Dis. 2021, 27, 322–324. [Google Scholar] [CrossRef]
- Faccini-Martínez, Á.A.; Muñoz-Leal, S.; Labruna, M.B.; Angerami, R.N. Borrelioses in Brazil: Is it time to consider tick-borne relapsing fever a neglected disease in Brazil? Rev. Soc. Bras. Med. Trop. 2021, 54, e0443. [Google Scholar] [CrossRef]
- da Silva, J.M.C.; Leal, I.R.; Tabarelli, M. Caatinga, the Largest Tropical Dry Forest Region in South America; Springer: Cham, Switzerland, 2017; pp. 1–482. [Google Scholar]
- Dantas-Torres, F.; Fernandes Martins, T.; Muñoz-Leal, S.; Onofrio, V.C.; Barros-Battesti, D.M. Ticks (Ixodida: Argasidae, Ixodidae) of Brazil: Updated species checklist and taxonomic keys. Ticks Tick Borne Dis. 2019, 10, 101252. [Google Scholar] [CrossRef]
- Labruna, M.B.; Nava, S.; Marcili, A.; Barbieri, A.R.; Nunes, P.H.; Horta, M.C.; Venzal, J.M. A new argasid tick species (Acari: Argasidae) associated with the rock cavy, Kerodon rupestris Wied-Neuwied (Rodentia: Caviidae), in a semiarid region of Brazil. Parasit. Vectors 2016, 9, 1–15. [Google Scholar] [CrossRef]
- Muñoz-Leal, S.; Venzal, J.M.; Jorge, F.R.; Teixeira, B.M.; Labruna, M.B. A new species of soft tick from dry tropical forests of Brazilian Caatinga. Ticks Tick-Borne Dis. 2021, 12, 101748. [Google Scholar] [CrossRef]
- Sangioni, L.A.; Horta, M.C.; Vianna, M.C.; Gennari, S.M.; Soares, R.M.; Galvão, M.A.; Schumaker, T.T.; Ferreira, F.; Vidotto, O.; Labruna, M.B. Rickettsial infection in animals and Brazilian spotted fever endemicity. Emerg. Infect. Dis. 2005, 11, 265–270. [Google Scholar] [CrossRef]
- Mangold, A.J.; Bargues, M.D.; Mas-Coma, S. Mitochondrial 16S rDNA sequences and phylogenetic relationships of species of Rhipicephalus and other tick genera among Metastriata (Acari: Ixodidae). Parasitol. Res. 1998, 84, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Sands, A.F.; Apanaskevich, D.A.; Matthee, S.; Horak, I.G.; Matthee, C.A. The effect of host vicariance and parasite life history on the dispersal of the multi-host ectoparasite, Hyalomma truncatum. J. Biogeogr. 2017, 44, 1124–1136. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Margos, G.; Gatewood, A.G.; Aanensen, D.M.; Hanincová, K.; Terekhova, D.; Vollmer, S.A.; Cornet, M.; Piesman, J.; Donaghy, M.; Bormane, A.; et al. MLST of housekeeping genes captures geographic population structure and suggests a European origin of Borrelia burgdorferi. Proc. Natl. Acad. Sci. USA 2008, 105, 8730–8735. [Google Scholar] [CrossRef]
- Ataliba, A.C.; Resende, J.S.; Yoshinari, N.; Labruna, M.B. Isolation and molecular characterization of a Brazilian strain of Borrelia anserina, the agent of fowl spirochaetosis. Res. Vet. Sci. 2007, 83, 145–149. [Google Scholar] [CrossRef]
- Ras, N.M.; Lascola, B.; Postic, D.; Cutler, S.J.; Rodhain, F.; Baranton, G.; Raoult, D. Phylogenesis of relapsing fever Borrelia spp. Int. J. Syst. Bacteriol. 1996, 46, 859–865. [Google Scholar] [CrossRef]
- Schwan, T.G.; Raffel, S.J.; Schrumpf, M.E.; Policastro, P.F.; Rawlings, J.A.; Lane, R.S.; Breitschwerdt, E.B.; Porcella, S.F. Phylogenetic analysis of the spirochetes Borrelia parkeri and Borrelia turicatae and the potential for tick-borne relapsing fever in Florida. J. Clin. Microbiol. 2005, 43, 3851–3859. [Google Scholar] [CrossRef]
- Stromdahl, E.Y.; Williamson, P.C.; Kollars, T.M.; Evans, S.R.; Barry, R.K.; Vince, M.A.; Dobbs, N.A. Evidence of Borrelia lonestari DNA in Amblyomma americanum (Acari: Ixodidae) removed from humans. J. Clin. Microbiol. 2003, 41, 5557–5562. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. Maximum-likelihood models for combined analyses of multiple sequence data. J. Mol. Evol. 1996, 42, 587–596. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Larget, B.; Alfaro, M.E. Bayesian phylogenetic model selection using reversible jump Markov chain Monte Carlo. Mol. Biol. Evol. 2004, 21, 1123–1133. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Schwarz, G. Estimating the dimension of a model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Rannala, B. Frequentist properties of Bayesian posterior probabilities of phylogenetic trees under simple and complex substitution models. Syst. Biol. 2004, 53, 904–913. [Google Scholar] [CrossRef]
- Kisinza, W.N.; McCall, P.J.; Mitani, H.; Talbert, A.; Fukunaga, M. A new identified tick-borne Borrelia species and relapsing fever in Tanzania. Lancet 2003, 362, 1283–1284. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Nakao, R.; Hang’ombe, B.M.; Sato, K.; Kajihara, M.; Kanchela, S.; Changula, K.; Eto, Y.; Ndebe, J.; Sasaki, M.; et al. Human borreliosis caused by a new world relapsing fever borrelia-like organism in the old world. Clin. Infect. Dis. 2019, 69, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.J.; Rudenko, N.; Golovchenko, M.; Cramaro, W.J.; Kirpach, J.; Savic, S.; Christova, I.; Amaro, A. Diagnosing borreliosis. Vector-Borne Zoonotic Dis. 2017, 17, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.L.; Fischer, R.J.; Raffel, S.J.; Schwan, T.G. Host associations and genomic diversity of Borrelia hermsii in an endemic focus of tick-borne relapsing fever in western North America. Parasit. Vectors 2016, 9, 575. [Google Scholar] [CrossRef]
- Rowe, D.L.; Honeycutt, R.L. Phylogenetic relationships, ecological correlates, and molecular evolution within the Cavioidea (Mammalia, Rodentia). Mol. Biol. Evol. 2002, 19, 263–277. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).