Abstract
Globally, tomato is the second most cultivated vegetable crop next to potato, preferentially grown in temperate climates. Processing tomatoes are generally produced in field conditions, in which soilborne pathogens have serious impacts on tomato yield and quality by causing diseases of the tomato root system. Major processing tomato-producing countries have documented soilborne diseases caused by a variety of pathogens including bacteria, fungi, nematodes, and oomycetes, which are of economic importance and may threaten food security. Recent field surveys in the Australian processing tomato industry showed that plant growth and yield were significantly affected by soilborne pathogens, especially Fusarium oxysporum and Pythium species. Globally, different management methods have been used to control diseases such as the use of resistant tomato cultivars, the application of fungicides, and biological control. Among these methods, biocontrol has received increasing attention due to its high efficiency, target-specificity, sustainability and public acceptance. The application of biocontrol is a mix of different strategies, such as applying antagonistic microorganisms to the field, and using the beneficial metabolites synthesized by these microorganisms. This review provides a broad review of the major soilborne fungal/oomycete pathogens of the field processing tomato industry affecting major global producers, the traditional and biological management practices for the control of the pathogens, and the various strategies of the biological control for tomato soilborne diseases. The advantages and disadvantages of the management strategies are discussed, and highlighted is the importance of biological control in managing the diseases in field processing tomatoes under the pressure of global climate change.
1. Introduction
Tomato (Solanum lycopersicum) has been under global cultivation for several centuries having emerged from western South America [1]. Due to its wide cultivation and unique nutritive value, tomato has become the world’s second most cultivated vegetable after potato and the most popular canned vegetable [2,3].
Tomatoes can either be marketed for fresh consumption or as processed products. Around 40 Mt tomatoes are processed annually, making tomatoes the most processed vegetable by weight [4]. The major product of the processing tomato industry is aseptic paste [5], and there are other products like diced and whole peeled tomatoes made into canned food, as well as tomato juice [6]. Processing tomatoes are generally grown in field conditions [7].
Tomatoes are susceptible to over 200 diseases [8] with soilborne diseases being a major component. Fusarium oxysporum has been regarded as one of the most important threats to both field and greenhouse-grown tomatoes worldwide with 10–80% of yield loss [8,9]. Another fungal pathogen, Pyrenochaeta lycopersici, recently renamed Pseudopyrenochaeta lycopersici [10,11], that causes corky root rot, in cooler climates such as northern Europe, can cause up to 75% yield loss of affected tomatoes [12]. Soilborne pathogens are believed to be hard to control for their persistence in soil and wide host range [13].
Traditionally, management methods for tomato diseases use resistant cultivars, chemicals like fungicides and pesticides, physical methods such as soil solarization and soil heating, and cultural methods such as crop rotation and hygiene [9,14,15]. Chemical treatments sometimes have adverse effects on the environment [16,17] and human health and physical control are often laborious and costly.
Breeding for disease resistance is a major objective for most public and private tomato breeders [18]. Breeders usually rely on the genetic diversity within wild tomato species to incorporate desirable traits, especially the resistance to different diseases, by crossing wild tomatoes with cultivated tomatoes [19]. Though genetic resistances usually have high efficiency, the screening for the traits and breeding process can be laborious and time-consuming [20,21], which may hamper the improvement progress of commercial tomato production.
Biological control, also known as biocontrol, is a method of controlling pests and diseases using other organisms (biocontrol agents) through the importation, augmentation, or conservation of agents in the environment [22,23], and is gaining increasing acceptance. Compared with conventional disease/pest control methods, its major advantages are the minimization of the public concerns of impacts on health and environment [24,25], self-sustainability, host-specificity, and cost-efficiency [26,27]. The commercialization of biocontrol for plant disease is relatively a new concept, with the majority of biocontrol agents registered in the United States Environmental Protection Agency after 2000 [28].
This review provides a comprehensive description of the tomato production systems and the major soilborne fungal/oomycete diseases of the global leading processing tomato-producing countries, analysis of the different control strategies for the management of the respective diseases with an emphasis on biocontrol, and proposes a future perspective for disease management within processing tomato industries which are under the threat of emerging pathogens due to climate change.
2. The Major Countries Producing Processing Tomatoes and Their Major Soilborne Fungal/Oomycete Diseases
2.1. Northern Hemisphere
World tomato production is more concentrated in the northern hemisphere, with an estimated amount of over 34 million metric tonnes (mT) for processing in 2022, with the southern hemisphere producing less than 2.7 million metric tonnes [4]. This is likely to be the result of the combined effects of climate, human and economic factors. The majority of the total processing tomato production (over 65%) is shared by the United States (California), China, and Italy [29].
2.1.1. The United States Tomato Industry and Major Soilborne/Fungal Diseases
Though only third in total tomato production size, the United States is the largest producer of processed tomatoes globally. California is the dominant state of US processed tomato production, which accounts for 95% of the nation’s tomatoes [30], with the production of 9.8 million mT in 2021 [29]. In California, the production of processing tomatoes is carried out on large farms, with 225 growers owning about 277,000 acres of land [31]. The production starts with the wet soil condition in spring and the harvest and processing in late July until October [32]. Processing tomatoes are mostly irrigated with sprinkler systems, with the increasing application of drip irrigation in the areas threatened by rising saline underground water [33]. All the harvests are performed mechanically, with the fruits graded by inspection before processing [33]. The other USA States with large processing tomato production are Florida and Indiana, but their production scale is very small compared with that of California [34].
Fresh tomato accounts for 8% of total U.S. tomato production [35], which is cultivated nationwide in the USA, with California and Florida being the major production sites, contributing to around 80% of the national production [36]. In California, tomatoes are produced year-round except in winter with the wide application of greenhouse production [36], while in Florida, the growing season is from October to June [35]. In other states, tomato production runs through the summer months when Florida tomatoes are off the market [35].
One important soilborne disease, buckeye rot, is reported to be caused by several oomycete pathogens, including Phytophthora nicotianae var. parasitica, P. capsici and P. cryptogea [37,38,39]. Buckeye rot was first reported in Florida in 1915, causing about 15% yield reduction [40], and was reported in California about two decades later [41]. Buckeye rot was found to cause significant tomato yield reduction in the furrow-irrigated zones of California in 1984, while the major pathogen was P. parasitica [39]. However, P. capsici was later found to be more virulent to tomatoes, which can also infect P. parasitica-resistant tomato varieties [42,43]. In recent years, buckeye rot is most common in the southeast and central states of the U.S. [44].
P. parasitica and P. capsici can also induce tomato Phytophthora root rot by invading plant roots. Reported in California in 1955, the disease had repeated outbreaks in the major processing tomato production zones of the state, leading to the almost complete destruction of the crop [45]. It was also reported that P. parasitica was more frequently isolated from Californian processing tomato production sites while P. capsici was more abundant in regions growing fresh tomatoes [43,45].
Corky root rot caused by the soilborne fungus P. lycopersici is considered to be another major disease limiting Californian tomato production [46]. Campbell [47] tested the pathogen on fresh market tomatoes and harvested 73% fewer large fruits in infected plots without any treatments, which was coherent with the estimation of processing tomato growers consulted in the study. Also, in another study covering nine organic farms and 18 conventional farms in the Central Valley of California, corky root rot was the disease found on most plants in most sampling locations [48]. Recently, P. lycopersici was also found to form a disease complex with other pathogens such as Colletotrichum coccodes in Ohio, causing severe wilting of tomatoes both in fields and glasshouses [49].
Fusarium wilt caused by F. oxysporum f. sp. lycopersici (Fol) also affects tomatoes throughout the United States. All three known physiological races of Fol have been reported in the United States. Race 1 was initially reported in 1886, which severely threatened commercial tomato production in Arkansas [50]. Later, Fol race 2 which overcame the tomato variety resistant to race 1 was reported in Ohio in 1945 [51]. Fol race 3 was reported across the United States [52], causing up to 80% disease incidence in tomatoes resistant to Fol race 1 and 2 when first reported in Florida in 1982 [53], while also significantly affecting the processing tomatoes in California [54]. Since the 2000s, Fusarium wilt has become a destructive disease in the south-eastern parts of the United States [44].
Fusarium crown and root rot (FCRR) is also a major soilborne disease of both U.S. field and greenhouse tomato production. The pathogen, Fusarium oxysporum f. sp. radicis–lycopersici (Forl), was first reported in Japan in 1974 and was later reported across the United States from California to Florida [55,56]. In Florida, the disease was reported to cause a tomato yield reduction of 15–65% [56].
The disease Verticillium wilt caused by Verticillium dahliae was first reported in California in 1926 [57]. With the resistance Ve gene incorporated into commercial tomato varieties, race 2 of V. dahliae pathogenic overcoming resistant tomatoes in Ohio and then California was reported [58,59]. Due to the fact that no resistant gene is available in tomato germplasm against V. dahliae race 2, this pathogen is now considered one of the diseases limiting the productivity of the Californian tomato industry [46]. In California, the yield loss caused by V. dahliae was up to 67% in a susceptible cultivar, with race 2 reducing V. dahliae race 1-resistant tomato yield by up to 25% at 100% disease incidence [59,60]. Later, Verticillium wilt also became an increasing problem for the Pacific Northwest (Washington, Oregon, and Idaho) states of the U.S., influencing not only tomatoes but also watermelons [61].
2.1.2. The Chinese Tomato Industry and Major Soilborne Fungal/Oomycete Diseases
Globally, China ranks first in total tomato production [32] and comes second in terms of estimated 2022 processing tomato production [4]. The major production site of the Chinese tomato industry is Xinjiang province, which accounts for over 80% of the total production size [4]. The major production site of processed tomatoes is the north-western provinces, including Xinjiang, Inner Mongolia and Gansu [32]. The cultivation in Xinjiang is from spring to summer (April to July), while the production is processed between August and October [62].
Tomato for fresh consumption is produced across China, with four provinces, Hebei, Henan, Shandong, and Xinjiang, producing the vast majority [63]. Due to the continuous huge market demand, the fresh tomato industry relies on the utilization of solar-type greenhouses with a year-round production cycle, with Shandong producing over 60% of fresh tomatoes in winter [32].
At least 10 diseases are reported to cause significant tomato yield loss in China [64], with one of them being Fusarium wilt. Among the three major physiological races of Fol, race 1 is the main pathogen of tomato Fusarium wilt in China according to field surveys conducted in several northern provinces including Heilongjiang, Shaanxi and Shanxi [65], while race 2 was only reported in Zhejiang, causing 30–60% tomato yield loss [66].
Recently, symptoms of putative wilt and root rot diseases were reported in several regions growing tomato varieties resistant to Fol race 1 and 2 from 2016 to 2018 [67]. Ye et al. [67] conducted field sampling in high-incidence provinces in Zhejiang, Hainan, Shanxi, and Shandong, and found that among the 64 collected F. oxysporum isolates, 35 were Fol race 3 isolates and 13 were Forl isolates. Therefore, Fol race 3 and Forl seem to have emerged as the major Fusaria pathogens of the Chinese tomato industry, thus more effective management strategies may be required.
2.1.3. The Italian Tomato Industry and Major Soilborne Fungal/Oomycete Diseases
In the 2022 industry estimation [4], Italy had the third largest processing tomato industry globally. Over 95% of Italian tomato production is processed [32]. Tomato production is throughout the country, with differences in production methods in different geographical regions. In the northern part of the country, the production is completely mechanized in large farms, with manual harvesting abolished by the early 1990s [68]. This area is the major production site for processing tomatoes with 2.885 million tonnes of estimated yield in 2022 [4]. In southern Italy, farms are usually owned by families, with smaller sizes of 4–10 ha and around 50% of them still practice manual harvesting [32,68]. The center-south region is Italy’s second-largest processing tomato production zone, and its estimated yield in 2022 was 2.59 million tonnes [4]. Tomatoes are planted in early May, and harvested in mid-July and the season ends in mid-September.
Tomato production in southern Italy is significantly affected by corky root rot. This disease has been reported since the 1960s in European glasshouse production, which became more severe in later years. In glasshouses in a Mediterranean environment, this disease can cause a 30–40% yield reduction [69].
In northern Italy, P. capsici was found to be the causal agent of Phytophthora root rot repeatedly observed on grafted tomatoes [70]. This disease was found to cause the sudden collapse of around 25% of the plants within 60 days of transplant [70].
Fusarium wilt is also an important tomato disease in Italy. Fol race 1 has been long reported in Italy, with race 2 first reported in 1999 [71]. Nowadays, Fusarium wilt caused by this pathogen is considered a major cause of economic loss in tomato production in Italy [72,73,74].
FCRR caused by Forl is another important soilborne disease for Italian tomato production. FCRR was reported in northern Italy as early as 1984 and then occurred as an outbreak in Sicily in 1986 [75]. This disease was found to cause substantial yield loss in both greenhouse and soilless fresh market production in several Italian tomato-producing regions such as Calabria, Emilia Romagna, Liguria, Sardinia, and Sicily [76]. FCRR was present in 24% of the greenhouses of Sicily and 66% of greenhouses in Sardinia in surveys conducted during the 1990s [77,78], with a up to 60% mortality rate during severe outbreaks.
2.2. Southern Hemisphere
Though tomatoes originated from the Latin American region of the southern hemisphere, the tomato production size of southern hemisphere countries has a comparatively small proportion in the global production. However, when compared with the major producers located in the northern hemisphere, the tomato industry in the southern hemisphere, especially field tomatoes are unique due to several factors. One major factor is the timing of summer in the southern hemisphere, which coincides with winter in the northern hemisphere, thus the most favorable season for tomato production between the two hemispheres is chronologically different. Also, the southern hemisphere generally has a milder climate compared with that of the northern hemisphere due to the higher ocean coverage [79], which may also affect the growth and development of cultivated tomatoes. Therefore, studies on soilborne pathogens threatening tomatoes under the unique climate and production process of the southern hemisphere may also be important for the understanding of the impact of a changing global climate on the disease cycle of these pathogens.
2.2.1. The Brazilian Tomato Industry and Major Soilborne Fungal/Oomycete Diseases
In the Southern hemisphere, Brazil is the largest processing tomato producer, with around 1.5 million tonnes in 2021 [29]. Most of Brazil’s tomato production is located in the southern provinces close to the coast, such as Goiás, São Paulo, Minas Gerais, Paraná, and Bahia [80]. The processing tomato production is generally based in the central provinces, with Goiás and the Cerrado region contributing to 99% of the production size due to the favorable soil and climatic conditions, and mostly irrigated with conventional methods [81]. To manage the problem caused by whitefly, the Ministry of Agriculture in Brazil set up a rule of a two-month tomato-free period for the processing industry, so the transplanting can only be carried out between 1 February and 30 June, with harvest completed by November [82].
Fusarium wilt caused by Fol is widely present in Brazil. Race 1 of Fol was first reported in São Paulo State in 1941, with only races 1 and 2 detected in all surveys in the 20th century [83]. In 2005, race 3 of Fol was also isolated from field samples from wilted hybrid tomato plants with resistance to Fol race 1 and 2 from Espirito Santo province [83], which then shortly after spread into other provinces [84]. The severe outbreaks of Fol race 3 have led to the widespread replacement of susceptible tomato hybrids with new cultivars carrying the resistant gene i-3 in Brazil [85].
Fol was believed to be the only pathogenic Fusarium related to tomato in Brazil until the 2010s. More recently, there were frequent and simultaneous outbreaks of Fusarium oxysporum disease capable of infecting Fol-resistant tomato cultivars [85]. Subsequently, Forl was reported in surveys on field tomato plants showing crown-rot and vascular discoloration symptoms with disease indices ranging from 10 to 50% from three Southeast Brazilian states and two northern states [86].
Verticillium wilt is another important soilborne fungal disease for the Brazilian tomato industry. Despite the fact that V. dahliae and V. albo-atrum were found to cause substantial economic losses in Brazilian vegetable production, only V. dahliae was reported on tomatoes [87]. Both races 1 and 2 of V. dahliae were reported in the 1980s and late 1990s [88].
2.2.2. The Chilean Tomato Industry and Major Soilborne Fungal/Oomycete Diseases
Chile has the second largest tomato production among the southern hemisphere countries, with around 1 million tonnes for processing annually [4]. The majority of Chilean tomato production is for processing, with around 5000 ha for the fresh market and 8000 ha for the processing industry [89,90]. The production area is concentrated in the middle part of the country between 30° and 34° latitude south, benefiting from the local mid-terranean climate optimizing tomato growth [89]. Chilean processing tomato industry is fully contracted, which primarily uses transplanted seedlings sown in July, then transplanted from mid-September to mid-November [91]. Though production was heavily reliant on human labor in the last century, the growing adaptation of mechanized production has led to increased yield and size of individual contracts [91].
Similar to Brazil, Fusarium oxysporum is an emerging pathogen for the Chilean tomato industry. Though cultivars resistant to Fol race 1 and 2 are commonly used, Fusarium wilt still occurred in important production zones such as Azapa valley in northern Chile, and the pathogens were later identified as Fol race 3 and Forl [92].
Forl is also becoming an increasing threat to the Chilean tomato industry, with fresh tomato production affected most severely. FCRR was frequently present in northern Chile and central Chile, with the former practicing both net-house and open-field crops, and the latter growing monocultural tomato crops in polyhouses [93].
2.2.3. The Australian Tomato Industry and Major Soilborne Fungal/Oomycete Diseases
Though Australia has a relatively small total tomato production size, it still has the largest processing tomato production outside the Latin America region in the southern hemisphere [4]. Tomatoes in Australia are almost always grown from imported seeds [94]. The fresh and processed tomatoes are produced by two distinctive industries in Australia. The fresh tomatoes are grown in either field or hydroponic environments and are harvested by hand all year round, with Queensland and Victoria as the major production sites [95], while 97% of processing tomatoes are produced in Victoria [96].
The Australian processing tomato industry is sited primarily in the Goulburn/Murray River areas of northern Victoria, with a minority in the Riverina region of New South Wales, which is a seasonal industry (February to April) and harvested by machinery [95]. According to the Australian Processing Tomato Research Council [97], 46% of the total tomato production was sent to the processing industry in 2020.
Fusarium wilt caused by Fol is considered an economically important disease for the Australian fresh tomato industry, causing huge losses [98]. Also, among the three major races of Fol species threatening the global tomato industry, the third race was first reported in Australia in 1979 [99] in Queensland.
Recent disease surveys carried out in the Australian processing tomato industry in Victoria found that processing tomatoes were experiencing an estimated 10% yield loss from soil-borne pathogens, with the two most abundant pathogens being F. oxysporum and Pythium spp. [100,101]. The F. oxysporum strains collected in the survey by Callaghan [100] produced symptoms of rot on tomato roots and crowns such as those caused by Forl, a species not reported in Australia, with slight differences in growth temperature, a wider host range and variable pathogenicity [100]. Thus, the disease caused by this isolate was named chocolate streak disease (CSD) to differentiate it from FCRR caused by Forl.
Among the eight pathogenic Pythium species causing root rot and seedling damping-off, Callaghan et al. [101] found that P. irrgulare was one of the most aggressive pathogens, as confirmed in the pre-emergence and early seedling phases of tomato plant growth.
3. Control Strategies of the Major Soilborne Fungal/Oomycete Diseases
As discussed above, the major soilborne fungal/oomycete pathogens of the major processing tomato producers are generally similar, with Fusarium wilt being the most common disease, followed by FCRR, with Verticillium wilt, corky root, and Phytophthora root rot also common in the northern hemisphere (Figure 1). In this section, a description of the different diseases and their different management methods is provided.
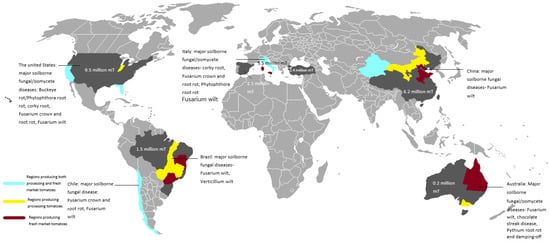
Figure 1.
Global production of processing tomatoes, major soilborne fungal/oomycete diseases, and the estimated processing tomato production size in 2022.
3.1. Tomato Corky Root Rot
Tomato corky root rot is generally caused by the fungus P. lycopersici. The roots of infected plants show dark brown, banded lesions [49]. With the development of the symptoms, even larger roots become infected, with extensive swollen and cracked brown lesions, giving them the distinctive corky look appearance [102]. The growth of the plant may be stunted and slow, but the disease usually does not kill plants, resulting in reduced yield [103].
P. lycopersici is an ascomycete, which overwinters as microconidia and hyphae, or produces microsclerotia which can withstand harsh conditions, survive in host root and soil, and maintain pathogenicity for up to 15 years [103]. When the environment becomes favorable, the microsclerotia will germinate and produce hyphae [104]. After reaching the host roots, the hyphae penetrate the host epidermis and gradually colonize the whole root [104]. This pathogen is soil-transmitted, which favors monoculture soil without proper disease management [104,105].
3.1.1. Conventional Control Methods
Cultural Control
The disease development of corky rot is at optimum at 15.5–20 °C [106]. Thus, it is better to plant tomatoes in spring when the soils start to become warm.
Though effective against many other pathogens, crop rotation alone may not be effective in controlling corky root rot, for P. lycopersici has a wide host range including cucumber, eggplant, lettuce, melons, and pepper [103].
Physical Control
Soil solarization by covering the field with plastic film for a long period is a practical method for the control of corky root rot. In Italy, Vitale et al. [69] found that solarization performed with ethylene-vinyl-acetate film has an identical level of control effect on corky rot symptoms as compared with fumigation with methyl bromide, which was better than that of metham sodium and metham potassium fumigation. However, the level of success of solarization depends on the combination of high ambient temperatures, maximum solar radiation, and optimum soil moisture as well as the existing inoculum and disease levels [107,108]. Therefore, solarization usually has varying effectiveness, and is generally less effective in climates where high summer temperatures coincide with the rainy season due to the cooling effect of rainfall and extensive clouds blocking the solar radiation [107].
Chemical Control
In fields previously reported to have corky root rot, a preplant treatment with soil fumigation was shown to reduce disease in the subsequent tomato crop [69]. Methyl bromide (MBr) used to be a preferred chemical, but it was proved to be an ozone-depletion agent which is more destructive to stratospheric ozone than chlorine [109], thus its use has been phased out in developed countries by 2005 and in 2015 by the less developed countries as required by Montreal Protocol [16]. Potential alternative chemicals such as chloropicrin, metam sodium, metam potassium, and dazomet [69,110,111] can only provide a lower control level of corky root rot compared with MBr treatment. For example, Vitale et al. [69] found that metham sodium fumigation (MS, 353 litres a.i. ha−1) and metham potassium fumigation (MK, 350 litres a.i. ha−1) did not reduce the disease incidence of corky root rot in their trial. Therefore, with reduced efficiency of chemical controls, the management of corky root rot may require the addition of more effective methods such as the use of resistant cultivars and biocontrol.
Resistance Breeding
Though breeding for resistant cultivars is a common strategy for the control of crop disease, commercial variants of both processing and fresh consumption tomatoes are susceptible to corky root disease [112]. So far, only one single recessive gene (pyl) was shown to confer resistance to corky root rot and was introgressed into Lycopersicon esculentum from L. peruvianum [113]. The pyl gene is later found to possibly be a recessive allele of a susceptibility gene [114] and it has not been cloned yet.
3.1.2. Biological Control
Some fungivorous nematodes have been recorded as potential biocontrol agents for corky root rot. Hasna et al. [115] tested two fungivorous nematodes, Aphelenchus avenae and Aphelenchoides spp. against P. lycopersici, and concluded only A. avenae was able to significantly reduce the severity of tomato root rot in greenhouse trials with a population of 3 or 23 nematodes mL−1 soil. However, in a later on-farm trial covering two tomato seasons in Sweden, Hasna et al. [105] found even at a higher inoculation rate of 50 nematodes mL−1 soil, the application of A. avanae into infested soil did not reduce corky root disease severity. Thus, the potential of nematodes to control corky root rot may not be dismissed, but the application method may still need improvements.
In greenhouse trials, bacterial antagonists such as Streptomyces spp. have been found to effectively suppress corky root disease of tomatoes and enhance plant growth, resulting in higher yields. Bubici et al. [116] evaluated the antagonism of twenty-six Streptomyces spp. against corky root rot on tomatoes in both glasshouse and field conditions and found the most effective strain can reduce disease severity up to 64% in the glasshouse and 48% in the field.
Antagonistic fungi may also be used in the biocontrol against corky root rot. Fiume and Fiume [112] conducted glasshouse trials against corky root rot using Trichoderma viride, Bacillus subtilis, and Streptomyces spp., and concluded that the application of all three microorganisms significantly reduced the corky root symptoms in terms of disease index, with T. viride having the best results, followed by Streptomyces spp. Besoain et al. [117] performed UV on native T. harzianum to obtain mutants and found the mutants ThF1-2 and ThF4-4 inhibited the growth of P. lycopersici in vitro by 1.3 and 5 fold, respectively. Sánchez-Téllez et al. [118] further tested the mutant ThF1-2 in greenhouse tomato trials and found applying solid formulation ThF1-2 resulted in a significantly lower root damage caused by P. lycopersici compared with a previous trial using MBr. The control of T. harzianum against P. lycopersici seems to be correlated to the differential expression of extracellular fungal cell wall hydrolytic enzymes between isolates [119].
Organic amendments may also help in the control of corky root rot. Workneh et al. [48] found that the application of green manure and compost reduced the corky root rot severity in organic farm tomatoes by stimulating microbial activities in a field survey. However, P. lycopersici responds differently to different amendments. Hasna et al. [120] tested composts consisting of green manure, garden waste, and horse manure against corky root rot in greenhouse tomatoes and found that garden waste compost significantly reduced the disease, whereas horse manure compost significantly stimulated disease, while the green manure compost had no effect on the disease despite the increased microbial activity. It was concluded that the disease severity of corky root rot can be suppressed by composts with a low concentration of ammonium nitrogen and a high concentration of calcium, but further studies may be necessary to further prove this perspective.
3.2. Fusarium Crown and Root Rot (FCRR) of Tomato
FCRR is caused by the pathogenic fungus Fusarium oxysporum f. sp. radicis-lycopersici (Forl). Though both being classified as Fusarium oxysporum, Forl and Fol, the agent of tomato Fusarium wilt, are two different formae speciales, which are informal taxonomic groupings based on the differences in the host range [121]. The pathogen invades the host via wounds and natural openings created by newly emerging roots [122]. Infected seedlings usually show symptoms of stunting and yellowing with premature abscission of lower leaves. With the development of the disease further via the xylem, necrotic lesions may gridle the crown, with the roots becoming rotted and discolored, and the seedlings then develop wilting symptoms [123]. On older plants, the first symptom may be the yellowing and collapse of the oldest leaves. The symptoms then develop upwards and infect young leaves. Older plants may be stunted and wilted, but still alive at harvest [56].
F. oxysporum is an ascomycete without an observable sexual stage [124]. This fungus can aggressively colonize both host roots and organic matter [125]. Though being able to persist in the soil as mycelial fragments, the highly durable chlamydospore is the major form of survival of F. oxysporum in the absence of a host [124,126]. The germination of the spore is usually triggered by the exudates from growing plant roots [124,126].
3.2.1. Conventional Control Methods
Cultural Control
Hygiene and sanitation of the seeds and transplant seedlings are important for Forl management. For example, Muslim et al. [127] found that plants not challenged with the pathogen still become infected by FCRR, which is probably due to incomplete soil sterilization. It is also strongly recommended that all equipment coming in direct contact with soil is cleaned and disinfected [56]. The pathogen may also use colonized and infected plants as carrying vectors, thus the infected plants and their roots should be removed immediately.
Crop rotation with a non-host crop may also prevent FCRR. Crops susceptible to Forl such as eggplants and peppers should be avoided in the rotation [128], while non-hosts such as lettuce may be useful to reduce inoculum levels in the soil [129]. However, the efficiency of crop rotation may be limited for FCRR control, because the pathogen can survive as chlamydospores in the soil for a long time [130].
Physical Control
FCRR is favored by cooler temperatures, thus planting in warm periods and using warm water in irrigation is recommended to restrict the development of disease [131]. Soil solarization has also been demonstrated to control FCRR. In studies testing several solarization methods, soil solarization generally reduced populations of Forl down to a depth of 5 cm [56].
Chemical Control
Before the 2010s, the most effective method for FCRR control was soil disinfection using methyl bromide (MBr) [132,133]. However, MBr has been phased out globally since 2015. The ban on MBr prompted the study of alternative chemicals for the control of soilborne pests including Forl. So far, the tested alternatives include 1,3-dichloropropene, chloropicrin, dozamet, fosthiazate, and metam sodium, with similar effects on Forl compared with MBr [56,134,135]. For example, McGovern et al. [134] tested the application of metam sodium in field tomatoes and found that rotovation of metam sodium at 935 L/ha into preformed beds consistently reduced FCRR incidence equal to those achieved by methyl bromide-chloropicrin. Also, 1,3-dichloropropene+chloropicrin (60.5% and 33.3%, w/w) was tested on Italian field tomatoes [135] and was able to achieve a good tomato yield using drip application in sandy loam soils with slight Forl infections and severe infections of Fol and galling nematodes, which was similar to those of the plots treated with MBr.
However, there are still several factors that may reduce the efficiency of Forl chemical control. For example, Forl chlamydospores were found to survive in the soil at a depth beyond 50 cm, which is unreachable by soil fumigation [131]. Also, Forl can efficiently colonize sterilized soil [55]. Therefore, soil fumigation may instead create favorable soil conditions for Forl colonization by reducing microbial competition.
Resistance Breeding
Resistant tomato varieties can also be used to control FCRR. The resistance of tomatoes to FCRR is found to be controlled by a single dominant locus (Frl) on chromosome 9 [122,136]. This gene has been successfully crossed into commercial tomato lines, with many Forl-resistant cultivars currently available. However, no additional resistant genes have been identified.
3.2.2. Biocontrol
Forl is believed to have poor competitive fitness against other microorganisms [131], thus biocontrol via organic amendments or biocontrol agents may be effective for the management of Forl.
Several antagonistic microorganisms have been tested for their properties to control FCRR. Sivan et al. [137] applied Trichoderma harzianum as seed coating or wheat-bran/peat in tomatoes grown in FCRR-infested field and recorded a 26.2% increase in yield of treated plots compared with the control, with the control of Forl at the highest effect on root tips. Datnoff et al. [138] also applied T. harzianum and Glomus intraradices into tomato fields with FCRR history and recorded a significant reduction in disease severity and disease incidence of FCRR by applying the fungi both combined and separately. Several hypervirulent binucleate Rhizoctonia strains were also found to reduce the vascular discoloration caused by FCRR on tomatoes up to 100% in greenhouse conditions and up to 70% in the field [127]. Moreover, a non-pathogenic endophytic F. solani strain was reported to reduce disease incidence of Forl when applied alone in glasshouse tomato by 47%, the effects of which improved when combined with certain fungicides [139]. Pythium oligandurm was also found to trigger the host defence of greenhouse tomatoes when challenged by Forl in the form of deposition of newly formed barriers beyond the infection sites [140].
Several bacteria species may also control FCRR. Pseudomonas fluorescens was found to synthesize the antibiotic 2,4-diacetylphloroglucinol, which suppressed the growth of Forl in vitro [141]. A further study found that P. fluorescens WCS365 used chemotaxis towards Forl hyphae, enabling it to efficiently colonize Forl and achieve control effects [142]. In a later screening by Kamilova et al. [143], strong competitive biocontrol strains P. fluorescens PCL1751 and P. putida PCL1760 were found to successfully suppress FCRR under the soil and hydroponic conditions. In addition, Baysal et al. [123] assessed in a greenhouse trial the effect of two Bacillus subtilis bacteria strains QST713 and EU07, and concluded that EU07 had a better disease inhibitory effect (disease incidence reduced by 75%) compared with QST713 (disease incidence reduced by 52%), and the inhibition may be achieved by YrvN protein coded in the genome of EU07 as a subunit of protease enzyme. Lytic enzymes, cellulases, proteases, 1,4-b-glucanase, and hydrolases from the secreted proteins from B. subtilis EU07 and FZB24 and concluded these essential proteins of Bacillus bacteria play an important role in the control of Forl [144].
Organic amendments promoting microbial activity may also be used in FCRR management, but they do not have consistent effects in field conditions. Straw was incorporated into the soil to manage FCRR by Jarvis [131], but the Forl soil population increased around and inside the straw, which only started to fall when the straw decomposed. However, Kavroulakis et al. [145] concluded that a compost mix made from grape marc wastes and extracted olive press cake can enhance tomato defensive capacity under Forl stress by making the pathogen unable to penetrate and colonize the host root, resulting in a 40% reduction in the disease incidence compared to the control. However, the plants in this trial were grown completely in the compost, making large-size commercial applications likely unrealistic.
3.3. Fusarium wilt Disease of Tomato
Fol, the causal agent of tomato Fusarium wilt, is able to penetrate plant cell epidermis, thus infecting tomato plants through the roots and colonizing the xylem for further colonization of the root system [9]. The symptoms are initially characterized as yellowing of the older leaves [50], followed by browning and wilting. Browning will also be visible in the vascular tissue, and this discoloration will extend to the apex of the plant [50]. The infected plant will experience stunted growth and drastically reduced yield, which will often die before maturity [9].
3.3.1. Conventional Control Methods
Cultural Control
Crop rotation can be used to manage Fusarium wilt, and it is recommended not to plant the same or related type of crop for at least four years if one crop is severely infected by Fusarium wilt [146]. The recommended crops for rotation are grasses and cereals [147].
Hygiene should also be practiced for Fol control. Disease-affected plants should be removed immediately. Used farming tools should be disinfected and cleaned before reuse. The use of sanitized footwear and clothes on the farm may help prevent the transportation of infected soils between paddocks [146]. Fallowing is another strategy for Fol control. Briefly, the land is left uncultivated for a period, and for Fol, it is recommended to practice fallowing during the summer months to let the high temperature and excessive drying reduce soil levels of Fol [9].
Physical Control
Soil solarization can also be used to control Fol residing in soil, preferably performed in the summertimes. However, since the development of Fusarium wilt favors warm temperatures (27–28 °C) [148], this strategy may not work in zones with cool climates.
Chemical Control
Soil fumigation with MBr was an effective method for Fol management however, with the phase-out of MBr the value of chemical control on Fol has drastically reduced. Though alternative chemicals such as chloropicrin, dimethyl disulfide, metam sodium, and 1,3-dichloropropene are available, they all lack the broad-spectrum volatile characteristics of MBr, which made it highly effective [149]. Systemic fungicides such as benomyl, thiabendazole, and thiophanate have also been used to control tomato Fusarium wilt [9], but it was believed that there are no fungicides especially effective for the control of this disease [146].
Resistance Breeding
The application of tomato cultivars resistant to Fusarium wilt is currently the most feasible management method.
The resistance to Fol was first identified by Bohn and Tucker in 1939 [150], who identified one single, dominant resistance locus later named I gene from one wild tomato accession of S. pimpinellifolium, Missouri accession 160 [151]. This gene was crossed into the first commercial Fol-resistant tomato cultivar and was located at tomato chromosome 11 [152].
Later, the second race of Fol, named Fol2 was reported to spread widely in Florida in the 1960s [153], which led to another screening for the corresponding resistant gene. The resistant gene was again found in wild tomato relatives- a natural hybrid PI126915, which was name I-2 and mapped to chromosome 11 [154].
In 1979, the third race of Fol–Fol3 was reported in Australia in fresh tomato production [99]. McGrath et al. [155] were the first to identify resistance to Fol3 in the S. pennellii accession PI414773 in 1987, and Scott and Jones [156] later identified a dominant Fol3 resistance locus in the S. pennellii accession LA716. This newly discovered gene was later named I-3 and used as the primary source of Fol3 resistance in commercial varieties. Gene I-3 was mapped to chromosome 7 [157], and McGrath et al. located another gene I-7 gene in chromosome 8 [158].
Three additional genes with partial resistance to Fol2 were also found by Sela-Buurlage et al. [152]. These researchers studied 53 introgression lines with chromosomes from LA716 and identified alleles I-4 and locus I-5 on chromosome 2, with locus I-6 on chromosome 10 of S. pennellii. However, none of these genes have their effects validated nor used for commercial purposes so far.
3.3.2. Biological Control
Potential biocontrol agents against Fol on tomatoes have been actively tested in a large number of studies. The most commonly used biocontrol agents belonged to various microbial genera including fungi (Aspergillus spp., Chaetomium spp., Glomus spp., non-pathogenic Fusarium spp., Trichoderma spp. and Penicillium spp.) and bacteria (Bacillus spp., Pseudomonas spp., Streptomyces spp., and Serratia spp.) [159].
Among the different genera of biocontrol microorganisms, non-pathogenic Fusarium strains are of high interest. In 1993, Alabouvette et al. [160] concluded that among the many groups of microorganisms tested for biocontrol activity, only non-pathogenic Fusarium species and fluorescent Pseudomonads showed consistent responses. In a later review by Ajilogba et al., these strains were found to be involved in most research conducted on plant biological enhancement using fungal endophytes [146]. One representative strain, F. oxysporum Fo47, was successfully tested against Fol [161,162,163], with the major mode of function being the induction of systemic resistance and priming of the plant defence reaction.
Another review by Raza et al. [159] analyzed biocontrol trials conducted between 2000 and 2014 and concluded that non-pathogenic Fusarium species and Pseudomonas species were supported by most research to be more effective in controlling Fusarium wilt in natural soil, while Penicillium, Streptomyces, and Aspergillus strains were more effective in growth media. However, the authors also found that 79% of the tests on tomatoes were conducted in greenhouse conditions, with 12% conducted in the field condition. Thus, for processing tomatoes grown predominately in field conditions, further field tests on the efficiency of different biocontrol agents are necessary.
Organic amendments are another group of biocontrol agents. For example, Borrego-Benjumea et al. [164] tested poultry manure, olive residue compost, and pelletised poultry manure for tomatoes grown in natural sandy soil and concluded that the combination of pelletized poultry manure with heating or solarization achieved the greatest reduction in Fusarium wilt severity. In a later study by Zhao et al. [165] testing chicken manure, rice straw, and vermicompost in a long-term tomato monocultural soil, vermicompost addition significantly increased soil pH, ammonium nitrogen, soil organic matter, and dissolved organic carbon, which promoted beneficial bacteria suppressing Fol. Organic amendments are often applied in combination with biocontrol microorganisms for better effects in different studies [159,166,167]. It was also suggested that the combined application of biocontrol organisms and amendments can increase the biocontrol efficiency of various genera of fungi and bacteria, with the exceptions of Pseudomonas and Penicillium [159].
3.4. Phytophthora Root Rot of Tomato
The oomycete P. capsici is the major pathogen causing Phytophthora root rot on tomatoes. This soilborne pathogen can invade tomatoes via root, inducing root and crown rot which can be visualized by the brown lesions on the plant’s lower part [168]. With sufficient rainfall, the whole plant can be infected. The root infection may cause damping-off of seedlings, stunting, wilting and eventual death in older plants [168]. The pathogen can also infect fruit in contact with the ground or via irrigation splash, and the disease, with symptoms of light-to-dark, water-soaked brown concentric rings on the fruit, is called buckeye rot instead [37,41,169].
As an oomycete, P. capsci can reproduce both sexual oospores with antheridium oogonium and asexual zoospores with sporangia [168,170,171]. Chlamydospores are also occasionally produced [168]. Most stages of P. capsci require the presence of a host, thus P. capsica seems only survive in the soil for long terms as thick-walled oospores [168]. The germination of oospores is facilitated by both chemical and mechanical stimulations, by either growing a germ tube or forming sporangium [168,171]. The sporangium then germinates directly or releases motile zoospores if immersed in water [170]. The zoospores are swimmable, which can be transmitted by rainfall or irrigation, and form germ tubes after contacting hosts [170].
3.4.1. Conventional Control Methods
Cultural Control
Crop rotation is often used to manage P. capsici along with many other soilborne pathogens, but its effectiveness is limited by the long survival of oomycetes in the soil and the wide host range of P. capsici. The host range of P. capsici was reported to cover at least 45 species of cultivated plants and weeds from 14 families of flowering plants [170], thus the selection of rotation crops for P. capsici is very narrow. Also, Lamour and Hausbeck [172] found P. capsici can survive as oospores for a 30-month nonhost period during crop rotation. Therefore, long rotations are required even if non-host crops are available, which may make crop rotations economically unfeasible.
It is very difficult to control P. capsici once the pathogen becomes established in the field. Thus, most control strategies are aimed at limiting free water to minimize inoculum spread and crop loss, which includes planting at well-drained sites or on a raised bed with controlled irrigation [168].
Physical Control
Soil solarization was found to be effective against Phytophthora root rot on tomatoes. From a trial in Florida a soil solarization treatment that heated the soil to a maximum of 47 °C at 10-cm depth had similar effects to MBr treatment at the same site in reducing the P. capsici population [107].
Chemical Control
The application of chemicals has been another approach to managing P. capsici. However, the phasing out of MBr has reduced the cost-efficiency of chemical control [173]. Other chemicals frequently applied include cyazofamid, dimethomorph, fluopicolide, fosetyl-Al, mandipropamid and mefenoxam (metalaxyl) [174,175,176,177]. Despite the various choices of chemicals, extensive use of fungicide has led to the emergence of resistant P. capsici strains, which makes it very hard to protect crops from P. capsici. For example, Lamour and Hausbeck [172] collected 141 isolates of P. capsici in Michigan and found around 60% to be intermediately sensitive or insensitive to mefenoxam. Even more recent groups of chemicals such as fluopicolide and cyazofamid have resulted in the fast emergence of pathogen resistance. Jackson et al. [175] concluded that among the 40 P. capsici isolates tested, all were either intermediately sensitive or resistant to cyazofamid at 100 μg/mL application rate. More recently, Siegenthaler and Hansen [177] found that out of 184 P. capsici isolates collected in Tennessee, 84 were resistant to fluopicolide.
Resistance Breeding
Until the 2010s, only several tomato strains moderately resistant to P. capsici were commercially available. Quesada-Ocampo and Hausbeck [173] screened 42 tomato cultivars and wild relatives for their resistance against P. capsici, and found Solanum habrochaites accession LA407, was resistant to all P. capsici isolates tested, with four additional cultivars having moderate resistance. However, the authors analyzed the genes of these cultivars and found a lack of correlation between genetic clusters and susceptibility to P. capsici, indicating that resistance was distributed in several tomato lineages. In a subsequent study, Quesada-Ocampo et al. [178] generated 62 backcross lines using LA407, and tested their resistance against different P. capsici strains and used annotated markers to locate genes related to the resistance. Though the researchers found that the resistance had a good inheritability among the population, they failed to find any annotated markers strongly associated with P. capsici resistance, with genes with annotation linked to disease resistance responses mapped to all chromosomes segregated among the population with the exceptions for 8, 9, 11, and 12. Therefore, the resistance of tomatos to P. capsici has not been related to specific gene/loci so far, and further studies are required.
3.4.2. Biocontrol
With insufficient levels of conventional control measures against Phytophthora root rot of tomatoes, antagonistic microbes and organic amendments have been tested to find feasible biocontrol approaches. Bacteria species are frequently studied for their biocontrol properties against Phytophthora root rot. Moataza [179] tested five Pseudomonas fluorescences strains against Rhizoctonia solani and P. capsici in tomato pot trials, and concluded that two strains, NRC1 and NRC3 had strong lytic activities leading to the destruction of the pathogens, but the method used in this research was seed coating, which may not be commercially feasible. In another study, Sharma et al. [180] tested 20 Bacillus strains against P. capsici on tomatoes grown in net house, and found one species, B. subtilis showed the best efficiency in terms of decreased disease severity. Furthermore, Syed-Ab-Rahman et al. [181] tested three bacteria- B. amyloliquefaciens, B. velezensis and Acinetobacter sp. on tomato, and concluded all three bacteria promoted tomato growth while significantly reducing the P. capsici load in their roots. An oomycete, Pythium oligandrum was also tested, and was believed to synthesize two Necrosis- and ethylene-inducing peptide 1 (Nep1)-like proteins PyolNLP5 and PyolNLP7, which induced the expression of antimicrobial tomato defensin genes against P. capsici [182].
The application of organic amendments is another approach to biocontrol. For P. capsici management, Nicol and Burlakoti [183] aerated compost and water and produced four aerobic compost teas. When tested in the glasshouse, the researchers concluded that if these products were drenched in potting mix before and after P. capsici inoculation, the disease progression was reduced by over 70%, with improved plant growth. Other efforts of using composts against P. capsici have generally been attempted on pepper [184,185,186], so the effects of these composts on tomatoes are unknown.
3.5. Pythium Root Rot and Damping-Off
The oomycete Pythium species tend to infect and cause rot of seeds, rootlets, root tips, and root hairs, with a preference for younger tissue at the root elongation zone and lateral roots [187]. The infection may cause small, brown, water-soaked lesions and can affect the entire root system [188]. With the focus on younger tissues, Pythium species infection often causes seedling damping-off at both pre- and post-germination stages [189,190], while infected older plants may also show stunted growth.
Pythium species overwinter in the soil as oospores or in plant debris as mycelium [191,192]. The germination of the oospore is facilitated by the exudates of germinating seeds and roots [192]. Similar to P. capsici, the oospore can produce a germ tube or form a sporangium, which germinates on its own or releases motile zoospores to contact and invade plant roots [191]. During the invasion, the oomycete hyphae release enzymes to destroy and feed on the host tissues [191]. After the invasion, Pythium species can either repeat the infection cycle in a new host with sporangia or survive as mycelia with sexual and asexual structures or dormant oospores until the next growing season [191,192].
3.5.1. Conventional Control Methods
Cultural Control
The application of pathogen-free seedlings and the control of irrigation are found to be effective forms for tomato Pythium disease management [193,194].
For Pythium species, crop rotation is generally not considered to be effective in the control of tomato infections because most Pythium species have a wide host range [195]. However, one study on wheat found that 3–4-year rotation cycles using wheat, canola and legume resulted in a significantly smaller disease incidence compared with less diverse rotations such as two-year wheat-canola [196]. The reason behind this finding may be that different crops have significantly different susceptibilities to Pythium infection, which may restrict the soilborne pathogen inoculum build-up after each crop, and eventually reducing the disease incidence in the next crop.
Physical Control
Soil solarization is an effective method for Pythium control with a long-period (six weeks to 60 days) of solarization during the summertime having been shown to significantly reduce the soilborne population of P. aphanidermatum in tropic zones [197,198]. In a field trial on tomatoes infected by Pythium spp., solarized soil showed a significantly lower mean damping-off incidence compared with un-solarized soil (2.15% compared with 68%) [199].
Chemical Control
Several chemicals have been used to manage Pythium species, including hymexazol, mefenoxam (metalaxyl), phosphonate, thiram and 8-Hydroxyquinoline [200,201,202,203,204]. The chemicals can be applied as seed treatment [205,206] or soil drenching [207] for seedlings of tomato.
In addition to the common economic and environmental concerns of chemical control, several major Pythium species collected from the production of various crops have developed resistance against several chemicals, especially mefenoxam. For example, Porter et al. [208] reported over 50% of the Pythium soil population consisted of mefenoxam-resistant isolates in ten of 64 potato fields from Oregon and Washington. Del Castillo Munera and Hausbeck [209] tested a total of 202 Pythium spp. isolates collected from Michigan, and found 39% of these, mostly P. ultimum and P. cylindrosporum isolates were intermediate to highly resistant to mefenoxam. For another major species P. irregulare, Aegerter et al. [210] tested four P. irregulare isolates from a greenhouse extensively applying mefenoxam and found no inhibition of growth of any isolate occurred at mefenoxam concentrations of 10 μg/mL or less. For other Pythium species such as P. aphanidermatum, resistance to mefenoxam was also reported [211,212]. In a rare case, Garzón et al. [203] even reported that the disease severity of a mefenoxam-resistant P. aphanidermatum on geranium can be stimulated by sublethal doses of mefenoxam.
Resistance Breeding
Though the deployment of resistant cultivars is a common and effective strategy for crop disease management, currently there is no Pythium-resistant tomato. The only potentially useful genetic resource against Pythium is the genes encoding pathogenesis-related (PR) proteins, with PR-1 protein showing antifungal activity against oomycetes [213]. Tomato has two related genes, PR1b1 and PR1a2, each encoding a basic and an acidic PR-1 protein [214], but the resistance of PR proteins is not pathogen-specific, with only limited effects against Pythium species.
3.5.2. Biocontrol
For biocontrol of Pythium disease on tomatoes, several bacteria strains have been studied. Postma et al. [215] tested four bacteria strains against P. aphanidermatum and found three strains, Pseudomonas chlororaphis, Peanibacillus polymyxa and Streptomyces pseudovenezuelae, significantly controlled P. aphanidermatum in under greenhouse conditions. The effect of Streptomyces bacteria was also supported by the study of Hassanisaadi et al. [195], who found two root-symbiont Streptomyces species significantly decreased disease incidence and improved performance of greenhouse tomato under P. aphanidermatum in stress out of the 116 tested species. For Bacillus bacteria, Martinez et al. [216] tested one B. subtilis strain MBI600 in a peat-based potting mix and concluded the addition of this strain significantly reduce tomato and sweet pepper damping-off and root rot while promoting root growth. Samaras et al. [204] also tested MBI600 on greenhouse tomatoes and concluded that the application of this strain achieved satisfactory control efficacy compared to chemical treatment with 8-Hydroxyquinoline.
For the application of fungal antagonists, the current focus seems to be on the Trichoderma species. Caron et al. [217] tested one local T. harzianum strain MAUL-20 on greenhouse tomatoes and found that it significantly reduced P. ultimum disease incidence, with a better effect compared with Rootshield™, a biofungicide based on T. harzianum KRL-AG2. Cuevas et al. [202] also tested T. parceramosum, T. pseudokoningii and T. harzianum respectively, and found the application of the Trichoderma pellets into the field before seeding can minimize the activity of Pythium spp., with a higher seed germination rate compared with the treatment using chemical fungicide mancozeb. Elshahawy and El-Mohamedy [188] tested the effects of five Trichoderma strains on P. aphanidermatum damping-off of tomatoes and concluded that under field conditions the combined application of the five isolates reduced by half the root rot severity while almost doubling the survival of tomato. This was thought to be through activating tomato defence enzymes and increasing leaf chlorophyll content, with an increased yield.
Interestingly, even arbuscular mycorrhizal fungi suppressing plant growth may also be used to control Pythium species. Larsen et al. [218] pre-treated greenhouse tomato seedlings with Glomus intraradices, G. mosseae, G. claroideum, and then challenged the seedlings with P. aphanidermatum, with the hypothesis that the application of growth-suppressive fungi may trigger plant defence response in terms of PR-1 expression to prepare the plants for Pythium infection. However, the application of arbuscular mycorrhizal fungi did not affect PR-1 gene expression, with only G. intraradices reducing the pathogen root infection level of P. aphanidermatum, thus the hypothesis was not confirmed.
Several organic amendments have also been tested against Pythium, such as canola residues and composts (animal bone charcoal, compost tea, solid green wastes, or green waste +manure) [215,219,220,221]. Also, Jayaraj et al. [222] found that formulating amendments such as lignite with biocontrol agents such as B. subtilis can greatly increase their shelf life, with good effects on Pythium suppression and plant growth promotion.
3.6. Tomato Verticillium Wilt
Verticillium. dahliae and V. albo-atrum are soilborne fungi that can induce vascular wilt diseases in over 200 dicotyledonous species, including those economically important such as tomato [223]. These species invade the host through roots [224,225] and then attack the vascular system via xylem vessels. This leads to wilting, vascular discoloration, early senescence, and the eventual death of the infected plant [224].
Though the two Verticillium species have similar lifecycles, V. dahliae causes monocyclic disease with only one disease cycle in a growing season [224]. In contrast, V. albo-atrum can produce conidia on infected plant tissues, which can be airborne and contribute to polycyclic diseases during one growing season [224]. The lifecycles of the two species both have a dormant, a parasitic, and a saprophytic stage [224]. During the dormant stage, the resting structures such as microsclerotia and mycelium in soil or plant debris are under microbiostasis or mycostasis and unable to germinate [224]. The germination of the pathogens is stimulated by root exudates from both host and non-host plants [226]. The pathogens then enter the parasitic stage by invading hosts through the root tip or elongation region to invade the xylem and vascular systems [226]. After the necrosis of the infected tissue, the saprophytic stage begins, in which the pathogens extend their colonization to shoots and roots and produce conidia or microsclerotia to repeat the cycle [224]. The pathogens can be spread by the transport of infected planting stock or by soil cultivation and soil movement by wind or water [226].
3.6.1. Conventional Control Methods
Cultural Control
Crop rotation with non-host crops is an effective strategy for Verticillium wilt management. The known non-host crops include small grain crops such as wheat and corn [227], and long rotations lasting over four years are recommended [44].
Hygiene is also important for Verticillium wilt control. pathogen-free seed and disease-free transplants should be used [44], with infected crop debris removed and destroyed away from the field. Equipment and foot ware should be washed to prevent the movement of infested soil between fields. Verticillium also prefers humid soil, thus maintaining well-drained soil, and eliminating excessive soil moisture may also limit the development of the pathogen [228].
Physical Control
Verticillium prefers cool temperatures for survival and developing symptoms, thus heating the soil through solarization could be an effective control method. Currently, solarization against Verticillium wilt is practiced generally in Mediterranean, desert, and tropical climates, because these climates allow the accumulation of adequate heat to neutralize the pathogen [229]. However, the data on solarization alone showed poorer performance compared with the MBr application, which can be improved when combined with the fumigation using MBr alternatives [230].
Chemical Control
Soil fumigation is also used to control Verticillium wilt. MBr alternatives such as chloropicrin (CP) (trichloronitromethane) are traditionally used as in formulations together with MBr to achieve a broader spectrum of activity [230]. In a trial by Gullino et al. [72], CP applied by shank injection at rates ≥30 g/m2 induced a satisfactory and consistent control of tomato Verticillium wilt, with no phytotoxicity, but the efficiency was slightly lower than standard MBr application and may have been influenced by soil type and organic matter content. Metam-sodium and 1,3-dichloropropene are other alternative soil fumigants, which have been applied in combination or with metam-sodium alone in the United States to reduce soil populations of V. dahliae [231]. Several other chemicals such as fungicides including azoxystrobin, benomyl, captan, thiram, and trifloxystrobin, and a plant defense activator, acibenzolar-S-methyl were also recommended [116,230,232].
Resistance Breeding
By far, the most feasible and economic control for Verticillium wilt is the application of resistant cultivars. The resistance gene in tomato to V. dahliae was first identified as a single dominant factor in the reciprocal crosses between the wilt-resistant variety W6 (Peru Wild × Century) and Moscow, a susceptible variety, and named as Ve in 1951 [233]. Ve was found to be a locus, which contains two genes, Ve1 and Ve2, with only Ve1 found to mediate resistance in tomato [223]. The strains of V. dahliae resistant to Ve1 and V. albo-atrum were assigned to race 2 [223]. The Ve1 gene has been incorporated into many commercial cultivars. However, all the current verticillium-resistant gene resources are against V. dahliae race 1, thus all race 2 strains of V. dahliae and V. albo-atrum can still infect the resistant cultivars.
3.6.2. Biocontrol
Biological control may be a promising method to control Verticillium wilt, given that most current management methods have limited efficiency. Various microorganisms have been tested against V. dahliae, such as bacteria Bacillus subtilis and B. velezensis [234], and fungi including Burkholderia gladioli [235], Gliocladium spp., Penicillium sp. [236,237], Trichoderma spp. [238], Talaromyces flavus [239], and even V. klebahnii and V. isaacii with low pathogenicity [240]. Though most of the microorganisms are found to be effective in trials, most of the trials were carried out in greenhouses or with sterilized soil, with only a few verified in field conditions. Larena et al. [237] conducted a field assay using P. oxalicum and concluded that seedlings needed to be treated with 106–107 CFU/g of the biocontrol agent around a week before transplanting to achieve a sufficient level of control, but only in a certain soil type (loam soil, pH = 7.0), and the formulation may not be feasible for tomato mass production due to the high CFU density requirement.
The application of organic amendments is known as another approach for crop disease biocontrol. It has long been known that bloodmeal and fishmeal can eliminate the incidence of Verticillium wilt in tomato [230]. Compared to animal-based amendments (manure), plant-based amendments not only support beneficial microbial activities but also have greater efficiency on pathogens due to deleterious chemicals produced by the plants, in addition to supporting beneficial microbial activities [241]. Giotis et al. [12] concluded that fresh Brassica tissue, household waste compost, and composted cow manure significantly reduced soilborne disease severity of tomato Verticillium wilt, with enhanced plant growth. Similar results were also achieved by Kadoglidou et al. [242], who applied soil incorporated spearmint and oregano-dried plant material, which caused disease suppression resulting in increased fruit yields of tomatoes inoculated with V. dahliae. Moreover, Ait Rahou et al. [243] used compost based on green waste (quackgrass) to greenhouse tomatoes inoculated with Verticillium and concluded that growth regulators directly produced by the microorganisms in the compost improved plant growth significantly. However, when Lazarovits et al. [244] applied compost made from sewage sludge to suppress V. dahliae in tomato plants, phytotoxicity was detected over one month, which may have been due to the excessive accumulation of plant-toxic heavy metals in soils. To conclude, though organic amendments may be useful for Verticillium wilt management, they may also carry toxic compounds which may lead to undesired effects.
4. Conclusions and Future Perspective
Among the field processing tomato producing countries covered in this review, the major soilborne fungal/oomycete pathogens affecting their tomato production are Fol, Forl, P. lycopersici and P. capsici, with the pathogenic Verticillium spp., and Pythium spp. being also important in certain countries. The various management methods (Table 1) generally have variable levels of effectiveness on the diseases (Table 2). For cultural controls, hygiene is fundamental to disease control, while the effectiveness of crop rotation is affected by the host range and longevity of the corresponding pathogen in soil and crop debris. Physical control represented by soil solarization is generally effective except for Fol which can withstand high temperatures, but the level of success is affected by several environmental and biological factors, thus it may work better as a part of integrated disease management. The situation of chemical control is more problematic. The industry used to rely heavily on the broad-spectrum, cost-efficient MBr soil fumigation effective against all soilborne pathogens, but it turned out to be a lose-lose. MBr heavily damaged the ozone layer, which resulted in it being banned globally, while the ban in turn caused enormous losses to the industry. For example, Cao et al. [132] estimated that in Florida, the phase-out of MBr and the introduction of replacements may have caused a 20% tomato yield reduction and a $1656 decrease in profit per acre, which substantially harmed the competitiveness and sustainability of the tomato industry of Florida. As discussed above, alternative chemicals are available for the control of all diseases, but these generally have reduced spectrum and cost-efficiency, and in some cases, the pathogens have already developed certain levels of resistance. The situation is further exacerbated by the decreasing public acceptance of the chemical application, and the fact that even some alternatives to MBr, such as metam sodium, have been banned or are scheduled for a phase-out in certain areas [245]. Breeding for disease resistance remains highly effective, but it is traditionally laborious and time-consuming, and genes for complete resistance to P. lycopersici, Pythium sp., V. dahilae race 2, and V. albo-atrum have not been identified.

Table 1.
Different management methods and their examples are mentioned in this review.
Table 2.
Advantages and disadvantages of the management methods concluded from this review.
Though relatively new for crop disease management, biological control seems to show promise. Although most biocontrol agents are still in the greenhouse trial stage, assessment has shown satisfying levels of disease control, many of which directly attack pathogens or initiate the plant’s own defence mechanisms, and some directly improve the growth and development of crops. Also, the use of organic amendments may not only improve the plant growth, but also utilize unwanted organic products, which may go to waste otherwise. Therefore, the advantages of biocontrol may minimize the adverse effects on the environment while being highly appealing to the public.
Disease management has long been challenging for the tomato industry. Cultivated tomato is considered to have low genetic diversity due to the population bottleneck effect of domestication and artificial selection [246], making them vulnerable to destructive pathogens. Also, temperate climates and adequate humidity is preferred by cultivated tomatoes [32], which may also facilitate the flourishing of soilborne microorganisms, including pathogens. Moreover, the fast development of international trade in recent years has brought new threats by allowing the emergence of pathogens into new geographical locations through transporting infected products [247], which can adapt to the new environment via mutation or merging with their local native relatives. Also, climate change may also facilitate the natural movement of pathogens. As demonstrated by Bebber et al. [248], global warming has made crop pathogens move poleward, affecting the zones that used to be too cold for them to survive. More importantly, climate change is particularly challenging for the processing tomato industry. Cammarano et al. [249] made predictions of global tomato yield based on five potential future global warming scenarios and concluded that around 6% decline in processing tomato production may take place in the three major producing areas (California, China, and Italy) by 2050 due to the increased temperature, with little differences between the scenarios.
With conventional disease management strategies becoming inadequate for the challenges brought about by pathogen resistance, global trade, and climate change, innovative control strategies are needed. Biocontrol, with its good potential in disease control efficiency, public acceptance, and crop yield improvement, should be incorporated into the integrated disease management of field processing tomato. This will complement cultural practices, physical disease management, and modern breeding techniques such as gene editing and CRISPR/Cas, to develop management practices emphasizing sustainability and the security of both food and the environment, and hence greatly reduced reliance on synthesized chemical application.
Author Contributions
Conceptualization, M.M., P.W.J.T. and J.-Z.H.; resources, P.W.J.T., D.C. and J.-Z.H.; writing—original draft preparation, M.M.; writing—review and editing, all authors; supervision, P.W.J.T., D.C., N.V. and J.-Z.H.; project administration, M.M., P.W.J.T. and J.-Z.H.; funding acquisition, D.C. and J.-Z.H. All authors have read and agreed to the published version of the manuscript.
Funding
ARC Research Hub for Smart Fertilizers (IH200100023).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rick, C.M. The tomato. Sci. Am. 1978, 239, 76–89. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#home (accessed on 30 November 2022).
- Barringer, S. Canned tomatoes: Production and storage. In Handbook of Vegetable Preservation and Processing, 1st ed.; Hui, Y.H., Ghazala, S., Graham, D.M., Murrell, K.D., Nip, W., Eds.; CRC Press: New York, NY, USA, 2004; pp. 109–120. [Google Scholar]
- WPTC. WPTC Crop Update as of 25 October 2022. Available online: https://www.tomatonews.com/force_doc.php?file=c50d46545467b815c44e583ddcdd7731c1c4ae2f.pdf (accessed on 30 November 2022).
- Luster, C., III. A rapid and sensitive sterility monitoring technique for aseptically processed bulk tomato paste. J. Food Sci. 1978, 43, 1046–1048. [Google Scholar] [CrossRef]
- Barrett, D.M. Future innovations in tomato processing. In Proceedings of the XIII International Symposium on Processing Tomato, Sirmione, Italy, 8 June 2014; Volume 1081, pp. 49–55. [Google Scholar]
- Johnstone, P.R.; Hartz, T.K.; LeStrange, M.; Nunez, J.J.; Miyao, E.M. Managing fruit soluble solids with late-season deficit irrigation in drip-irrigated processing tomato production. HortScience 2005, 40, 1857–1861. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, A.K.; Kumar, A. Disease management of tomato through PGPB: Current trends and future perspective. 3 Biotech 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Bawa, I. Management strategies of Fusarium wilt disease of tomato incited by Fusarium oxysporum f. sp. lycopersici(Sacc.) A Review. Int. J. Adv. Acad. Res. 2016, 2, 32–42. [Google Scholar]
- Valenzuela-Lopez, N.; Cano-Lira, J.F.; Guarro, J.; Sutton, D.A.; Wiederhold, N.; Crous, P.W.; Stchigel, A.M. Coelomycetous Dothideomycetes with emphasis on the families Cucurbitariaceae and Didymellaceae. Stud. Mycol. 2018, 90, 1–69. [Google Scholar] [CrossRef]
- d’Errico, G.; Marra, R.; Crescenzi, A.; Davino, S.W.; Fanigliulo, A.; Woo, S.L.; Lorito, M. Integrated management strategies of Meloidogyne incognita and Pseudopyrenochaeta lycopersici on tomato using a Bacillus firmus-based product and two synthetic nematicides in two consecutive crop cycles in greenhouse. Crop. Prot. 2019, 122, 159–164. [Google Scholar] [CrossRef]
- Giotis, C.; Markelou, E.; Theodoropoulou, A.; Toufexi, E.; Hodson, R.; Shotton, P.; Leifert, C. Effect of soil amendments and biological control agents (BCAs) on soil-borne root diseases caused by Pyrenochaeta lycopersici and Verticillium albo-atrum in organic greenhouse tomato production systems. Eur. J. Plant Pathol. 2009, 123, 387–400. [Google Scholar] [CrossRef]
- Cheng, H.; Zhang, D.; Ren, L.; Song, Z.; Li, Q.; Wu, J.; Cao, A. Bio-activation of soil with beneficial microbes after soil fumigation reduces soil-borne pathogens and increases tomato yield. Environ. Pollut. 2021, 283, 117160. [Google Scholar] [CrossRef]
- Baysal-Gurel, F.; Gardener, B.M.; Miller, S.A. Soil Borne Disease Management in Organic Vegetable Production. Org. Agric. Available online: https://eorganic.org/node/7581. (accessed on 30 November 2022).
- Spadaro, D.; Gullino, M.L. Improving the efficacy of biocontrol agents against soilborne pathogens. Crop. Prot. 2005, 24, 601–613. [Google Scholar] [CrossRef]
- Gareau, B.J. Lessons from the Montreal Protocol delay in phasing out methyl bromide. J. Environ. Stud. Sci. 2015, 5, 163–168. [Google Scholar] [CrossRef]
- Piel, C.; Pouchieu, C.; Carles, C.; Beziat, B.; Boulanger, M.; Bureau, M.; Baldi, I. Agricultural exposures to carbamate herbicides and fungicides and central nervous system tumour incidence in the cohort AGRICAN. Environ. Int. 2019, 130, 104876. [Google Scholar] [CrossRef]
- Foolad, M.R.; Merk, H.L.; Ashrafi, H. Genetics, genomics and breeding of late blight and early blight resistance in tomato. Crit. Rev. Plant Sci. 2008, 27, 75–107. [Google Scholar] [CrossRef]
- Menda, N.; Strickler, S.R.; Edwards, J.D.; Bombarely, A.; Dunham, D.M.; Martin, G.B.; Mueller, L.A. Analysis of wild-species introgressions in tomato inbreds uncovers ancestral origins. BMC Plant Biol. 2014, 14, 1–16. [Google Scholar] [CrossRef] [PubMed]
- de Toledo Thomazella, D.P.; Brail, Q.; Dahlbeck, D.; Staskawicz, B. CRISPR-Cas9 mediated mutagenesis of a DMR6 ortholog in tomato confers broad-spectrum disease resistance. BioRxiv. Available online: https://www.biorxiv.org/content/10.1101/064824v3.full. (accessed on 30 November 2022).
- Panthee, D.R.; Brown, A.F.; Yousef, G.G.; Ibrahem, R.; Anderson, C. Novel molecular marker associated with Tm2a gene conferring resistance to tomato mosaic virus in tomato. Plant Breed. 2013, 132, 413–416. [Google Scholar] [CrossRef]
- Flint, M.L.; Dreistadt, S.H. Natural Enemies Handbook: The Illustrated Guide to Biological Pest Control; Univ of California Press: Berkely, CA, USA, 1998; Volume 3386, pp. 3–6. [Google Scholar]
- Landis, D.A.; Orr, D.B. Biological Control: Approaches and Applications. Electronic IPM Textbook. Available online: https://ipmworld.umn.edu/landis (accessed on 30 November 2022).
- Rechcigl, J.E.; Rechcigl, N.A. (Eds.) Biological and Biotechnological Control of Insect Pests; CRC Press: New York, NY, USA, 1999; pp. 3–5. [Google Scholar]
- van Lenteren, J.C.; Bolckmans, K.; Köhl, J.; Ravensberg, W.J.; Urbaneja, A. Biological control using invertebrates and microorganisms: Plenty of new opportunities. BioControl 2018, 63, 39–59. [Google Scholar] [CrossRef]
- Singh, S.; Bhatnagar, S.; Choudhary, S.; Nirwan, B.; Sharma, K. Fungi as biocontrol agent: An alternate to chemicals. In Fungi and Their Role in Sustainable Development: Current Perspectives, 1st ed.; Gehlot, P., Singh, J., Eds.; Springer: Singapore, 2018; pp. 23–33. [Google Scholar]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Sharma, A. Trichoderma: The “secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Junaid, J.M.; Dar, N.A.; Bhat, T.A.; Bhat, A.H.; Bhat, M.A. Commercial biocontrol agents and their mechanism of action in the management of plant pathogens. Int. J. Mod. Plant Anim. Sci. 2013, 1, 39–57. [Google Scholar]
- Hort Innovation. Tomato Topics September 2022. Available online: https://www.aptrc.asn.au/wp-content/uploads/2022/10/Tomato-Topics-September-2022-1.pdf (accessed on 30 November 2022).
- Pathak, T.B.; Stoddard, C.S. Climate change effects on the processing tomato growing season in California using growing degree day model. Model. Earth Syst. Environ. 2018, 4, 765–775. [Google Scholar] [CrossRef]
- Jones, J.B., Jr. Tomato Plant Culture: In the Field, Greenhouse, and Home Garden; CRC Press: New York, NY, USA, 2007; pp. 55–80. [Google Scholar]
- Costa, J.M.; Heuvelink, E.P. The global tomato industry. In Tomatoes, 2nd ed.; Heuvelink, E., Ed.; CABI: Cambridge, MA, USA, 2018; pp. 1–26. [Google Scholar]
- Hartz, T.; Miyao, G.; Mickler, J.; Lestrange, M.; Stoddard, S.; Nuñez, J.; Aegerter, B. Processing Tomato Production in California. Available online: https://escholarship.org/content/qt4hc350c9/qt4hc350c9.pdf (accessed on 30 November 2022).
- The Top 10 Tomato Producing States in the United States. Available online: https://www.worldatlas.com/articles/the-top-10-tomato-producing-states-in-the-united-states.html (accessed on 30 November 2022).
- Guan, Z.; Biswas, T.; Wu, F. The US Tomato Industry: An Overview of Production and Trade. Available online: https://journals.flvc.org/edis/article/download/105009/118652/ (accessed on 30 November 2022).
- Baskins, S.; Bond, J.; Minor, T. Unpacking the Growth in per Capita Availability of Fresh Market Tomatoes. Available online: https://www.ers.usda.gov/webdocs/outlooks/92442/vgs-19c-01.pdf?v=5296.1 (accessed on 30 November 2022).
- Fernández-Pavía, S.P.; Rodríguez-Alvarado, G.; Sánchez-Yáñez, J.M. Buckeye rot of tomato caused by Phytophthora capsici in Michoacan, Mexico. Plant Dis. 2003, 87, 872. [Google Scholar] [CrossRef]
- Gupta, S.K.; Sachin, U.; Sharma, R.C. Biology, epidemiology and management of buckeye rot of tomato. In Challenging Problems in Horticultural and Forest Pathology, 1st ed.; Sharma, R.C., Sarma, G.N., Eds.; Indus Publishing Company: New Delhi, India, 2006; pp. 183–199. [Google Scholar]
- Hoy, M.W.; Ogawa, J.M.; Duniway, J.M. Effects of irrigation on buckeye rot of tomato fruit caused by Phytophthora parasitica. Phytopathology 1984, 74, 474–478. [Google Scholar] [CrossRef]
- Kendrick, J.B. Phytophthora rot of tomato, eggplant, and pepper. Proc. Indiana Acad. Sci. 1922, 32, 299–306. [Google Scholar]
- Tompkins, C.M.; Tucker, C.M. Buckeye rot of tomato in California. J. Agric. Res. 1941, 62, 467–474. [Google Scholar]
- Bolkan, H. A technique to evaluate tomatoes for resistance to Phytophthora root rot in the greenhouse. Plant Dis. 1985, 69, 708–709. [Google Scholar] [CrossRef]
- Filho, A.C.; Duniway, J.M. Dispersal of Phytophthora capsici and P. parasitica in furrow-irrigated rows of bell pepper, tomato and squash. Plant Pathol. 1995, 44, 1025–1032. [Google Scholar] [CrossRef]
- Babadoost, M. Important fungal diseases of tomato in the United States of America. In Proceedings of the III International Symposium on Tomato Diseases, Ischia, Italy, 25–30 July 2010; Volume 914, pp. 85–92. [Google Scholar]
- Ioannou, N.; Grogan, R.G. Control of Phytophthora root rot of processing tomato with ethazol and metalaxyl. Plant Dis. 1984, 68, 429–435. [Google Scholar] [CrossRef]
- Stamova, L. Resistance to Two Important Tomato Diseases in California. In Proceedings of the XV Meeting of the EUCARPIA Tomato Working Group, Bari, Italy, 20–23 September 2005; Volume 789, pp. 87–94. [Google Scholar]
- Campbell, R.N. Corky Root of Tomato in California Caused by Pyrenochaeta lycopersici and Control by Soil Fumigation. Plant Dis. 1981, 66, 657–661. [Google Scholar] [CrossRef]
- Workneh, F.; Van Bruggen AH, C.; Drinkwater, L.E.; Shennan, C. Variables associated with corky root and Phytophthora root rot of tomatoes in organic and conventional farms. Phytopathology 1993, 83, 581–589. [Google Scholar] [CrossRef]
- Vrisman, C.M.; Testen, A.L.; Elahi, F.; Miller, S.A. First report of tomato brown root rot complex caused by Colletotrichum coccodes and Pyrenochaeta lycopersici in Ohio. Plant Dis. 2017, 101, 247. [Google Scholar] [CrossRef]
- Srinivas, C.; Devi, D.N.; Murthy, K.N.; Mohan, C.D.; Lakshmeesha, T.R.; Singh, B.; Srivastava, R.K. Fusarium oxysporum f. sp. lycopersici causal agent of vascular wilt disease of tomato: Biology to diversity–A review. Saudi J. Biol. Sci. 2019, 26, 1315–1324. [Google Scholar] [CrossRef]
- Alexander, L. Physiologic specialization in the tomato wilt. J. Agric. Res. 1945, 70, 303. [Google Scholar]
- Cai, G.H.; Gale, L.R.; Schneider, R.W.; Kistler, H.C.; Davis, R.M.; Elias, K.S.; Miyao, E.M. Origin of race 3 of Fusarium oxysporum f. sp. lycopersici at a single site in California. Phytopathology 2003, 93, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Volin, R.B.; Jones, J.P. A new race of Fusarium wilt of tomato in Florida and sources of resistance. Proc. Fla. State Hortic. Soc. 1982, 95, 268–269. [Google Scholar]
- Davis, R.M.; Kimble, K.A.; Farrar, J.J. A third race of Fusarium oxysporum f. sp. lycopersici identified in California. Plant Dis. 1988, 72, 453. [Google Scholar] [CrossRef]
- Benhamou, N.; Charest, P.M.; Jarvis, W.R. Biology and Host-Parasite Relations of Fusarium oxysporum f. sp. radicis-lycopersici. In Vascular Wilt Diseases of Plants, 1st ed.; Tjamos, E.C., Beckman, C.H., Eds.; Springer: Berlin, Germany, 1986; pp. 95–105. [Google Scholar]
- Ozbay, N.; Newman, S.E. Fusarium crown and root rot of tomato and control methods. Plant Pathol. J. 2004, 3, 9–18. [Google Scholar] [CrossRef]
- Rudolph, B.A. Verticillium wilt of tomatoes in California. sclerotia of Verticillium albo-atrum and its influence on infection. Phytopathology 1926, 16, 234. [Google Scholar]
- Alexander, L.J. Susceptibility of certain Verticillium-resistant Tomato varieties to an Ohio isolate of the pathogen. Phytopathology 1962, 52, 998–1000. [Google Scholar]
- Grogan, R.G.; Ioannou, N.; Schneider, R.W.; Sall, M.A.; Kimble, K.A. Verticillium wilt on resistant tomato cultivars in California: Virulence of isolates from plants and soil and relationship of inoculum density to disease incidence. Phytopathology 1979, 69, 1176–1180. [Google Scholar] [CrossRef]
- Ashworth, L.J.; Huisman, O.C.; Harper, D.M.; Stromberg, L.K. Verticillium wilt disease of tomato: Influence of inoculum density and root extension upon disease severity. Phytopathology 1979, 69, 490–492. [Google Scholar] [CrossRef]
- Buller, S.; Inglis, D.; Miles, C. Plant growth, fruit yield and quality, and tolerance to verticillium wilt of grafted watermelon and tomato in field production in the Pacific Northwest. HortScience 2013, 48, 1003–1009. [Google Scholar] [CrossRef]
- Costa, J.M.; Heuvelink, E. Introduction: The tomato crop and industry. In Tomatoes, 1st ed.; Heuvelink, E., Ed.; CABI: Cambridge, MA, USA, 2005; pp. 1–20. [Google Scholar]
- Zhang, X.X.; Qiu, H.; Huang, Z. Farm structure of the apple and tomato production in the EU and China. In Apple and Tomato Chains in China and the EU. LEI 26, 1st ed.; Zhang, X.X., Qiu, H., Huang, Z., Eds.; LEI: The Hague, The Netherlands, 2010; pp. 21–27. [Google Scholar]
- Wang, Y.; Zhang, Y.; Gao, Z.; Yang, W. Breeding for resistance to tomato bacterial diseases in China: Challenges and prospects. Hortic. Plant J. 2018, 4, 193–207. [Google Scholar] [CrossRef]
- Chang, Y.D.; Du, B.; Wang, L.; Ji, P.; Xie, Y.J.; Li, X.F.; Wang, J.M. A study on the pathogen species and physiological races of tomato Fusarium wilt in Shanxi, China. J. Integr. Agric. 2018, 17, 1380–1390. [Google Scholar] [CrossRef]
- Xu, Z.; Huang, K.; Shou, W.; Zhou, S.; Li, G.; Chen, L.; Jin, B. The determination of physiological race of Fusarium oxysporum f. sp. lycopersici of tomato in Zhejiang, China. Acta Physiol. Plant. 2000, 22, 356–358. [Google Scholar] [CrossRef]
- Ye, Q.; Wang, R.; Ruan, M.; Yao, Z.; Cheng, Y.; Wan, H.; Zhou, G. Genetic diversity and identification of wilt and root rot pathogens of tomato in China. Plant Dis. 2020, 104, 1715–1724. [Google Scholar] [CrossRef]
- Perrotta, D. Processing tomatoes in the era of the retailing revolution: Mechanization and migrant labour in northern and southern Italy. In Migration and Agriculture, 1st ed.; Corrado, A., de Castro, C., Perrotta, D., Eds.; Routledge: Oxfordshire, UK, 2016; pp. 82–100. [Google Scholar]
- Vitale, A.; Castello, I.; Cascone, G.; D’Emilio, A.; Mazzarella, R.; Polizzi, G. Reduction of corky root infections on greenhouse tomato crops by soil solarization in South Italy. Plant Dis. 2011, 95, 195–201. [Google Scholar] [CrossRef]
- Garibaldi, A.; Gilardi, G.; Baudino, M.; Ortu, G.; Gullino, M.L. Phytophthora capsici: A soilborne pathogen dangerous on grafted tomato (Solanum lycopersicum × S. hirsutum) in Italy. Plant Dis. 2012, 96, 1830. [Google Scholar] [CrossRef]
- Stravato, V.M.; Buonaurio, R.; Cappelli, C. First Report of Fusarium oxysporum f. sp. lycopersici Race 2 on Tomato in Italy. Plant Dis. 1999, 83, 967. [Google Scholar] [CrossRef]
- Gullino, M.L.; Minuto, A.; Gilardi, G.; Garibaldi, A.; Ajwa, H.; Duafala, T. Efficacy of preplant soil fumigation with chloropicrin for tomato production in Italy. Crop Prot. 2002, 21, 741–749. [Google Scholar] [CrossRef]
- Rongai, D.; Pulcini, P.; Pesce, B.; Milano, F. Antifungal activity of pomegranate peel extract against fusarium wilt of tomato. Eur. J. Plant Pathol. 2017, 147, 229–238. [Google Scholar] [CrossRef]
- Srinivasan, K.; Gilardi, G.; Garibaldi, A.; Gullino, M.L. Bacterial antagonists from used rockwool soilless substrates suppress Fusarium wilt of tomato. J. Plant Pathol. 2009, 91, 147–154. [Google Scholar]
- Di Primo, P.; Cartia, G.; Katan, T. Vegetative compatibility and heterokaryon stability in Fusarium oxysporum f. sp. radicis-lycopersici from Italy. Plant Pathol. 2001, 50, 371–382. [Google Scholar] [CrossRef]
- Garibaldi, A.; Minuto, A.; Minuto, G. Influence of Recycled Substrates on the Severity of Crown Rot on Soilless Tomato. In Proceedings of the XV Meeting of the EUCARPIA Tomato Working Group, Bari, Italy, 20–23 September 2005; Volume 789, pp. 167–170. [Google Scholar]
- Cartia, G.; Asero, C. Investigation on spread and pathogenicity of Fusarium oxysporum f. sp. radicis lycopersici in Sicilian greenhouses. Colt. Protette 1994, 23, 75–78. [Google Scholar]
- Franceschini, A.; Maddau, L.; Corda, P.; Ionta, G. Observations on Fusarium crown and root rot of tomato and its control in Sardinia (Italy). Dif. Delle Piante 1996, 19, 125–138. [Google Scholar]
- Frenzel, B. Atlas of Paleoclimates and Paleoenvironments of the Northern Hemisphere. Available online: https://epic.awi.de/id/eprint/29922/1/Fre1992a.pdf (accessed on 30 November 2022).
- Elias, M.A.; Borges, F.J.; Bergamini, L.L.; Franceschinelli, E.V.; Sujii, E.R. Climate change threatens pollination services in tomato crops in Brazil. Agric. Ecosyst. Environ. 2017, 239, 257–264. [Google Scholar] [CrossRef]
- Marouelli, W.A.; Silva, W.L. Water tension thresholds for processing tomatoes under drip irrigation in Central Brazil. Irrig. Sci. 2007, 25, 411–418. [Google Scholar] [CrossRef]
- Inoue-Nagata, A.K.; Lima, M.F.; Gilbertson, R.L. A review of geminivirus diseases in vegetables and other crops in Brazil: Current status and approaches for management. Hortic. Bras. 2016, 34, 8–18. [Google Scholar] [CrossRef]
- Reis, A.; Costa, H.; Boiteux, L.S.; Lopes, C.A. First report of Fusarium oxysporum f. sp. lycopersici race 3 on tomato in Brazil. Fitopatol. Bras. 2005, 30, 426–428. [Google Scholar] [CrossRef]
- Reis, A.; Boiteux, L.S. Outbreak of Fusarium oxysporum f. sp. lycopersici race 3 in commercial fresh-market tomato fields in Rio de Janeiro State, Brazil. Hortic. Bras. 2007, 25, 451–454. [Google Scholar] [CrossRef]
- Gonçalves, A.M.; Costa, H.; Fonseca, M.E.N.; Boiteux, L.S.; Lopes, C.A.; Reis, A. Variability and geographical distribution of Fusarium oxysporum f. sp. lycopersici physiological races and field performance of resistant sources in Brazil. In Proceedings of the V International Symposium on Tomato Diseases: Perspectives and Future Directions in Tomato Protection, Málaga, Spain, 13–16 June 2016; Volume 1207, pp. 45–50. [Google Scholar]
- Cabral, C.S.; Gonçalves, A.M.; Fonseca ME, N.; Urben, A.F.; Costa, H.; Lourenço, V.; Reis, A. First detection of Fusarium oxysporum f. sp. radicis–lycopersici across major tomato–producing regions in Brazil. Phytoparasitica 2020, 48, 545–553. [Google Scholar] [CrossRef]
- Suaste-Dzul, A.P.; Costa, H.; Fonseca ME, N.; Boiteux, L.S.; Reis, A. Mating types and physiological races of Verticillium dahliae in Solanaceae crops in Brazil. Eur. J. Plant Pathol. 2022, 164, 1–14. [Google Scholar] [CrossRef]
- Suaste-Dzul, A.P.; Veloso, J.S.; Costa, H.; Boiteux, L.S.; Lourenço Jr, V.; Lopes, C.A.; Reis, A. Verticillium diseases of vegetable crops in Brazil: Host range, microsclerotia production, molecular haplotype network, and pathogen species determination. Plant Pathol. 2022, 71, 1417–1430. [Google Scholar] [CrossRef]
- Valdés, V. The tomato industry in Chile. In Proceedings of the IV International Symposium on Processing Tomatoes, Mendoza, Argentina, 18–21 February 1991; Volume 301, pp. 59–62. [Google Scholar]
- Valenzuela, M.; Fuentes, B.; Alfaro, J.F.; Galvez, E.; Salinas, A.; Besoain, X.A.; Seeger, M. First Report of bacterial speck caused by Pseudomonas syringae pv. tomato Race 1 affecting tomato in different Regions of Chile. Plant Dis. 2021, 106, 1979. [Google Scholar] [CrossRef] [PubMed]
- Saavedra Del, R.G.; Escaff, G.M.; Cortacáns, P.D.; Ruiz-Tagle, C. Recent developments in processing tomato production in Chile. In Proceedings of the IX International Symposium on the Processing Tomato, Melbourne, Australia, 15–18 November 2004; Volume 724, pp. 335–338. [Google Scholar]
- Sepúlveda-Chavera, G.; Huanca, W.; Salvatierra-Martínez, R.; Latorre, B.A. First report of Fusarium oxysporum f. sp. lycopersici race 3 and F. oxysporum f. sp. radicis-lycopersici in tomatoes in the Azapa Valley of Chile. Plant Dis. 2014, 98, 1432. [Google Scholar] [CrossRef] [PubMed]
- Elizondo-Pasten, E.; Boix-Ruiz, A.; Gómez-Tenorio, M.A.; Ruiz-Olmos, C.; Marín-Guirao, J.I.; Tello-Marquina, J.C.; Camacho-Ferre, F. Two complementary techniques allow detection of Fusarium oxysporum f. sp. radicis-lycopersici in soils from two different tomato-cultivated areas of Chile. In Proceedings of the V International Symposium on Tomato Diseases: Perspectives and Future Directions in Tomato Protection, Málaga, Spain, 13–16 June 2016; Volume 1207, pp. 315–318. [Google Scholar]
- Constable, F.; Chambers, G.; Penrose, L.; Daly, A.; Mackie, J.; Davis, K.; Gibbs, M. Viroid-infected tomato and capsicum seed shipments to Australia. Viruses 2019, 11, 98. [Google Scholar] [CrossRef]
- Australian Processing Tomato Grower. Tomatoes. Available online: https://www.accc.gov.au/system/files/public-registers/documents/D11%2B2315841.pdf (accessed on 30 November 2022).
- Plant Health Australia. Processing Tomatoes. Available online: https://www.planthealthaustralia.com.au/industries/processing-tomatoes/ (accessed on 30 November 2022).
- Australian Processing Tomato Research Council. Annual Industry Survey 2020. Available online: https://www.aptrc.asn.au/wp-content/uploads/2020/12/Industry-Survey-2020-ed.pdf (accessed on 30 November 2022).
- Lim, G.T.T.; Wang, G.P.; Hemming, M.N.; Basuki, S.; McGrath, D.J.; Carroll, B.J.; Jones, D.A. Mapping the I-3 gene for resistance to Fusarium wilt in tomato: Application of an I-3 marker in tomato improvement and progress towards the cloning of I-3. Australas. Plant Pathol. 2006, 35, 671–680. [Google Scholar] [CrossRef]
- Grattidge, R.; O’Brien, R.G. Occurrence of a third race of Fusarium wilt of tomatoes in Queensland. Plant Dis. 1982, 66, 165–166. [Google Scholar] [CrossRef]
- Callaghan, S.E. Root and Collar Rot Pathogens Associated with Yield Decline of Processing Tomatoes in Victoria, Australia. Doctoral Dissertation, The University of Melbourne, Melbourne, Australia, 2020. [Google Scholar]
- Callaghan, S.E.; Burgess, L.W.; Ades, P.K.; Tesoriero, L.A.; Taylor, P.W.J. Diversity and pathogenicity of Pythium species associated with reduced yields of processing tomatoes (Solanum lycopersicum ) in Victoria, Australia. Plant Dis. 2022, 106, 1632–1638. [Google Scholar] [CrossRef]
- Watterson, J.C. Diseases. In The Tomato Crop, 1st ed.; Atherton, J.G., Rudich, J., Eds.; Springer: Dordrecht, Germany, 1986; pp. 443–484. [Google Scholar]
- Ekengren, S.K. Cutting the Gordian knot: Taking a stab at corky root rot of tomato. Plant Biotechnol. 2008, 25, 265–269. [Google Scholar] [CrossRef][Green Version]
- Aragona, M.; Minio, A.; Ferrarini, A.; Valente, M.T.; Bagnaresi, P.; Orrù, L.; Delledonne, M. De novo genome assembly of the soil-borne fungus and tomato pathogen Pyrenochaeta Lycopersici. BMC Genom. 2014, 15, 1–12. [Google Scholar] [CrossRef]
- Hasna, M.K.; Ögren, E.; Persson, P.; Mårtensson, A.; Rämert, B. Management of corky root disease of tomato in participation with organic tomato growers. Crop Prot. 2009, 28, 155–161. [Google Scholar] [CrossRef]
- Shankar, R.; Harsha, S.; Bhandary, R. A Practical Guide to Identification and Control of Tomato Diseases. Available online: https://www.researchgate.net/file.PostFileLoader.html?id=589eb904615e2793034a4db2&assetKey=AS%3A460474400677895%401486797060834 (accessed on 30 November 2022).
- Coelho, L.; Chellemi, D.O.; Mitchell, D.J. Efficacy of solarization and cabbage amendment for the control of Phytophthora spp. in North Florida. Plant Dis. 1999, 83, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Yücel, S.; Özarslandan, A.; Colak, A.; Ay, T.; Can, C. Effect of solarization and fumigant applications on soilborne pathogens and root-knot nematodes in greenhouse-grown tomato in Turkey. Phytoparasitica 2007, 35, 450–456. [Google Scholar] [CrossRef]
- Manö, S.; Andreae, M.O. Emission of methyl bromide from biomass burning. Science 1994, 263, 1255–1257. [Google Scholar] [CrossRef] [PubMed]
- Rosskopf, E.N.; Chellemi, D.O.; Kokalis-Burelle, N.; Church, G.T. Alternatives to methyl bromide: A Florida perspective. Plant Health Prog. 2005, 6, 19. [Google Scholar] [CrossRef]
- Locascio, S.J.; Gilreath, J.P.; Dickson, D.W.; Kucharek, T.A.; Jones, J.P.; Noling, J.W. Fumigant alternatives to methyl bromide for polyethylene-mulched. HortScience 1997, 32, 1208–1211. [Google Scholar] [CrossRef]
- Fiume, G.; Fiume, F. Biological control of corky root in tomato. Commun. Agric. Appl. Biol. Sci. 2008, 73, 233–248. [Google Scholar]
- Doganlar, S.; Dodson, J.; Gabor, B.; Beck-Bunn, T.; Crossman, C.; Tanksley, S.D. Molecular mapping of the py-1 gene for resistance to corky root rot (Pyrenochaeta lycopersici) in tomato. Theor. Appl. Genet. 1998, 97, 784–788. [Google Scholar] [CrossRef]
- Milc, J.; Bagnaresi, P.; Aragona, M.; Valente, M.T.; Biselli, C.; Infantino, A.; Pecchioni, N. Comparative transcriptome profiling of the response to Pyrenochaeta lycopersici in resistant tomato cultivar Mogeor and its background genotype—Susceptible Moneymaker. Funct. Integr. Genom. 2019, 19, 811–826. [Google Scholar] [CrossRef]
- Hasna, M.K.; Lagerlöf, J.; Rämert, B. Effects of fungivorous nematodes on corky root disease of tomato grown in compost-amended soil. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2008, 58, 145–153. [Google Scholar] [CrossRef]
- Bubici, G.; Marsico, A.D.; D’Amico, M.; Amenduni, M.; Cirulli, M. Evaluation of Streptomyces spp. for the biological control of corky root of tomato and Verticillium wilt of eggplant. Appl. Soil Ecol. 2013, 72, 128–134. [Google Scholar] [CrossRef]
- Besoain, X.A.; Pérez, L.M.; Araya, A.; Lefever, L.; Montealegre, J.R. New strains obtained after UV treatment and protoplast fusion of native Trichoderma harzianum: Their biocontrol activity on Pyrenochaeta lycopersici. Electron. J. Biotechnol. 2007, 10, 604–617. [Google Scholar] [CrossRef][Green Version]
- Sánchez-Téllez, S.; Herrera-Cid, R.A.; Besoain-Canales, X.A.; Pérez-Roepke, L.M.; Montealegre-Andrade, J.R. In vitro and in vivo inhibitory effect of solid and liquid Trichoderma harzianum formulations on biocontrol of Pyrenochaeta lycopersici. Interciencia 2013, 38, 425–429. [Google Scholar]
- Pérez, L.; Besoaín, X.; Reyes, M.; Pardo, G.; Montealegre, J. The expression of extracellular fungal cell wall hydrolytic enzymes in different Trichoderma harzianum isolates correlates with their ability to control Pyrenochaeta Lycopersici. Biol. Res. 2002, 35, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Hasna, M.K.; Mårtensson, A.; Persson, P.; Rämert, B. Use of composts to manage corky root disease in organic tomato production. Ann. Appl. Biol. 2007, 151, 381–390. [Google Scholar] [CrossRef]
- Edel-Hermann, V.; Lecomte, C. Current status of Fusarium oxysporum formae speciales and races. Phytopathology 2019, 109, 512–530. [Google Scholar] [CrossRef] [PubMed]
- Mutlu, N.; Demirelli, A.; Ilbi, H.; Ikten, C. Development of co-dominant SCAR markers linked to resistant gene against the Fusarium oxysporum f. sp. radicis-lycopersici. Theor. Appl. Genet. 2015, 128, 1791–1798. [Google Scholar] [CrossRef]
- Baysal, Ö.; Çalışkan, M.; Yeşilova, Ö. An inhibitory effect of a new Bacillus subtilis strain (EU07) against Fusarium oxysporum f. sp. radicis-lycopersici. Physiol. Mol. Plant Pathol. 2008, 73, 25–32. [Google Scholar] [CrossRef]
- Gordon, T.R. Fusarium oxysporum and the Fusarium wilt syndrome. Annu. Rev. Phytopathol. 2017, 55, 23–39. [Google Scholar] [CrossRef]
- Gordon, T.R.; Okamoto, D. Population structure and the relationship between pathogenic and nonpathogenic strains of Fusarium oxysporum. Phytopathology 1992, 82, 73–77. [Google Scholar] [CrossRef]
- Olivain, C.; Humbert, C.; Nahalkova, J.; Fatehi, J.; l’Haridon, F.; Alabouvette, C. Colonization of tomato root by pathogenic and nonpathogenic Fusarium oxysporum strains inoculated together and separately into the soil. Appl. Environ. Microbiol. 2006, 72, 1523–1531. [Google Scholar] [CrossRef]
- Muslim, A.; Horinouchi, H.; Hyakumachi, M. Control of Fusarium crown and root rot of tomato with hypovirulent binucleate Rhizoctonia in soil and rock wool systems. Plant Dis. 2003, 87, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Altinok, H.H.; Yüksel, G.; Altinok, M.A. Pathogenicity and phylogenetic analysis of Fusarium oxysporum f. sp. capsici isolates from pepper in Turkey. Can. J. Plant Pathol. 2020, 42, 279–291. [Google Scholar] [CrossRef]
- Zhang, S.; Roberts, P.D.; McGovern, R.J.; Datnoff, L.E. Fusarium Crown and Root Rot of Tomato in Florida. Institute of Food and Agricultural Sciences (IFAS), Publication PP52. Available online: https://edis.ifas.ufl.edu/publication/PG082 (accessed on 30 November 2022).
- McGovern, R.J. Management of tomato diseases caused by Fusarium oxysporum. Crop. Prot. 2015, 73, 78–92. [Google Scholar] [CrossRef]
- Jarvis, W.R. Epidemiology of Fusarium oxysporum f. sp. radicis-lycopersici. In Vascular Wilt Diseases of Plants, 1st ed.; Tjamos, E.C., Beckman, C.H., Eds.; Springer: Berlin, Germany, 1989; pp. 397–411. [Google Scholar]
- Cao, X.; Guan, Z.; Vallad, G.E.; Wu, F. Economics of fumigation in tomato production: The impact of methyl bromide phase-out on the Florida tomato industry. Int. Food Agribus. Manag. Rev. 2019, 22, 589–600. [Google Scholar] [CrossRef]
- Horinouchi, H.; Katsuyama, N.; Taguchi, Y.; Hyakumachi, M. Control of Fusarium crown and root rot of tomato in a soil system by combination of a plant growth-promoting fungus, Fusarium equiseti, and biodegradable pots. Crop. Prot. 2008, 27, 859–864. [Google Scholar] [CrossRef]
- McGovern, R.J.; Vavrina, C.S.; Noling, J.W.; Datnoff, L.A.; Yonce, H.D. Evaluation of application methods of metam sodium for management of Fusarium crown and root rot in tomato in southwest Florida. Plant Dis. 1998, 82, 919–923. [Google Scholar] [CrossRef]
- Minuto, A.; Gullino, M.L.; Lamberti, F.; D’addabbo, T.; Tescari, E.; Ajwa, H.; Garibaldi, A. Application of an emulsifiable mixture of 1, 3-dichloropropene and chloropicrin against root knot nematodes and soilborne fungi for greenhouse tomatoes in Italy. Crop Prot. 2006, 25, 1244–1252. [Google Scholar] [CrossRef]
- Fazio, G.; Stevens, M.R.; Scott, J.W. Identification of RAPD markers linked to fusarium crown and root rot resistance (Frl) in tomato. Euphytica 1999, 105, 205–210. [Google Scholar] [CrossRef]
- Sivan, A.; Ucko, O.; Chet, I. Biological control of Fusarium crown rot of tomato by Trichoderma harzianum under field conditions. Plant Dis. 1987, 71, 587–592. [Google Scholar] [CrossRef]
- Datnoff, L.E.; Nemec, S.; Pernezny, K. Biological control of Fusarium crown and root rot of tomato in Florida using Trichoderma harzianum and Glomus intraradices. Biol. Control 1995, 5, 427–431. [Google Scholar] [CrossRef]
- Malandrakis, A.; Daskalaki, E.R.; Skiada, V.; Papadopoulou, K.K.; Kavroulakis, N. A Fusarium solani endophyte vs. fungicides: Compatibility in a Fusarium oxysporum f. sp. radicis-lycopersici–tomato pathosystem. Fungal Biol. 2018, 122, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Benhamou, N.; Rey, P.; Chérif, M.; Hockenhull, J.; Tirilly, Y. Treatment with the mycoparasite Pythium oligandrum triggers induction of defense-related reactions in tomato roots when challenged with Fusarium oxysporum f. sp. radicis-lycopersici. Phytopathology 1997, 87, 108–122. [Google Scholar] [CrossRef] [PubMed]
- Duffy, B.K.; Défago, G. Zinc improves biocontrol of Fusarium crown and root rot of tomato by Pseudomonas fluorescens and represses the production of pathogen metabolites inhibitory to bacterial antibiotic biosynthesis. Phytopathology 1997, 87, 1250–1257. [Google Scholar] [CrossRef]
- de Weert, S.; Kuiper, I.; Lagendijk, E.L.; Lamers, G.E.; Lugtenberg, B.J. Role of chemotaxis toward fusaric acid in colonization of hyphae of Fusarium oxysporum f. sp. radicis-lycopersici by Pseudomonas fluorescens WCS365. Mol. Plant-Microbe Interact. 2004, 17, 1185–1191. [Google Scholar] [CrossRef] [PubMed]
- Kamilova, F.; Validov, S.; Lugtenberg, B. Biological control of tomato foot and root rot caused by Fusarium oxysporum f. sp. radicis-lycopersici by Pseudomonas bacteria. In Proceedings of the II International Symposium on Tomato Diseases, Kusadasi, Turkey, 8–12 October 2007; Volume 808, pp. 317–320. [Google Scholar]
- Baysal, Ö.; Lai, D.; Xu, H.H.; Siragusa, M.; Çalışkan, M.; Carimi, F.; Tör, M. A proteomic approach provides new insights into the control of soil-borne plant pathogens by Bacillus species. PLoS ONE 2013, 8, e53182. [Google Scholar] [CrossRef]
- Kavroulakis, N.; Ehaliotis, C.; Ntougias, S.; Zervakis, G.I.; Papadopoulou, K.K. Local and systemic resistance against fungal pathogens of tomato plants elicited by a compost derived from agricultural residues. Physiol. Mol. Plant Pathol. 2005, 66, 163–174. [Google Scholar] [CrossRef]
- Ajilogba, C.F.; Babalola, O.O. Integrated management strategies for tomato Fusarium wilt. Biocontrol Sci. 2013, 18, 117–127. [Google Scholar] [CrossRef]
- Miller, S.A.; Rowe, R.C.; Riedel, R.M. Fusarium and Verticillium Wilts of Tomato, Potato, Pepper, and Eggplant. The Ohio State University Extension. Available online: https://www.cabdirect.org/cabdirect/abstract/20127800677 (accessed on 30 November 2022).
- Larkin, R.P.; Fravel, D.R. Effects of varying environmental conditions on biological control of Fusarium wilt of tomato by nonpathogenic Fusarium spp. Phytopathology 2002, 92, 1160–1166. [Google Scholar] [CrossRef]
- Yu, J.; Land, C.J.; Vallad, G.E.; Boyd, N.S. Tomato tolerance and pest control following fumigation with different ratios of dimethyl disulfide and chloropicrin. Pest Manag. Sci. 2019, 75, 1416–1424. [Google Scholar] [CrossRef]
- Bohn, G.W.; Tucker, C.M. Immunity to Fusarium wilt in the tomato. Science 1939, 89, 603–604. [Google Scholar] [CrossRef]
- Takken, F.; Rep, M. The arms race between tomato and Fusarium oxysporum. Mol. Plant Pathol. 2010, 11, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Sela-Buurlage, M.; Budai-Hadrian, O.; Pan, Q.; Carmel-Goren, L.; Vunsch, R.; Zamir, D.; Fluhr, R. Genome-wide dissection of Fusarium resistance in tomato reveals multiple complex loci. Mol. Genet. Genom. 2001, 265, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Katan, T.; Shlevin, E.; Katan, J. Sporulation of Fusarium oxysporum f. sp. lycopersici on stem surfaces of tomato plants and aerial dissemination of inoculum. Phytopathology 1997, 87, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Sarfatti, M.; Katan, J.; Fluhr, R.; Zamir, D. An RFLP marker in tomato linked to the Fusarium oxysporum resistance gene I2. Theor. Appl. Genet. 1989, 78, 755–759. [Google Scholar] [CrossRef] [PubMed]
- McGrath, D.J.; Gillespie, D.; Vawdrey, L. Inheritance of resistance to Fusarium oxysporum f. sp. lycopersici races 2 and 3 in Lycopersicon pennellii. Aust. J. Agric. Res. 1987, 38, 729–733. [Google Scholar] [CrossRef]
- Scott, J.W.; Jones, J.P. Monogenic resistance in tomato to Fusarium oxysporum f. sp. lycopersici race 3. Euphytica 1989, 40, 49–53. [Google Scholar] [CrossRef]
- Catanzariti, A.M.; Lim, G.T.; Jones, D.A. The tomato I-3 gene: A novel gene for resistance to Fusarium wilt disease. New Phytol. 2015, 207, 106–118. [Google Scholar] [CrossRef]
- Chitwood-Brown, J.; Vallad, G.E.; Lee, T.G.; Hutton, S.F. Breeding for resistance to Fusarium wilt of tomato: A review. Genes 2021, 12, 1673. [Google Scholar] [CrossRef]
- Raza, W.; Ling, N.; Zhang, R.; Huang, Q.; Xu, Y.; Shen, Q. Success evaluation of the biological control of Fusarium wilts of cucumber, banana, and tomato since 2000 and future research strategies. Crit. Rev. Biotechnol. 2017, 37, 202–212. [Google Scholar] [CrossRef]
- Alabouvette, C.; Lemanceau, P.; Steinberg, C. Recent advances in the biological control of Fusarium wilts. Pestic. Sci. 1993, 37, 365–373. [Google Scholar] [CrossRef]
- Aimé, S.; Cordier, C.; Alabouvette, C.; Olivain, C. Comparative analysis of PR gene expression in tomato inoculated with virulent Fusarium oxysporum f. sp. lycopersici and the biocontrol strain F. oxysporum Fo47. Physiol. Mol. Plant Pathol. 2008, 73, 9–15. [Google Scholar] [CrossRef]
- de Lamo, F.J.; Spijkers, S.B.; Takken, F.L. Protection to tomato wilt disease conferred by the nonpathogen Fusarium oxysporum Fo47 is more effective than that conferred by avirulent strains. Phytopathology 2021, 111, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Duijff, B.J.; Pouhair, D.; Olivain, C.; Alabouvette, C.; Lemanceau, P. Implication of systemic induced resistance in the suppression of fusarium wilt of tomato by Pseudomonas fluorescens WCS417r and by nonpathogenic Fusarium oxysporum Fo47. Eur. J. Plant Pathol. 1998, 104, 903–910. [Google Scholar] [CrossRef]
- Borrego-Benjumea, A.; Basallote-Ureba, M.J.; Abbasi, P.A.; Lazarovits, G.; Melero-Vara, J.M. Effects of incubation temperature on the organic amendment-mediated control of Fusarium wilt of tomato. Ann. Appl. Biol. 2014, 164, 453–463. [Google Scholar] [CrossRef]
- Zhao, F.; Zhang, Y.; Dong, W.; Zhang, Y.; Zhang, G.; Sun, Z.; Yang, L. Vermicompost can suppress Fusarium oxysporum f. sp. lycopersici via generation of beneficial bacteria in a long-term tomato monoculture soil. Plant Soil 2019, 440, 491–505. [Google Scholar] [CrossRef]
- Barakat, R.M.; Al-Masri, M.I. Trichoderma harzianum in combination with sheep manure amendment enhances soil suppressiveness of Fusarium wilt of tomato. Phytopathol. Mediterr. 2009, 48, 385–395. [Google Scholar]
- Mwangi, M.W.; Muiru, W.M.; Narla, R.D.; Kimenju, J.W.; Kariuki, G.M. Effect of soil sterilisation on biological control of Fusarium oxysporum f. sp. lycopersici and Meloidogyne javanica by antagonistic fungi and organic amendment in tomato crop. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2018, 68, 656–661. [Google Scholar] [CrossRef]
- Lamour, K.H.; Stam, R.; Jupe, J.; Huitema, E. The oomycete broad-host-range pathogen Phytophthora capsici. Mol. Plant Pathol. 2012, 13, 329–337. [Google Scholar] [CrossRef]
- Wani, A.H. An overview of the fungal rot of tomato. Mycopath 2011, 9, 33–38. [Google Scholar]
- Hausbeck, M.K.; Lamour, K.H. Phytophthora capsici on vegetable crops: Research progress and management challenges. Plant Dis. 2004, 88, 1292–1303. [Google Scholar] [CrossRef]
- Ristaino, J.B.; Johnston, S.A. Ecologically based approaches to management of Phytophthora blight on bell pepper. Plant Dis. 1999, 83, 1080–1089. [Google Scholar] [CrossRef] [PubMed]
- Lamour, K.H.; Hausbeck, M.K. Effect of crop rotation on the survival of Phytophthora capsici in Michigan. Plant Dis. 2003, 87, 841–845. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Ocampo, L.M.; Hausbeck, M.K. Resistance in tomato and wild relatives to crown and root rot caused by Phytophthora capsici. Phytopathology 2010, 100, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Bower, L.A.; Coffey, M.D. Development of laboratory tolerance to phosphorous acid, fosetyl-Al, and metalaxyl in Phytophthora capsici. Can. J. Plant Pathol. 1985, 7, 1–6. [Google Scholar] [CrossRef]
- Jackson, K.L.; Yin, J.; Ji, P. Sensitivity of Phytophthora capsici on vegetable crops in Georgia to mandipropamid, dimethomorph, and cyazofamid. Plant Dis. 2012, 96, 1337–1342. [Google Scholar] [CrossRef] [PubMed]
- Kousik, C.S.; Keinath, A.P. First report of insensitivity to cyazofamid among isolates of Phytophthora capsici from the southeastern United States. Plant Dis. 2008, 92, 979. [Google Scholar] [CrossRef] [PubMed]
- Siegenthaler, T.B.; Hansen, Z.R. Sensitivity of Phytophthora capsici from Tennessee to mefenoxam, fluopicolide, oxathiapiprolin, dimethomorph, mandipropamid, and cyazofamid. Plant Dis. 2021, 105, 3000–3007. [Google Scholar] [CrossRef]
- Quesada-Ocampo, L.M.; Vargas, A.M.; Naegele, R.P.; Francis, D.M.; Hausbeck, M.K. Resistance to crown and root rot caused by Phytophthora capsici in a tomato advanced backcross of Solanum habrochaites and Solanum lycopersicum. Plant Dis. 2016, 100, 829–835. [Google Scholar] [CrossRef]
- Moataza, M.S. Destruction of Rhizoctonia solani and Phytophthora capsici causing tomato root-rot by Pseudomonas fluorescences lytic enzymes. Res. J. Agric. Biol. Sci. 2006, 2, 274–281. [Google Scholar]
- Sharma, R.; Chauhan, A.; Shirkot, C.K. Characterization of plant growth promoting Bacillus strains and their potential as crop protectants against Phytophthora capsici in tomato. Biol. Agric. Hortic. 2015, 31, 230–244. [Google Scholar] [CrossRef]
- Syed-Ab-Rahman, S.F.; Xiao, Y.; Carvalhais, L.C.; Ferguson, B.J.; Schenk, P.M. Suppression of Phytophthora apsica infection and promotion of tomato growth by soil bacteria. Rhizosphere 2019, 9, 72–75. [Google Scholar] [CrossRef]
- Yang, K.; Dong, X.; Li, J.; Wang, Y.; Cheng, Y.; Zhai, Y.; Dou, D. Type 2 Nep1-like proteins from the biocontrol oomycete Pythium oligandrum suppress Phytophthora capsici infection in solanaceous plants. J. Fungi 2021, 7, 496. [Google Scholar] [CrossRef] [PubMed]
- Nicol, R.W.; Burlakoti, P. Effect of aerobic compost tea inputs and application methods on protecting tomato from Phytophthora capsici. In Proceedings of the IV International Symposium on Tomato Diseases, Orlando, FL, USA, 24–27 June 2013; Volume 1069, pp. 229–233. [Google Scholar]
- González-Hernández, A.I.; Suárez-Fernández, M.B.; Pérez-Sánchez, R.; Gómez-Sánchez M, Á.; Morales-Corts, M.R. Compost tea induces growth and resistance against Rhizoctonia solani and Phytophthora capsici in pepper. Agronomy 2021, 11, 781. [Google Scholar] [CrossRef]
- Jiang, Z.Q.; Guo, Y.H.; Li, S.M.; Qi, H.Y.; Guo, J.H. Evaluation of biocontrol efficiency of different Bacillus preparations and field application methods against Phytophthora blight of bell pepper. Biol. Control 2006, 36, 216–223. [Google Scholar] [CrossRef]
- Kim, K.D.; Nemec, S.; Musson, G. Control of Phytophthora root and crown rot of bell pepper with composts and soil amendments in the greenhouse. Appl. Soil Ecol. 1997, 5, 169–179. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef]
- Elshahawy, I.E.; El-Mohamedy, R.S. Biological control of Pythium damping-off and root-rot diseases of tomato using Trichoderma isolates employed alone or in combination. J. Plant Pathol. 2019, 101, 597–608. [Google Scholar] [CrossRef]
- Manjunath, M.; Prasanna, R.; Nain, L.; Dureja, P.; Singh, R.; Kumar, A.; Kaushik, B.D. Biocontrol potential of cyanobacterial metabolites against damping off disease caused by Pythium aphanidermatum in solanaceous vegetables. Arch. Phytopathol. Plant Prot. 2010, 43, 666–677. [Google Scholar] [CrossRef]
- Mdee, L.K.; Masoko, P.; Eloff, J.N. The activity of extracts of seven common invasive plant species on fungal phytopathogens. S. Afr. J. Bot. 2009, 75, 375–379. [Google Scholar] [CrossRef]
- Beckerman, J. Pythium Root Rot of Herbaceous Plants. Available online: https://bch.cbd.int/api/v2013/documents/7efc7c33-9c4d-4589-95b0-959650a561fa/attachments/20505/Pythium%20Root%20Rot%20of%20herbaceous%20plants.pdf (accessed on 30 November 2022).
- Arora, H.; Sharma, A.; Sharma, S.; Haron, F.F.; Gafur, A.; Sayyed, R.Z.; Datta, R. Pythium damping-off and root rot of capsicum annuum l.: Impacts, diagnosis, and management. Microorganisms 2021, 9, 823. [Google Scholar] [CrossRef]
- Male, M.F.; Vawdrey, L.L. Efficacy of fungicides against damping-off in papaya seedlings caused by Pythium Aphanidermatum. Australas. Plant Dis. Notes 2010, 5, 103–104. [Google Scholar] [CrossRef]
- Paulitz, T.C.; Zhou, T.; Rankin, L. Selection of rhizosphere bacteria for biological control of Pythium aphanidermatum on hydroponically grown cucumber. Biol. Control 1992, 2, 226–237. [Google Scholar] [CrossRef]
- Hassanisaadi, M.; Shahidi Bonjar, G.H.; Hosseinipour, A.; Abdolshahi, R.; Ait Barka, E.; Saadoun, I. Biological control of Pythium aphanidermatum, the causal agent of tomato root rot by two Streptomyces root symbionts. Agronomy 2021, 11, 846. [Google Scholar] [CrossRef]
- Harvey, P.; Lawrence, L. Managing Pythium root disease complexes to improve productivity of crop rotations. Outlooks Pest Manag. 2008, 19, 127. [Google Scholar] [CrossRef]
- Triki, M.A.; Priou, S.; El Mahjoub, M. Effects of soil solarization on soil-borne populations of Pythium aphanidermatum and Fusarium solani and on the potato crop in Tunisia. Potato Res. 2001, 44, 271–279. [Google Scholar] [CrossRef]
- Reddy, G.S.; Rao, V.K.; Sitaramaiah, K.; Chalam, T.V. Soil Solarization for Control of Soil-borne Pathogen Complex due to Meloidogyne incognita and Pythium aphanidermatum. Indian J. Nematol. 2001, 31, 136–138. [Google Scholar]
- Jayaraj, J.; Radhakrishnan, N.V. Enhanced activity of introduced biocontrol agents in solarized soils and its implications on the integrated control of tomato damping-off caused by Pythium spp. Plant Soil 2008, 304, 189–197. [Google Scholar] [CrossRef]
- Abbasi, P.A.; Lazarovits, G. Seed treatment with phosphonate (AG3) suppresses Pythium damping-off of cucumber seedlings. Plant Dis. 2006, 90, 459–464. [Google Scholar] [CrossRef]
- Al-Balushi, Z.M.; Agrama, H.; Al-Mahmooli, I.H.; Maharachchikumbura, S.S.; Al-Sadi, A.M. Development of resistance to hymexazol among Pythium species in cucumber greenhouses in Oman. Plant Dis. 2018, 102, 202–208. [Google Scholar] [CrossRef]
- Cuevas, V.C.; Sinohin, A.M. Performance of selected Philippine species of Trichoderma as biocontrol agents of damping off pathogens and as growth enhancer of vegetables in farmers’ field. Philipp. Agric. Sci. 2005, 88, 63–71. [Google Scholar]
- Garzón, C.D.; Molineros, J.E.; Yánez, J.M.; Flores, F.J.; del Mar Jiménez-Gasco, M.; Moorman, G.W. Sublethal doses of mefenoxam enhance Pythium damping-off of geranium. Plant Dis. 2011, 95, 1233–1238. [Google Scholar] [CrossRef] [PubMed]
- Samaras, A.; Roumeliotis, E.; Ntasiou, P.; Karaoglanidis, G. Bacillus subtilis MBI600 promotes growth of tomato plants and induces systemic resistance contributing to the control of soilborne pathogens. Plants 2021, 10, 1113. [Google Scholar] [CrossRef] [PubMed]
- Rajendraprasad, M.; Vidyasagar, B.; Devi, G.U.; Rao, S.K. Biological control of tomato damping off caused by Pythium debaryanum. Int. J. Chem. Stud. 2017, 5, 447–452. [Google Scholar]
- Salman, M.; Abuamsha, R. Potential for integrated biological and chemical control of damping-off disease caused by Pythium ultimum in tomato. BioControl 2012, 57, 711–718. [Google Scholar] [CrossRef]
- Dukare, A.S.; Prasanna, R.; Dubey, S.C.; Nain, L.; Chaudhary, V.; Singh, R.; Saxena, A.K. Evaluating novel microbe amended composts as biocontrol agents in tomato. Crop. Prot. 2011, 30, 436–442. [Google Scholar] [CrossRef]
- Porter, L.D.; Hamm, P.B.; David, N.L.; Gieck, S.L.; Miller, J.S.; Gundersen, B.; Inglis, D.A. Metalaxyl-M-resistant Pythium species in potato production areas of the Pacific Northwest of the USA. Am. J. Potato Res. 2009, 86, 315–326. [Google Scholar] [CrossRef]
- Del Castillo Múnera, J.; Hausbeck, M.K. Characterization of Pythium species associated with greenhouse floriculture crops in Michigan. Plant Dis. 2016, 100, 569–576. [Google Scholar] [CrossRef]
- Aegerter, B.J.; Greathead, A.S.; Pierce, L.E.; Davis, R.M. Mefenoxam-resistant isolates of Pythium irregulare in an ornamental greenhouse in California. Plant Dis. 2002, 86, 692. [Google Scholar] [CrossRef]
- Lee, S.; Garzón, C.D.; Moorman, G.W. Genetic structure and distribution of Pythium aphanidermatum populations in Pennsylvania greenhouses based on analysis of AFLP and SSR markers. Mycologia 2010, 102, 774–784. [Google Scholar] [CrossRef]
- Lookabaugh, E.C.; Kerns, J.P.; Shew, B.B. Evaluating Fungicide Selections to Manage Pythium Root Rot on Poinsettia Cultivars with Varying Levels of Partial Resistance. Plant Dis. 2021, 105, 1640–1647. [Google Scholar] [CrossRef]
- Niderman, T.; Genetet, I.; Bruyere, T.; Gees, R.; Stintzi, A.; Legrand, M.; Mosinger, E. Pathogenesis-related PR-1 proteins are antifungal (isolation and characterization of three 14-kilodalton proteins of tomato and of a basic PR-1 of tobacco with inhibitory activity against Phytophthora infestans). Plant Physiol. 1995, 108, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Tornero, P.; Gadea, J.; Conejero, V.; Vera, P. Two PR-1 genes from tomato are differentially regulated and reveal a novel mode of expression for a pathogenesis-related gene during the hypersensitive response and development. Mol. Plant-Microbe Interact. 1997, 10, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Postma, J.; Clematis, F.; Nijhuis, E.H.; Someus, E. Efficacy of four phosphate-mobilizing bacteria applied with an animal bone charcoal formulation in controlling Pythium aphanidermatum and Fusarium oxysporum f. sp. radicis lycopersici in tomato. Biol. Control 2013, 67, 284–291. [Google Scholar] [CrossRef]
- Martinez, C.; Bourassa, A.; Roy, G.; Desbiens, M.C.; Bussières, P. Efficacy of PRO-MIX® with Biofungicide against Root Diseases caused by Pythium spp. and Rhizoctonia spp. In Proceedings of the XXVII International Horticultural Congress-IHC2006: International Symposium on Sustainability through Integrated and Organic, Seoul, Republic of Korea, 13–19 August 2006; Volume 767, pp. 185–192. [Google Scholar]
- Caron, J.; Laverdière, L.; Thibodeau, P.O.; Bélanger, R.R. Utilisation d’une souche indigène de Trichoderma harzianum contre cinq agents pathogènes chez le concombre et la tomate de serre au Québec. Phytoprotection 2002, 83, 73–87. [Google Scholar] [CrossRef][Green Version]
- Larsen, J.; Graham, J.H.; Cubero, J.; Ravnskov, S. Biocontrol traits of plant growth suppressive arbuscular mycorrhizal fungi against root rot in tomato caused by Pythium aphanidermatum. Eur. J. Plant Pathol. 2012, 133, 361–369. [Google Scholar] [CrossRef]
- St Martin, C.C.G.; Dorinvil, W.; Brathwaite, R.A.I.; Ramsubhag, A. Effects and relationships of compost type, aeration and brewing time on compost tea properties, efficacy against Pythium ultimum, phytotoxicity and potential as a nutrient amendment for seedling production. Biol. Agric. Hortic. 2012, 28, 185–205. [Google Scholar] [CrossRef]
- Jenana RK, B.; Haouala, R.; Triki, M.A.; Godon, J.J.; Hibar, K.; Khedher, M.B.; Henchi, B. Composts, compost extracts and bacterial suppressive action on Pythium aphanidermatum in tomato. Pak. J. Bot. 2009, 41, 315–327. [Google Scholar]
- Postma, J.; Nijhuis, E.H. Pseudomonas chlororaphis and organic amendments controlling Pythium infection in tomato. Eur. J. Plant Pathol. 2019, 154, 91–107. [Google Scholar] [CrossRef]
- Jayaraj, J.; Radhakrishnan, N.V.; Kannan, R.; Sakthivel, K.; Suganya, D.; Venkatesan, S.; Velazhahan, R. Development of new formulations of Bacillus subtilis for management of tomato damping-off caused by Pythium aphanidermatum. Biocontrol Sci. Technol. 2005, 15, 55–65. [Google Scholar] [CrossRef]
- Fradin, E.F.; Zhang, Z.; Juarez Ayala, J.C.; Castroverde, C.D.; Nazar, R.N.; Robb, J.; Thomma, B.P. Genetic dissection of Verticillium wilt resistance mediated by tomato Ve1. Plant Physiol. 2009, 150, 320–332. [Google Scholar] [CrossRef]
- Fradin, E.F.; Thomma, B.P. Physiology and molecular aspects of Verticillium wilt diseases caused by V. dahliae and V. albo-atrum. Mol. Plant Pathol. 2006, 7, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Mandelc, S.; Timperman, I.; Radišek, S.; Devreese, B.; Samyn, B.; Javornik, B. Comparative proteomic profiling in compatible and incompatible interactions between hop roots and Verticillium albo-atrum. Plant Physiol. Biochem. 2013, 68, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Berlanger, I.; Powelson, M.L. Verticillium Wilt. Available online: https://www.apsnet.org/edcenter/disandpath/fungalasco/pdlessons/Pages/VerticilliumWilt.aspx (accessed on 30 November 2022).
- Butterfield, E.J.; DeVay, J.E.; Garber, R.H. The influence of several crop sequences on the incidence of Verticillium wilt of cotton and on the populations of Verticillium dahliae in field soil. Phytopathology 1978, 68, 1217–1220. [Google Scholar] [CrossRef]
- Iott, M.C. Utility of Grafting and Evaluation of Rootstocks for the Management of Verticillium Wilt in Tomato Production in Western North Carolina. Master’s Thesis, North Carolina State University, Raleigh, NC, USA, 2013. [Google Scholar]
- Stapleton, J.J. Soil solarization in various agricultural production systems. Crop. Prot. 2000, 19, 837–841. [Google Scholar] [CrossRef]
- Goicoechea, N. To what extent are soil amendments useful to control Verticillium wilt. Pest Manag. Sci. Former. Pestic. Sci. 2009, 65, 831–839. [Google Scholar] [CrossRef]
- Rowe, R.C.; Powelson, M.L. Potato early dying: Management challenges in a changing production environment. Plant Dis. 2002, 86, 1184–1193. [Google Scholar] [CrossRef]
- Ordentlich, A.; Nachmias, A.; Chet, I. Integrated control of Verticillium dahliae in potato by Trichoderma harzianum and captan. Crop. Prot. 1990, 9, 363–366. [Google Scholar] [CrossRef]
- Baergen, K.D.; Hewitt, J.D.; Clair, D.S. Resistance of tomato genotypes to four isolates of Verticillium dahliae race 2. HortScience 1993, 28, 833–836. [Google Scholar] [CrossRef]
- Dhouib, H.; Zouari, I.; Abdallah, D.B.; Belbahri, L.; Taktak, W.; Triki, M.A.; Tounsi, S. Potential of a novel endophytic Bacillus velezensis in tomato growth promotion and protection against Verticillium wilt disease. Biol. Control 2019, 139, 104092. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Sakr, S.; Bufo, S.A.; Camele, I. An attempt of biocontrol the tomato-wilt disease caused by Verticillium dahliae using Burkholderia gladioli pv. agaricicola and its bioactive secondary metabolites. Int. J. Plant Biol. 2017, 8, 7263. [Google Scholar] [CrossRef]
- Jabnoun-Khiareddine, H.; Daami-Remadi, M.; Ayed, F.; El Mahjoub, M. Biocontrol of tomato Verticillium wilt by using indigenous Gliocladium spp. and Penicillium sp. isolates. Dyn. Soil Dyn. Plant 2009, 3, 70–79. [Google Scholar]
- Larena, I.; Sabuquillo, P.; Melgarejo, P.; De Cal, A. Biocontrol of Fusarium and Verticillium wilt of tomato by Penicillium oxalicum under greenhouse and field conditions. J. Phytopathol. 2003, 151, 507–512. [Google Scholar] [CrossRef]
- Jabnoun-Khiareddine, H.; Daami-Remadi, M.; Ayed, F.; El Mahjoub, M. Biological control of tomato Verticillium wilt by using indigenous Trichoderma spp. Afr. J. Plant Sci. Biotechnol. 2009, 3, 26–36. [Google Scholar]
- Naraghi, L.; Heydari, A.; Rezaee, S.; Razavi, M.; Jahanifar, H.; Khaledi, E. Biological control of tomato Verticillium wilt disease by Talaromyces flavus. J. Plant Prot. Res. 2010, 50, 360–365. [Google Scholar] [CrossRef]
- Puri, K.D.; Hu, X.; Gurung, S.; Short, D.P.; Sandoya, G.V.; Schild, M.; Subbarao, K.V. Verticillium klebahnii and V. isaacii Isolates Exhibit Host-dependent Biological Control of Verticillium Wilt Caused by V. dahliae. PhytoFrontiers 2021, 1, 276–290. [Google Scholar] [CrossRef]
- Acharya, B.; Ingram, T.W.; Oh, Y.; Adhikari, T.B.; Dean, R.A.; Louws, F.J. Opportunities and challenges in studies of host-pathogen interactions and management of Verticillium dahliae in tomatoes. Plants 2020, 9, 1622. [Google Scholar] [CrossRef]
- Kadoglidou, K.; Chatzopoulou, P.; Maloupa, E.; Kalaitzidis, A.; Ghoghoberidze, S.; Katsantonis, D. Mentha and oregano soil amendment induces enhancement of tomato tolerance against soilborne diseases, yield and quality. Agronomy 2020, 10, 406. [Google Scholar] [CrossRef]
- Ait Rahou, Y.; Ait-El-Mokhtar, M.; Anli, M.; Boutasknit, A.; Ben-Laouane, R.; Douira, A.; Meddich, A. Use of mycorrhizal fungi and compost for improving the growth and yield of tomato and its resistance to Verticillium dahliae. Arch. Phytopathol. Plant Prot. 2021, 54, 665–690. [Google Scholar] [CrossRef]
- Lazarovits, G.; Conn, K.; Tenuta, M. Control of Verticillium dahliae with soil amendments: Efficacy and mode of action. In Advances in Verticillium Research and Disease Management, 1st ed.; Tjamos, E.C., Rowe, R.C., Heale, J.B., Fravel, D.R., Eds.; APS Press: St Paul, MN, USA, 2000; pp. 274–291. [Google Scholar]
- Yuce, E.K.; Yigit, S.; Tosun, N. Efficacy of solarization combined with metam sodium and hydrogen peroxide in control of Fusarium oxysporum f. sp. radicis-lycopersici and Clavibacter michiganensis subsp. michiganensis in Tomato greenhouse. In Proceedings of the III International Symposium on Tomato Diseases, Ischia, Italy, 25–30 July 2010; Volume 914, pp. 385–391. [Google Scholar]
- Kulus, D. Genetic resources and selected conservation methods of tomato. J. Appl. Bot. Food Qual. 2018, 91, 135–144. [Google Scholar]
- Fones, H.N.; Bebber, D.P.; Chaloner, T.M.; Kay, W.T.; Steinberg, G.; Gurr, S.J. Threats to global food security from emerging fungal and oomycete crop pathogens. Nat. Food 2020, 1, 332–342. [Google Scholar] [CrossRef]
- Bebber, D.P.; Field, E.; Gui, H.; Mortimer, P.; Holmes, T.; Gurr, S.J. Many unreported crop pests and pathogens are probably already present. Glob. Chang. Biol. 2019, 25, 2703–2713. [Google Scholar] [CrossRef] [PubMed]
- Cammarano, D.; Jamshidi, S.; Hoogenboom, G.; Ruane, A.C.; Niyogi, D.; Ronga, D. Processing tomato production is expected to decrease by 2050 due to the projected increase in temperature. Nat. Food 2022, 3, 347–444. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).