

Effect of Ciprofloxacin on the Composition of Intestinal Microbiota in Sarcophaga peregrina (Diptera: Sarcophagidae)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Cultivation of S. peregrina
2.3. 16S rRNA Microbial Community Analysis
2.4. Quality Control of Sequencing Data
2.5. Taxonomic Classification of OTUs
2.6. Statistical Analysis
3. Results
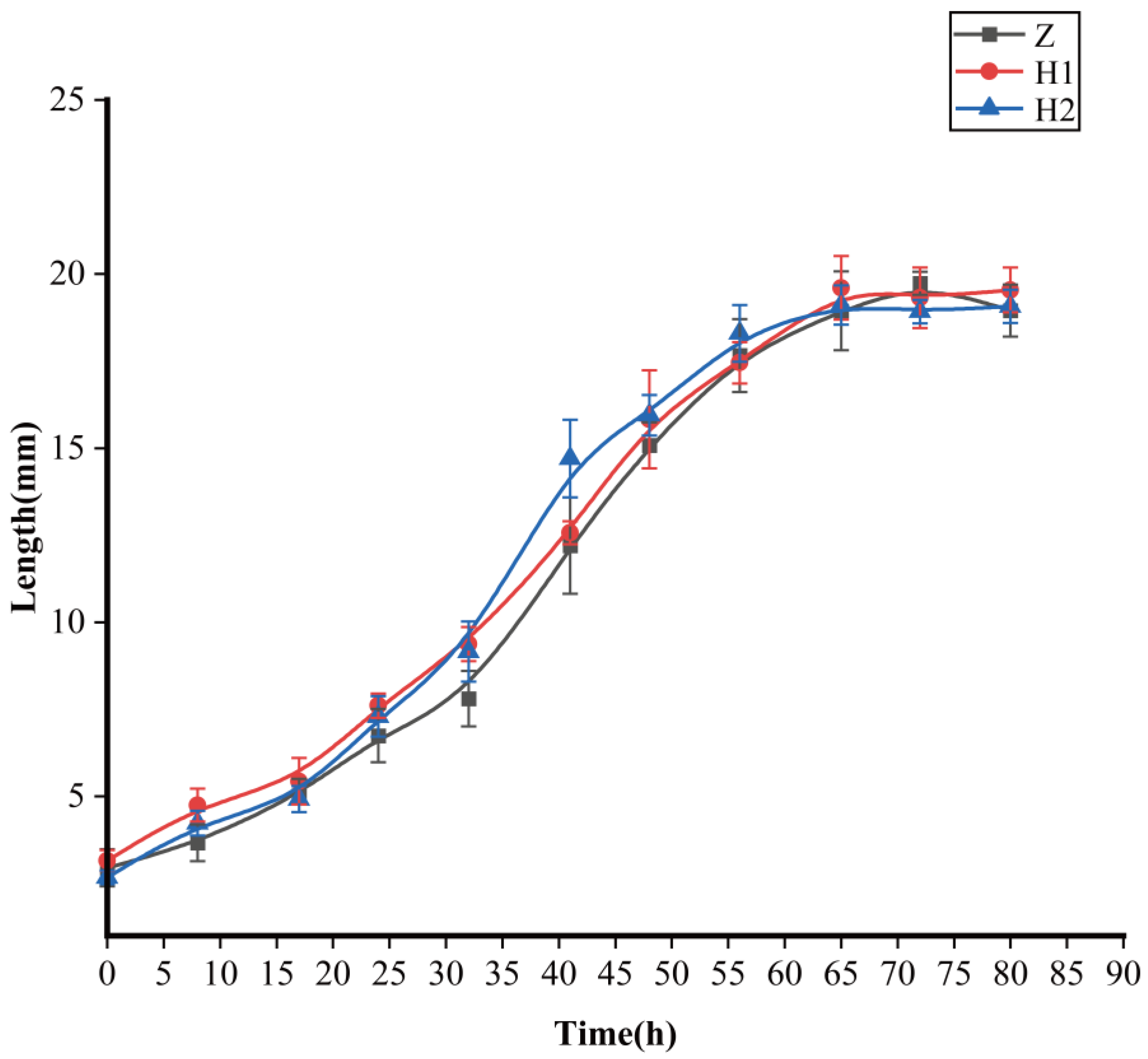
3.1. Effects of Ciprofloxacin on Body Length and Body Weight of S. peregrina
3.2. Gut Microbiota Sequencing Results and Data Analysis
3.2.1. Sequencing Results
3.2.2. Gut microbiota Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cirimotich:, C.M.; Ramirez, J.L.; Dimopoulos, G. Native Microbiota Shape Insect Vector Competence for Human Pathogens. Cell Host Microbe 2011, 10, 307–310. [Google Scholar] [CrossRef]
- Ceja-Navarro, J.A.; Vega, F.E.; Karaoz, U.; Hao, Z.; Jenkins, S.; Lim, H.C.; Kosina, P.; Infante, F.; Northen, T.R.; Brodie, E.L. Gut microbiota mediate caffeine detoxification in the primary insect pest of coffee. Nat. Commun. 2015, 6, 7618. [Google Scholar] [CrossRef]
- Lee, J.B.; Park, K.E.; Lee, S.A.; Jang, S.H.; Eo, H.J.; Jang, H.A.; Kim, C.H.; Ohbayashi, T.; Matsuura, Y.; Kikuchi, Y.; et al. Gut symbiotic bacteria stimulate insect growth and egg production by modulating and gene expression. Dev. Comp. Immunol. 2017, 69, 12–22. [Google Scholar] [CrossRef]
- Goane, L.; Salgueiro, J.; Pereyra, P.M.; Arce, O.E.A.; Ruiz, M.J.; Nussenbaum, A.L.; Segura, D.F.; Vera, M.T. Antibiotic treatment reduces fecundity and nutrient content in females of (Diptera: Tephritidae) in a diet dependent way. J. Insect. Physiol. 2022, 139, 104396. [Google Scholar] [CrossRef]
- Shin, S.C.; Kim, S.H.; You, H.; Kim, B.; Kim, A.C.; Lee, K.A.; Yoon, J.H.; Ryu, J.H.; Lee, W.J. Microbiome Modulates Host Developmental and Metabolic Homeostasis via Insulin Signaling. Science 2011, 334, 670–674. [Google Scholar] [CrossRef]
- Storelli, G.; Defaye, A.; Erkosar, B.; Hols, P.; Royet, J.; Leulier, F. Promotes Systemic Growth by Modulating Hormonal Signals through TOR-Dependent Nutrient Sensing. Cell Metab. 2011, 14, 403–414. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Reddy, D.N. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef]
- Ramsey, J.S.; Chavez, J.D.; Johnson, R.; Hosseinzadeh, S.; Mahoney, J.E.; Mohr, J.P.; Robison, F.; Zhong, X.; Hall, D.G.; MacCoss, M.; et al. Protein interaction networks at the host-microbe interface in, the insect vector of the citrus greening pathogen. R. Soc. Open Sci. 2017, 4, 160545. [Google Scholar] [CrossRef]
- Douglas, A.E. Omics and the metabolic function of insect-microbial symbioses. Curr. Opin. Insect. Sci. 2018, 29, 1–6. [Google Scholar] [CrossRef]
- Singh, B.; Crippen, T.; Zheng, L.Y.; Fields, A.T.; Yu, Z.N.; Ma, Q.; Wood, T.K.; Dowd, S.E.; Flores, M.; Tomberlin, J.K.; et al. A metagenomic assessment of the bacteria associated with Lucilia sericata and Lucilia cuprina (Diptera: Calliphoridae). Appl. Microbiol. Biotechnol. 2015, 99, 869–883. [Google Scholar] [CrossRef]
- Wohlfahrt, D.; Woolf, M.S.; Singh, B. A survey of bacteria associated with various life stages of primary colonizers: Lucilia sericata and Phormia regina. Sci. Justice 2020, 60, 173–179. [Google Scholar] [CrossRef]
- Palevich, N.; Maclean, P.H.; Carvalho, L.; Jauregui, R. Bacterial Diversity Profiling of the New Zealand Parasitic Blowfly Based on 16S rRNA Gene Amplicon Sequencing. Microbiol. Resour. Announc. 2021, 10, e00257-21. [Google Scholar] [CrossRef]
- Gasz, N.E.; Geary, M.J.; Doggett, S.L.; Harvey, M.L. Bacterial association observations in Lucilia sericata and Lucilia cuprina organs through 16S rRNA gene sequencing. Appl. Microbiol. Biotechnol. 2021, 105, 2979–2980. [Google Scholar] [CrossRef]
- Junqueira, A.C.M.; Ratan, A.; Acerbi, E.; Drautz-Moses, D.I.; Premkrishnan, B.N.V.; Costea, P.I.; Linz, B.; Purbojati, R.W.; Paulo, D.F.; Gaultier, N.E.; et al. The microbiomes of blowflies and houseflies as bacterial transmission reservoirs. Sci. Rep. 2017, 7, 16324. [Google Scholar] [CrossRef]
- Bahrndorff, S.; de Jonge, N.; Skovgård, H.; Nielsen, J.L. Bacterial Communities Associated with Houseflies (L.) Sampled within and between Farms. PLoS ONE 2017, 12, e0169753. [Google Scholar] [CrossRef]
- Uriel, Y.; Gries, R.; Tu, L.; Carroll, C.; Zhai, H.M.; Moore, M.; Gries, G. The fly factor phenomenon is mediated by interkingdom signaling between bacterial symbionts and their blow fly hosts. Insect. Sci. 2020, 27, 256–265. [Google Scholar] [CrossRef]
- Marei, N.; Elwahy, A.H.M.; Salah, T.A.; El Sherif, Y.; Abd El-Samie, E. Enhanced antibacterial activity of Egyptian local insects’ chitosan-based nanoparticles loaded with ciprofloxacin-HCl. Int. J. Biol. Macromol. 2019, 126, 262–272. [Google Scholar] [CrossRef]
- Cacicedo, M.L.; Pacheco, G.; Islan, G.A.; Alvarez, V.A.; Barud, H.S.; Castro, G.R. Chitosan-bacterial cellulose patch of ciprofloxacin for wound dressing: Preparation and characterization studies. Int. J. Biol. Macromol. 2020, 147, 1136–1145. [Google Scholar] [CrossRef]
- Appelbaum, P.C.; Hunter, P.A. The fluoroquinolone antibacterials: Past, present and future perspectives. Int. J. Antimicrob. Agents 2000, 16, 5–15. [Google Scholar] [CrossRef]
- Hooper, D.C. Mechanisms of action of antimicrobials: Focus on fluoroquinolones. Clin. Infect. Dis. 2001, 32, S9–S15. [Google Scholar] [CrossRef]
- Roy, D.C.; Tomblyn, S.; Burmeister, D.M.; Wrice, N.L.; Becerra, S.C.; Burnett, L.R.; Saul, J.M.; Christy, R.J. Ciprofloxacin-Loaded Keratin Hydrogels Prevent Infection and Support Healing in a Porcine Full-Thickness Excisional Wound. Adv. Wound Care 2015, 4, 457–468. [Google Scholar] [CrossRef]
- Contardi, M.; Heredia-Guerrero, J.A.; Perotto, G.; Valentini, P.; Pompa, P.P.; Spanò, R.; Goldoni, L.; Bertorelli, R.; Athanassiou, A.; Bayer, I.S. Transparent ciprofloxacin-povidone antibiotic films and nanofiber mats as potential skin and wound care dressings. Eur. J. Pharm. Sci. 2017, 104, 133–144. [Google Scholar] [CrossRef]
- Najafabadi, S.A.A.; Shirazaki, P.; Kharazi, A.Z.; Varshosaz, J.; Tahriri, M.; Tayebi, L. Evaluation of sustained ciprofloxacin release of biodegradable electrospun gelatin/poly(glycerol sebacate) mat membranes for wound dressing applications. Asia-Pac. J. Chem. Eng. 2018, 13, e2255. [Google Scholar] [CrossRef]
- Chatterjee, S.N.; Das, D.; Dangar, T.K. Efficacy of Bacillus sphaericus against Aedes (Stegomyia) aegypti, Mansonia indiana (Edward) and Culex vishnui under laboratory and field conditions. Entomon 2008, 33, 181–187. [Google Scholar]
- Ferraz, A.C.P.; Dallavecchia, D.L.; da Silva, D.C.; de Carvalho, R.P.; da Silva, R.G.; Aguiar-Coelho, V.M. Evaluation of the influence of the antibiotic ciprofloxacin in the development of an Old World screwworm fly, Chrysomya putoria. J. Insect. Sci. 2014, 14, 3. [Google Scholar] [CrossRef]
- Chatterjee, S.; Ghosh, T.S. Characterization of Ampicillin Resistant sp Isolated from the Midgut of (Van der Wulp) and its Role on Larval Development. J. Pure Appl. Microbiol. 2010, 4, 875–878. [Google Scholar]
- Preusser, D.; Bröring, U.; Fischer, T.; Juretzek, T. Effects of antibiotics ceftriaxone and levofloxacin on the growth of L. (Diptera: Calliphoridae) and effects on the determination of the post-mortem interval. J. Forensic Leg. Med. 2021, 81, 102207. [Google Scholar] [CrossRef]
- Ferraz, A.C.P.; Dallavecchia, D.L.; Silva, D.C.; Figueiredo, A.L.; Proença, B.; Silva, R.G.; Aguiar, V.M. Effects of the Antibiotics Gentamicin on the Postembryonic Development of (Diptera: Calliphoridae). J. Insect. Sci. 2014, 14, 279. [Google Scholar] [CrossRef]
- Natori, S. Molecules participating in insect immunity of Sarcophaga peregrina. Proc. Jpn. Acad. Ser. B 2010, 86, 927–938. [Google Scholar] [CrossRef]
- Shang, Y.J.; Amendt, J.; Wang, Y.; Ren, L.P.; Yang, F.Q.; Zhang, X.Y.; Zhang, C.Q.; Guo, Y.D. Multimethod combination for age estimation of (Diptera: Sarcophagidae) with implications for estimation of the postmortem interval. Int. J. Leg. Med. 2023, 137, 329–344. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microb. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Preusser, D.; Fischer, T.; Juretzek, T. Effects of antibiotics ceftriaxone and levofloxacin on the growth of Lucilia sericata (Diptera: Calliphoridae). Med. Vet. Entomol. 2023, 37, 805–815. [Google Scholar] [CrossRef]
- Sherman, R.A.; Wyle, F.A.; Thrupp, L. Effects of seven antibiotics on the growth and development of Phaenicia sericata (Diptera: Calliphoridae) larvae. J. Med. Entomol. 1995, 32, 646–649. [Google Scholar] [CrossRef]
- Gupta, A.K.; Rastogi, G.; Nayduch, D.; Sawant, S.S.; Bhonde, R.R.; Shouche, Y.S. Molecular phylogenetic profiling of gut-associated bacteria in larvae and adults of flesh flies. Med. Vet. Entomol. 2014, 28, 345–354. [Google Scholar] [CrossRef]
- Pais, I.S.; Valente, R.S.; Sporniak, M.; Teixeira, L. Drosophila melanogaster establishes a species-specific mutualistic interaction with stable gut-colonizing bacteria. PLoS Biol. 2018, 16, e2005710. [Google Scholar] [CrossRef]
- Ventura, C.; Briones-Roblero, C.I.; Hernández, E.; Rivera-Orduña, F.N.; Zúñiga, G. Comparative Analysis of the Gut Bacterial Community of Four Fruit Flies (Diptera: Tephritidae) Based on Pyrosequencing. Curr. Microbiol. 2018, 75, 966–976. [Google Scholar] [CrossRef]
- Blow, F.; Gioti, A.; Starns, D.; Ben-Yosef, M.; Pasternak, Z.; Jurkevitch, E.; Vontas, J.; Darby, A.C. Draft Genome Sequence of the Bactrocera oleae Symbiont “Candidatus Erwinia dacicola”. Genome Announc. 2016, 4, e00896–e00916. [Google Scholar] [CrossRef]
- Yun, J.H.; Roh, S.W.; Whon, T.W.; Jung, M.J.; Kim, M.S.; Park, D.S.; Yoon, C.; Nam, Y.D.; Kim, Y.J.; Choi, J.H.; et al. Insect Gut Bacterial Diversity Determined by Environmental Habitat, Diet, Developmental Stage, and Phylogeny of Host. Appl. Environ. Microbiol. 2014, 80, 5254–5264. [Google Scholar] [CrossRef]
- Zurek, K.; Nayduch, D. Bacterial Associations Across House Fly Life History: Evidence for Transstadial Carriage From Managed Manure. J. Insect Sci. 2016, 16, 2. [Google Scholar] [CrossRef]
- Winans, N.J.; Walter, A.; Chouaia, B.; Chaston, J.M.; Douglas, A.E.; Newell, P.D. A genomic investigation of ecological differentiation between free-living and -associated bacteria. Mol. Ecol. 2017, 26, 4536–4550. [Google Scholar] [CrossRef]
- Su, Z.J.; Zhang, M.J.; Liu, X.; Tong, L.; Huang, Y.D.; Li, G.H.; Pang, Y. Comparison of Bacterial Diversity in Wheat Bran and in the Gut of Larvae and Newly Emerged Adult of (Diptera: Muscidae) by Use of Ethidium Monoazide Reveals Bacterial Colonization. J. Econ. Entomol. 2010, 103, 1832–1841. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Greatest Length | Pupal Weight | Adult Weight | |
|---|---|---|---|
| C | 19.74 ± 0.33 a | 68.82 ± 4.07 b | 41.60 ± 3.69 b |
| H1 | 19.60 ± 0.92 a | 59.14 ± 1.55 a | 38.17 ± 5.02 b |
| H2 | 19.11 ± 0.56 a | 62.28 ± 2.19 a | 22.00 ± 6.75 a |
| df | 2 | 2 | 2 |
| F | 1.32 | 15.43 | 22.88 |
| Df | Sums of Sqs | MeanSqs | F.Model | R2 | p-Value (>F) | Signif | |
|---|---|---|---|---|---|---|---|
| Group factor | 14 | 9.115 | 0.65107 | 5.6351 | 0.7245 | 0.001 | *** |
| Residuals | 30 | 3.4662 | 0.11554 | 0.2755 | |||
| Total | 44 | 12.5812 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, H.; Zhang, X.; Yang, F.; Zhang, C.; Ngando, F.J.; Ren, L.; Guo, Y. Effect of Ciprofloxacin on the Composition of Intestinal Microbiota in Sarcophaga peregrina (Diptera: Sarcophagidae). Microorganisms 2023, 11, 2867. https://doi.org/10.3390/microorganisms11122867
Tang H, Zhang X, Yang F, Zhang C, Ngando FJ, Ren L, Guo Y. Effect of Ciprofloxacin on the Composition of Intestinal Microbiota in Sarcophaga peregrina (Diptera: Sarcophagidae). Microorganisms. 2023; 11(12):2867. https://doi.org/10.3390/microorganisms11122867
Chicago/Turabian StyleTang, Haojie, Xiangyan Zhang, Fengqin Yang, Changquan Zhang, Fernand Jocelin Ngando, Lipin Ren, and Yadong Guo. 2023. "Effect of Ciprofloxacin on the Composition of Intestinal Microbiota in Sarcophaga peregrina (Diptera: Sarcophagidae)" Microorganisms 11, no. 12: 2867. https://doi.org/10.3390/microorganisms11122867
APA StyleTang, H., Zhang, X., Yang, F., Zhang, C., Ngando, F. J., Ren, L., & Guo, Y. (2023). Effect of Ciprofloxacin on the Composition of Intestinal Microbiota in Sarcophaga peregrina (Diptera: Sarcophagidae). Microorganisms, 11(12), 2867. https://doi.org/10.3390/microorganisms11122867