

Characterization of Cronobacter sakazakii and Cronobacter malonaticus Strains Isolated from Powdered Dairy Products Intended for Consumption by Adults and Older Adults

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Isolation and Identification Methods of Cronobacter spp.
2.3. Whole-Genome Sequencing (WGS)
2.4. Sequence Type (ST) and Core Genome Multilocus Sequence Typing (cgMLST) of Cronobacter Isolates
2.5. Determination of Serotypes
2.6. Antibiotic Susceptibility
2.7. Adherence and Invasion Assays
2.8. Detection of Antibiotic Resistance and Virulence Genes
2.9. Detection of Plasmids and Mobile Genetic Elements (MGEs)
3. Results
3.1. Sampling and Identification of Isolates
3.2. Antibiotic Resistance Profile
3.3. Adherence and Invasion Assays
3.4. Detection of Antibiotic Resistance and Virulence Genes
3.5. Detection of Plasmids and Mobile Genetic Elements (MGEs)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iversen, C.; Mullane, N.; Mc Cardell, B.; Tall, B.; Lehner, A.; Fanning, S.; Stephan, R.; Joosten, H. Cronobacter gen. nov., a new genus to accommodate the biogroups of Enterobacter sakazakii, and proposal of Cronobacter sakazakii gen. nov.comb. nov., C. malonaticus sp. nov., C. turicensis sp. nov., C. muytjensii sp. nov., C. dublinensis sp. nov., Cronobacter genomospecies 1, and of three subspecies, C. dublinensis sp. nov.subsp. dublinensis subsp. nov., C. dublinensis sp. nov.subsp. lausannensis subsp. nov., and C. dublinensis sp. nov.subsp. lactaridi subsp. nov. Int. J. Syst. Evol. Microbiol. 2008, 58, 1442–1447. [Google Scholar] [PubMed]
- Joseph, S.; Cetinkaya, E.; Drahovska, H.; Levican, A.; Figueras, M.; Forsythe, S. Cronobacter condimenti sp. nov., isolated from spiced meat, and Cronobacter universalis sp. nov., a species designation for Cronobacter sp. genomospecies 1, recovered from a leg infection, water and food ingredients. Int. J. Syst. Evol. Microbiol. 2012, 62, 1277–1283. [Google Scholar] [CrossRef] [PubMed]
- Stephan, R.; Grim, C.; Gopinath, G.; Mammel, M.; Sathyamoorthy, V.; Trach, L.; Chase, H.; Fanning, S.; Tall, B. Re-examination of the taxonomic status of Enterobacter helveticus, Enterobacter pulveris and Enterobacter turicensis as members of the genus Cronobacter and their reclassification in the genera Franconibacter gen. nov. and Siccibacter gen. nov. as Franconibacter helveticus comb. nov., Franconibacter pulveris comb. nov. and Siccibacter turicensis comb. nov., respectively. Int. J. Syst. Evol. Microbiol. 2014, 64, 3402–3410. [Google Scholar] [PubMed]
- Forsythe, S.J. Updates on the Cronobacter genus. Annu. Rev. Food Sci. Technol. 2018, 25, 23–44. [Google Scholar] [CrossRef]
- Hariri, S.; Joseph, S.; Forsythe, S.J. Cronobacter sakazakii ST4 strains and neonatal meningitis, United States. Emerg. Infect. Dis. 2013, 19, 175–177. [Google Scholar] [CrossRef]
- Holý, O.; Forsythe, S. Cronobacter spp. as emerging causes of healthcare-associated infection. J. Hosp. Infect. 2014, 86, 169–177. [Google Scholar] [CrossRef]
- Jaradat, Z.W.; Al Mousa, W.; Elbetieha, A.; Al Nabulsi, A.; Tall, B.D. Cronobacter spp. opportunistic food-borne pathogens. A review of their virulence and environmental-adaptive traits. J. Med. Microbiol. 2014, 63, 1023–1037. [Google Scholar] [CrossRef]
- Joseph, S.; Forsythe, S.J. Predominance of Cronobacter sakazakii sequence type 4 in neonatal infections. Emerg. Infect. Dis. 2011, 17, 1713–1715. [Google Scholar] [CrossRef]
- Alsonosi, A.; Hariri, S.; Kajsík, M.; Oriešková, M.; Hanulík, V.; Röderová, M.; Petrželová, J.; Kollárová, H.; Drahovská, H.; Forsythe, S.; et al. The speciation and genotyping of Cronobacter isolates from hospitalised patients. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1979–1988. [Google Scholar] [CrossRef]
- Ling, N.; Li, C.; Zhang, J.; Wu, Q.; Zeng, H.; He, W.; Ye, Y.; Wang, J.; Ding, Y.; Chen, M.; et al. Prevalence and molecular and antimicrobial characteristics of Cronobacter spp. isolated from raw vegetables in China. Front. Microbiol. 2018, 9, 1149. [Google Scholar] [CrossRef]
- Holý, O.; Petrželová, J.; Hanulík, V.; Chromá, M.; Matoušková, I.; Forsythe, S. Epidemiology of Cronobacter spp. isolates from patients admitted to the Olomouc University Hospital (Czech Republic). Epidemiol. Mikrobiol. Imunol. 2014, 63, 69–72. [Google Scholar] [PubMed]
- Yong, W.; Guo, B.; Shi, X.; Cheng, T.; Chen, M.; Jiang, X.; Ye, Y.; Wang, J.; Xie, G.; Ding, J. An investigation of an acute gastroenteritis outbreak: Cronobacter sakazakii, a potential cause of food-borne illness. Front. Microbiol. 2018, 9, 2549. [Google Scholar] [CrossRef] [PubMed]
- Parra-Flores, J.; Maury-Sintjago, E.; Rodriguez-Fernández, A.; Acuña, S.; Cerda, F.; Aguirre, J.; Holy, O. Microbiological quality of powdered infant formula in Latin America. J. Food Prot. 2020, 83, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Brandão, M.L.; Umeda, N.S.; Jackson, E.; Forsythe, S.J.; de Filippis, I. Isolation, molecular and phenotypic characterization, and antibiotic susceptibility of Cronobacter spp. from Brazilian retail foods. Food Microbiol. 2017, 63, 129–138. [Google Scholar] [CrossRef]
- Vojkovska, H.; Karpiskova, R.; Orieskova, M.; Drahovska, H. Characterization of Cronobacter spp. isolated from food of plant origin and environmental samples collected from farms and from supermarkets in the Czech Republic. Int. J. Food Microbiol. 2016, 217, 130–136. [Google Scholar] [CrossRef]
- Franco, A.A.; Hu, L.; Grim, C.J.; Gopinath, G.; Sathyamoorthy, V.; Jarvis, K.G.; Lee, C.; Sadowski, J.; Kim, J.; Kothary, M.H.; et al. Characterization of putative virulence genes on the related RepFIB plasmids harbored by Cronobacter spp. Appl. Environ. Microbiol. 2011, 77, 3255–3267. [Google Scholar] [CrossRef]
- Aly, M.A.; Domig, K.; Kneifel, W.; Reimhult, E. Whole genome sequencing-based comparison of food isolates of Cronobacter sakazakii. Front. Microbiol. 2019, 10, 1464. [Google Scholar] [CrossRef]
- Cruz, A.; Xicohtencatl, J.; Gonzalez, B.; Bobadilla, M.; Eslava, C.; Rosas, I. Virulence traits in Cronobacter species isolated from different sources. Can. J. Microbiol. 2011, 57, 735–744. [Google Scholar] [CrossRef]
- Holý, O.; Cruz-Cordova, A.; Xicohtencatl-Cortés, J.; Hochel, I.; Parra-Flores, J.; Petrzelova, J.; Facevicova, K.; Forsythe, S.; Alsonosi, A. Occurrence of virulence factors in Cronobacter sakazakii and Cronobacter malonaticus originated from clinical samples. Microb. Pathog. 2019, 127, 250–256. [Google Scholar] [CrossRef]
- Singh, N.; Goel, G.; Raghav, M. Insights into virulence factors determining the pathogenicity of Cronobacter sakazakii. Virulence 2015, 6, 433–440. [Google Scholar] [CrossRef]
- Cruz-Córdova, A.; Rocha-Ramírez, L.; Ochoa, S.; Gónzalez-Pedrajo, B.; Espinosa, N.; Eslava, C.; Hernández-Chiñas, U.; Mendoza-Hernández, G.; Rodríguez-Leviz, A.; Valencia-Mayoral, P.; et al. Flagella from five Cronobacter species induce pro-inflammatory cytokines in macrophage derivatives from human monocytes. PLoS ONE 2012, 7, e52091. [Google Scholar] [CrossRef] [PubMed]
- Aldubyan, M.; Almami, I.; Benslimane, F.; Alsonosi, A.; Forsythe, S. Comparative outer membrane protein analysis of high and low-invasive strains of Cronobacter malonaticus. Front. Microbiol. 2017, 8, 2268. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, H.; Winter, J.; Boocock, D.; De Girolamo, L.; Forsythe, S.J. Characterization of outer membrane vesicles from a neonatal meningitic strain of Cronobacter sakazakii. FEMS Microbiol. Lett. 2015, 362, fnv085. [Google Scholar] [CrossRef] [PubMed]
- Flores, J.P.; Arvizu, S.; Silva, J.; Fernández, E. Two cases of hemorrhagic diarrhea caused by Cronobacter sakazakii in hospitalized nursing infants associated with the consumption of powdered infant formula. J. Food Prot. 2011, 74, 2177–2181. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.D.; Park, J.; Chang, H. Detection, antibiotic susceptibility and biofilm formation of Cronobacter spp. from various foods in Korea. Food Control 2012, 24, 225–230. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Ling, N.; Shen, Y.; Zhang, D.; Liu, D.; Ou, D.; Wu, Q.; Ye, Y. Effects of tolC on tolerance to bile salts and biofilm formation in Cronobacter malonaticus. J. Dairy Sci. 2021, 104, 9521–9531. [Google Scholar] [CrossRef]
- Leopold, S.; Goering, R.; Witten, A.; Harmsen, D.; Mellmann, A. Bacterial whole-genome sequencing revisited: Portable, scalable, and standardized analysis for typing and detection of virulence and antibiotic resistance genes. J. Clin. Microbiol. 2014, 52, 2365–2370. [Google Scholar] [CrossRef]
- Iversen, C.; Forsythe, S.J. Isolation of Enterobacter sakazakii and other Enterobacteriaceae from powdered infant formula milk and related products. Food Microbiol. 2004, 21, 771–776. [Google Scholar] [CrossRef]
- Lepuschitz, S.; Sorschag, S.; Springer, B.; Allerberger, F.; Ruppitsch, W. Draft genome sequence of carbapenemase-producing Serratia marcescens isolated from a patient with chronic obstructive pulmonary disease. Genome Announc. 2017, 5, e01288-17. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bliss, C.M.; Bennett, J.S.; Bratcher, H.B.; Brehony, C.; Colles, F.M.; Wimalarathna, H.; Harrison, O.B.; Sheppard, S.K.; Cody, A.J.; et al. Ribosomal multilocus sequence typing: Universal characterization of bacteria from domain to strain. Microbiology 2012, 158, 1005–1015. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.R.; Enns, E.; Marinier, E.; Mandal, A.; Herman, E.K.; Chen, C.Y.; Graham, M.; Van Domselaar, G.; Stothard, P. Proksee: In-depth characterization and visualization of bacterial genomes. Nucleic Acids Res. 2023, 51, W484–W492. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Jünemann, S.; Sedlazeck, F.J.; Prior, K.; Albersmeier, A.; John, U.; Kalinowski, J.; Mellmann, A.; Goesmann, A.; von Haesler, A.; Stoye, J.; et al. Updating benchtop sequencing performance comparison. Nat. Biotechnol. 2013, 31, 294–296. [Google Scholar] [CrossRef]
- Ogrodzki, P.; Forsythe, S. Capsular profiling of the Cronobacter genus and the association of specific Cronobacter sakazakii and C. malonaticus capsule types with neonatal meningitis and necrotizing enterocolitis. BMC Genom. 2015, 16, 758. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI supplement M100; Clinical and Laboratory Standars Institute: Wayne, PA, USA, 2020; pp. 1–263. [Google Scholar]
- Bortolaia, V.; Kaas, R.F.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.R.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.; Lago, B.; Dave, B.; Pereira, S.; Sharma, A.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.H.; McDermott, P.F.; et al. Validating the AMRFinder tool and resistance gene database by using antimicrobial resistance genotype-phenotype correlations in a collection of isolates. Antimicrob. Agents Chemother. 2019, 63, e00483-19. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.; Nash, J.H.E. MOB-suite: Software tools for clustering, reconstruction and typing of plasmids from draft assemblies. Microb. Genom. 2018, 4, e000206. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Kadlicekova, V.; Kajsik, M.; Soltys, K.; Szemes, T.; Slobodnikova, L.; Janosikova, L.; Hubenakova, Z.; Ogrodzki, P.; Forsythe, S.; Turna, J.; et al. Characterisation of Cronobacter strains isolated from hospitalised adult patients. Antonie van Leeuwenhoek 2018, 111, 1073–1085. [Google Scholar] [CrossRef] [PubMed]
- Parra-Flores, J.; Cerda-Leal, F.; Contreras, A.; Valenzuela-Riffo, N.; Rodriguez, A.; Aguirre, J. Cronobacter sakazakii and microbiological parameters in dairy formulas associated with a food alert in Chile. Front. Microbiol. 2018, 9, 1708. [Google Scholar] [CrossRef]
- Lepuschitz, S.; Ruppitsch, W.; Pekard-Amenitsch, S.; Forsythe, S.J.; Cormican, M.; Mach, R.L.; Piérard, D.; Allerberger, F.; The EUCRONI Study Group. Multicenter study of Cronobacter sakazakii infections in humans, Europe, 2017. Emerg. Infect. Dis. 2019, 25, 515–522. [Google Scholar] [CrossRef]
- Ruppitsch, W.; Pietzka, A.; Prior, K.; Bletz, S.; Fernandez, H.L.; Allerberger, F.; Harmsen, D.; Mellmann, A. Defining and evaluating a core genome multilocus sequence typing scheme for whole-genome sequence-based typing of Listeria monocytogenes. J. Clin. Microbiol. 2015, 53, 2869–2876. [Google Scholar] [CrossRef]
- Serwecińska, L. Antimicrobials and Antibiotic-Resistant Bacteria: A risk to the environment and to Public Health. Water 2020, 12, 3313. [Google Scholar] [CrossRef]
- Molloy, C.; Cagney, C.; O’Brien, S.; Iversen, C.; Fanning, S.; Duffy, G. Surveillance and characterization by Pulsed-Field Gel Electrophoresis of Cronobacter spp in farming and domestic environments, food production animals and retails foods. Int. J. Food Microbiol. 2009, 136, 198–238. [Google Scholar] [CrossRef]
- Arslan, S.; Ertürk, H.G. Occurrence, virulence and antimicrobial susceptibility profiles of Cronobacter spp. from ready-to-eat foods. Curr. Microbiol. 2021, 78, 3403–3416. [Google Scholar] [CrossRef]
- Li, Q.; Li, C.; Ye, Q.; Gu, Q.; Wu, S.; Zhang, Y.; Wei, X.; Xue, L.; Chen, M.; Zeng, H.; et al. Occurrence, molecular characterization and antibiotic resistance of Cronobacter spp. isolated from wet rice and flour products in Guangdong, China. Curr. Res. Food Sci. 2023, 7, 100554. [Google Scholar] [CrossRef] [PubMed]
- López-Pueyo, M.J.; Barcenilla-Gaite, F.; Amaya-Villar, R.; Garnacho-Montero, J.J. Multirresistencia antibiótica en unidades de críticos. Med. Intensiva 2011, 35, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Wolfensberger, A.; Kuster, S.P.; Marchesi, M.; Zbinden, R.; Hombach, M. The effect of varying multidrug-resistence (MDR) definitions on rates of MDR gram-negative rods. Antimicrob. Resist. Infect. Control 2019, 8, 193. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Lei, T.; He, W.; Zhang, J.; Liang, B.; Li, C.; Ling, N.; Ding, Y.; Wu, S.; Wang, J.; et al. Novel multidrug-resistant Cronobacter sakazakii causing meningitis in neonate, China, 2015. Emerg. Infect. Dis. 2018, 24, 2121–2124. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.H.; Yu, B.; Xiang, Y.; Zhang, Z.; Zhang, T.; Zeng, Y.C.; Cui, Z.G.; Huo, X.X. Two cases of multi-antibiotic resistant Cronobacter spp. infections of infants in China. Biomed. Environ. Sci. 2017, 30, 601–605. [Google Scholar]
- Pakbin, B.; Brück, W.M.; Allahyari, S.; Rossen, J.W.A.; Mahmoudi, R. Antibiotic resistance and molecular characterization of Cronobacter sakazakii strains isolated from powdered infant formula milk. Foods 2022, 11, 1093. [Google Scholar] [CrossRef]
- Fei, P.; Jiang, Y.; Feng, J.; Forsythe, S.J.; Li, R.; Zhou, Y.; Man, C. Antibiotic and desiccation resistance of Cronobacter sakazakii and C. malonaticus isolates from powdered infant formula and processing environments. Front. Microbiol. 2017, 8, 316. [Google Scholar] [CrossRef]
- Gan, X.; Li, M.; Xu, J.; Yan, S.; Wang, W.; Li, F. Emerging of multidrug-resistant Cronobacter sakazakii isolated from infant supplementary food in China. Microbiol. Spectr. 2022, 10, e0119722. [Google Scholar] [CrossRef]
- Quintero-Villegas, M.; Wittke, A.; Hutkins, R. Adherence inhibition of Cronobacter sakazakii to intestinal epithelial cells by lactoferrin. Curr. Microbiol. 2014, 69, 574–579. [Google Scholar] [CrossRef]
- Mange, J.P.; Stephan, R.; Borel, L.; Wild, P.; Kim, K.S.; Pospischil, A.; Lenher, A. Adhesive properties of Enterobacter sakazakii to human epithelial and brain microvascular endothelial cells. BMC Microbiol. 2006, 6, 58–68. [Google Scholar] [CrossRef]
- Townsend, S.; Hurrell, E.; Gonzalez-Gomez, I.; Lowe, J.; Frye, J.; Forsythe, S.; Badger, J. Enterobacter sakazakii invades brain capillary endothelial cells, persists in human macrophages influencing cytokine secretion and induces severe brain pathology in the neonatal rat. Microbiology 2007, 153, 3538–3547. [Google Scholar] [CrossRef] [PubMed]
- Parra-Flores, J.; Aguirre, J.; Juneja, V.; Jackson, E.; Cruz, A.; Silva, J.; Forsythe, S. Virulence and antibiotic resistance profiles of Cronobacter sakazakii and Enterobacter spp. involved in the diarrheic hemorrhagic outbreak in Mexico. Front. Microbiol. 2018, 9, 2206. [Google Scholar] [CrossRef] [PubMed]
- Holý, O.; Parra-Flores, J.; Lepuschitz, S.; Alarcón-Lavín, M.P.; Cruz-Córdova, A.; Xicohtencatl-Cortes, J.; Mancilla-Rojano, J.; Ruppitsch, W.; Forsythe, S. Molecular characterization of Cronobacter sakazakii strains isolated from powdered milk. Foods 2021, 10, 20. [Google Scholar] [CrossRef]
- Müller, A.; Hächler, H.; Stephan, R.; Lehner, A. Presence of AmpC beta-lactamases, CSA-1, CSA-2, CMA-1, and CMA-2 conferring an unusual resistance phenotype in Cronobacter sakazakii and Cronobacter malonaticus. Microb. Drug Resist. 2014, 20, 275–280. [Google Scholar] [CrossRef]
- Jang, H.; Chase, H.R.; Gangiredla, J.; Grim, C.J.; Patel, I.R.; Kothary, M.H.; Jackson, S.A.; Mammel, M.K.; Carter, L.; Negrete, F.; et al. Analysis of the molecular diversity among Cronobacter species isolated from filth flies using targeted PCR, pan genomic DNA microarray, and whole genome sequencing analyses. Front. Microbiol. 2020, 11, 561204. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; Eshwar, A.; Lehner, A.; Gangiredla, J.; Patel, I.R.; Beaubrun, J.J.-G.; Chase, H.R.; Negrete, F.; Finkelstein, S.; Weinstein, L.M.; et al. Characterization of Cronobacter sakazakii strains originating from plant-origin foods using comparative genomic analyses and zebrafish infectivity studies. Microorganisms 2022, 10, 1396. [Google Scholar] [CrossRef]
- Fei, P.; Jing, H.; Ma, Y.; Dong, G.; Chang, Y.; Meng, Z.; Jiang, S.; Xie, Q.; Li, S.; Chen, X.; et al. Cronobacter spp. in commercial powdered infant formula collected from nine provinces in China: Prevalence, genotype, biofilm formation, and antibiotic susceptibility. Front. Microbiol. 2022, 13, 900690. [Google Scholar] [CrossRef]
- Jaradat, Z.W.; Al-Mousa, W.A.; Elbetieha, A.M.; Ababneh, Q.O.; Al-Nabulsi, A.A.; Jang, H.; Gangiredla, J.; Patel, I.R.; Gopinath, G.R.; Tall, B.D. Virulence, antimicrobial susceptibility and phylogenetic analysis of Cronobacter sakazakii isolates of food origins from Jordan. J. Appl. Microbiol. 2022, 133, 2528–2546. [Google Scholar] [CrossRef]
- Shi, L.; Liang, Q.; Zhan, Z.; Feng, J.; Zhao, Y.; Chen, Y.; Huang, M.; Tong, Y.; Wu, W.; Chen, W.; et al. Co-occurrence of 3 different resistance plasmids in a multi-drug resistant Cronobacter sakazakii isolate causing neonatal infections. Virulence 2018, 9, 110–112. [Google Scholar] [CrossRef]
- Parra-Flores, J.; Holý, O.; Riffo, F.; Lepuschitz, S.; Ruppitsch, W.; Forsythe, S. Draft Genome Sequences of seven Cronobacter sakazakii strains carrying the mcr 9.1 gene isolated in Chile. Microbiol. Resour. Announc. 2021, 10, e0050621. [Google Scholar] [CrossRef]
- Franco, A.A.; Kothary, M.; Gopinath, G.; Jarvis, K.; Grim, C.J.; Hu, L.; Datta, R.; McCardell, B.; Tall, B. Cpa, the outer membrane protease of Cronobacter sakazakii, activates plasminogen and mediates resistance to serum bactericidal activity. Infect. Immun. 2011, 79, 1578–1587. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.; Hariri, S.; Masood, N.; Forsythe, S. Sialic acid utilization by Cronobacter sakazakii. Microb. Inform. Exp. 2013, 3, e3. [Google Scholar] [CrossRef] [PubMed]
- Severi, E.; Hood, D.W.; Thomas, G.H. Sialic acid utilization by bacterial pathogens. Microbiology 2007, 153, 2817–2822. [Google Scholar] [CrossRef]
- Sohanpal, B.K.; Friar, S.; Roobol, J.; Plumbridge, J.A.; Blomfield, I.C. Multiple co-regulatory elements and IHF are necessary for the control of fimB expression in response to sialic acid and N-acetylglucosamine in Escherichia coli K-12. Mol. Microbiol. 2007, 63, 1223–1236. [Google Scholar] [CrossRef]
- Esteban-Kenel, V.; Ochoa, S.; Curiel-Quesada, E.; Quezada, H.; Medina-Contreras, O.; Fernández-Rendón, E.; Rosas-Pérez, I.; Arellano-Galindo, J.; Cisneros, B.; Hernandez-Castro, R.; et al. Proteomics profiles of Cronobacter sakazakii and a fliF mutant: Adherence and invasion in mouse neuroblastoma cells. Microb. Pathog. 2020, 149, 104595. [Google Scholar]
- Kim, K.; Kim, K.P.; Choi, J.; Lim, J.A.; Lee, J.; Hwang, S.; Ryu, S. Outer membrane proteins A (OmpA) and X (OmpX) are essential for basolateral invasion of Cronobacter sakazakii. Appl. Environ. Microbiol. 2010, 76, 5188–5198. [Google Scholar] [CrossRef] [PubMed]
- Mohan Nair, M.K.; Venkitanarayanan, K. Role of bacterial OmpA and host cytoskeleton in the invasion of human intestinal epithelial cells by Enterobacter sakazakii. Pediatr. Res. 2007, 62, 664–669. [Google Scholar] [CrossRef]
- Walling, L.R.; Butler, J.S. Toxins targeting transfer RNAs: Translation inhibition by bacterial toxin–antitoxin systems. Wiley Interdiscip. Rev. RNA 2019, 10, e1506. [Google Scholar] [CrossRef]
- Finkelstein, S.; Negrete, F.; Jang, H.; Gangiredla, J.; Mammel, M.; Patel, I.R.; Chase, H.R.; Woo, J.; Lee, Y.; Wang, C.Z.; et al. Prevalence, distribution, and phylogeny of type two toxin-antitoxin genes possessed by Cronobacter species where C. sakazakii homologs follow sequence type lineages. Microorganisms 2019, 7, 554. [Google Scholar] [CrossRef]
- Lachowska, M.; Izdebski, R.; Urbanowicz, P.; Żabicka, D.; Królak-Olejnik, B. Infection of Cronobacter sakazakii ST1 producing SHV-12 in a premature infant born from triplet pregnancy. Microorganisms 2021, 9, 1878. [Google Scholar] [CrossRef]
- Yan, Q.; Power, K.A.; Cooney, S.; Fox, E.; Gopinath, G.R.; Grim, C.J.; Tall, B.D.; McCusker, M.P.; Fanning, S. Complete genome sequence and phenotype microarray analysis of Cronobacter sakazakii SP291: A persistent isolate cultured from a powdered infant formula production facility. Front. Microbiol. 2013, 4, 256. [Google Scholar] [CrossRef] [PubMed]
- Moine, D.; Kassam, M.; Baert, L.; Tang, Y.; Barretto, C.; Bru, C.N.; Klijn, C.; Descombes, P. Fully closed genome sequences of five type strains of the genus Cronobacter and one Cronobacter sakazakii strain. Genome Announc. 2016, 4, e00142-16. [Google Scholar] [CrossRef] [PubMed]
- Svobodová, B.; Vlach, J.; Junková, P.; Karamonová, L.; Blažková, M.; Fukal, L. Novel method for reliable identification of Siccibacter and Franconibacter strains: From "Pseudo-Cronobacter" to New Enterobacteriaceae genera. Appl. Environ. Microbiol. 2017, 83, e00234-17. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Forsythe, S.J.; Yang, X.; Fu, S.; Man, C.; Jiang, Y. Invited review: Stress resistance of Cronobacter spp. affecting control of its growth during food production. J. Dairy Sci. 2021, 104, 11348–11367. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, Z.E.; Hunt, K.; Koolman, L.; Butler, F.; Fanning, S. Cronobacter species in the built food production environment: A review on persistence, pathogenicity, regulation and detection methods. Microorganisms 2023, 11, 1379. [Google Scholar] [CrossRef]
 .
.
.
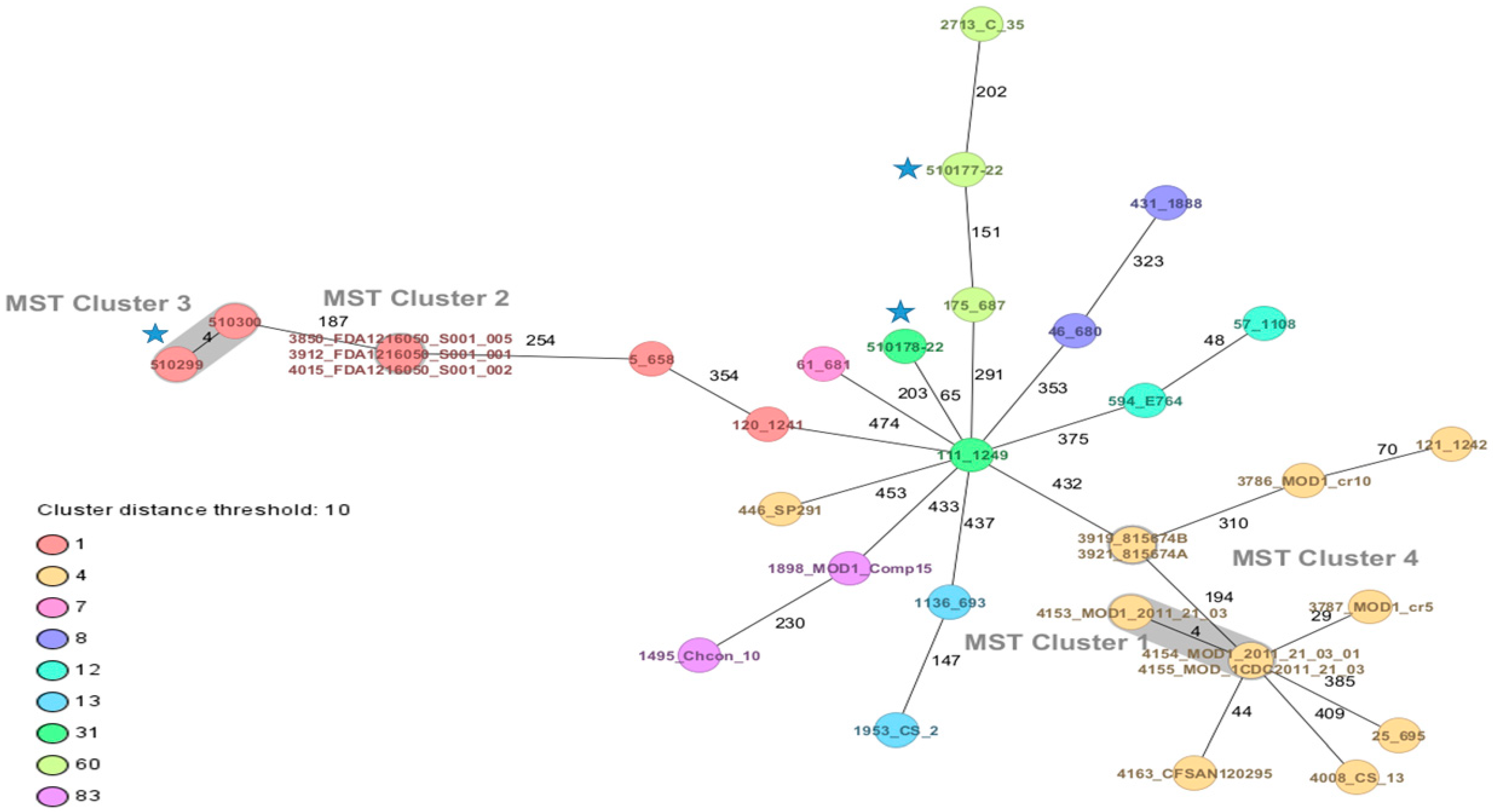
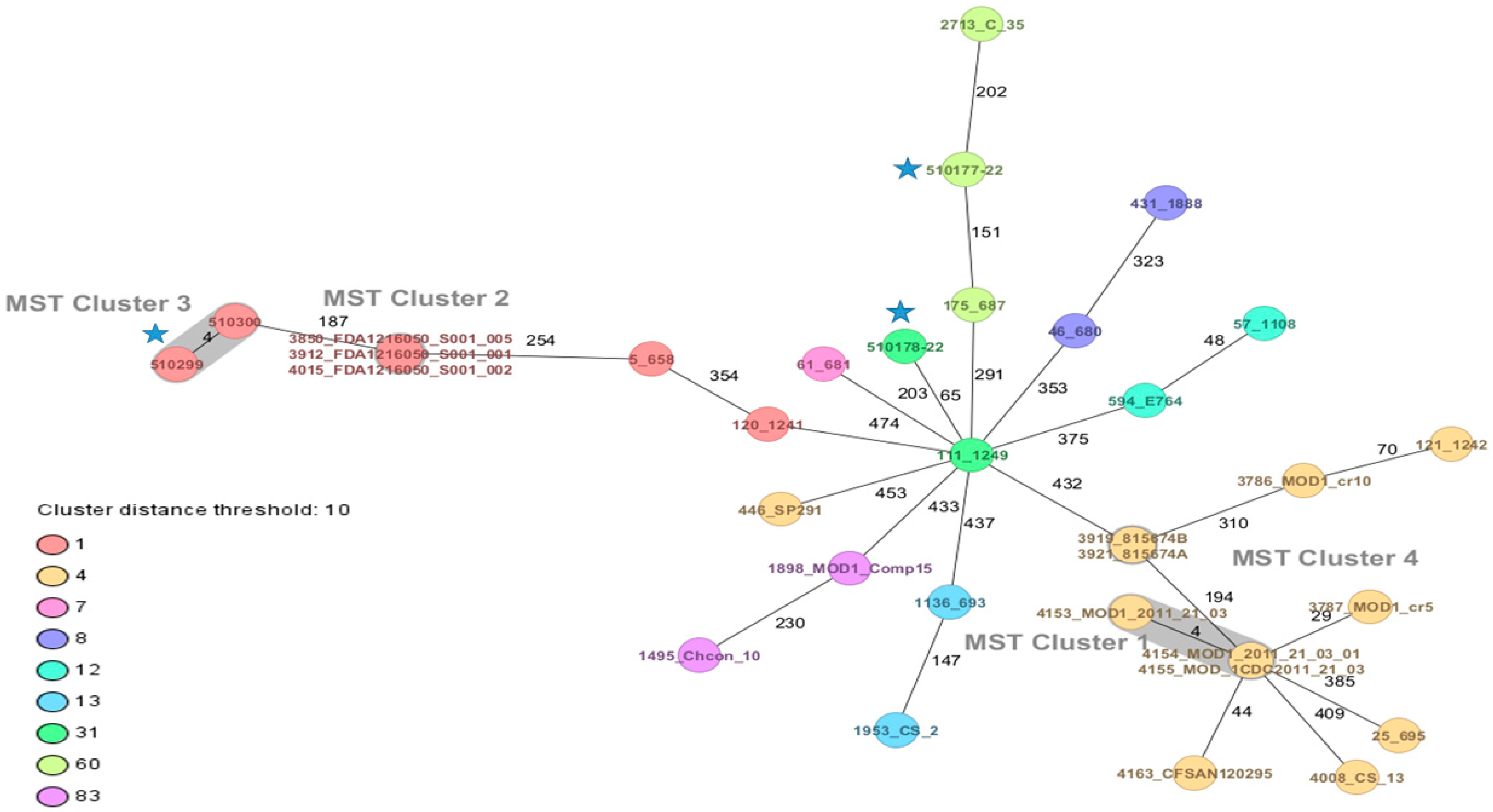
.
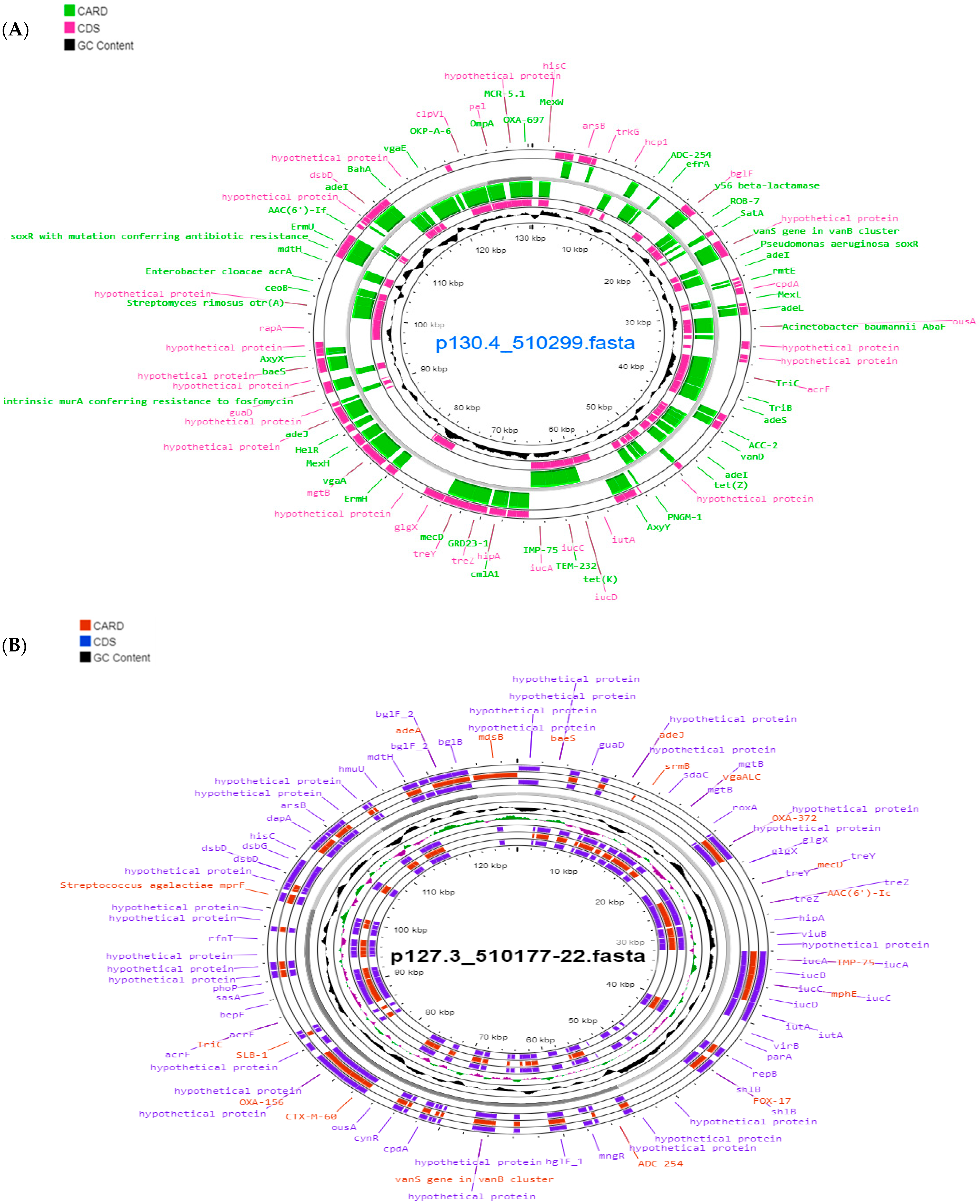

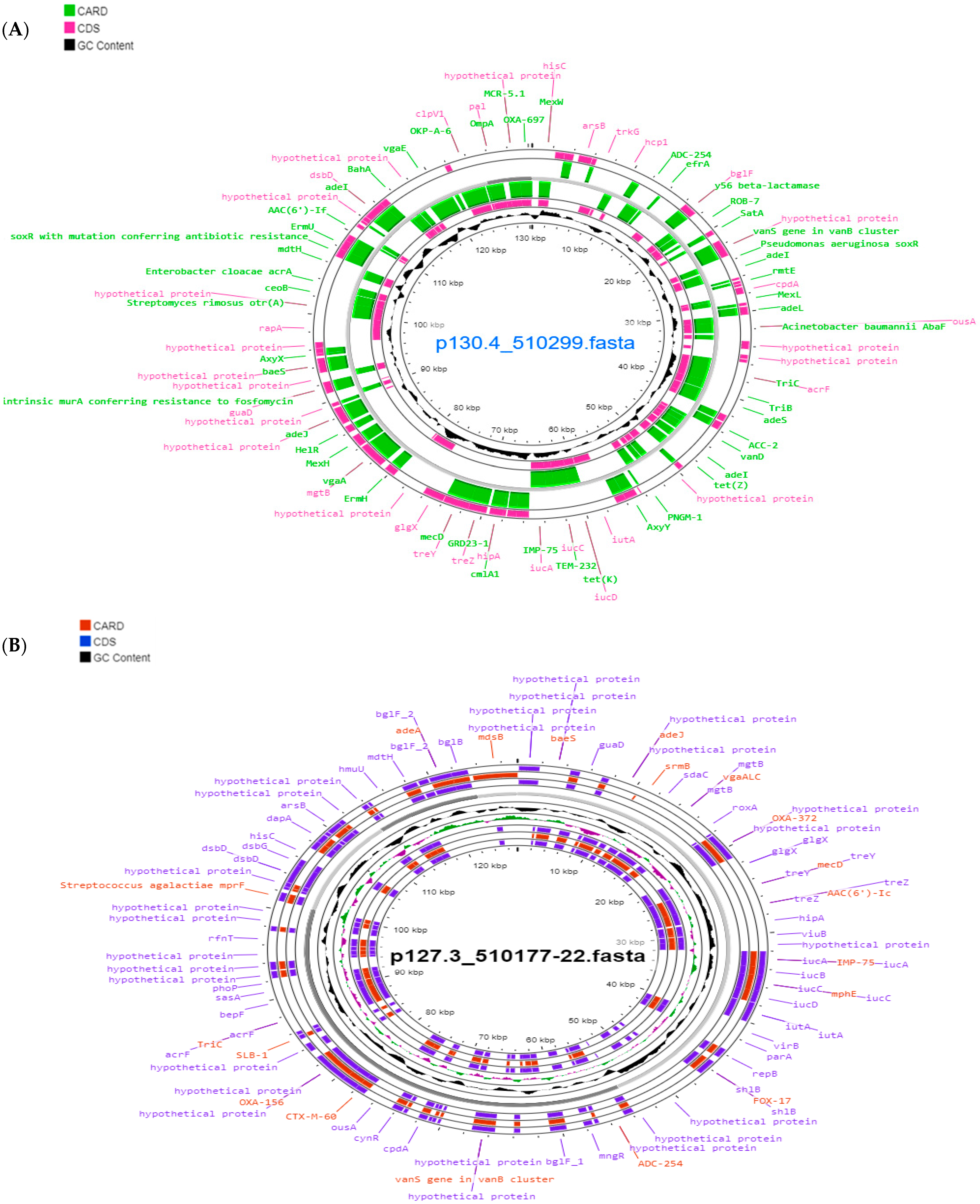
{kind=link}
{kind=link}
| Sample ID | PubMLST ID * | MALDI-TOF | WGS rMLST Result ** | ST | CC | Serotype (gnd-galF Alleles) | Collection Date |
|---|---|---|---|---|---|---|---|
| 510299 | 4183 | Cronobacter spp. | Cronobacter sakazakii | 1 | 1 | Csak O:1 | 2018 |
| 510300 | 4184 | Cronobacter spp. | Cronobacter sakazakii | 1 | 1 | Csak O:1 | 2018 |
| 510178-22 | 4187 | Cronobacter spp. | Cronobacter sakazakii | 31 | 31 | Csak O:2 | 2019 |
| 510177-22 | 4188 | Cronobacter spp. | Cronobacter malonaticus | 60 | 60 | Cmal O:1 | 2019 |
| 510441-19 | 4189 | Franconibacter helveticus | Franconibacter helveticus | 345 | ND | Fhelv O:1 | 2019 |
| Strains | Species | AK (30 µg) | AM (10 µg) | AMC (20/10 µg) | CAZ (30 µg) | CIP (5 µg) | CL (30 µg) | CTX (30 µg) | GE (10 µg) | KF (30 µg) | TE (30 µg) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 510299 | C. sakazakii ST1 | R | R | S | R | S | S | S | S | R | S |
| 510300 | C. sakazakii ST1 | R | R | S | R | S | S | S | S | R | S |
| 510178-22 | C. sakazakii ST31 | S | S | S | S | S | S | S | S | R | S |
| 510177-22 | C. malonaticus ST60 | S | R | R | S | S | S | S | S | R | S |
| 510441-19 | F. helveticus ST345 | S | S | S | R | S | S | S | S | S | S |
| Strain ID | ST | Resistance Profile | Resistance Genes |
|---|---|---|---|
| 510299 | ST1 | AK, AM, CAZ, KF | mcr-9.1, CSA-1, adeF, CRP, emrBR, EF-Tu, GlpT, fosA8, H-NS, PBP3, KpnF, marA, msbA, qacG, rsmA, AcrAB-TolC with MarR mutations |
| 510300 | ST1 | AK, AM, CAZ, KF | mcr-9.1, CSA-1, adeF, CRP, emrB, EF-Tu, GlpT, fosA8, H-NS, PBP3, KpnFE, marA, msbA, qacG, rsmA, vanG, AcrAB-TolC with MarR mutations |
| 510178-22 | ST31 | KF | CSA-1, KpnEF, marA, qacG, CRP, adeF, vanG, emrRB, rsmA, H-NS, fosA8, msbA, PBP3, GlpT, AcrAB-TolC with MarR mutations |
| 510177-22 | ST60 | AM, AMC, KF | CMA-1, adeF, rsmA, H-NS, adeF, KpnEF, marA, vanG, CRP, qacG, fosA8, msbA, emrBR, PBP3, GlpT |
| 510441-19 | ND | CAZ | fosA5, qacJ, marA, KpnFEH, adeF, baeR, rsmA, emrR, msbA, CRP, GlpT, PBP3, AcrAB-TolC with MarR mutations |
| Virulence Gene | Function | C. sakazakii ST1 (510299) | C. sakazakii ST1 (510300) | C. sakazakii S31 (510178-22) | C. malonaticus ST60 (510177-22) |
|---|---|---|---|---|---|
| flgB | motility | + | + | + | + |
| flgK | flagellar hook-associated protein 1 | + | + | + | + |
| flgL | flagellar hook-associated protein 3 | + | + | + | + |
| flgM | negative regulator of flagellin synthesis | + | + | + | + |
| flgN | flagellar synthesis FlgN protein | + | + | + | + |
| flhD | flagellar hook-associated protein 2 | + | + | + | + |
| fliA | flagellar operon FliA | + | + | + | + |
| fliC | flagellin | + | + | + | + |
| fliD | flagellar hook-associated protein 2 | + | + | + | + |
| fliR | flagellar biosynthetic FliR protein | + | + | + | + |
| fliT | flagellar FliT protein | + | + | + | + |
| fliZ | FliZ protein | + | + | + | + |
| lolA | outer membrane lipoprotein carrier protein | + | + | + | + |
| motB | chemotaxis MotA protein | + | + | + | + |
| sdiA | LuxR family transcriptional regulator | + | + | + | + |
| slyB | outer membrane lipoprotein SlyB | + | + | + | + |
| tolC | outer membrane channel protein | + | + | + | + |
| msbA | survival in macrophage | + | + | + | + |
| mviN | protective immunity and colonization | + | + | + | + |
| cpa | plasminogen activator | + | + | + | - |
| hem | hemolysins | + | + | + | + |
| ompA | adhesion cell; biofilm formation | + | + | + | + |
| ompX | adhesion cell | + | + | + | + |
| cheR | chemotaxis protein methyltransferase | + | + | + | + |
| cheY | response regulator of chemotaxis family | + | + | + | + |
| cheB | desiccation tolerance | + | + | + | + |
| lpxA | epithelial cell invasion and lipid A production | + | + | + | + |
| nanA,K,T | exogenous sialic acid utilization | + | + | + | - |
| ibpA | small heat shock protein | + | + | + | + |
| wzzB | desiccation tolerance | + | + | + | + |
| fic | cell filamentation protein | + | + | + | + |
| hsp20 | small shock protein | - | - | - | - |
| relB | RelE antitoxin | + | + | + | + |
| ID Strain | Specie | ST | Plasmid | Plasmid Accession Number | Size (Kb) | Mobile Genetic Elements |
|---|---|---|---|---|---|---|
| 510299 | C. sakazakii | 1 | pESA3 BR10-DEC | CP000785 CP035364 | 130,366 13,688 | IS5075, ISEsa2, ISEsa1, IS26, IS903, ISPpu12, IS102 |
| 510300 | C. sakazakii | 1 | pESA3 pCS36-4CPA | CP000785 KM373703 | 130,600 3959 | IS5075, IS26, ISPpu12, IS102, ISEsa2, ISEsa1 |
| 510178-22 | C. sakazakii | 31 | p109.3 (pSP291-1) p49.7 (pCMA2) | CP004092 CP013942 | 109,330 49,749 | ISEsa1 |
| 510177-22 | C. malonaticus | 60 | p127.3 (pCMA1) | CP013941 | 127,318 | ISSen4 |
| 510441-19 | F. helveticus | 345 | p24.7_510441-19 p14.9_510441-19 | CP023876 CP035364 | 24,653 14,863 | ISPpu12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parra-Flores, J.; Flores-Soto, F.; Flores-Balboa, C.; Alarcón-Lavín, M.P.; Cabal-Rosel, A.; Daza-Prieto, B.; Springer, B.; Cruz-Córdova, A.; Leiva-Caro, J.; Forsythe, S.; et al. Characterization of Cronobacter sakazakii and Cronobacter malonaticus Strains Isolated from Powdered Dairy Products Intended for Consumption by Adults and Older Adults. Microorganisms 2023, 11, 2841. https://doi.org/10.3390/microorganisms11122841
Parra-Flores J, Flores-Soto F, Flores-Balboa C, Alarcón-Lavín MP, Cabal-Rosel A, Daza-Prieto B, Springer B, Cruz-Córdova A, Leiva-Caro J, Forsythe S, et al. Characterization of Cronobacter sakazakii and Cronobacter malonaticus Strains Isolated from Powdered Dairy Products Intended for Consumption by Adults and Older Adults. Microorganisms. 2023; 11(12):2841. https://doi.org/10.3390/microorganisms11122841
Chicago/Turabian StyleParra-Flores, Julio, Fernanda Flores-Soto, Carolina Flores-Balboa, María P. Alarcón-Lavín, Adriana Cabal-Rosel, Beatriz Daza-Prieto, Burkhard Springer, Ariadnna Cruz-Córdova, José Leiva-Caro, Stephen Forsythe, and et al. 2023. "Characterization of Cronobacter sakazakii and Cronobacter malonaticus Strains Isolated from Powdered Dairy Products Intended for Consumption by Adults and Older Adults" Microorganisms 11, no. 12: 2841. https://doi.org/10.3390/microorganisms11122841
APA StyleParra-Flores, J., Flores-Soto, F., Flores-Balboa, C., Alarcón-Lavín, M. P., Cabal-Rosel, A., Daza-Prieto, B., Springer, B., Cruz-Córdova, A., Leiva-Caro, J., Forsythe, S., & Ruppitsch, W. (2023). Characterization of Cronobacter sakazakii and Cronobacter malonaticus Strains Isolated from Powdered Dairy Products Intended for Consumption by Adults and Older Adults. Microorganisms, 11(12), 2841. https://doi.org/10.3390/microorganisms11122841