
Influence of the Antibiotic Oxytetracycline on the Morphometric Characteristics and Endophytic Bacterial Community of Lettuce (Lactuca sativa L.)
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Hydroponic Experiment Design
2.2. Soil Experiment Design
2.3. DNA Extraction, qPCR, and Sequencing
2.4. Statistical Analysis
3. Results and Discussion
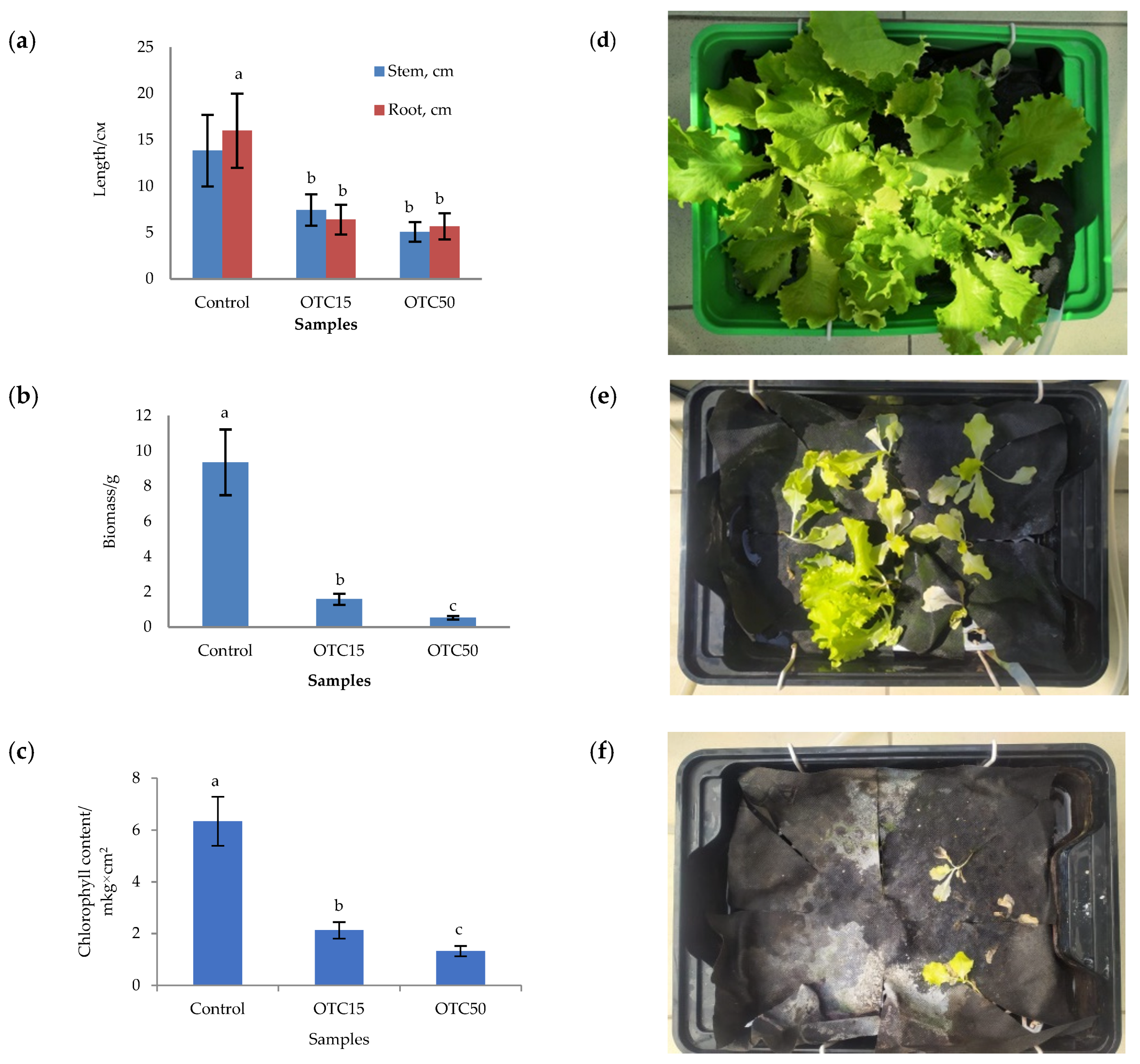
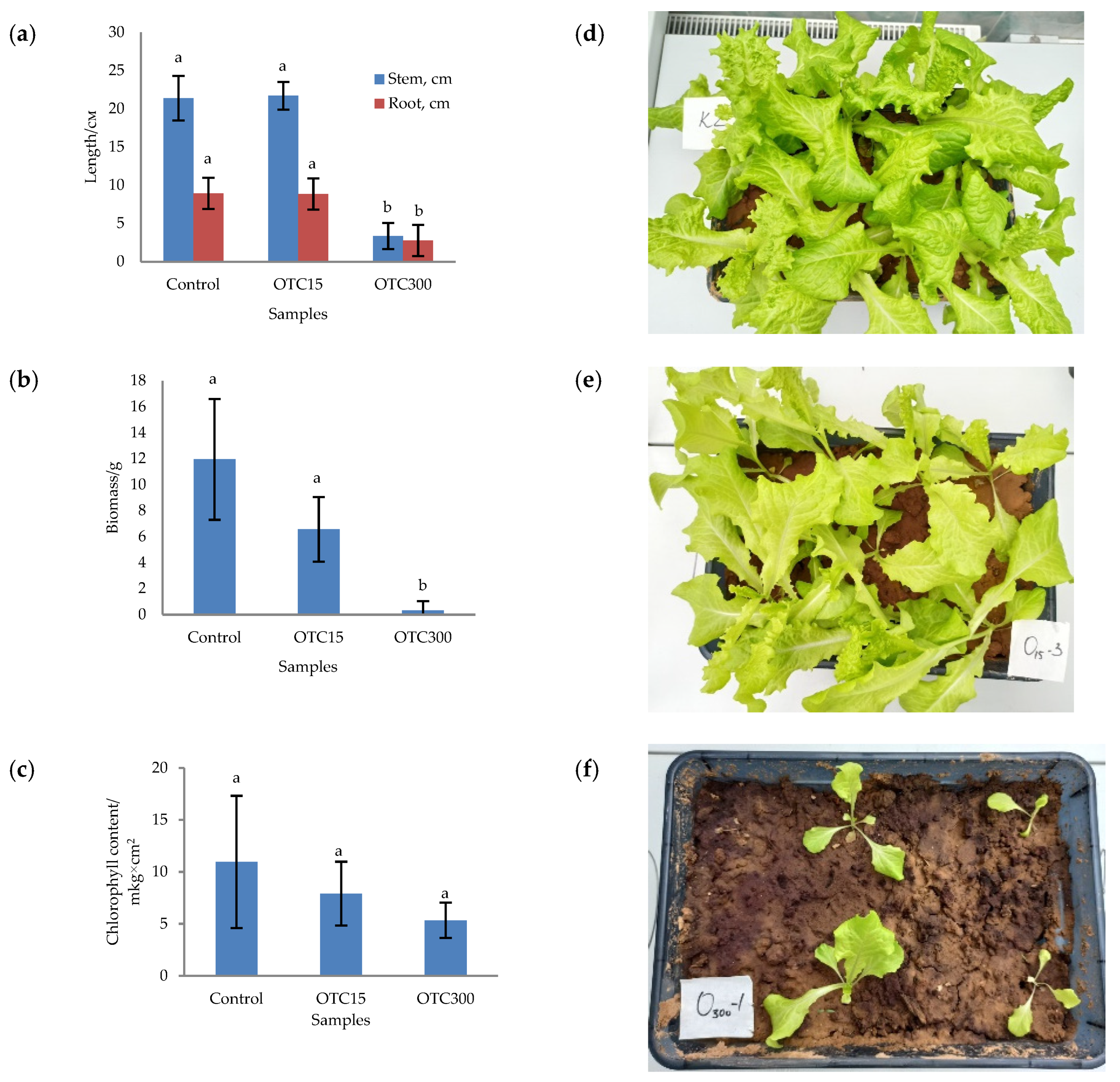
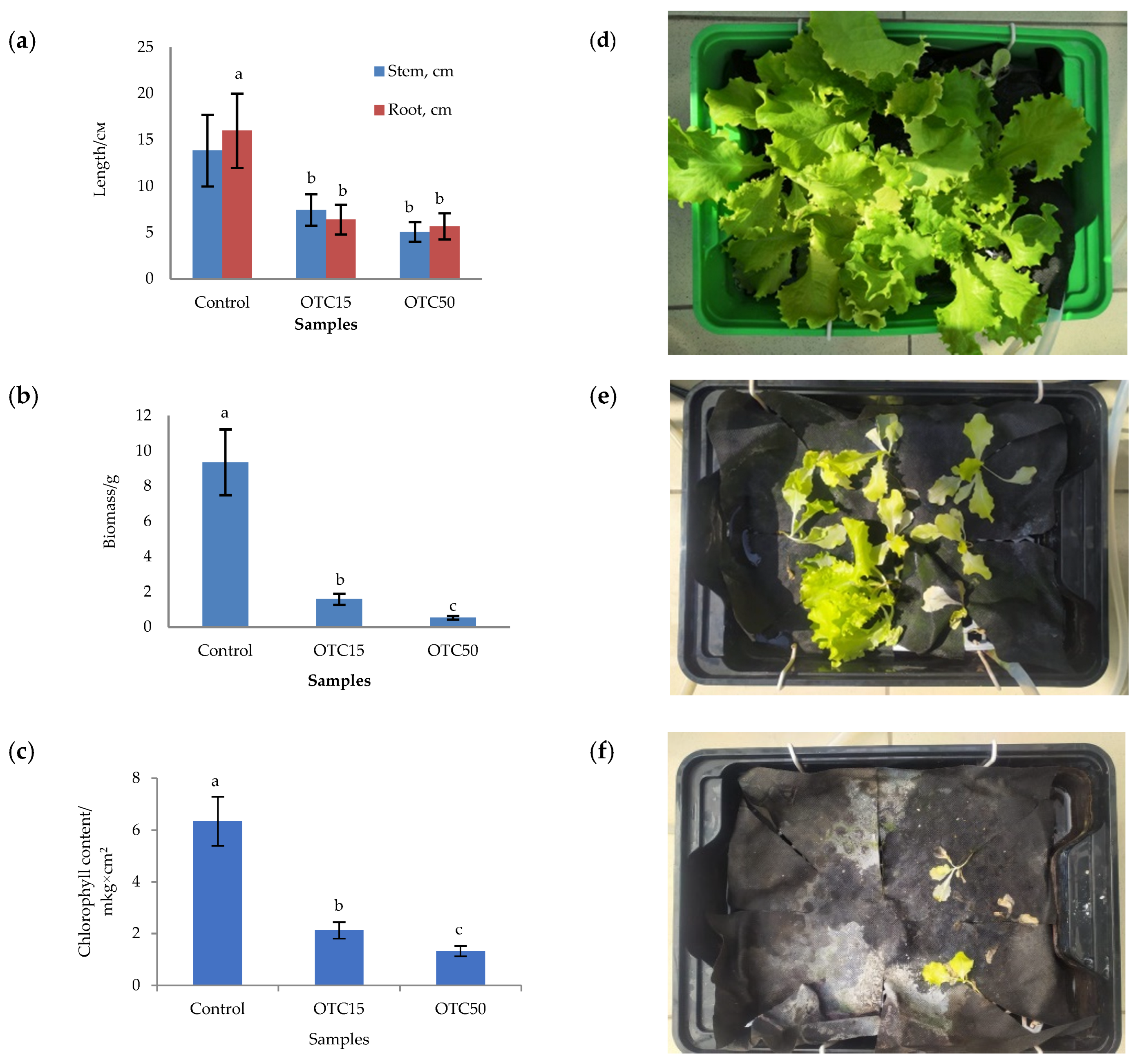
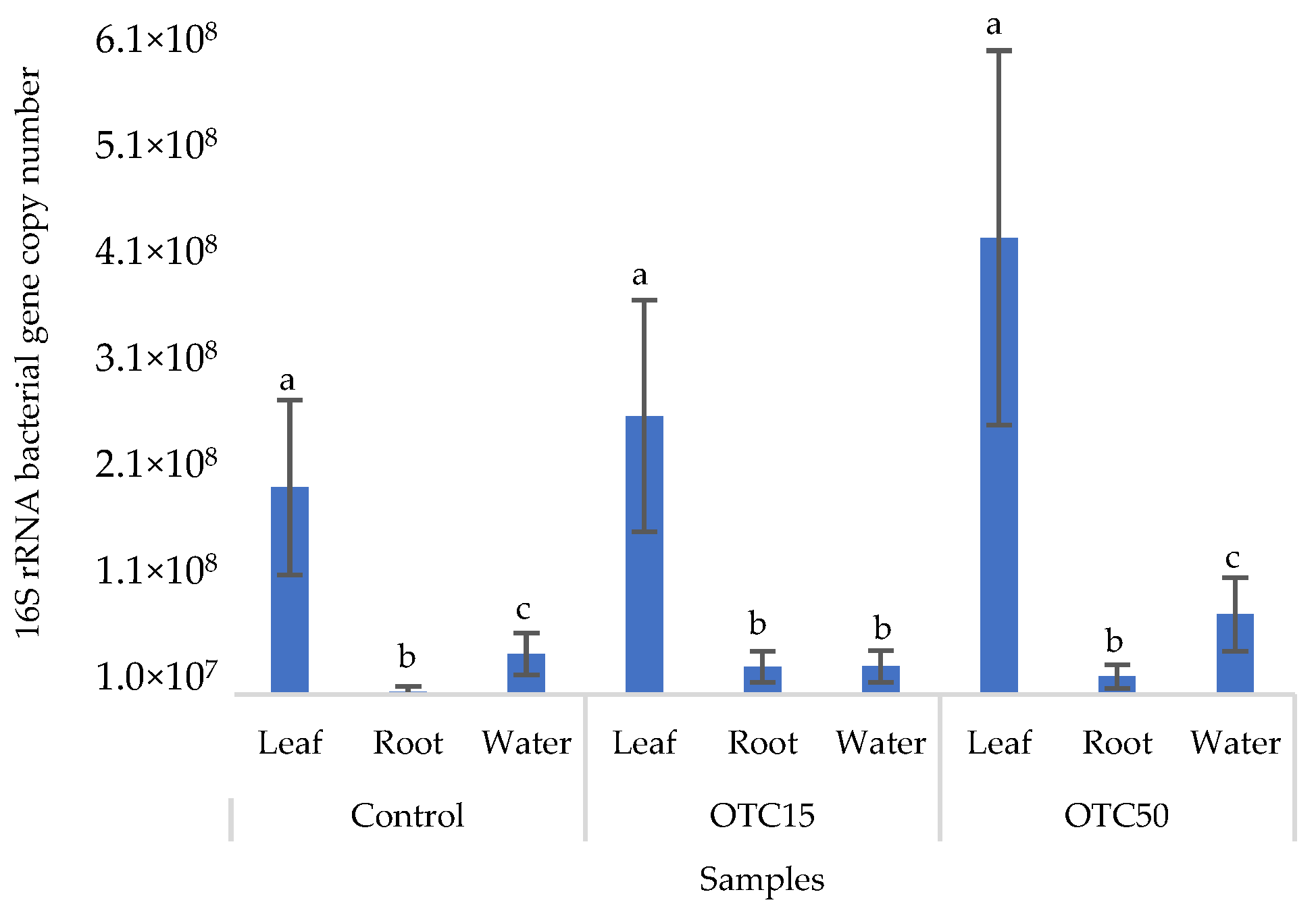
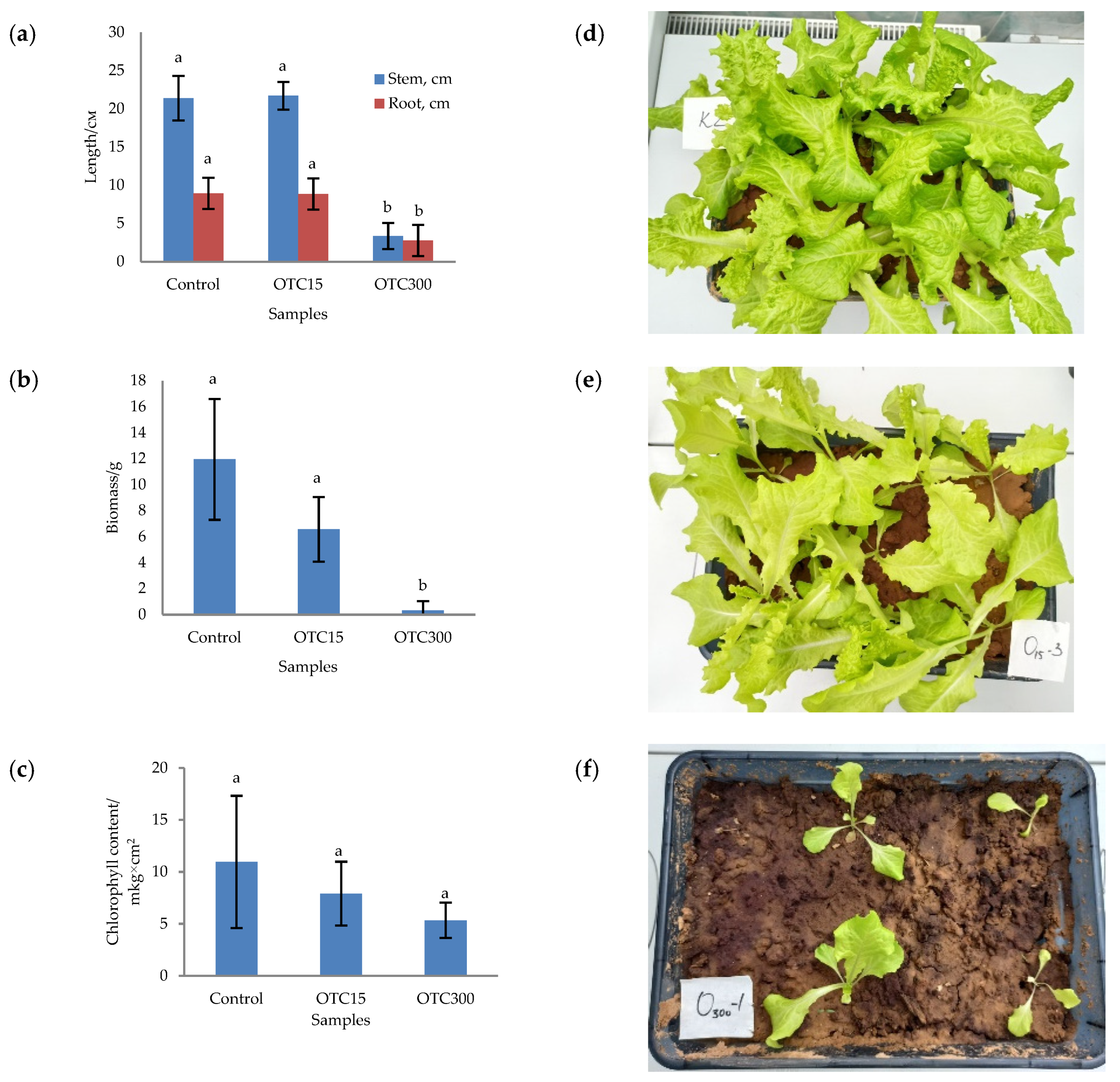
3.1. Hydroponic Experiment
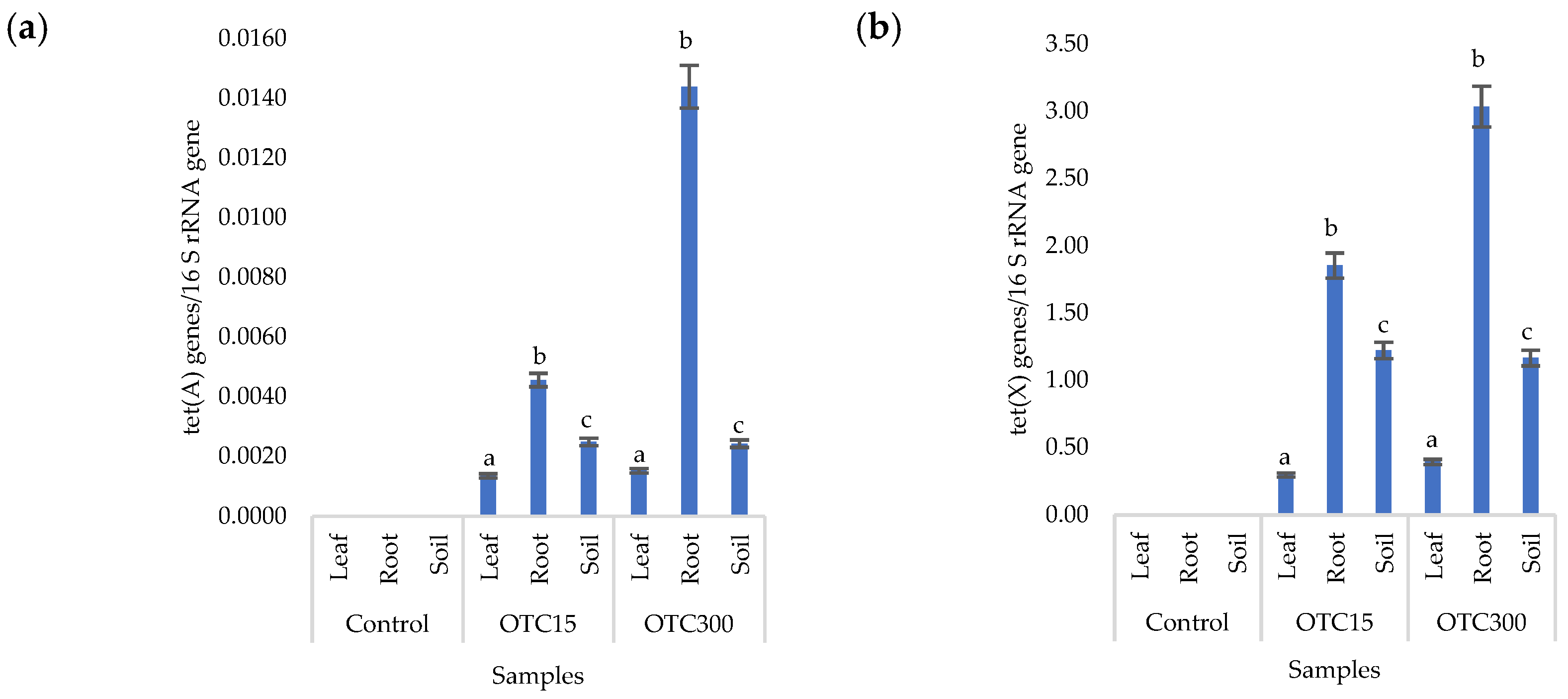
3.2. Soil Experiment
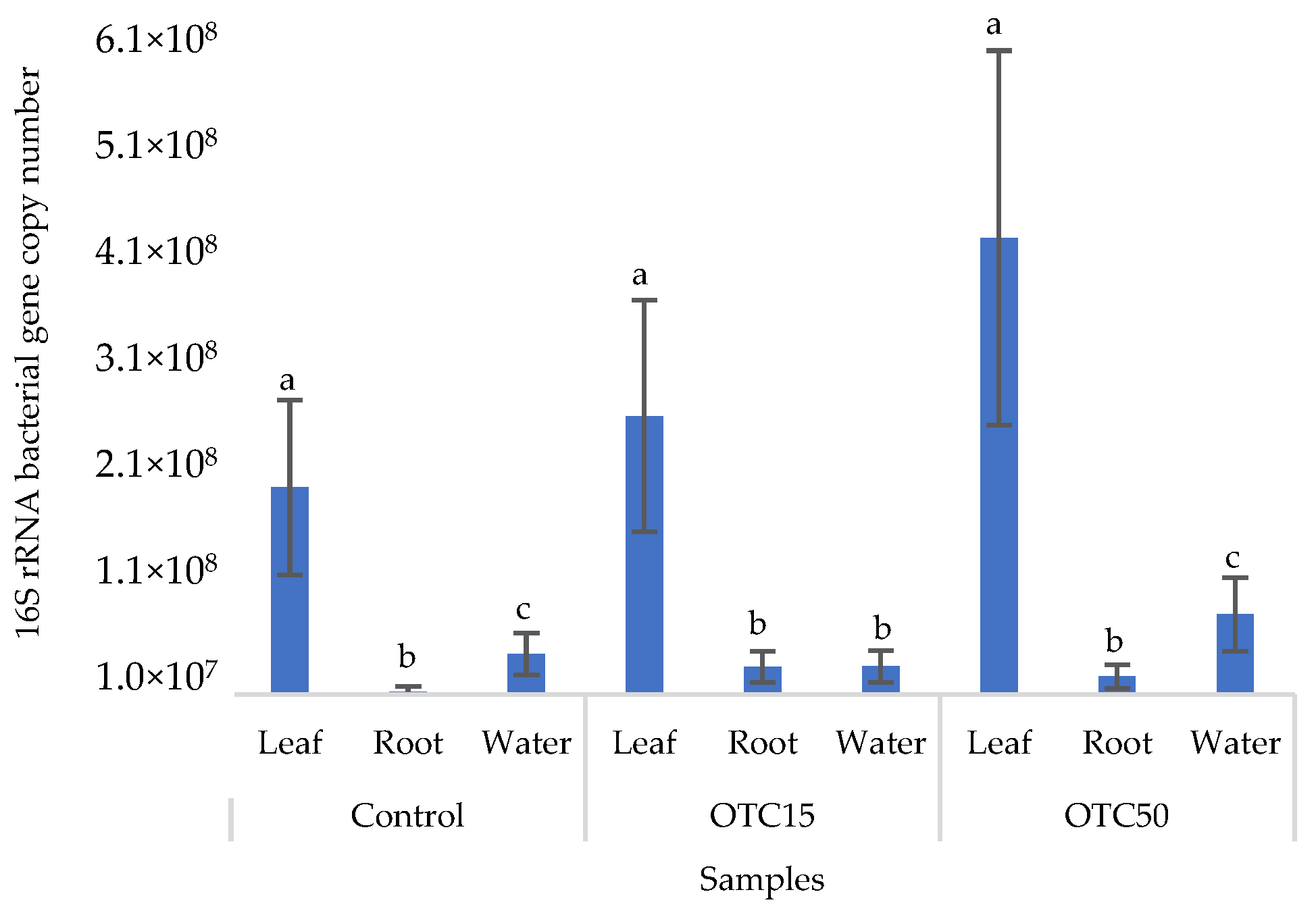
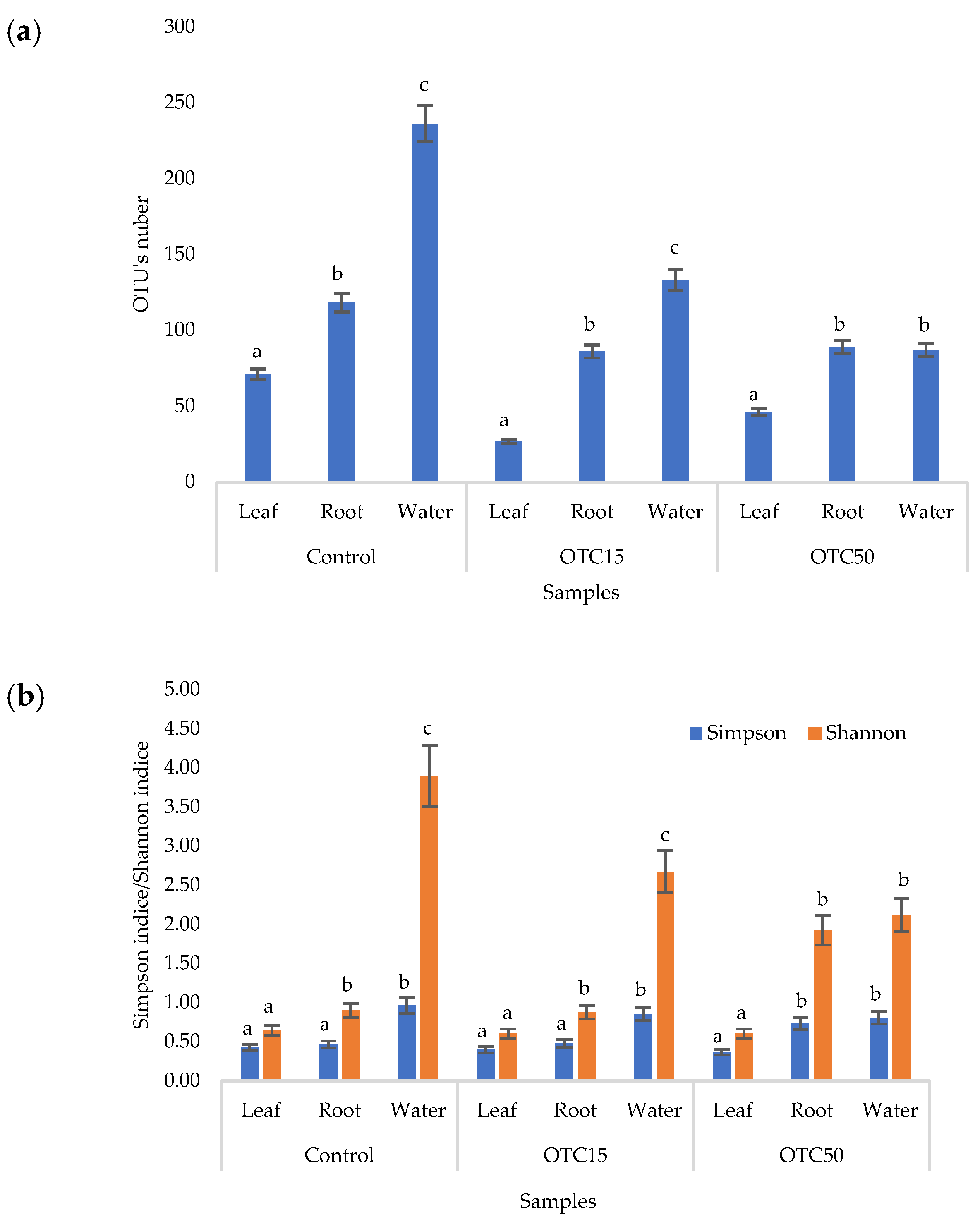
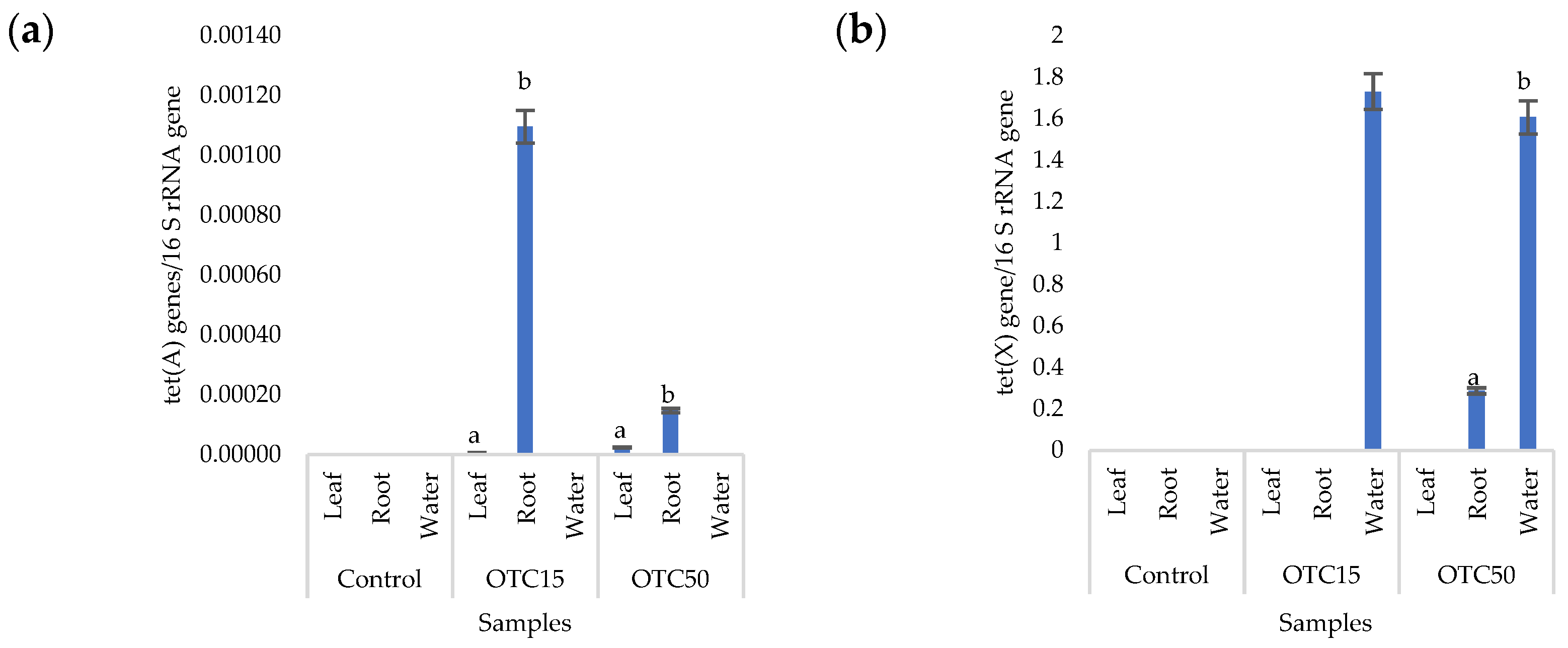
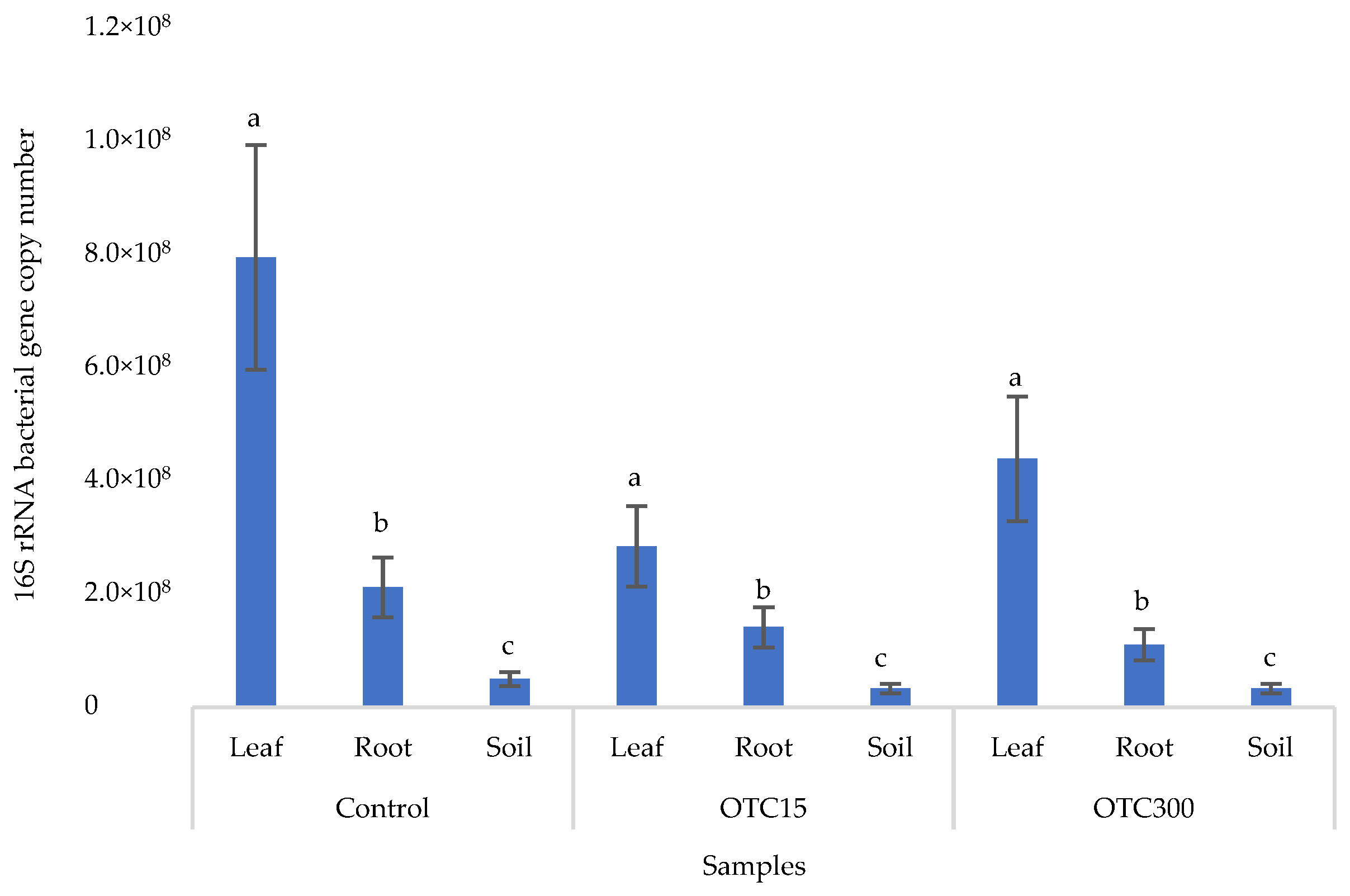
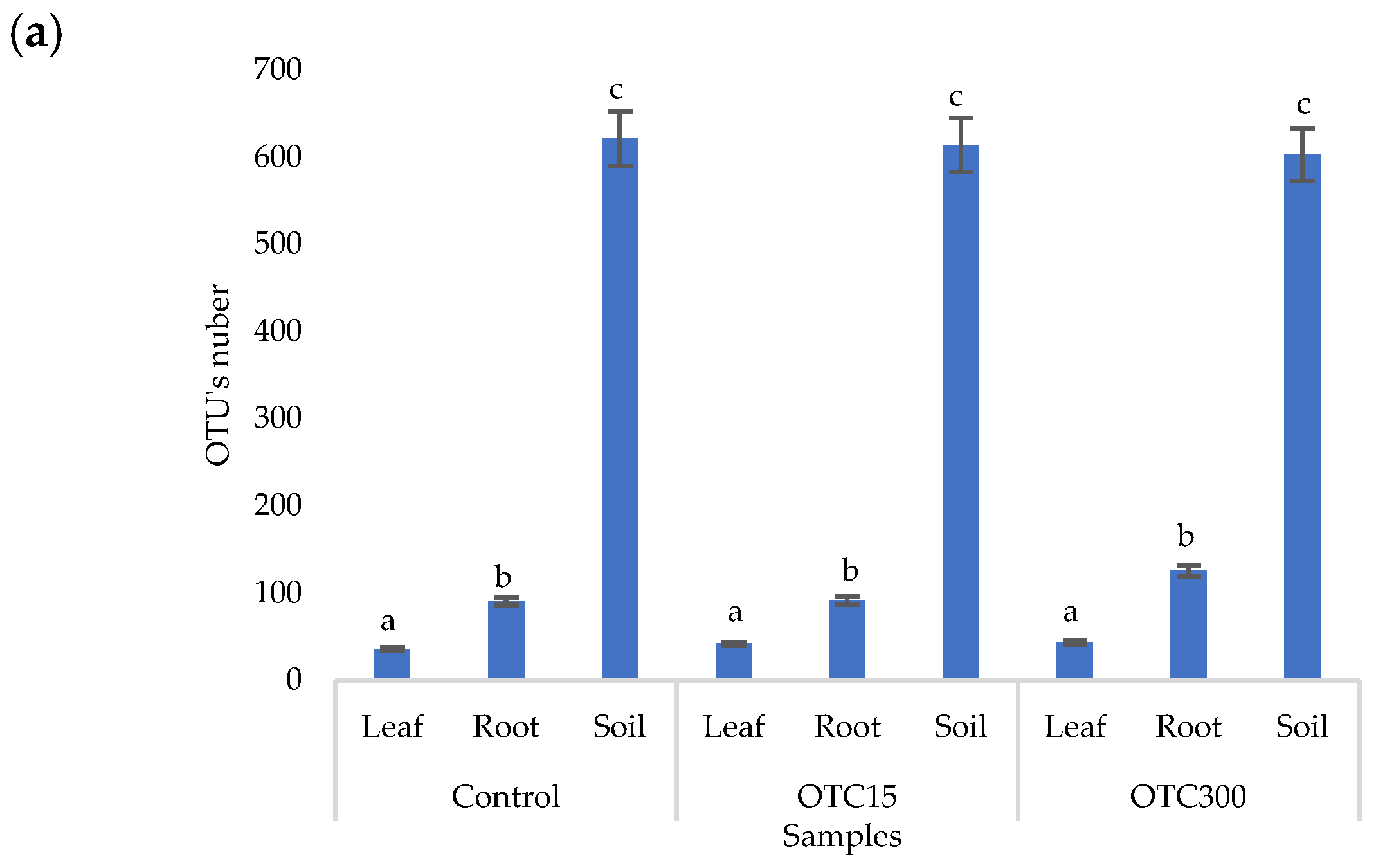
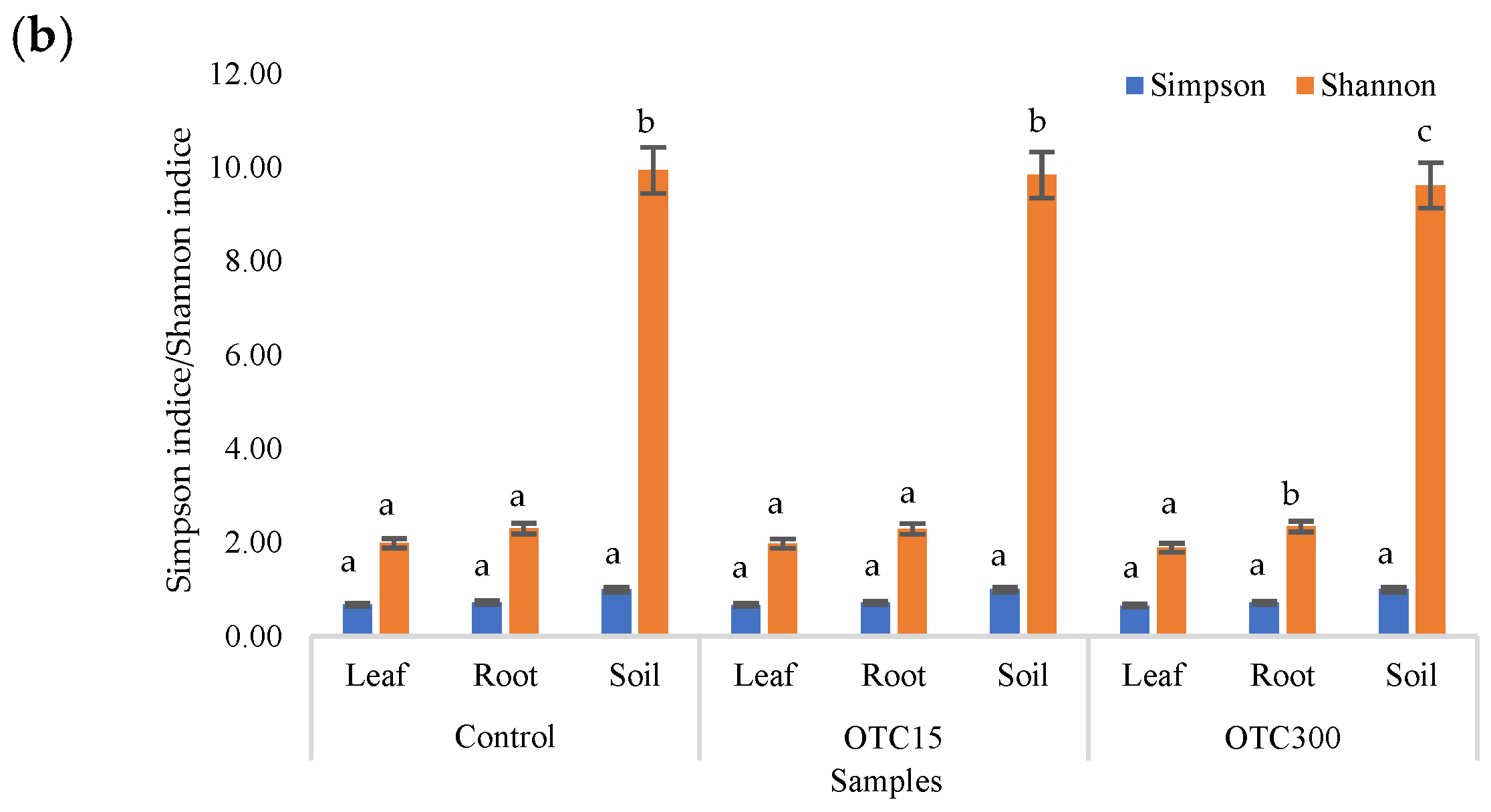
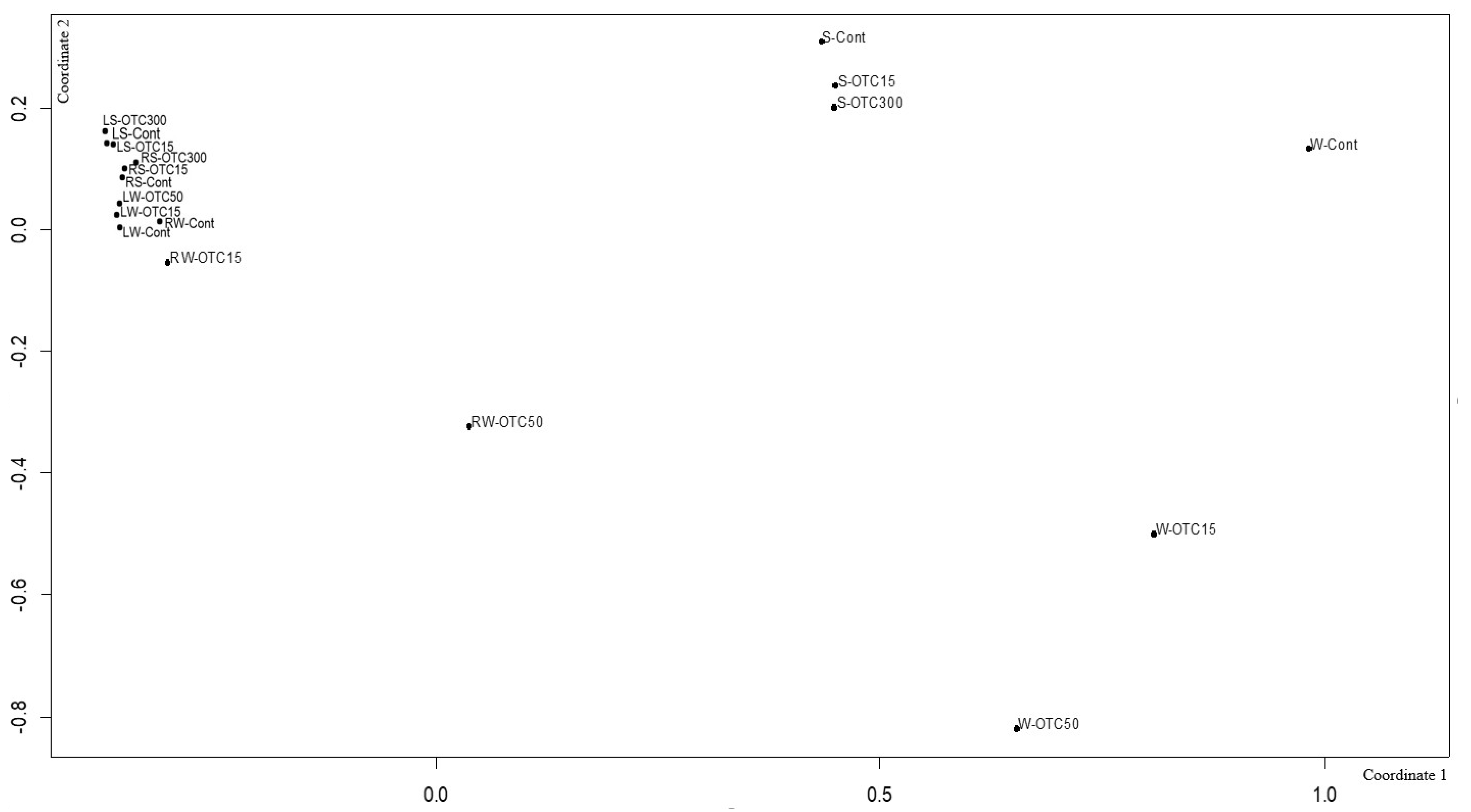
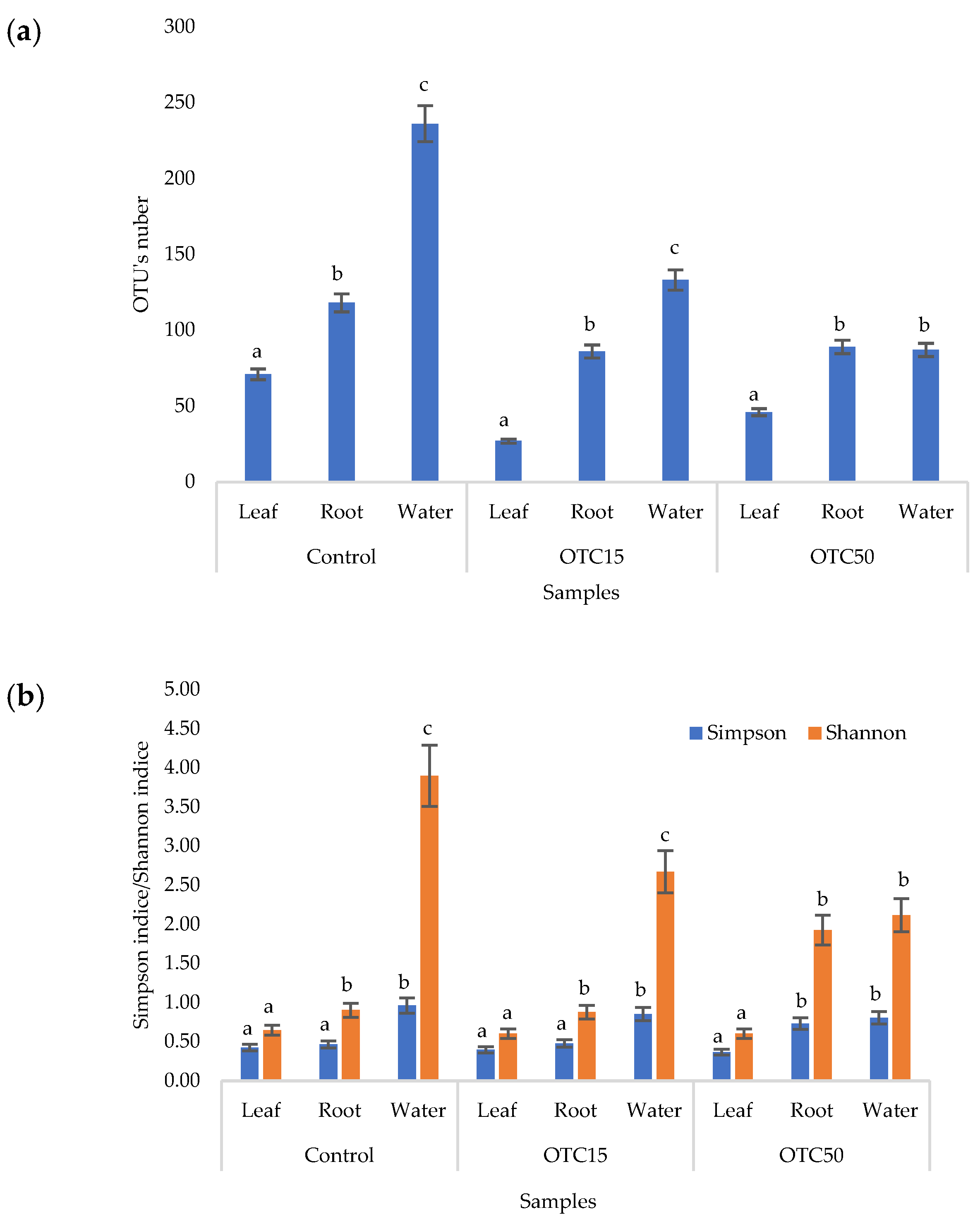
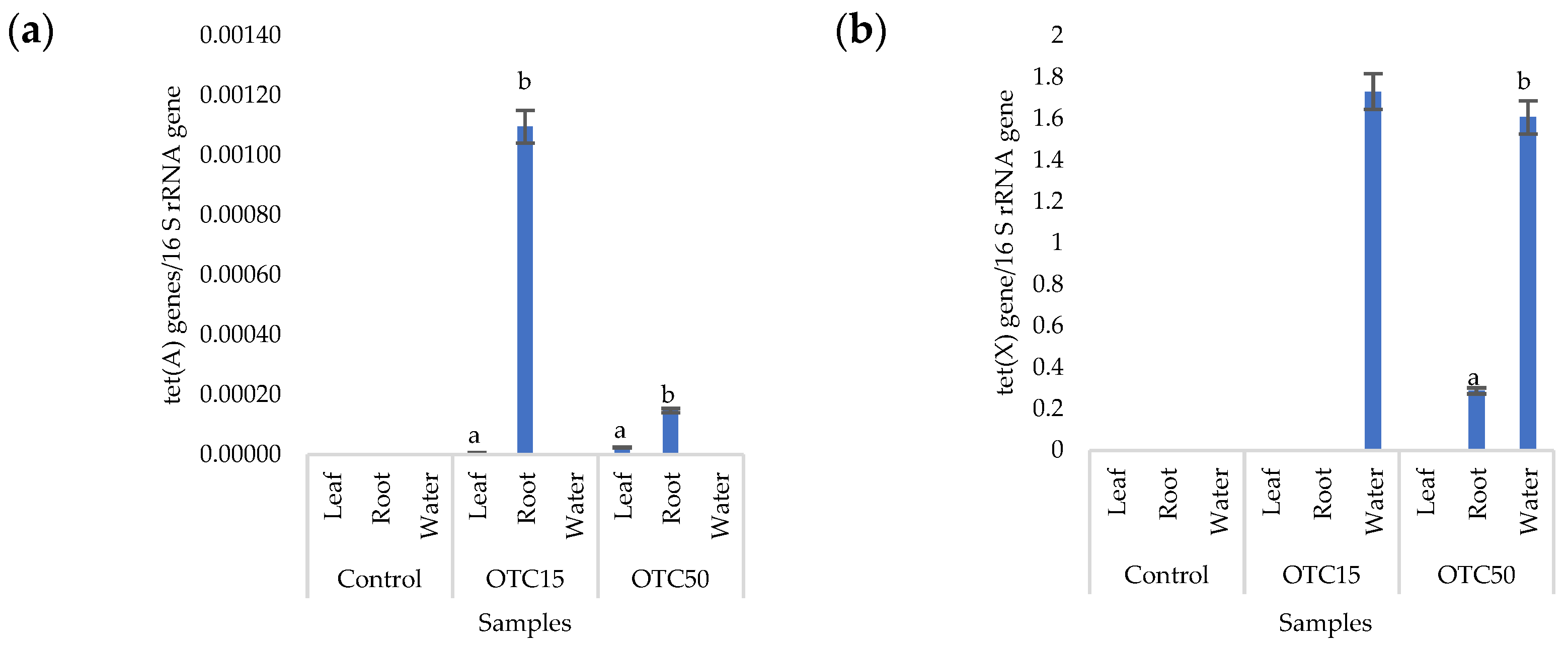
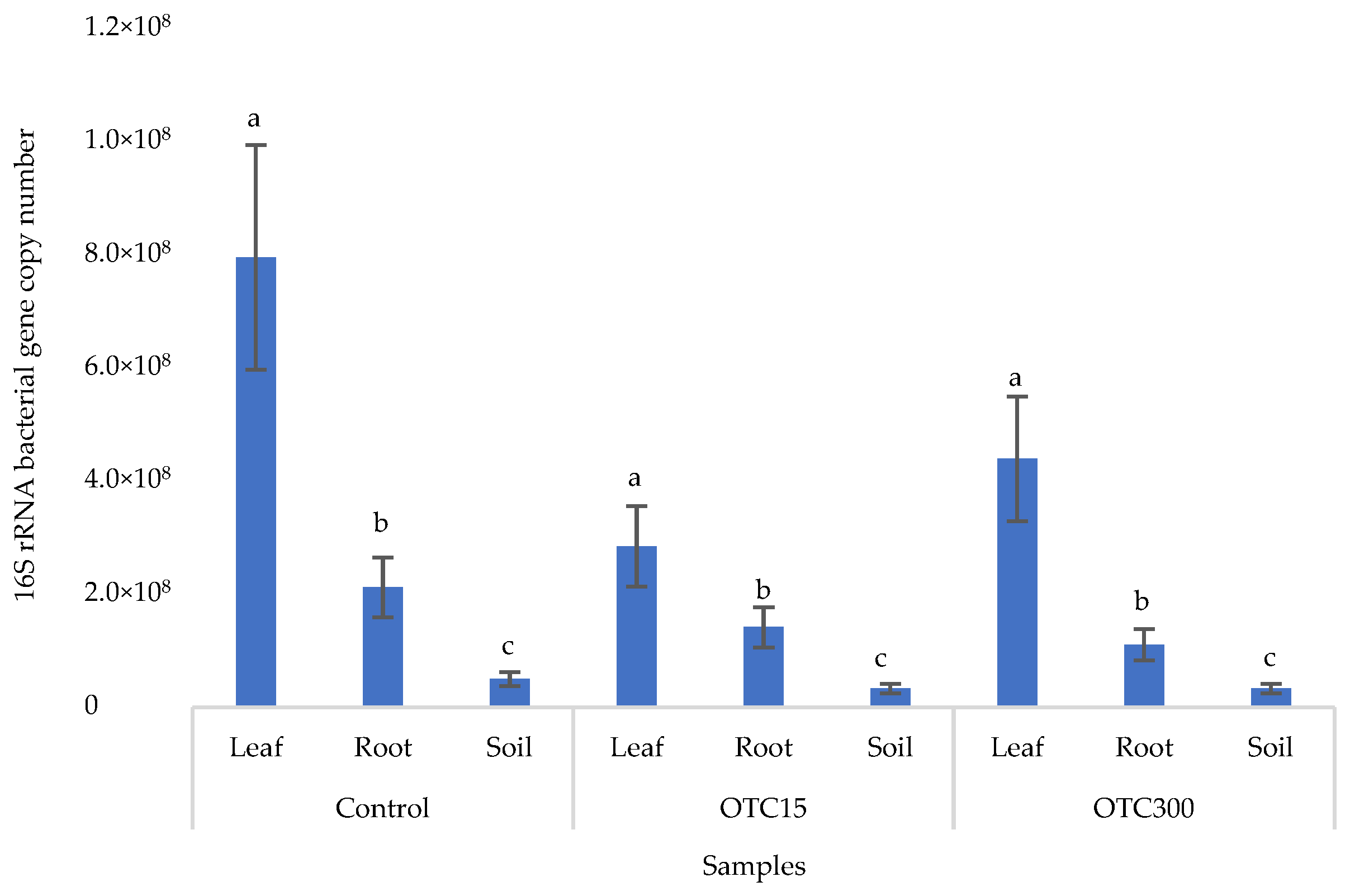
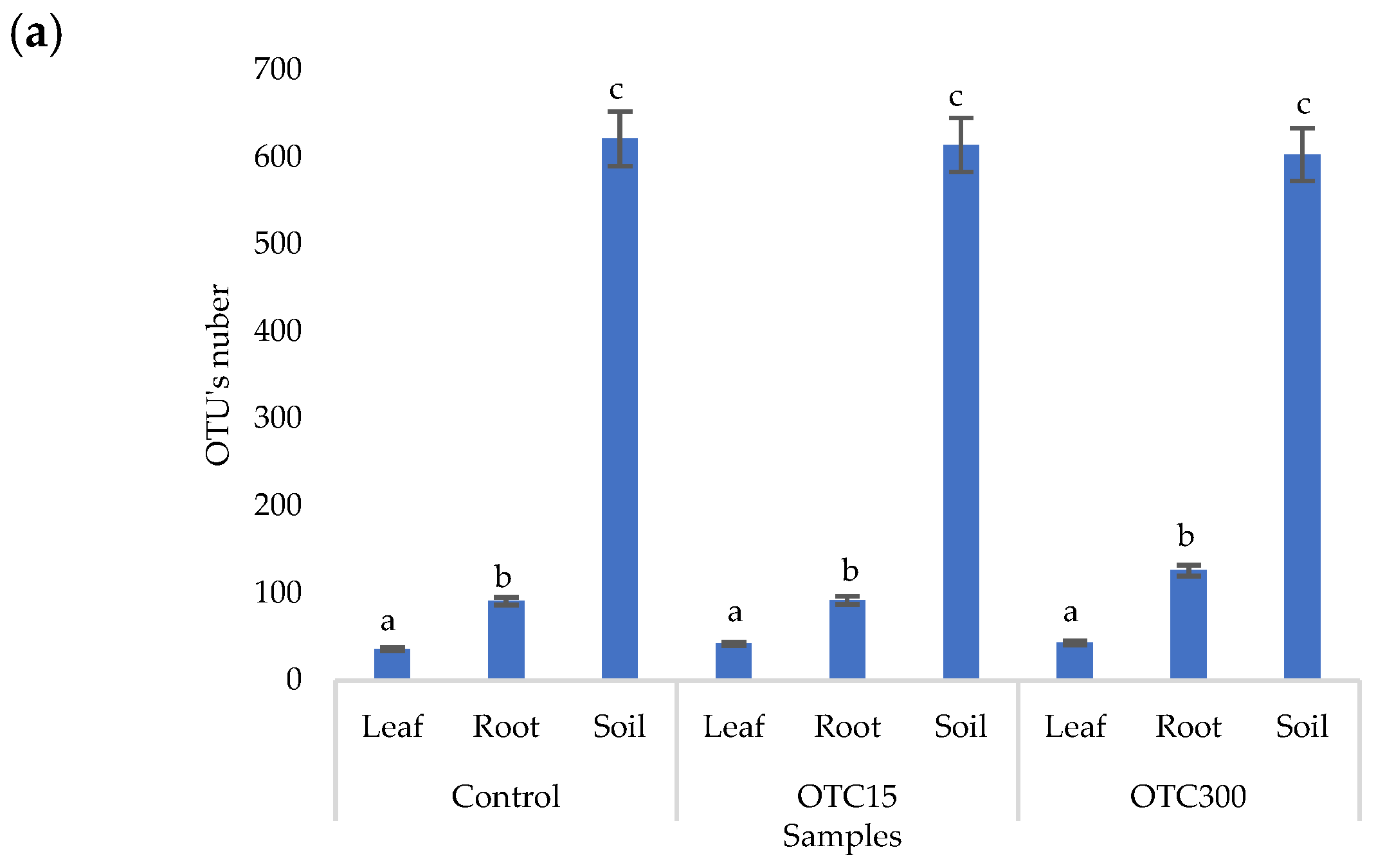
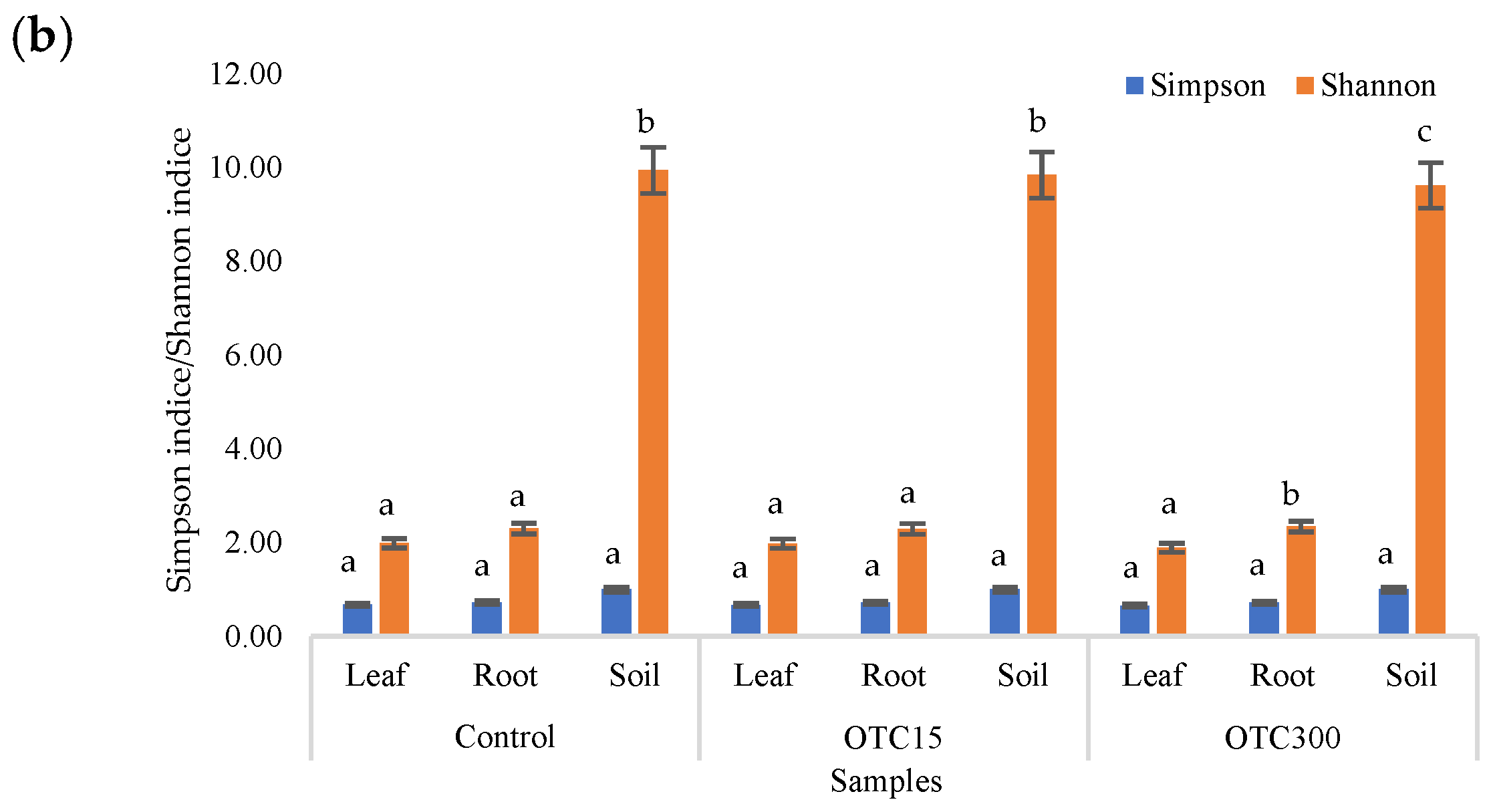
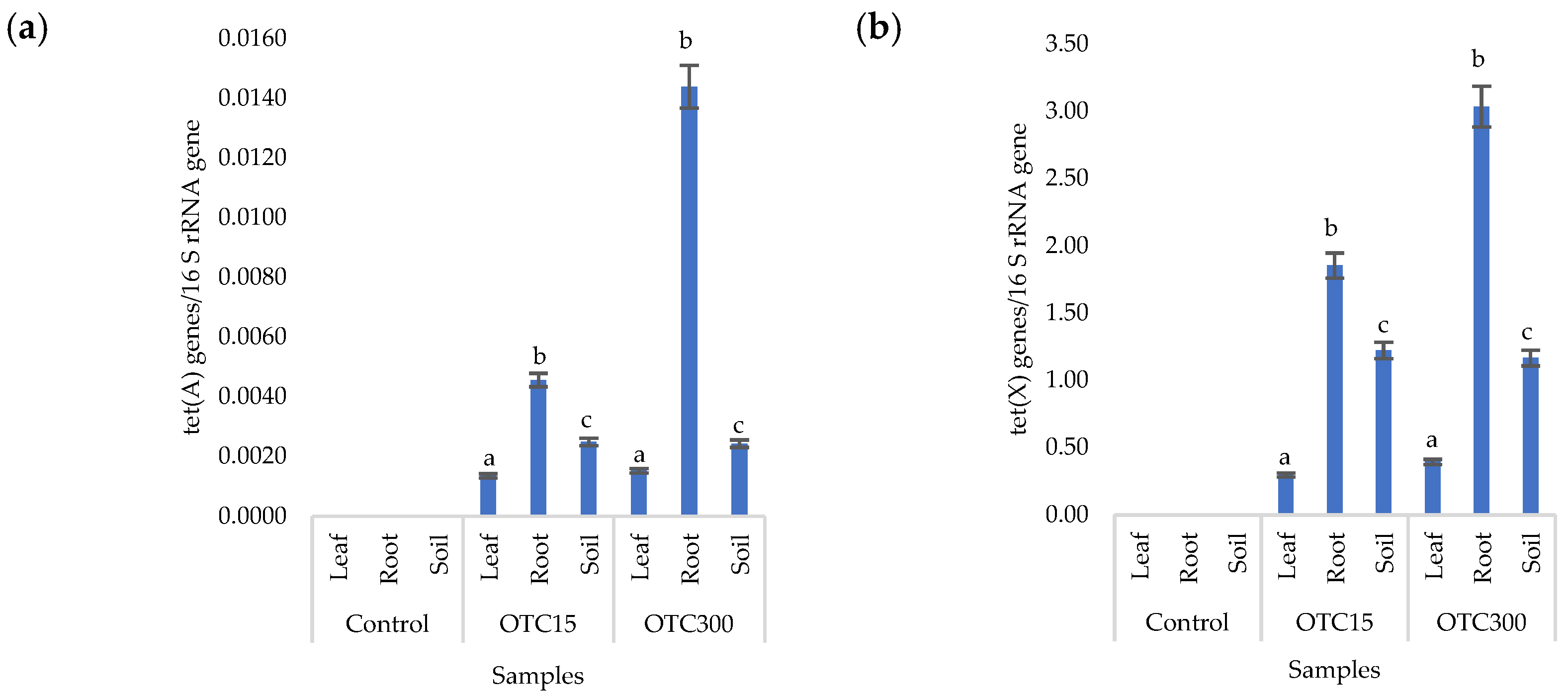
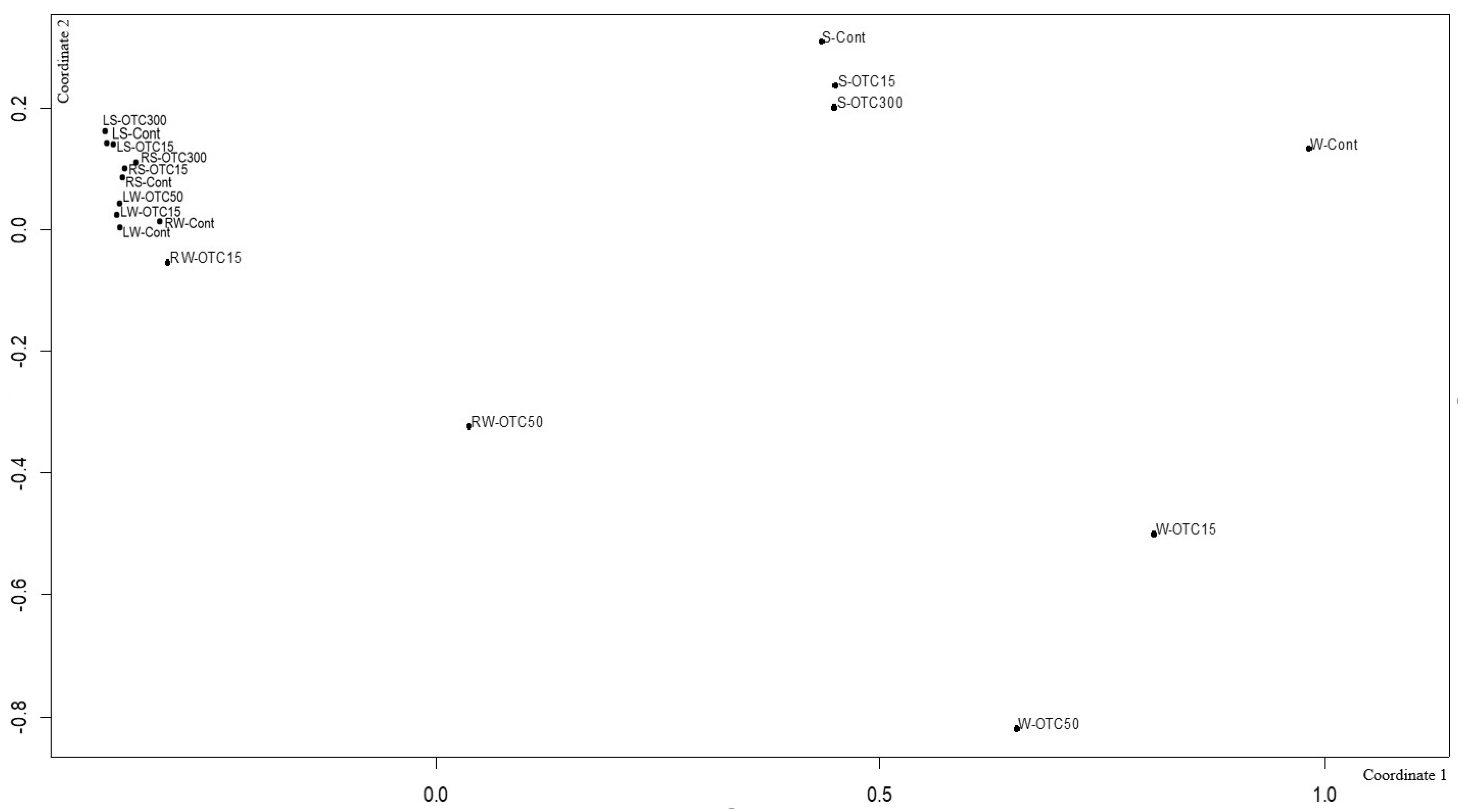
3.3. Bacterial Community Analysis
Endospheric Bacterial Communities of Lettuce Plants
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, X.; Wang, J.; Zhu, L.; Wang, J. Field-based evidence for enrichment of antibiotic resistance genes and mobile genetic elements in manure-amended vegetable soils. Sci. Total Environ. 2019, 654, 906–913. [Google Scholar] [CrossRef]
- Duan, M.; Li, H.; Gu, J.; Tuo, X.; Sun, W.; Qian, X.; Wang, X. Effects of biochar on reducing the abundance of oxytetracycline, antibiotic resistance genes, and human pathogenic bacteria in soil and lettuce. Environ. Pollut. 2017, 224, 787–795. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Hu, H.-W.; Chen, Q.-L.; Singh, B.K.; Yan, H.; Chen, D.; He, J.-Z. Transfer of antibiotic resistance from manure-amended soils to vegetable microbiomes. Environ. Int. 2019, 130, 104912. [Google Scholar] [CrossRef]
- Li, H.; Zheng, X.; Tan, L.; Shao, Z.; Cao, H.; Xu, Y. The vertical migration of antibiotic-resistant genes and pathogens in soil and vegetables after the application of different fertilizers. Environ. Res. 2022, 203, 111884. [Google Scholar] [CrossRef]
- Jechalke, S.; Heuer, H.; Siemens, J.; Amelung, W.; Smalla, K. Fate and effects of veterinary antibiotics in soil. Trends Microbiol. 2014, 22, 536–545. [Google Scholar] [CrossRef]
- Heuer, H.; Focks, A.; Lamshöft, M.; Smalla, K.; Matthies, M.; Spiteller, M. Fate of sulfadiazine administered to pigs and its quantitative effect on the dynamics of bacterial resistance genes in manure and manured soil. Soil Biol. Biochem. 2008, 40, 1892–1900. [Google Scholar] [CrossRef]
- Halling-Sørensen, B.; Nielsen, S.N.; Lanzky, P.; Ingerslev, F.; Lützhøft, H.H.; Jørgensen, S. Occurrence, fate and effects of pharmaceutical substances in the environment—A review. Chemosphere 1998, 36, 357–393. [Google Scholar] [CrossRef]
- Granados-Chinchilla, F.; Rodríguez, C. Tetracyclines in Food and Feedingstuffs: From Regulation to Analytical Methods, Bacterial Resistance, and Environmental and Health Implications. J. Anal. Methods Chem. 2017, 2017, 1–24. [Google Scholar] [CrossRef]
- Mulchandani, R.; Wang, Y.; Gilbert, M.; Van Boeckel, T.P. Global trends in antimicrobial use in food-producing animals: 2020 to 2030. PLoS Glob. Public Health 2023, 3, e0001305. [Google Scholar] [CrossRef]
- Zhou, Y.; Niu, L.; Zhu, S.; Lu, H.; Liu, W. Occurrence, abundance, and distribution of sulfonamide and tetracycline resistance genes in agricultural soils across China. Sci. Total Environ. 2017, 599–600, 1977–1983. [Google Scholar] [CrossRef]
- Smalla, K.; Wieland, G.; Buchner, A.; Zock, A.; Parzy, J.; Kaiser, S.; Roskot, N.; Heuer, H.; Berg, G. Bulk and Rhizosphere Soil Bacterial Communities Studied by Denaturing Gradient Gel Electrophoresis: Plant-Dependent Enrichment and Seasonal Shifts Revealed. Appl. Environ. Microbiol. 2001, 67, 4742–4751. [Google Scholar] [CrossRef]
- Schreiter, S.; Ding, G.-C.; Heuer, H.; Neumann, G.; Sandmann, M.; Grosch, R.; Kropf, S.; Smalla, K. Effect of the soil type on the microbiome in the rhizosphere of field-grown lettuce. Front. Microbiol. 2014, 5, 144. [Google Scholar] [CrossRef]
- Tien, Y.C.; Li, B.; Zhang, T.; Scott, A.; Murray, R.; Sabourin, L.; Topp, E. Impact of dairy manure pre-application treatment on manure composition, soil dynamics of antibiotic resistance genes, and abundance of antibiotic-resistance genes on vegetables at harvest. Sci. Total Environ. 2017, 581–582, 32–39. [Google Scholar] [CrossRef]
- Brandt, K.K.; Sjøholm, O.R.; Krogh, K.A.; Halling-Sørensen, B.; Nybroe, O. Increased Pollution-Induced Bacterial Community Tolerance to Sulfadiazine in Soil Hotspots Amended with Artificial Root Exudates. Environ. Sci. Technol. 2009, 43, 2963–2968. [Google Scholar] [CrossRef]
- Heuer, H.; Smalla, K. Plasmids foster diversification and adaptation of bacterial populations in soil. FEMS Microbiol. Rev. 2012, 36, 1083–1104. [Google Scholar] [CrossRef]
- Minden, V.; Deloy, A.; Volkert, A.M.; Leonhardt, S.D.; Pufal, G. Antibiotics impact plant traits, even at small concentrations. AoB Plants 2017, 9, plx010. [Google Scholar] [CrossRef]
- Migliore, L.; Rotini, A.; Cerioli, N.L.; Cozzolino, S.; Fiori, M. Phytotoxic Antibiotic Sulfadimethoxine Elicits a Complex Hormetic Response in the Weed Lythrum Salicaria L. Dose-Response 2010, 8, 414–427. [Google Scholar] [CrossRef]
- Michelini, L.; Reichel, R.; Werner, W.; Ghisi, R.; Thiele-Bruhn, S. Sulfadiazine Uptake and Effects on Salix fragilis L. and Zea mays L. Plants. Water Air Soil Pollut. 2012, 223, 5243–5257. [Google Scholar] [CrossRef]
- Piotrowicz-Cieślak, A.I.; Adomas, B.; Nałęcz-Jawecki, G.; Michalczyk, D.J. Phytotoxicity of Sulfamethazine Soil Pollutant to Six Legume Plant Species. J. Toxicol. Environ. Health Part A 2010, 73, 1220–1229. [Google Scholar] [CrossRef]
- Liu, F.; Ying, G.G.; Tao, R.; Zhao, J.L.; Yang, J.F.; Zhao, L.F. Effects of six selected antibiotics on plant growth and soil microbial and enzymatic activities. Environ. Pollut. 2009, 157, 1636–1642. [Google Scholar] [CrossRef]
- Zhao-Jun, L.I.; Xiao-Yu, X.I.E.; Zhang, S.Q.; Liang, Y.C. Wheat growth and photosynthesis as affected by oxytetracycline as a soil contaminant. Pedosphere 2011, 21, 244–250. [Google Scholar]
- Puccinelli, M.; Landi, M.; Maggini, R.; Pardossi, A.; Incrocci, L. Iodine biofortification of sweet basil and lettuce grown in two hydroponic systems. Sci. Hortic. 2020, 276, 109783. [Google Scholar] [CrossRef]
- de Boer, M.; Heuer, C.; Hussein, H.; McDougall, S. Minimum inhibitory concentrations of selected antimicrobials against Escherichia coli and Trueperella pyogenes of bovine uterine origin. J. Dairy Sci. 2015, 98, 4427–4438. [Google Scholar] [CrossRef]
- ISO 17601:2016; Soil Quality—Estimation of Abundance of Selected Microbial Gene Sequences by Quantitative PCR from DNA Directly Extracted from Soil. International Organization for Standard: Geneva, Switzerland, 2016.
- Zhang, W.; Huang, M.-H.; Qi, F.-F.; Sun, P.-Z.; Van Ginkel, S.W. Effect of trace tetracycline concentrations on the structure of a microbial community and the development of tetracycline resistance genes in sequencing batch reactors. Bioresour. Technol. 2013, 150, 9–14. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Thiele-Bruhn, S. Pharmaceutical antibiotic compounds in soils—A review. J. Plant Nutr. Soil Sci. 2003, 166, 145–167. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, J.; Zhang, W.; Wang, Z.; Xie, Y.; Zhang, H. Influence of tetracycline exposure on the growth of wheat seedlings and the rhizosphere microbial community structure in hydroponic culture. J. Environ. Sci. Health 2010, 45, 190–197. [Google Scholar] [CrossRef]
- Bradel, B.G.; Preil, W.; Jeske, H. Remission of the Free-branching Pattern of Euphorbia pulcherrima by Tetracycline Treatment. J. Phytopathol. 2000, 148, 587–590. [Google Scholar]
- Michelini, L.; La Rocca, N.; Rascio, N.; Ghisi, R. Structural and functional alterations induced by two sulfonamide antibiotics on barley plants. Plant Physiol. Biochem. 2013, 67, 55–62. [Google Scholar] [CrossRef]
- Qiu, J.; Chen, Y.; Feng, Y.; Li, X.; Xu, J.; Jiang, J. Adaptation of Rhizosphere Microbial Communities to Continuous Exposure to Multiple Residual Antibiotics in Vegetable Farms. Int. J. Environ. Res. 2023, 20, 3137. [Google Scholar] [CrossRef]
- Sarmah, A.K.; Meyer, M.T.; Boxall, A.B. A global perspective on the use, sales, exposure pathways, occurrence, fate and effects of veterinary antibiotics (VAs) in the environment. Chemosphere 2006, 65, 725–759. [Google Scholar] [CrossRef]
- Feranchuk, S.; Belkova, N.; Potapova, U.; Kuzmin, D.; Belikov, S. Evaluating the use of diversity indices to distinguish between microbial communities with different traits. Res. Microbiol. 2018, 169, 254–261. [Google Scholar] [CrossRef]
- Galieva, G.; Galitskaya, P.; Selivanovskaya, S. Plant Microbiome: Origin, Composition, and Functions. Uchenye Zap. Kazan. Univ. Seriya Estestv. Nauk. 2023, 165, 231–262. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, W.; Yang, L.; Stedtfeld, R.D.; Peng, A.; Gu, C.; Li, H. Antibiotic resistance genes and bacterial communities in cornfield and pasture soils receiving swine and dairy manures. Environ. Pollut. 2019, 248, 947–957. [Google Scholar] [CrossRef]
- Hund-Rinke, K.; Simon, M.; Lukow, T. Effects of tetracycline on the soil microflora: Function, diversity, resistance. J. Soils Sediments 2004, 4, 11–16. [Google Scholar] [CrossRef]
- Al-Rimawi, F.; Hijaz, F.; Nehela, Y.; Batuman, O.; Killiny, N. Uptake, Translocation, and Stability of Oxytetracycline and Streptomycin in Citrus Plants. Antibiotics 2019, 8, 196. [Google Scholar] [CrossRef]
- Boto, R.E.F.; Almeida, P.; Queiroz, J.A. Degradation patterns of tetracycline antibiotics in reverse micelles and water. Biomed. Chromatogr. 2006, 288, 278–288. [Google Scholar]
- Xuan, R.; Arisi, L.; Wang, Q.; Yates, S.R.; Biswas, K.C. Hydrolysis and photolysis of oxytetracycline in aqueous solution. J. Environ. Sci. Health 2010, 45, 73–81. [Google Scholar] [CrossRef]
- Møller, T.S.; Overgaard, M.; Nielsen, S.S.; Bortolaia, V.; Sommer, M.O.; Guardabassi, L.; Olsen, J.E. Relation between tetR and tetA expression in tetracycline resistant Escherichia coli. BMC Microbiol. 2016, 16, 39. [Google Scholar] [CrossRef]
- Wang, T.; Sun, S.; Xu, Y.; Waigi, M.G.; Odinga, E.S.; Vasilyeva, G.K.; Hu, X. Nitrogen Regulates the Distribution of Antibiotic Resistance Genes in the Soil–Vegetable System. Front. Microbiol. 2022, 13, 848750. [Google Scholar] [CrossRef]
- Mandic-Mulec, I.; Stefanic, P.; van Elsas, J.D. Ecology of bacillaceae. Microbiol. Spect. 2016, 3, 59–85. [Google Scholar]
- Guo, P.; Zhang, K.; Ma, X.; He, P. Clostridium species as probiotics: Potentials and challenges. J. Anim. Sci. Biotechnol. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Ryan, M.P.; Tony Pembroke, J. The genus ochrobactrum as major opportunistic pathogens. Microorganisms 2020, 8, 1797. [Google Scholar] [CrossRef]
- Young, J.P.W.; Moeskjær, S.; Afonin, A.; Rahi, P.; Maluk, M.; James, E.K.; Perry, B.J. Defining the Rhizobium leguminosarum Species Complex. Genes 2021, 12, 111. [Google Scholar] [CrossRef]
- Wang, Z.; Hu, H.; Zhu, T.; Zheng, J.; Gänzle, M.G.; Simpson, D.J. Ecology and Function of the Transmissible Locus of Stress Tolerance in Escherichia coli and Plant-Associated Enterobacteriaceae. mSystems 2021, 6, e00378-21. [Google Scholar] [CrossRef]
- Dworkin, M.; Falkow, S.; Rosenberg, E.; Schleifer, K.; Stackebrandt, E. The Prokaryotes, 3rd ed.; Springer: New York, NY, USA, 2014; pp. 321–333. [Google Scholar]
- Linke, D.; Goldman, A. Bacterial adhesion. Adv. Exp. Med. Biol. 2011, 715, 271–284. [Google Scholar]
- Kim, J.S.; Kang, S.W.; Lee, J.H.; Park, S.H.; Lee, J.S. The evolution and competitive strategies of Akkermansia muciniphila in gut. Gut Microbes 2022, 14, 2025017. [Google Scholar] [CrossRef]
- Besaury, L.; Floret, J.; Rémond, C. Sphingobacterium prati sp. nov., isolated from agricultural soil and involved in lignocellulose deconstruction. Int. J. Syst. Evol. Microbiol. 2021, 71, 004963. [Google Scholar] [CrossRef]
- Barahona, F.; Slim, J. Sphingobacterium multivorum: Case report and literature review. New Microbes New Infect. 2015, 7, 33–36. [Google Scholar] [CrossRef]
- Rosenberg, E.; DeLong, E.; Lory, S.; Stackebrandt, E.; Thompson, F. The Prokaryotes. In Zootaxa; Springer: Berlin, Germany, 2012; Volume 56, pp. 47–62. [Google Scholar]
- Lovering, A.L.; Sockett, R.E. Microbe Profile: Bdellovibrio bacteriovorus: A specialized bacterial predator of bacteria. Microbiology 2021, 167, 001043. [Google Scholar] [CrossRef]
- Wegst-Uhrich, S.R.; Navarro, D.A.; Zimmerman, L.; Aga, D.S. Assessing antibiotic sorption in soil: A literature review and new case studies on sulfonamides and macrolides. Chem. Central J. 2014, 8, 1–12. [Google Scholar] [CrossRef]
- Kumar, K.; Gupta, S.C.; Baidoo, S.K.; Chander, Y.; Rosen, C.J. Antibiotic Uptake by Plants from Soil Fertilized with Animal Manure. J. Environ. Qual. 2005, 34, 2082–2085. [Google Scholar] [CrossRef]
- Zhang, H.; Li, X.; Yang, Q.; Sun, L.; Yang, X.; Zhou, M.; Deng, R.; Bi, L. Plant Growth, Antibiotic Uptake, and Prevalence of Antibiotic Resistance in an Endophytic System of Pakchoi under Antibiotic Exposure. Int. J. Environ. Res. Public Health 2017, 14, 1336. [Google Scholar] [CrossRef]
- Kim, K.R.; Owens, G.; Kwon, S.I.; So, K.H.; Lee, D.B.; Ok, Y.S. Occurrence and environmental fate of veterinary antibiotics in the terrestrial environment. Water Air Soil Pollut. 2011, 214, 163–174. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Rott, M.; Schlaeppi, K.; van Themaat, E.V.L.; Ahmadinejad, N.; Assenza, F.; Rauf, P.; Huettel, B.; Reinhardt, R.; Schmelzer, E.; et al. Revealing structure and assembly cues for Arabidopsis root-inhabiting bacterial microbiota. Nature 2012, 488, 91–95. [Google Scholar] [CrossRef]
- Franklin, A.M.; Williams, C.; Andrews, D.M.; Watson, J.E. Sorption and desorption behavior of four antibiotics at concentrations simulating wastewater reuse in agricultural and forested soils. Chemosphere 2021, 289, 133038. [Google Scholar] [CrossRef]
- Danilova, N.; Galitskaya, P.; Selivanovskaya, S. Veterinary antibiotic oxytetracycline’s effect on the soil microbial community. J. Ecol. Environ. 2020, 44, 1–9. [Google Scholar] [CrossRef]
- Fogler, K.; Guron, G.K.P.; Wind, L.L.; Keenum, I.M.; Hession, W.C.; Krometis, L.-A.; Strawn, L.K.; Pruden, A.; Ponder, M.A. Microbiota and Antibiotic Resistome of Lettuce Leaves and Radishes Grown in Soils Receiving Manure-Based Amendments Derived From Antibiotic-Treated Cows. Front. Sustain. Food Syst. 2019, 3, 22. [Google Scholar] [CrossRef]
- Zhu, B.; Chen, Q.; Chen, S.; Zhu, Y.G. Does organically produced lettuce harbor higher abundance of antibiotic resistance genes than conventionally produced? Environ. Int. 2017, 98, 152–159. [Google Scholar] [CrossRef]
- Kovalakova, P.; Cizmas, L.; McDonald, T.J.; Marsalek, B.; Feng, M.; Sharma, V.K. Occurrence and toxicity of antibiotics in the aquatic environment: A review. Chemosphere 2020, 251, 126351. [Google Scholar] [CrossRef]
- Martin-Laurent, F.; Topp, E.; Billet, L.; Batisson, I.; Malandain, C.; Besse-Hoggan, P.; Morin, S.; Artigas, J.; Bonnineau, C.; Kergoat, L.; et al. Environmental risk assessment of antibiotics in agroecosystems: Ecotoxicological effects on aquatic microbial communities and dissemination of antimicrobial resistances and antibiotic biodegradation potential along the soil-water continuum. Environ. Sci. Pollut. Res. 2019, 26, 18930–18937. [Google Scholar] [CrossRef]
- Qiu, D.; Xu, N.; Zhang, Q.; Zhou, W.; Wang, Y.; Zhang, Z.; Qian, H. Negative effects of abamectin on soil microbial communities in the short term. Front. Microbiol. 2022, 13, 1053153. [Google Scholar] [CrossRef]
- Marti, R.; Scott, A.; Tien, Y.-C.; Murray, R.; Sabourin, L.; Zhang, Y.; Topp, E. Impact of Manure Fertilization on the Abundance of Antibiotic-Resistant Bacteria and Frequency of Detection of Antibiotic Resistance Genes in Soil and on Vegetables at Harvest. Appl. Environ. Microbiol. 2013, 79, 5701–5709. [Google Scholar] [CrossRef]
- Chowdhury, F.; Langenkämper, G.; Grote, M. Studies on uptake and distribution of antibiotics in red cabbage. J. Für Verbraucherschutz Und Leb. 2016, 11, 61–69. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial OTU | Habitat/Short Description | Correlation Coefficient with tet(A) Gene Copies |
|---|---|---|
| f__Bacillaceae;g__;s__ | Soil, air, aquatic ecosystems, composts, salt lakes [43]. | 0.86 |
| g__Clostridium;s__ | Soil; gastrointestinal tracts of animals and humans. There are several pathogenic species [44]. | 0.84 |
| g__Ochrobactrum;s__ | Water, soil, plants, and animals. Pathogen [45]. | 0.84 |
| g__Rhizobium;s__leguminosarum | Formation of symbiosis with leguminous plants [46]. | 0.86 |
| f__Enterobacteriaceae;Other;Other | Soil, aquatic ecosystems, animal and human gastrointestinal tracts, plants. There are several pathogenic species [47]. | 0.85 |
| f__Enterobacteriaceae;g__;s__ | 0.83 | |
| g__Erwinia;s__ | Phytopathogen [48]. | 0.85 |
| f__Xanthomonadaceae;g__;s__ | Phytopathogen [49]. | 0.92 |
| g__Akkermansia;s__muciniphila | Human gastrointestinal tract [50]. | 0.85 |
| Bacterial OTU | Habitat/Short Description | Correlation Coefficient with tet(X) Gene Copies |
| g__Pedobacter;s__ | Soil [51] | 0.81 |
| g__Sphingobacterium;s__multvorum | Soil, aquatic ecosystems, plants, wastewater treatment plants [52]. | 0.92 |
| g__Bradyrhizobium;Other | Nitrogen fixation [53]. | 0.90 |
| g__Aminobacter;s__ | Soil [53]. | 0.92 |
| g__Mesorhizobium;Other | Nitrogen fixation [53]. | 0.81 |
| g__Mesorhizobium;s__ | 0.98 | |
| g__Bdellovibrio;Other | Different habitats [54]. | 0.97 |
| g__Bdellovibrio;s__bacteriovorus | 0.93 | |
| f__Moraxellaceae;g__;s__ | Aquatic ecosystems; soil [53]. | 0.92 |
| Bacterial OTU | Hydroponic Experiment | |||||
|---|---|---|---|---|---|---|
| Leaf | Root | |||||
| Control | OTC15 | OTC50 | Control | OTC15 | OTC50 | |
| Unassigned;Other;Other;Other;Other;Other;Other | 54.3 | 73.5 | 42.4 | 7.2 | 9.1 | 1.4 |
| f__Micrococcaceae;g__;s__ | 3.1 | 0.0 | 0.0 | 0.0 | 0.1 | 0.0 |
| g__Rhodococcus;s__ | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| f__Streptomycetaceae;g__;s__ | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| g__Streptomyces;s__ | 0.0 | 0.0 | 0.0 | 0.2 | 0.0 | 0.0 |
| g__Rubrobacter;s__ | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| g__Flavobacterium;s__ | 0.0 | 0.0 | 0.0 | 0.3 | 0.4 | 0.1 |
| o__Sphingobacteriales;f__;g__;s__ | 2.1 | 3.5 | 1.9 | 3.3 | 2.4 | 0.1 |
| g__Sphingobacterium;s__faecium | 0.0 | 0.8 | 0.0 | 0.0 | 0.4 | 3.4 |
| g__Sphingobacterium;s__multivorum | 0.0 | 0.0 | 1.3 | 0.1 | 0.6 | 4.7 |
| o__[Saprospirales];f__;g__;s__ | 0.0 | 0.3 | 0.6 | 0.7 | 0.9 | 0.0 |
| f__Clostridiaceae;Other;Other | 0.0 | 0.0 | 0.0 | 3.4 | 0.1 | 0.0 |
| f__Peptostreptococcaceae;g__;s__ | 0.0 | 0.4 | 1.9 | 3.1 | 0.0 | 0.0 |
| f__Caulobacteraceae;g__;s__ | 0.3 | 1.0 | 0.0 | 0.4 | 3.2 | 2.9 |
| o__Rhizobiales;f__;g__;s__ | 1.4 | 0.0 | 0.0 | 0.4 | 1.3 | 6.2 |
| f__Methylobacteriaceae;g__;s__ | 0.3 | 0.4 | 0.6 | 0.1 | 0.4 | 0.1 |
| g__Aminobacter;s__ | 0.3 | 0.7 | 0.6 | 0.1 | 1.6 | 3.2 |
| g__Agrobacterium;s__ | 0.7 | 0.0 | 0.0 | 5.0 | 1.8 | 0.1 |
| g__Rhizobium;s__leguminosarum | 0.0 | 0.0 | 0.0 | 1.2 | 6.2 | 1.5 |
| f__Rhodospirillaceae;g__;s__ | 2.9 | 2.0 | 0.6 | 0.6 | 0.6 | 0.1 |
| g__Sphingomonas;s__ | 5.0 | 6.1 | 7.6 | 8.8 | 6.9 | 0.4 |
| g__Sphingomonas;s__wittichii | 0.6 | 0.0 | 1.9 | 4.8 | 0.4 | 0.0 |
| g__Achromobacter;s__ | 0.0 | 0.0 | 0.0 | 0.1 | 3.2 | 6.0 |
| f__Comamonadaceae;g__;s__ | 1.0 | 0.0 | 1.3 | 1.2 | 1.1 | 0.2 |
| f__Oxalobacteraceae;g__;s__ | 0.5 | 0.0 | 0.0 | 11.5 | 0.4 | 0.0 |
| f__Methylophilaceae;Other;Other | 0.0 | 2.3 | 0.0 | 0.3 | 10.2 | 0.4 |
| f__Methylophilaceae;g__;s__ | 0.4 | 0.0 | 0.6 | 17.2 | 1.5 | 0.0 |
| g__Bdellovibrio;s__bacteriovorus | 0.0 | 0.3 | 2.5 | 0.0 | 2.4 | 8.6 |
| g__Helicobacter;s__pylori | 0.2 | 0.4 | 3.2 | 0.4 | 0.2 | 0.0 |
| g__Pseudomonas;s__ | 0.3 | 2.3 | 3.2 | 1.3 | 16.0 | 20.1 |
| g__Pseudomonas;s__fragi | 0.0 | 0.0 | 0.0 | 3.6 | 2.4 | 0.0 |
| g__Stenotrophomonas;s__ | 0.0 | 0.0 | 1.3 | 0.5 | 2.7 | 26.0 |
| c__TM7-3;o__;f__;g__;s__ | 6.2 | 0.0 | 0.6 | 0.3 | 0.0 | 0.0 |
| Unassigned;Other;Other;Other;Other;Other;Other | 0.0 | 0.4 | 0.4 | 0.1 | 0.4 | 0.2 |
| f__Micrococcaceae;g__;s__ | 0.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.1 |
| g__Rhodococcus;s__ | 6.0 | 5.6 | 13.2 | 7.1 | 7.6 | 7.1 |
| f__Streptomycetaceae;g__;s__ | 0.0 | 0.0 | 0.4 | 1.7 | 0.8 | 3.2 |
| g__Streptomyces;s__ | 0.0 | 0.0 | 0.0 | 0.7 | 0.7 | 4.8 |
| g__Rubrobacter;s__ | 1.7 | 3.3 | 2.3 | 1.9 | 2.2 | 1.2 |
| g__Flavobacterium;s__ | 0.0 | 0.0 | 0.2 | 5.2 | 2.3 | 0.3 |
| o__Sphingobacteriales;f__;g__;s__ | 3.9 | 5.2 | 6.5 | 4.2 | 3.8 | 3.6 |
| g__Sphingobacterium;s__faecium | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| g__Sphingobacterium;s__multivorum | 0.0 | 0.0 | 0.0 | 0.1 | 0.0 | 0.0 |
| o__[Saprospirales];f__;g__;s__ | 5.0 | 1.0 | 2.2 | 2.2 | 1.5 | 2.1 |
| f__Clostridiaceae;Other;Other | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| f__Peptostreptococcaceae;g__;s__ | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| f__Caulobacteraceae;g__;s__ | 2.1 | 1.5 | 2.3 | 2.8 | 1.0 | 1.4 |
| o__Rhizobiales;f__;g__;s__ | 0.4 | 0.2 | 0.4 | 0.4 | 0.2 | 0.4 |
| f__Methylobacteriaceae;g__;s__ | 1.5 | 1.6 | 0.6 | 1.7 | 3.0 | 0.5 |
| g__Aminobacter;s__ | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| g__Agrobacterium;s__ | 0.0 | 0.0 | 0.0 | 1.3 | 0.3 | 0.3 |
| g__Rhizobium;s__leguminosarum | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| f__Rhodospirillaceae;g__;s__ | 3.1 | 2.8 | 1.7 | 1.3 | 2.1 | 1.7 |
| g__Sphingomonas;s__ | 49.4 | 49.2 | 48.4 | 28.2 | 40.7 | 35.9 |
| g__Sphingomonas;s__wittichii | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| g__Achromobacter;s__ | 0.0 | 0.0 | 0.0 | 0.1 | 0.0 | 0.0 |
| f__Comamonadaceae;g__;s__ | 6.6 | 7.9 | 2.4 | 6.0 | 3.4 | 3.5 |
| f__Oxalobacteraceae;g__;s__ | 0.0 | 0.0 | 0.0 | 0.1 | 0.0 | 0.1 |
| f__Methylophilaceae;Other;Other | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| f__Methylophilaceae;g__;s__ | 0.0 | 0.0 | 0.0 | 3.3 | 0.5 | 0.0 |
| g__Bdellovibrio;s__bacteriovorus | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| g__Helicobacter;s__pylori | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| g__Pseudomonas;s__ | 3.1 | 7.1 | 6.5 | 3.7 | 8.4 | 6.9 |
| g__Pseudomonas;s__fragi | 0.0 | 0.0 | 0.0 | 1.6 | 0.0 | 0.7 |
| g__Stenotrophomonas;s__ | 0.0 | 0.0 | 0.0 | 0.0 | 0.1 | 0.3 |
| c__TM7-3;o__;f__;g__;s__ | 0.0 | 0.0 | 0.0 | 0.0 | 0.1 | 0.0 |
| Bacterial OTU | Water | Soil | ||||
|---|---|---|---|---|---|---|
| Control | OTC15 | OTC50 | Control | OTC15 | OTC300 | |
| Unassigned;Other;Other;Other;Other;Other;Other | 6 | 0 | 0 | 1 | 1 | 1 |
| o__Acidimicrobiales;f__;g__;s__ | 0 | 0 | 0 | 3 | 2 | 2 |
| g__Lentzea;Other | 0 | 0 | 0 | 2 | 1 | 1 |
| f__Micrococcaceae;g__;s__ | 1 | 0 | 0 | 6 | 6 | 6 |
| g__Arthrobacter;s__ | 16 | 0 | 0 | 3 | 3 | 3 |
| f__Nocardioidaceae;g__;s__ | 1 | 0 | 0 | 4 | 3 | 3 |
| g__Patulibacter;s__ | 0 | 4 | 0 | 0 | 0 | 0 |
| g__Flavobacterium;s__ | 5 | 0 | 0 | 1 | 1 | 1 |
| g__Pedobacter;s__ | 0 | 0 | 3 | 0 | 0 | 0 |
| g__Sphingobacterium;s__faecium | 0 | 3 | 0 | 0 | 0 | 0 |
| g__Sphingobacterium;s__multivorum | 0 | 2 | 17 | 0 | 0 | 0 |
| g__Candidatus Rhabdochlamydia;s__ | 4 | 0 | 0 | 0 | 0 | 0 |
| g__Bacillus;s__ | 0 | 0 | 0 | 3 | 2 | 2 |
| o__Rhizobiales;f__;g__;s__ | 2 | 2 | 1 | 1 | 1 | 1 |
| g__Devosia;s__ | 6 | 1 | 1 | 1 | 1 | 1 |
| g__Aminobacter;s__ | 0 | 16 | 15 | 1 | 2 | 2 |
| g__Agrobacterium;s__ | 3 | 0 | 0 | 0 | 0 | 0 |
| f__Rhodospirillaceae;g__;s__ | 3 | 0 | 0 | 0 | 0 | 1 |
| o__Rickettsiales;f__;g__;s__ | 4 | 0 | 0 | 0 | 0 | 0 |
| g__Kaistobacter;s__ | 0 | 0 | 0 | 2 | 2 | 2 |
| g__Novosphingobium;s__ | 2 | 0 | 0 | 0 | 0 | 0 |
| g__Sphingobium;s__ | 4 | 0 | 0 | 0 | 0 | 0 |
| g__Sphingomonas;s__wittichii | 7 | 0 | 0 | 0 | 0 | 0 |
| f__Oxalobacteraceae;g__;s__ | 1 | 0 | 0 | 2 | 2 | 3 |
| f__Methylophilaceae;Other;Other | 0 | 8 | 4 | 0 | 0 | 0 |
| f__Methylophilaceae;g__;s__ | 2 | 1 | 0 | 1 | 2 | 2 |
| g__Methyloversatilis;s__ | 0 | 5 | 0 | 0 | 0 | 0 |
| g__Bdellovibrio;s__bacteriovorus | 0 | 32 | 34 | 0 | 0 | 0 |
| f__Coxiellaceae;g__;s__ | 2 | 0 | 0 | 0 | 0 | 0 |
| g__Pseudomonas;s__ | 1 | 8 | 10 | 0 | 1 | 2 |
| f__Xanthomonadaceae;g__;s__ | 0 | 0 | 0 | 3 | 3 | 2 |
| g__Stenotrophomonas;s__ | 0 | 0 | 3 | 0 | 0 | 0 |
| c__TM7-3;o__;f__;g__;s__ | 3 | 0 | 0 | 0 | 0 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danilova, N.; Galieva, G.; Kuryntseva, P.; Selivanovskaya, S.; Galitskaya, P. Influence of the Antibiotic Oxytetracycline on the Morphometric Characteristics and Endophytic Bacterial Community of Lettuce (Lactuca sativa L.). Microorganisms 2023, 11, 2828. https://doi.org/10.3390/microorganisms11122828
Danilova N, Galieva G, Kuryntseva P, Selivanovskaya S, Galitskaya P. Influence of the Antibiotic Oxytetracycline on the Morphometric Characteristics and Endophytic Bacterial Community of Lettuce (Lactuca sativa L.). Microorganisms. 2023; 11(12):2828. https://doi.org/10.3390/microorganisms11122828
Chicago/Turabian StyleDanilova, Natalia, Gulnaz Galieva, Polina Kuryntseva, Svetlana Selivanovskaya, and Polina Galitskaya. 2023. "Influence of the Antibiotic Oxytetracycline on the Morphometric Characteristics and Endophytic Bacterial Community of Lettuce (Lactuca sativa L.)" Microorganisms 11, no. 12: 2828. https://doi.org/10.3390/microorganisms11122828
APA StyleDanilova, N., Galieva, G., Kuryntseva, P., Selivanovskaya, S., & Galitskaya, P. (2023). Influence of the Antibiotic Oxytetracycline on the Morphometric Characteristics and Endophytic Bacterial Community of Lettuce (Lactuca sativa L.). Microorganisms, 11(12), 2828. https://doi.org/10.3390/microorganisms11122828