

A Polyvalent Adhesin–Toxoid Multiepitope-Fusion-Antigen-Induced Functional Antibodies against Five Enterotoxigenic Escherichia coli Adhesins (CS7, CS12, CS14, CS17, and CS21) but Not Enterotoxins (LT and STa)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Bacteria Strains Used in the Study
2.3. Adhesin MEFA-IIb Construction and Characterization
2.4. Mouse Immunization
2.5. Mouse Antigen-Specific Antibody Titration
2.6. Rabbit Immunization and Challenge
2.7. Statistical Analysis
3. Results
3.1. Adhesin MEFA-IIb Presenting Epitopes of the Five ETEC Adhesins (CS7, CS12, CS14, CS17, and CS21), Two STa Toxoids, and an LT Epitope Was Constructed and Expressed
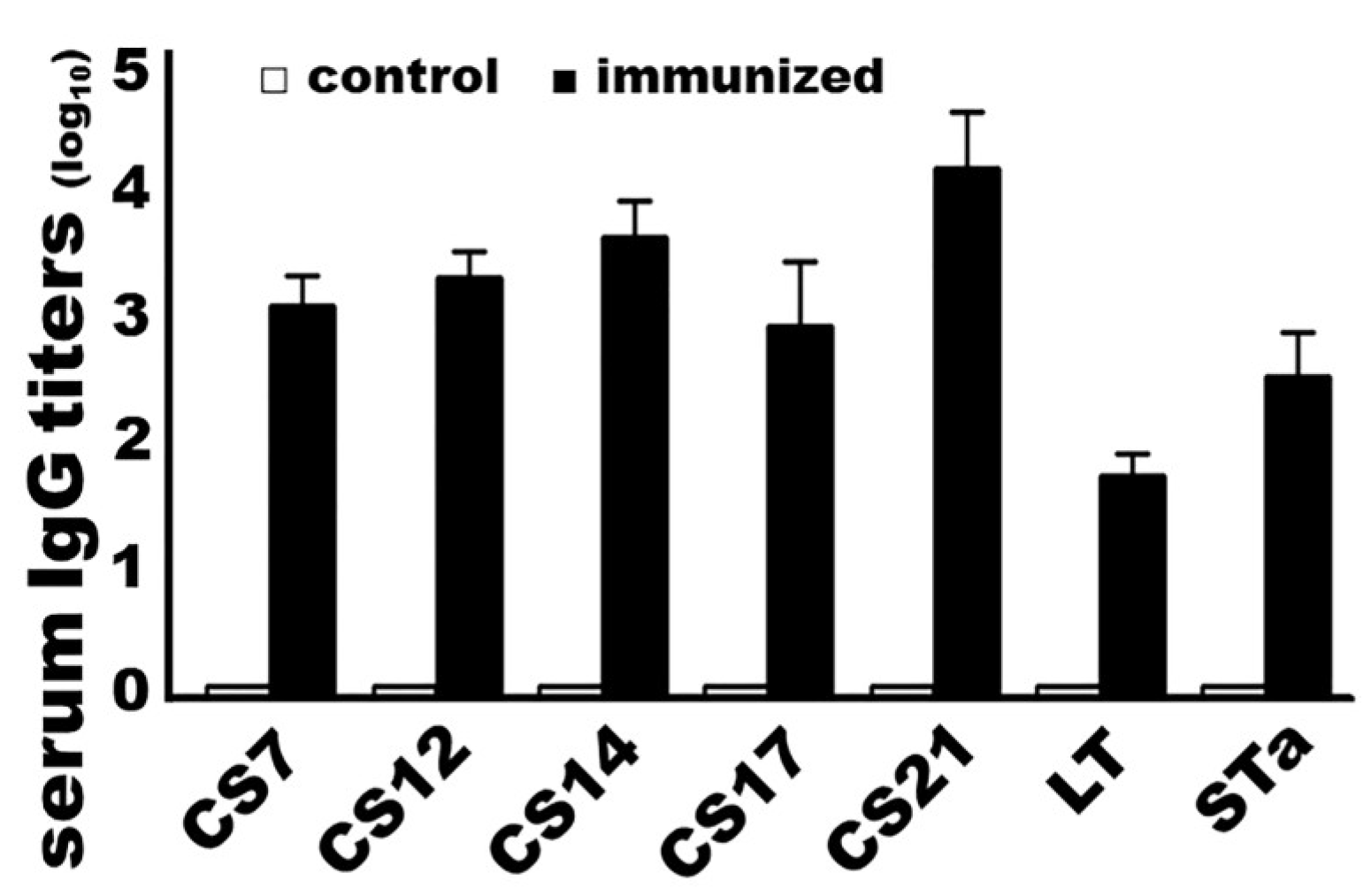
3.2. Mice Immunized with Adhesin MEFA-IIb Developed Strong IgG Titers to Five ETEC Adhesins (CS7, CS12, CS14, CS17, and CS21) as well as to Both ETEC Toxins (LT and STa)
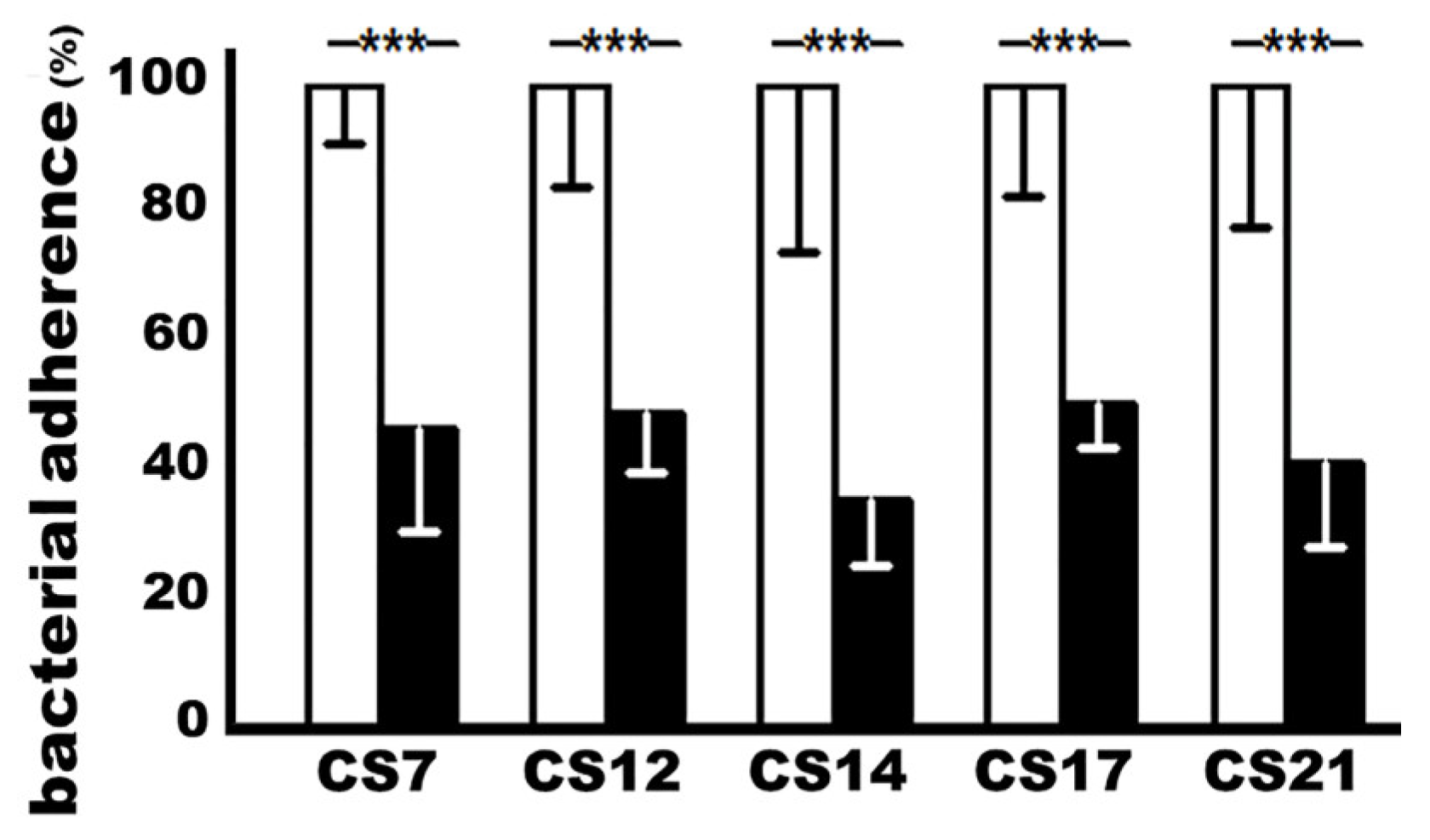
3.3. Adhesin MEFA-IIb-Induced Mouse Serum Antibodies Inhibited the Adherence of CS7, CS12, CS14, CS17, and CS21 Adhesins
3.4. Adhesin MEFA-IIb-Induced Antibodies Did Not Neutralize STa or CT Enterotoxicity
3.5. Adhesin MEFA-IIb Protein Induced Antigen-Specific Antibody Responses in the IM-Immunized Rabbits
3.6. Adhesin MEFA-IIb-Induced Antibodies in Rabbit Serum Samples Broadly Inhibited Adherence by ETEC Bacteria Expressing CS7, CS12, CS14, CS17, or CS21 Adhesin
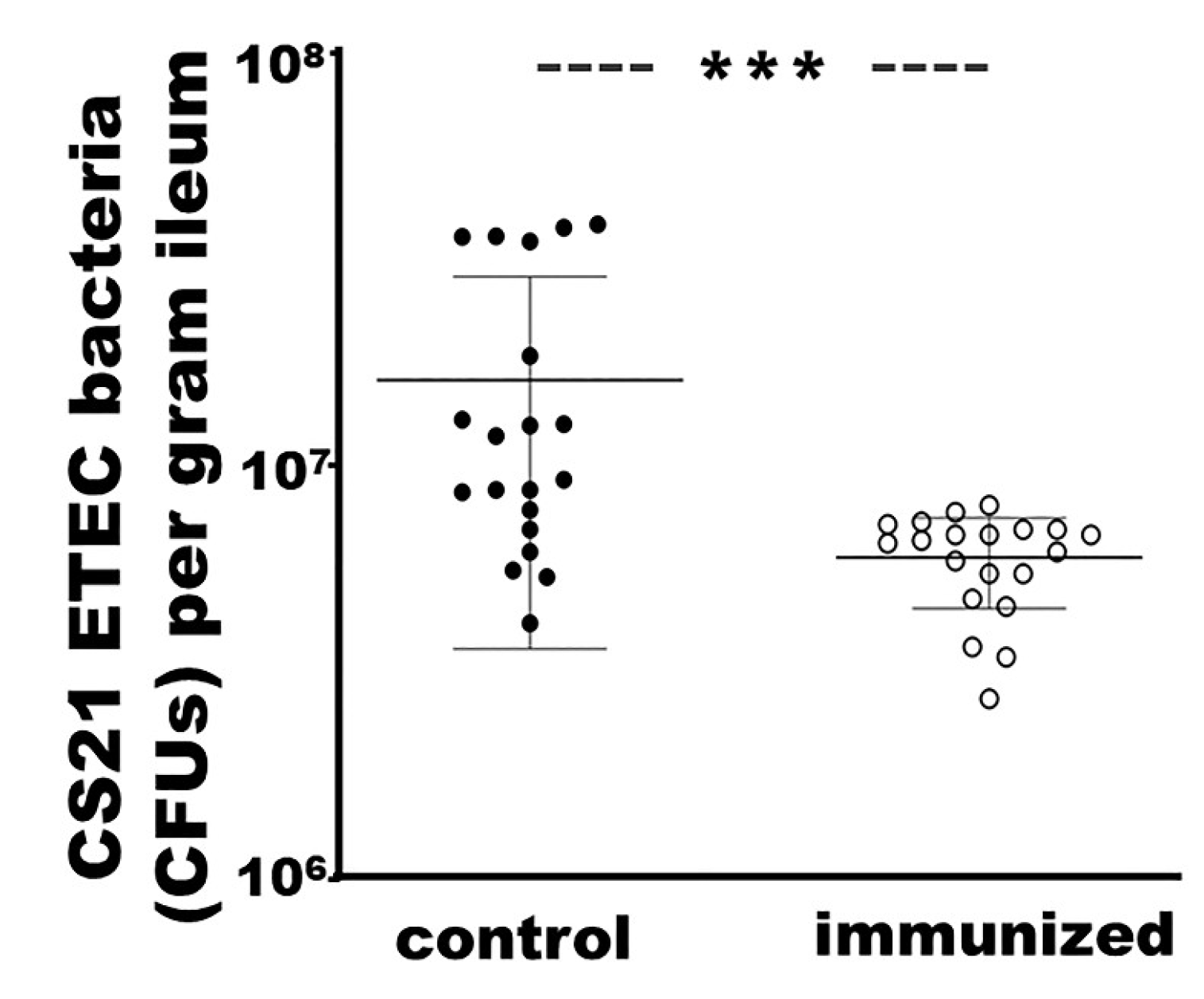
3.7. Rabbits IM Immunized with Adhesin MEFA-IIb Exhibited a Significant Reduction in CS21 ETEC Intestinal Colonization
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): A prospective, case-control study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Platts-Mills, J.A.; Liu, J.; Rogawski, E.T.; Kabir, F.; Lertsethtakarn, P.; Siguas, M.; Khan, S.S.; Praharaj, I.; Murei, A.; Nshama, R.; et al. Use of quantitative molecular diagnostic methods to assess the aetiology, burden, and clinical characteristics of diarrhoea in children in low-resource settings: A reanalysis of the MAL-ED cohort study. Lancet Glob. Health 2018, 6, E1309–E1318. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.D.; DuPont, H.L. Etiology of travellers’ diarrhea. J. Travel. Med. 2017, 24, S13–S16. [Google Scholar] [CrossRef]
- Seo, H.; Duan, Q.; Zhang, W. Vaccines against gastroenteritis, current progress and challenges. Gut Microbes 2020, 11, 1486–1517. [Google Scholar] [CrossRef] [PubMed]
- Svennerholm, A.-M. From cholera to enterotoxigenic Escherichia coli (ETEC) vaccine development. Indian J. Med. Res. 2011, 133, 188–196. [Google Scholar]
- Zhang, W.; Sack, D.A. Progress and hurdles in the development of vaccines against enterotoxigenic Escherichia coli in humans. Expert. Rev. Vaccines 2012, 11, 677–694. [Google Scholar] [CrossRef]
- Isidean, S.D.; Riddle, M.S.; Savarino, S.J.; Porter, C.K. A systematic review of ETEC epidemiology focusing on colonization factor and toxin expression. Vaccine 2011, 29, 6167–6178. [Google Scholar] [CrossRef]
- Qadri, F.; Das, S.K.; Faruque, A.S.; Fuchs, G.J.; Albert, M.J.; Sack, R.B.; Svennerholm, A.M. Prevalence of toxin types and colonization factors in enterotoxigenic Escherichia coli isolated during a 2-year period from diarrheal patients in Bangladesh. J. Clin. Microbiol. 2000, 38, 27–31. [Google Scholar] [CrossRef]
- Gaastra, W.; Svennerholm, A.M. Colonization factors of human enterotoxigenic Escherichia coli (ETEC). Trends Microbiol. 1996, 4, 444–452. [Google Scholar] [CrossRef]
- von Mentzer, A.; Connor, T.R.; Wieler, L.H.; Semmler, T.; Iguchi, A.; Thomson, N.R.; Rasko, D.A.; Joffre, E.; Corander, J.; Pickard, D.; et al. Identification of enterotoxigenic Escherichia coli (ETEC) clades with long-term global distribution. Nat. Genet. 2014, 46, 1321–1326. [Google Scholar] [CrossRef]
- Kuhlmann, F.M.; Martin, J.; Hazen, T.H.; Vickers, T.J.; Pashos, M.; Okhuysen, P.C.; Gomez-Duarte, O.G.; Cebelinski, E.; Boxrud, D.; Del Canto, F.; et al. Conservation and global distribution of non-canonical antigens in Enterotoxigenic Escherichia coli. PLoS Negl. Trop. Dis. 2019, 13, e0007825. [Google Scholar] [CrossRef] [PubMed]
- Vidal, R.M.; Muhsen, K.; Tennant, S.M.; Svennerholm, A.M.; Sow, S.O.; Sur, D.; Zaidi, A.K.M.; Faruque, A.S.G.; Saha, D.; Adegbola, R.; et al. Colonization factors among enterotoxigenic Escherichia coli isolates from children with moderate-to-severe diarrhea and from matched controls in the Global Enteric Multicenter Study (GEMS). PLoS Negl. Trop. Dis. 2019, 13, e0007037. [Google Scholar] [CrossRef] [PubMed]
- Kharat, V.B.; Ahmed, M.; Jiang, Z.D.; Riddle, M.S.; DuPont, H.L. Colonization Factors in Enterotoxigenic Escherichia coli Strains in Travelers to Mexico, Guatemala, and India Compared with Children in Houston, Texas. Am. J. Trop. Med. Hyg. 2017, 96, 83–87. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, R.; Porter, C.K.; Cantrell, J.A.; Denearing, B.; O’Dowd, A.; Grahek, S.L.; Sincock, S.A.; Woods, C.; Sebeny, P.; Sack, D.A.; et al. Volunteer challenge with enterotoxigenic Escherichia coli that express intestinal colonization factor fimbriae CS17 and CS19. J. Infect. Dis. 2011, 204, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Begum, Y.A.; Baby, N.I.; Faruque, A.S.; Jahan, N.; Cravioto, A.; Svennerholm, A.M.; Qadri, F. Shift in phenotypic characteristics of enterotoxigenic Escherichia coli (ETEC) isolated from diarrheal patients in Bangladesh. PLoS Negl. Trop. Dis. 2014, 8, e3031. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lee, K.H.; Zhang, W. Multiepitope fusion antigen: MEFA, an epitope- and structure-based vaccinology platform for multivalent vaccine development. In Methods in Molecular Biology; Bidmos, F., Bossé, J., Langford, P., Eds.; Bacterial Vaccines; Springer Nature: New York, NY, USA, 2022; Volume 2414, pp. 151–169. [Google Scholar]
- Upadhyay, I.; Parvej, S.M.D.; Li, S.; Lauder, K.L.; Shen, Y.; Zhang, W. Polyvalent Protein Adhesin MEFA-II Induces Functional Antibodies against Enterotoxigenic Escherichia coli (ETEC) Adhesins CS7, CS12, CS14, CS17, and CS21 and Heat-Stable Toxin (STa). Appl. Environ. Microbiol. 2023, 89, e0068323. [Google Scholar] [CrossRef] [PubMed]
- Ruan, X.; Knudsen, D.E.; Wollenberg, K.M.; Sack, D.A.; Zhang, W. Multiepitope fusion antigen induces broadly protective antibodies that prevent adherence of Escherichia coli strains expressing colonization factor antigen I (CFA/I), CFA/II, and CFA/IV. Clin. Vaccine Immunol. 2014, 21, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.; Garcia, C.; Ruan, X.; Duan, Q.; Sack, D.A.; Zhang, W. Preclinical characterization of immunogenicity and efficacy against diarrhea from MecVax, a multivalent enterotoxigenic E. coli vaccine candidate. Infect. Immun. 2021, 89, e0010621. [Google Scholar] [CrossRef]
- Jones, R.M., Jr.; Seo, H.; Zhang, W.; Sack, D.A. A multi-epitope fusion antigen candidate vaccine for Enterotoxigenic Escherichia coli is protective against strain B7A colonization in a rabbit model. PLoS Negl. Trop. Dis. 2022, 16, e0010177. [Google Scholar] [CrossRef]
- Upadhyay, I.; Lauder, K.L.; Li, S.; Ptacek, G.; Zhang, W. Intramuscularly Administered Enterotoxigenic Escherichia coli (ETEC) Vaccine Candidate MecVax Prevented H10407 Intestinal Colonization in an Adult Rabbit Colonization Model. Microbiol. Spectr. 2022, 10, e0147322. [Google Scholar] [CrossRef]
- Luo, Q.; Qadri, F.; Kansal, R.; Rasko, D.A.; Sheikh, A.; Fleckenstein, J.M. Conservation and immunogenicity of novel antigens in diverse isolates of enterotoxigenic Escherichia coli. PLoS Negl. Trop. Dis. 2015, 9, e0003446. [Google Scholar] [CrossRef]
- Upadhyay, I.; Li, S.; Ptacek, G.; Seo, H.; Sack, D.A.; Zhang, W. A polyvalent multiepitope protein cross-protects against Vibrio cholerae infection in rabbit colonization and passive protection models. Proc. Natl. Acad. Sci. USA 2022, 119, e2202938119. [Google Scholar] [CrossRef]
- Duan, Q.; Lu, T.; Garcia, C.; Yanez, C.; Nandre, R.M.; Sack, D.A.; Zhang, W. Co-administered tag-less toxoid fusion 3xSTaN12S-mnLTR192G/L211A and CFA/I/II/IV MEFA (multiepitope fusion antigen) induce neutralizing antibodies to 7 adhesins (CFA/I, CS1-CS6) and both enterotoxins (LT, STa) of enterotoxigenic Escherichia coli (ETEC). Front. Microbiol. 2018, 9, e1198. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.Y.; Seo, H.; Sack, D.A.; Zhang, W. Intradermally Administered Enterotoxigenic Escherichia coli Vaccine Candidate MecVax Induces Functional Serum Immunoglobulin G Antibodies against Seven Adhesins (CFA/I and CS1 through CS6) and Both Toxins (STa and LT). Appl. Environ. Microbiol. 2022, 88, e0213921. [Google Scholar] [CrossRef]
- Ruan, X.; Robertson, D.C.; Nataro, J.P.; Clements, J.D.; Zhang, W.; The STa Toxoid Vaccine Consortium Group. Characterization of heat-stable (STa) toxoids of enterotoxigenic Escherichia coli fused to a double mutant heat-labile toxin (dmLT) peptide in inducing neutralizing anti-STa antibodies. Infect. Immun. 2014, 82, 1823–1832. [Google Scholar] [CrossRef] [PubMed]
- Ruan, X.; Sack, D.A.; Zhang, W. Genetic fusions of a CFA/I/II/IV MEFA (multiepitope fusion antigen) and a toxoid fusion of heat-stable toxin (STa) and heat-labile toxin (LT) of enterotoxigenic Escherichia coli (ETEC) retain broad anti-CFA and antitoxin antigenicity. PLoS ONE 2015, 10, e0121623. [Google Scholar] [CrossRef] [PubMed]
- Nandre, R.; Ruan, X.; Duan, Q.; Zhang, W. Enterotoxigenic Escherichia coli heat-stable toxin and heat-labile toxin toxoid fusion 3xSTaN12S-dmLT induces neutralizing anti-STa antibodies in subcutaneously immunized mice. FEMS Microbiol. Lett. 2016, 363, fnw246. [Google Scholar] [CrossRef]
- Nandre, R.M.; Duan, Q.; Wang, Y.; Zhang, W. Passive antibodies derived from intramuscularly immunized toxoid fusion 3xSTaN12S-dmLT protect against STa+ enterotoxigenic Escherichia coli (ETEC) diarrhea in a pig model. Vaccine 2017, 35, 552–556. [Google Scholar] [CrossRef]
- Seo, H.; Lu, T.; Nandre, R.M.; Duan, Q.; Zhang, W. Immunogenicity characterization of genetically fused or chemically conjugated heat-stable toxin toxoids of enterotoxigenic Escherichia coli in mice and pigs. FEMS Microbiol. Lett. 2019, 366, fnz037. [Google Scholar] [CrossRef]
- Duan, Q.; Huang, J.; Xiao, N.; Seo, H.; Zhang, W. Neutralizing anti-STa antibodies derived from enterotoxigenic Escherichia coli (ETEC) toxoid fusions with heat-stable toxin (STa) mutant STaN12S, STaL9A/N12S or STaN12S/A14T show little cross-reactivity with guanylin or uroguanylin. Appl. Environ. Microbiol. 2017, 84, e01737-17. [Google Scholar] [CrossRef]
- Liu, M.; Ruan, X.; Zhang, C.; Lawson, S.R.; Knudsen, D.E.; Nataro, J.P.; Robertson, D.C.; Zhang, W. Heat-labile- and heat-stable-toxoid fusions (LTR192G-STaP13F of human enterotoxigenic Escherichia coli elicit neutralizing antitoxin antibodies. Infect. Immun. 2011, 79, 4002–4009. [Google Scholar] [CrossRef]
- Zhang, C.; Knudsen, D.E.; Liu, M.; Robertson, D.C.; Zhang, W.; The STa Toxoid Vaccine Consortium Group. Toxicity and immunogenicity of enterotoxigenic Escherichia coli heat-labile and heat-stable toxoid fusion 3xSTaA14Q-LTS63K/R192G/L211A in a murine model. PLoS ONE 2013, 8, e77386. [Google Scholar] [CrossRef]
- Huang, J.C.; Duan, Q.D.; Zhang, W.P. Significance of enterotoxigenic Escherichia coli (ETEC) heat-labile toxin (LT) enzymatic subunit epitopes in LT enterotoxicity and immunogenicity. Appl. Environ. Microbiol. 2018, 84, e00849-00818. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, A.; Joseph, S.S.; Reynolds, N.D.; Poncet, D.; Maciel, M., Jr.; Nunez, G.; Espinoza, N.; Nieto, M.; Castillo, R.; Royal, J.M.; et al. Evaluation of the immunogenicity and protective efficacy of a recombinant CS6-based ETEC vaccine in an Aotus nancymaae CS6 + ETEC challenge model. Vaccine 2021, 39, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Gutierrez, R.L.; Maciel, M.; Poole, S.; Testa, K.J.; Trop, S.; Duplessis, C.; Lane, A.; Riddle, M.S.; Hamer, M.; et al. Safety and immunogenicity of intramuscularly administered CS6 subunit vaccine with a modified heat-labile enterotoxin from enterotoxigenic Escherichia coli. Vaccine 2021, 39, 5548–5556. [Google Scholar] [CrossRef] [PubMed]
- Riddle, M.S.; Maciel, M., Jr.; Porter, C.K.; Poole, S.T.; Gutierrez, R.L.; Gormley, R.; Laird, R.M.; Sebeny, P.J.; Dori, K.E.; Greenleaf, M.E.; et al. A first in human clinical trial assessing the safety and immunogenicity of transcutaneously delivered enterotoxigenic Escherichia coli fimbrial tip adhesin with heat-labile enterotoxin with mutation R192G. Vaccine 2020, 38, 7040–7048. [Google Scholar] [CrossRef] [PubMed]
- Rollenhagen, J.E.; Woods, C.M.; O’Dowd, A.; Poole, S.T.; Tian, J.H.; Guebre-Xabier, M.; Ellingsworth, L.; Prouty, M.G.; Glenn, G.; Savarino, S.J. Evaluation of transcutaneous immunization as a delivery route for an enterotoxigenic E. coli adhesin-based vaccine with CfaE, the colonization factor antigen 1 (CFA/I) tip adhesin. Vaccine 2019, 37, 6134–6138. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Seo, H.; Upadhyay, I.; Zhang, W. A Polyvalent Adhesin–Toxoid Multiepitope-Fusion-Antigen-Induced Functional Antibodies against Five Enterotoxigenic Escherichia coli Adhesins (CS7, CS12, CS14, CS17, and CS21) but Not Enterotoxins (LT and STa). Microorganisms 2023, 11, 2473. https://doi.org/10.3390/microorganisms11102473
Li S, Seo H, Upadhyay I, Zhang W. A Polyvalent Adhesin–Toxoid Multiepitope-Fusion-Antigen-Induced Functional Antibodies against Five Enterotoxigenic Escherichia coli Adhesins (CS7, CS12, CS14, CS17, and CS21) but Not Enterotoxins (LT and STa). Microorganisms. 2023; 11(10):2473. https://doi.org/10.3390/microorganisms11102473
Chicago/Turabian StyleLi, Siqi, Hyesuk Seo, Ipshita Upadhyay, and Weiping Zhang. 2023. "A Polyvalent Adhesin–Toxoid Multiepitope-Fusion-Antigen-Induced Functional Antibodies against Five Enterotoxigenic Escherichia coli Adhesins (CS7, CS12, CS14, CS17, and CS21) but Not Enterotoxins (LT and STa)" Microorganisms 11, no. 10: 2473. https://doi.org/10.3390/microorganisms11102473
APA StyleLi, S., Seo, H., Upadhyay, I., & Zhang, W. (2023). A Polyvalent Adhesin–Toxoid Multiepitope-Fusion-Antigen-Induced Functional Antibodies against Five Enterotoxigenic Escherichia coli Adhesins (CS7, CS12, CS14, CS17, and CS21) but Not Enterotoxins (LT and STa). Microorganisms, 11(10), 2473. https://doi.org/10.3390/microorganisms11102473