
The Influence of the Genotype and Planting Density on the Structure and Composition of Root and Rhizosphere Microbial Communities in Maize


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Maize Accessions and Experimental Field Site
2.2. Planting Density Design
2.3. Rhizosphere and Root Sampling
2.4. DNA Extraction, Library Construction, and Sequencing
2.5. Bioinformatics Analysis
3. Results
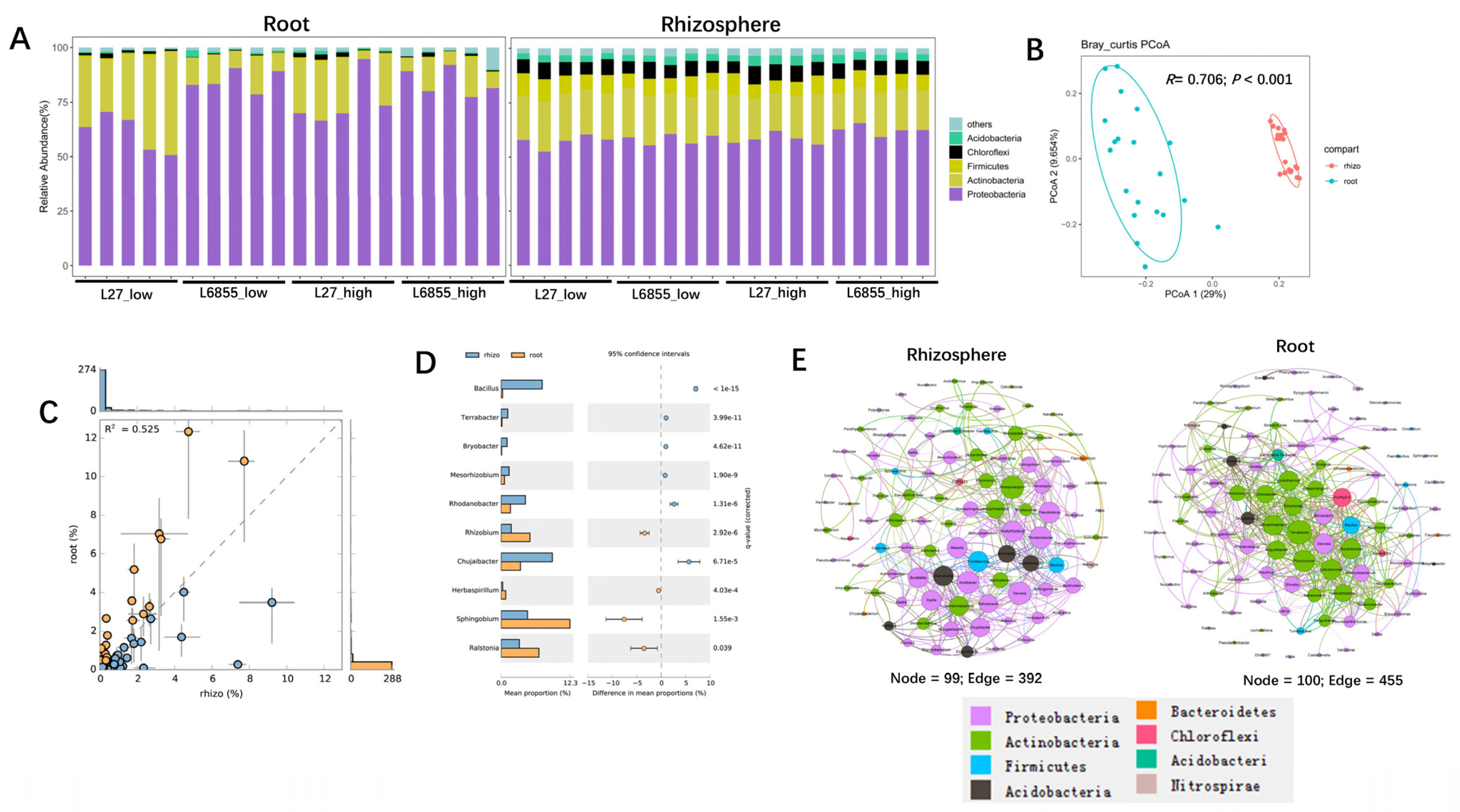
3.1. Bacterial Community Composition in the Rhizosphere and Roots of Maize
3.2. Microbes Specifically Selected by Maize Roots
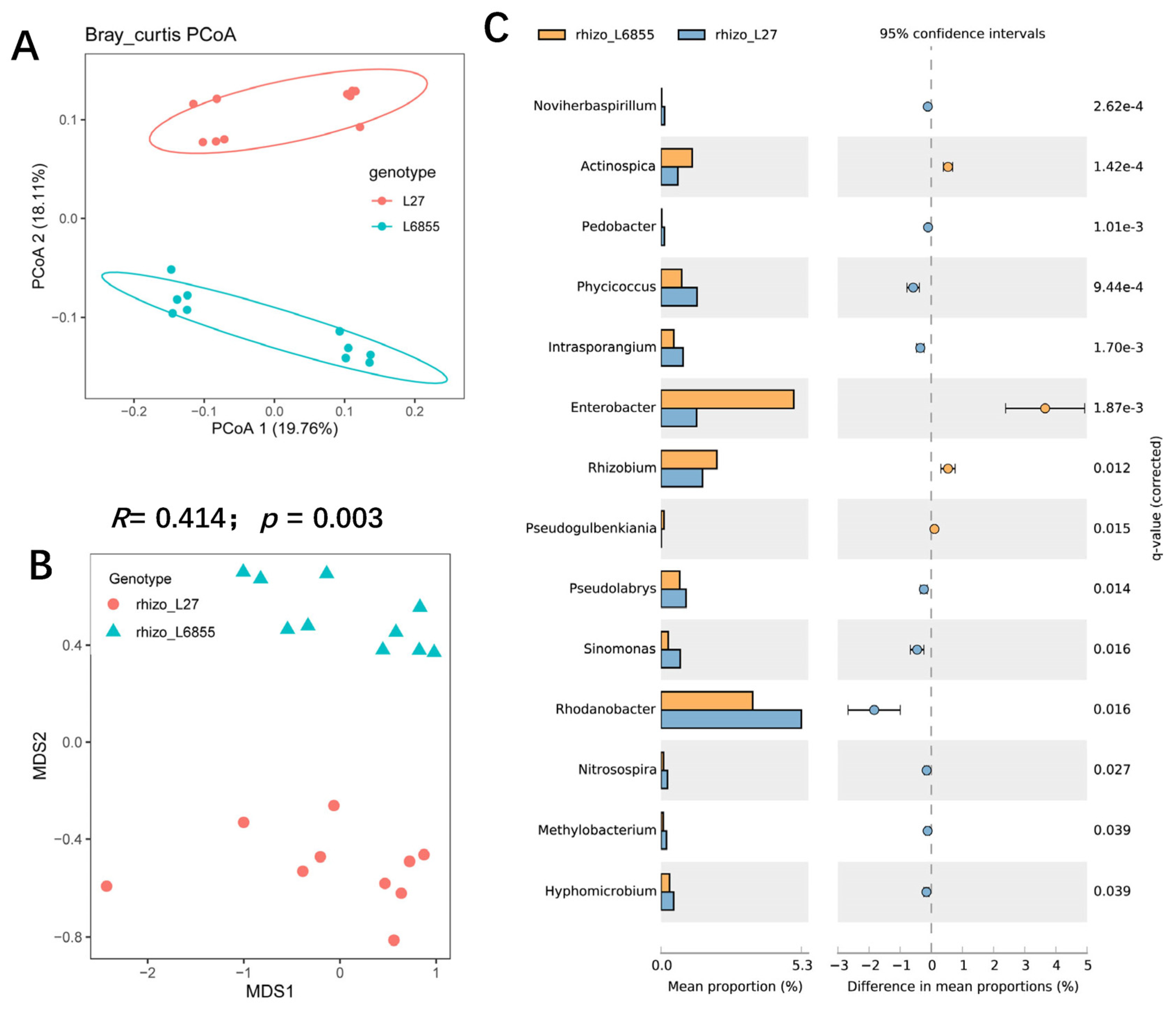
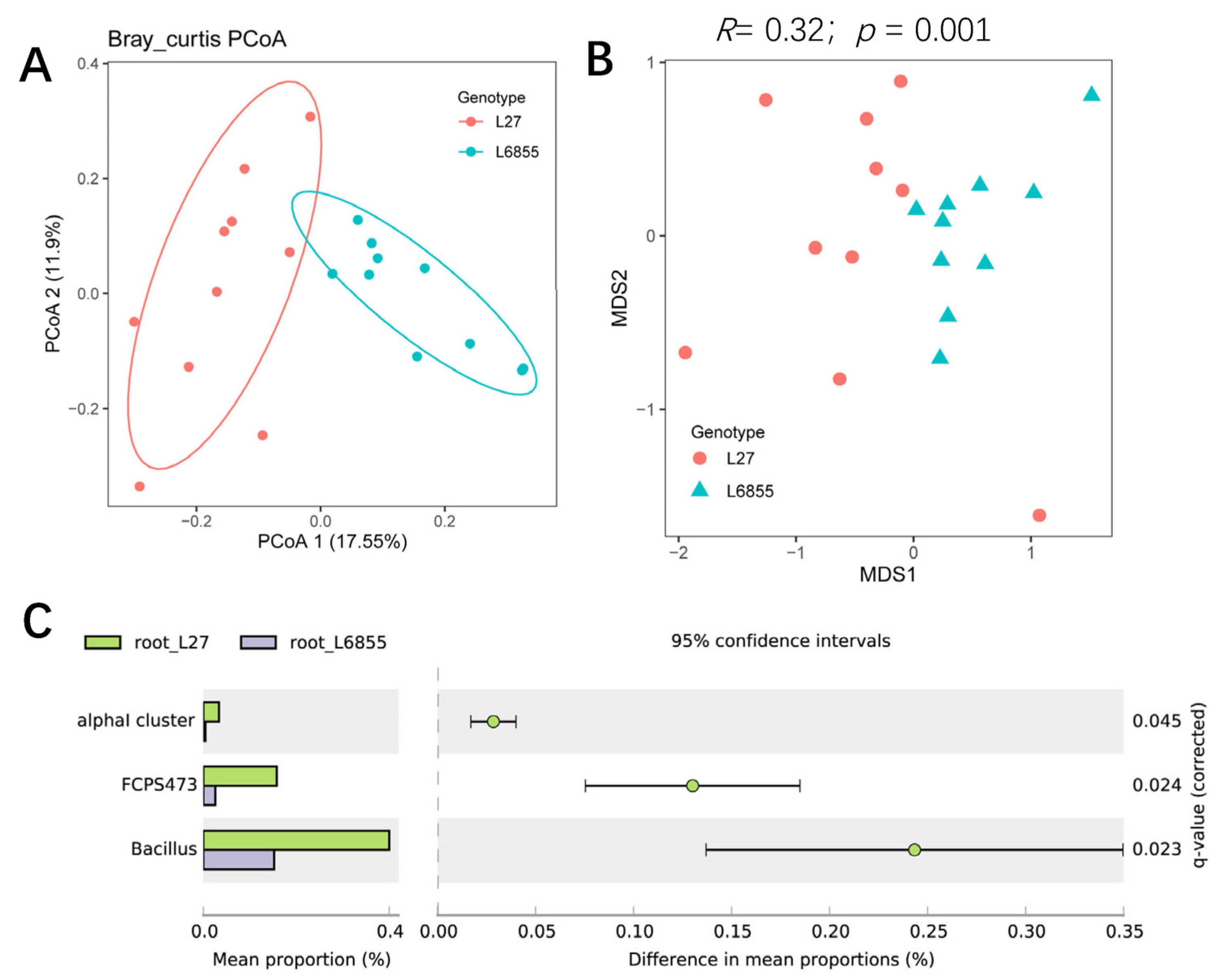
3.3. The Maize Genotypic Traitsthe Structure and Composition of Root-Associated Microbiota
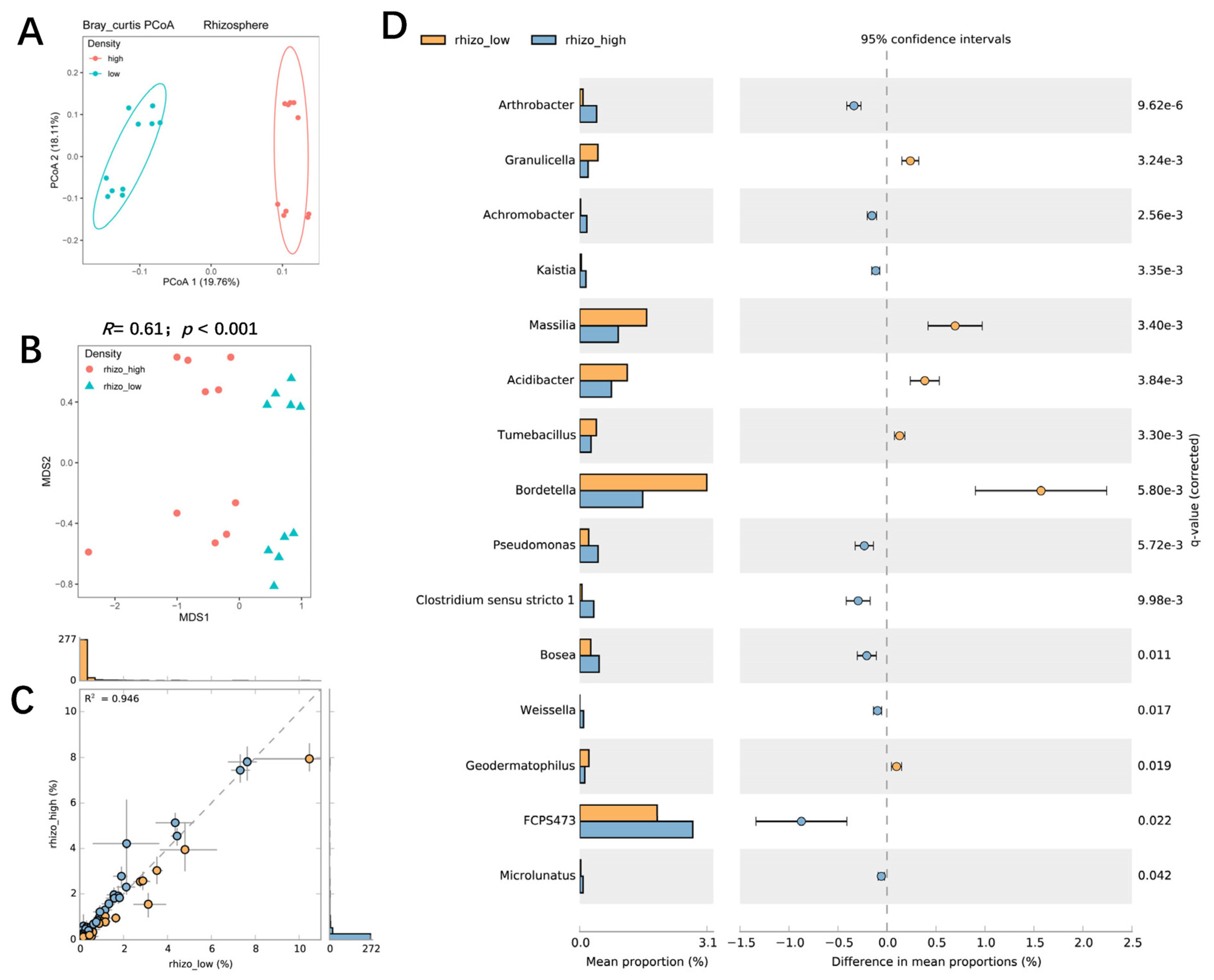
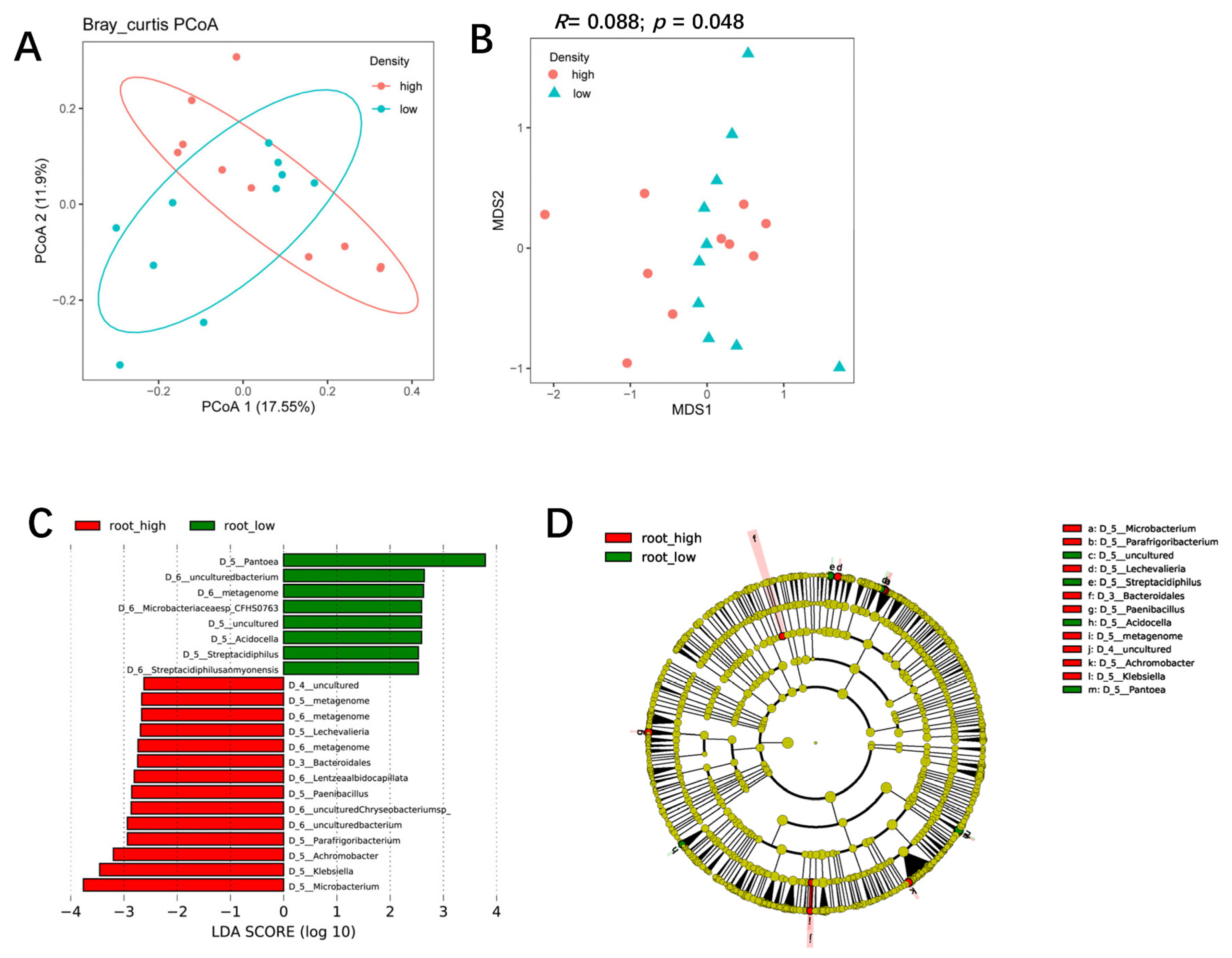
3.4. The Influence of Planting Density on Bacterial Communities in the Rhizosphere and Roots of Maize
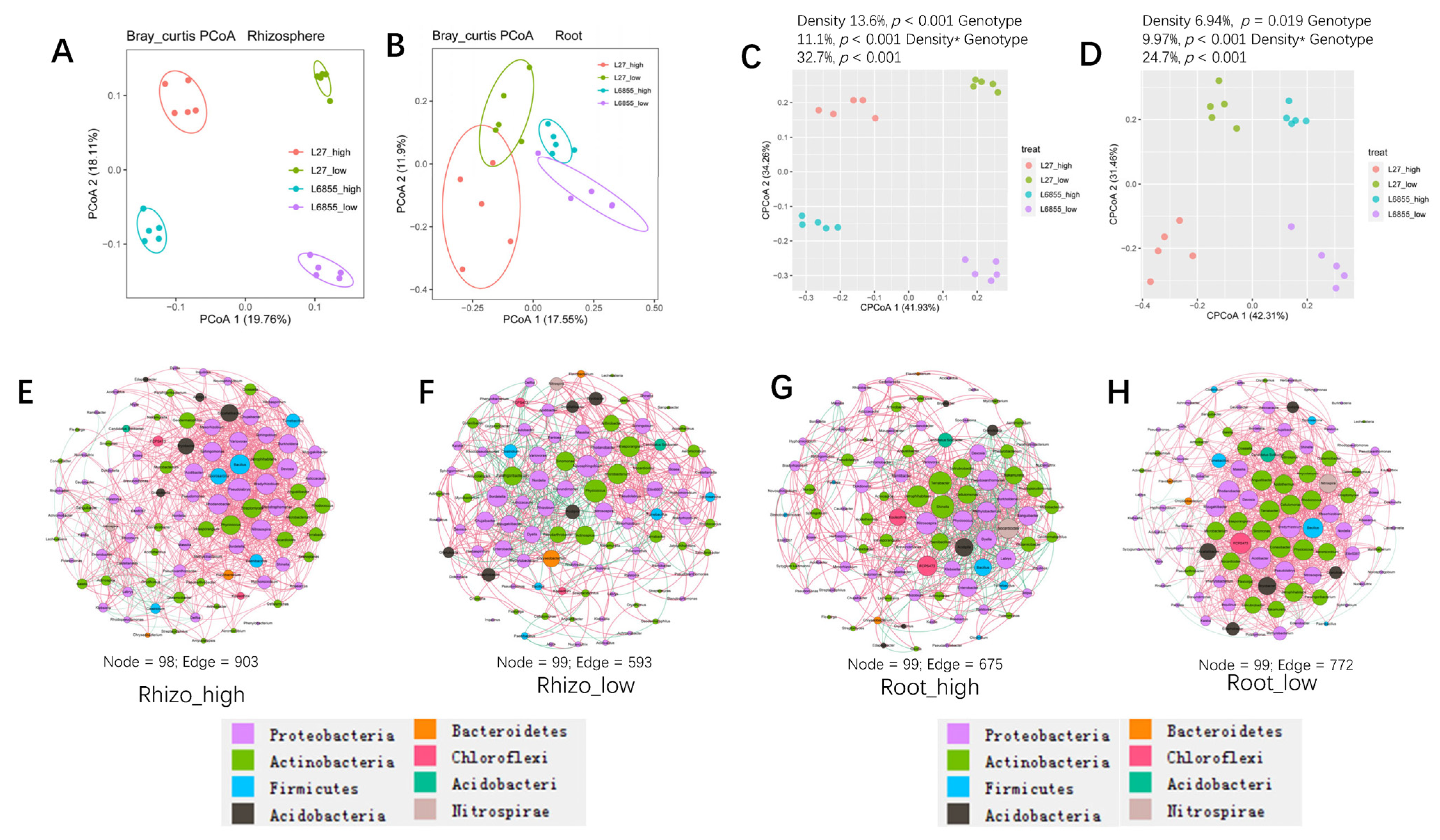
3.5. Genotype and Planting Density Have a Synergistic Effect on the Bacterial Community
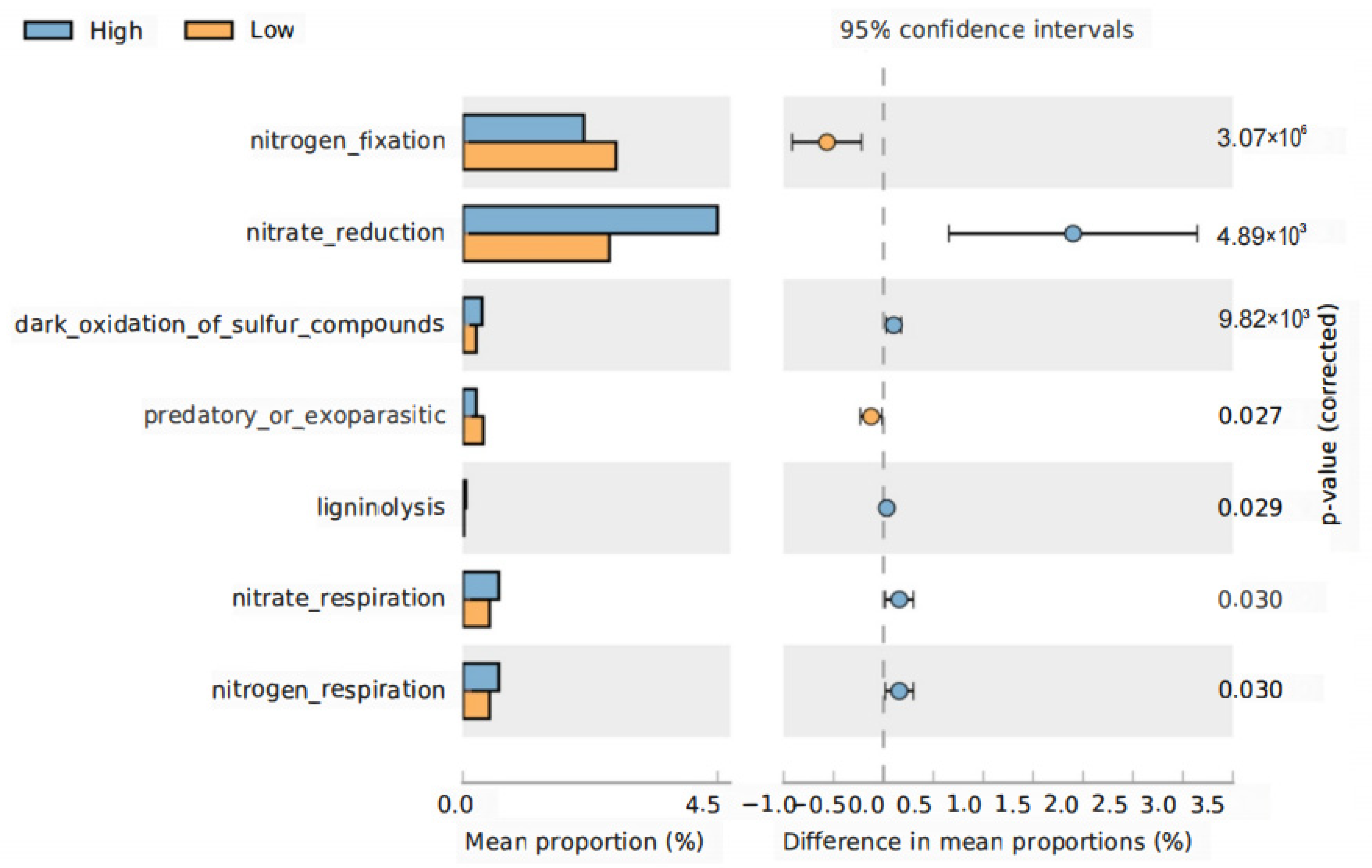
3.6. Functional Shifts in Rhizosphere Bacteria under Different Planting Densities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Philippot, L.; Griffiths, B.S.; Langenheder, S. Microbial community resilience across ecosystems and multiple disturbances. Microbiol. Mol. Biol. Rev. 2021, 85, e00026-20. [Google Scholar] [CrossRef]
- Li, W.; Niu, Z.; Chen, H.; Li, D. Characterizing canopy structural complexity for the estimation of maize LAI based on ALS data and UAV stereo images. Int. J. Remote Sens. 2017, 38, 2106–2116. [Google Scholar] [CrossRef]
- Li, S.K.; Zhao, J.R.; Dong, S.T.; Zhao, M.; Li, C.H.; Cui, Y.H. Advances and prospects of maize cultivation in China. Sci. Agric. Sin. 2017, 50, 1941–1959. [Google Scholar]
- Slattery, R.A.; Ort, D.R. Perspectives on improving light distribution and light use efficiency in crop canopies. Plant Physiol. 2021, 185, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Sun, L.; Mou, H.; Ali, S.; Liu, D.; Zhang, Y. Effects of planting patterns and sowing densities on grain-filling, radiation use efficiency and yield of maize (Zea mays L.) in semi-arid regions. Agric. Water Manag. 2018, 201, 287–298. [Google Scholar] [CrossRef]
- Li, T.; Liu, Y.; Shi, L.; Jiang, C.D. Systemic regulation of photosynthetic function in field-grown sorghum. Plant Physiol. Biochem. 2015, 94, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Bloor, J.M.G.; Grubb, P.J. Morphological plasticity of shade-tolerant tropical rainforest tree seedlings exposed to light changes. Funct. Ecol. 2004, 18, 337–348. [Google Scholar] [CrossRef]
- Li, G.H.; Liu, J.; Dong, S.T.; Liu, P.; Zhang, J.W.; Zhao, B. Effects of close planting and nitrogen application rates on grain yield and nitrogen utilization efficiency of different density-tolerance maize hybrids. Sci. Agric. Sin. 2017, 50, 2247–2258. [Google Scholar]
- Chapepa, B.; Mudada, N.; Mapuranga, R. The impact of plant density and spatial arrangement on light interception on cotton crop and seed cotton yield: An overview. J. Cotton Res. 2020, 3, 210–215. [Google Scholar] [CrossRef]
- Boomsma, C.R.; Santini, J.B.; Tollenaar, M. Maize more phophysiological responses to intense crowding and low nitrogen availability: An analysis and review. Agron. J. 2010, 101, 1426–1452. [Google Scholar] [CrossRef]
- Huot, B.; Yao, J.; Montgomery, B.L.; He, S.Y. Growth-defense tradeofs in plants: A balancing act to optimize fitness. Mol. Plant 2014, 7, 1267–1287. [Google Scholar] [CrossRef]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial interactions within the plant holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Kracher, B.; Hiruma, K.; Münch, P.C.; Thon, M.R.; Weimann, A.; Damm, U.; Dallery, J.-F.; Hainaut, M.; Henrissat, B.; Lespinet, O.; et al. Survival trade-ofs in plant roots during colonization by closely related benefcial and pathogenic fungi. Nat. Commun. 2016, 7, 11362. [Google Scholar]
- Castrillo, G.; Teixeira, P.; Paredes, S.H.; Law, T.F.; Lorenzo, L.; Feltcher, M.E. Root microbiota drive direct integration of phosphate stress and immunity. Nature 2017, 543, 513–518. [Google Scholar] [CrossRef]
- Stringlis, I.A.; Yu, K.; Feussner, K.J.; de Jonge, R.; Van Bentum, S.; Van Verk, M.C.; Berendsen, R.L.; Bakker, P.A.H.M.; Fewussner, I.; Pieterse, C.M.J. MYB72-dependent coumarin exudation shapes root microbiome assembly to promote plant health. Proc. Natl Acad. Sci. USA 2018, 115, E5213–E5222. [Google Scholar] [CrossRef] [PubMed]
- Hacquard, S.; Garrido-Oter, R.; Gonzalez, A.; Spaepen, S.; Ackermann, G.; Lebeis, S. Microbiota and host nutrition across plant and animal kingdoms. Cell Host Microbe 2015, 17, 603–616. [Google Scholar] [CrossRef]
- Jones, D.L.; Nguyen, C.; Finlay, D. Carbon flow in the rhizosphere: Carbon trading at the soil-root interface. Plant Soil 2009, 321, 5–33. [Google Scholar] [CrossRef]
- Hirsch, A.M.; Alvarado, J.; Bruce, D.; Chertkov, O.; Hoff, P.L.; Detter, J.C. Complete Genome Sequence of Micromonospora Strain L5, a Potential Plant-Growth-Regulating Actinomycete, Originally Isolated from Casuarina equisetifolia Root Nodules. Genome Announc. 2013, 5, e00759. [Google Scholar] [CrossRef] [PubMed]
- Nihorimbere, V.; Cawoy, H.; Seyer, A.; Brunelle, A.; Thonart, P.; Ongena, M. Impact of rhizosphere factors on cyclic lipopeptide signature from the plant beneficial strain Bacillus amyloliquefaciens S499. FEMS Microbiol. Ecol. 2012, 79, 176–191. [Google Scholar] [CrossRef]
- Dohrmann, A.B.; Küting, M.; Jünemann, S.; Jaenicke, S.; Schlüter, A.; Tebbe, C.C. Importance of rare taxa for bacterial diversity in the rhizosphere of Bt and conventional maize varieties. ISME J. 2013, 7, 37–49. [Google Scholar] [CrossRef]
- Zhong, Y.; Yang, Y.; Liu, P.; Xu, R.; Rensing, C.; Fu, X. Genotype and rhizobium inoculation modulate the assembly of soybean rhizobacterial communities. Plant Cell Environ. 2019, 42, 2028–2044. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Ver Loren Themaat, E.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 2012, 64, 807–838. [Google Scholar] [CrossRef]
- Lundberg, D.S.; Lebeis, S.L.; Paredes, S.H.; Yourstone, S.; Gehring, J.; Malfatti, S. Defining the core Arabidopsis thaliana root microbiome. Nature 2012, 488, 86–90. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Garrido-Oter, R.; Münch, P.C.; Weiman, A.; Dröge, J.; Pan, Y. Structure and function of the bacterial root microbiota in wild and domesticated barley. Cell Host Microbe 2015, 17, 392–403. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Amir, A.; Mcdonald, D.; Navas-Molina, J.A.; Kopylova, E.; Morton, J.T.; ZechXu, Z. Deblur rapidly resolves single-nucleotide community sequence patterns. mSystems 2017, 2, e00191-16. [Google Scholar] [CrossRef]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.B. Vegan: Community Ecology Package, R package version 2.3-0; R Foundation: Ames, IA, USA, 2015. [Google Scholar]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Rott, M.; Schlaeppi, K.; Ver Loren Van Themaat, E.; Ahmadinejad, N.; Assenza, F. Revealing structure and assembly cues for Arabidopsis root-inhabiting bacterial microbiota. Nature 2012, 488, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Lebeis, S.L.; Paredes, S.H.; Lundberg, D.S.; Breakfield, N.; Gehring, J.; McDonald, M. Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa. Science 2015, 349, 860–864. [Google Scholar] [CrossRef]
- Edwards, J.; Johnson, C.; Santos-Medellín, C.; Lurie, E.; Podishetty, N.; Bhatnagar, S. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, E911–E920. [Google Scholar] [CrossRef] [PubMed]
- Walters, W.A.; Jin, Z.; Nicholas, Y.; Wallace, J.G.; Jessica, S.; Zhang, W.; González-Peña, A.; Peiffer, J.; Koren, O.; Shi, Q.; et al. Large-scale replicated field study of maize rhizosphere identifies heritable microbes. Proc. Natl. Acad. Sci. USA 2018, 115, 7368–7373. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2014, 8, 790–803. [Google Scholar] [CrossRef]
- Schlaeppi, K.; Dombrowski, N.; Oter, R.G.; Ver Loren van Themaat, E.; Schulze-Lefert, P. Quantitative divergence of the bacterial root microbiota in Arabidopsis thaliana relatives. Proc. Natl. Acad. Sci. USA 2014, 111, 585–592. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.X.; Zhang, N.; Hu, B.; Jin, T.; Xu, H.; Qin, J.; Yan, P.; Zhang, X.; Guo, X.; et al. NRT1.1B is associated with root microbiota composition and nitrogen use in field-grown rice. Nat. Biotechnol. 2019, 37, 676–684. [Google Scholar] [CrossRef]
- Zarraonaindia, I.; Sarah, M.O.; Pamela, W.; Kristin, W.; Jarrad, H.; Simon, L.; Nicholas, A.B.; David, A.M.; Gilles, M.; Safiyh, T.; et al. The Soil Microbiome Influences Grapevine-Associated Microbiota. ASM J. 2015, 24, e02527-14. [Google Scholar] [CrossRef]
- Xu, H.; Yang, Y.; Tian, Y.; Xu, R.; Zhong, Y.; Liao, H. Rhizobium Inoculation Drives the Shifting of Rhizosphere Fungal Community in a Host Genotype Dependent Manner. Front. Microbiol. 2020, 10, 3135. [Google Scholar] [CrossRef] [PubMed]
- Kudjordjie, E.N.; Sapkota, R.; Steffensen, S.K.; Fomsgaard, I.S.; Nicolaisen, M. Maize synthesized benzoxazinoids affect the host associated microbiome. Microbiome 2019, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Matus-Acuña, V.; Caballero-Flores, G.; Martínez-Romero, E. The influence of maize genotype on the rhizosphere eukaryotic community. FEMS Microbiol. Ecol. 2021, 97, fiab066. [Google Scholar] [CrossRef]
- Lopes, L.D.; Wang, P.; Futrell, S.L.; Schachtman, D.P. Sugars and jasmonic acid concentration in root exudates affect maize rhizosphere bacterial communities. Appl. Environ. Microbiol. 2022, 88, e0097122. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Xiao, K.; Guo, X.; Zhang, F.; Zhao, C. Review on Maize Canopy Structure, Light Distributing and Canopy Photosynthesis. J. Maize Sci. 2005, 13, 55–59. [Google Scholar]
- Xue, J.Q.; Liang, Z.S.; Ma, G.S.; Lu, H.; Ren, J. Population physiological indices on density-tolerance of maize in different plant type. Chin. J. Appl. Ecol. 2002, 13, 55–59. [Google Scholar]
- Wang, X.K.; Wang, X.L.; Sun, S.L.; Tu, X.Y.; Lin, K.D.; Qin, L.; Wang, X.Y.; Li, G.; Zhong, S.L.; Li, P.H. Characterization of regulatory modules controlling leaf angle in maize. Plant Physiol. 2022, 190, 500–515. [Google Scholar] [CrossRef]
- Liu, T.; Chen, J.; Wang, Z.; Wu, X.; Ding, R.; Han, Q.; Cai, T.; Jia, Z. Ridge and furrow planting pattern optimizes canopy structure of summer maize and obtains higher grain yield. Field Crops Res. 2018, 219, 242–249. [Google Scholar] [CrossRef]
- Chang, X.; Wei, D.; Zeng, Y.; Zhao, X.; Hu, Y.; Wu, X.; Song, C.; Gong, G.; Chen, H.; Yang, C.; et al. Maize-soybean relay strip intercropping reshapes the rhizosphere bacterial community and recruits beneficial bacteria to suppress Fusarium root rot of soybean. Front. Microbiol. 2022, 13, 1009689. [Google Scholar] [CrossRef]
- Zhang, R.; Mu, Y.; Li, X.; Li, S.; Sang, P.; Wang, X.; Wu, H.; Xu, N. Response of the arbuscular mycorrhizal fungi diversity and community in maize and soybean rhizosphere soil and roots to intercropping systems with different nitrogen application rates. Sci. Total Environ. 2020, 740, 139810. [Google Scholar] [CrossRef]
- Zhong, Y.; Liang, L.; Xu, R.; Xu, H.; Sun, L.; Liao, H. Intercropping tea plantations with soybean and rapeseeds enhances nitrogen fixation through shifts in soil microbial communities. Front. Agric. Sci. Eng. 2022, 9, 344–355. [Google Scholar]
- Wang, X.; Feng, H.; Wang, Y.; Wang, M.; Xie, X.; Chang, H.; Wang, L.; Qu, J.; Sun, K.; He, W.; et al. Mycorrhizal symbiosis modulates the rhizosphere microbiota to promote rhizobia-legume symbiosis. Mol. Plant 2021, 14, 503–516. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.M.; Reynolds, H.L. Plant-soil feedbacks shift from negative to positive with decreasing light in forest understory species. Ecology 2015, 96, 2523–2532. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.C.; Graham, J.H.; Smith, F.A. Functioning of mycorrhizal associations along the mutualism-parasitism continuum. New Phytol. 1997, 135, 575–585. [Google Scholar] [CrossRef]
- Song, C.; Wang, W.; Gan, Y.; Wang, L.; Chang, X.; Wang, Y.; Yang, W. Growth promotion ability of phosphate-solubilizing bacteria from the soybean rhizosphere under maize-soybean intercropping systems. J. Sci. Food Agric. 2022, 102, 1430–1442. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Lin, J.; Chen, S.; Lu, H.; Liao, C. The Influence of the Genotype and Planting Density on the Structure and Composition of Root and Rhizosphere Microbial Communities in Maize. Microorganisms 2023, 11, 2443. https://doi.org/10.3390/microorganisms11102443
Zhang Y, Lin J, Chen S, Lu H, Liao C. The Influence of the Genotype and Planting Density on the Structure and Composition of Root and Rhizosphere Microbial Communities in Maize. Microorganisms. 2023; 11(10):2443. https://doi.org/10.3390/microorganisms11102443
Chicago/Turabian StyleZhang, Yang, Jianxin Lin, Shanhu Chen, Heding Lu, and Changjian Liao. 2023. "The Influence of the Genotype and Planting Density on the Structure and Composition of Root and Rhizosphere Microbial Communities in Maize" Microorganisms 11, no. 10: 2443. https://doi.org/10.3390/microorganisms11102443
APA StyleZhang, Y., Lin, J., Chen, S., Lu, H., & Liao, C. (2023). The Influence of the Genotype and Planting Density on the Structure and Composition of Root and Rhizosphere Microbial Communities in Maize. Microorganisms, 11(10), 2443. https://doi.org/10.3390/microorganisms11102443