

Metabolic Profiling and Comparative Proteomic Insight in Respect of Amidases during Iprodione Biodegradation

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Media
2.2. Sample Collection and Isolation of IPR-Degrading Bacteria
2.3. Inoculum Preparation
2.4. Screening of IPR-Degrading Bacteria
2.5. Characterization and Identification of IPR-Degrading Bacterial Strains
2.6. Bacterial Growth and IPR Removal
2.7. Proteome Preparation, Digestion, and Mass Spectrometry Analysis
2.8. Data Analysis
2.8.1. Bacterial Growth and IPR Degradation Analysis
2.8.2. Proteomic Data Analysis
2.8.3. Pathway Analysis
2.9. Statistical Analysis
3. Results
3.1. Isolation and Selection of IPR-Degrading Bacteria
3.2. Characterization and Molecular Identification
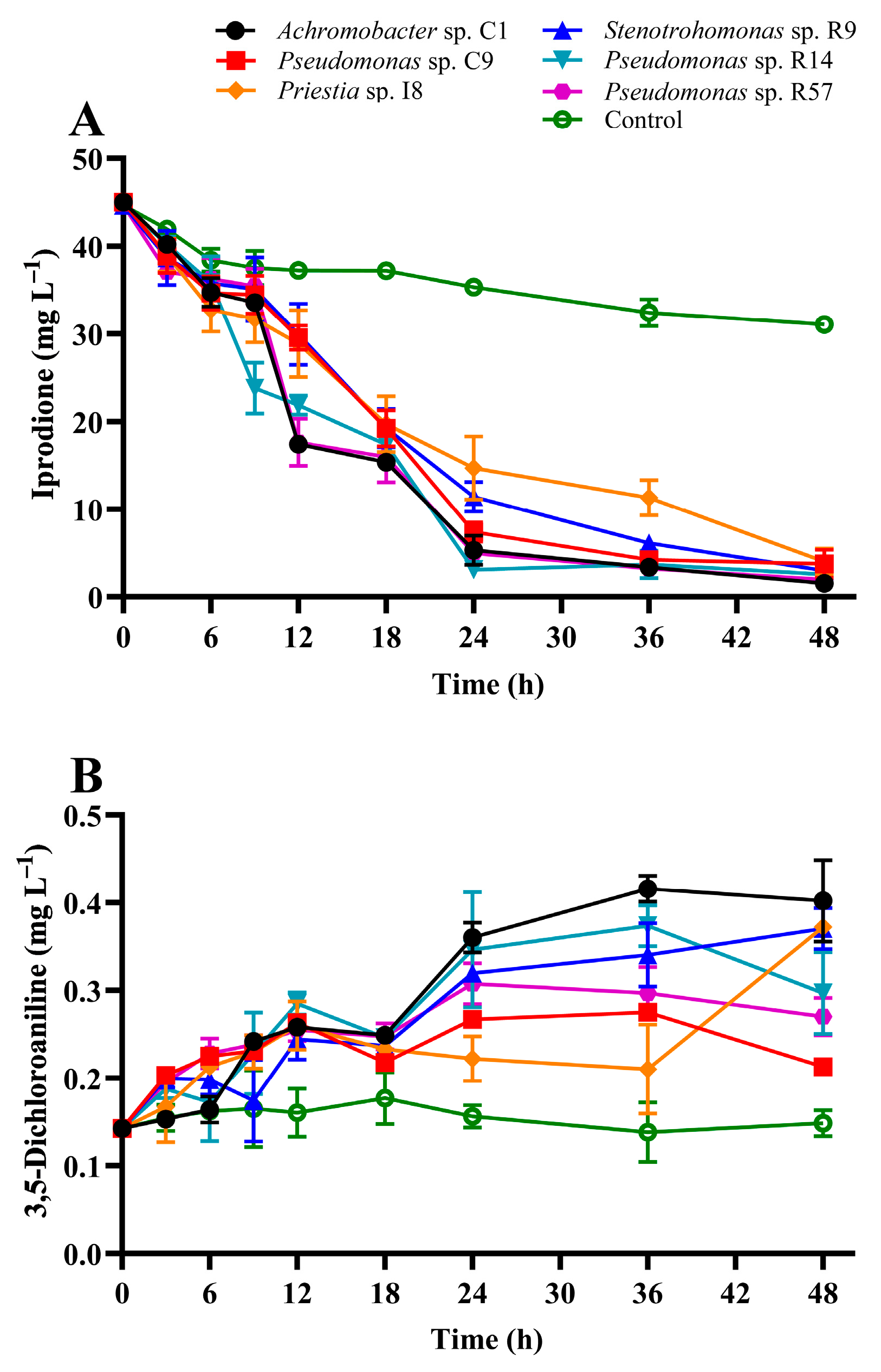
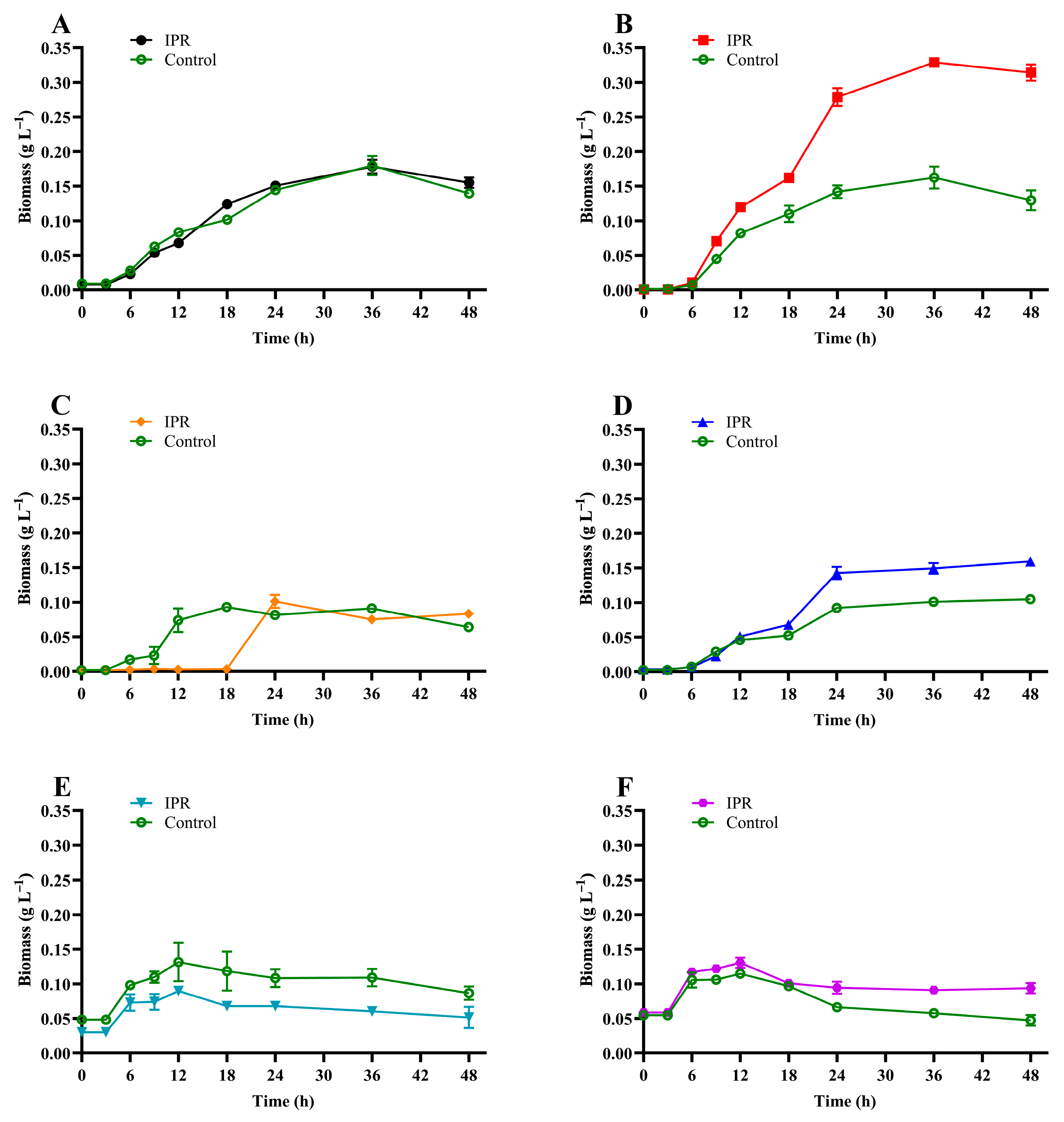
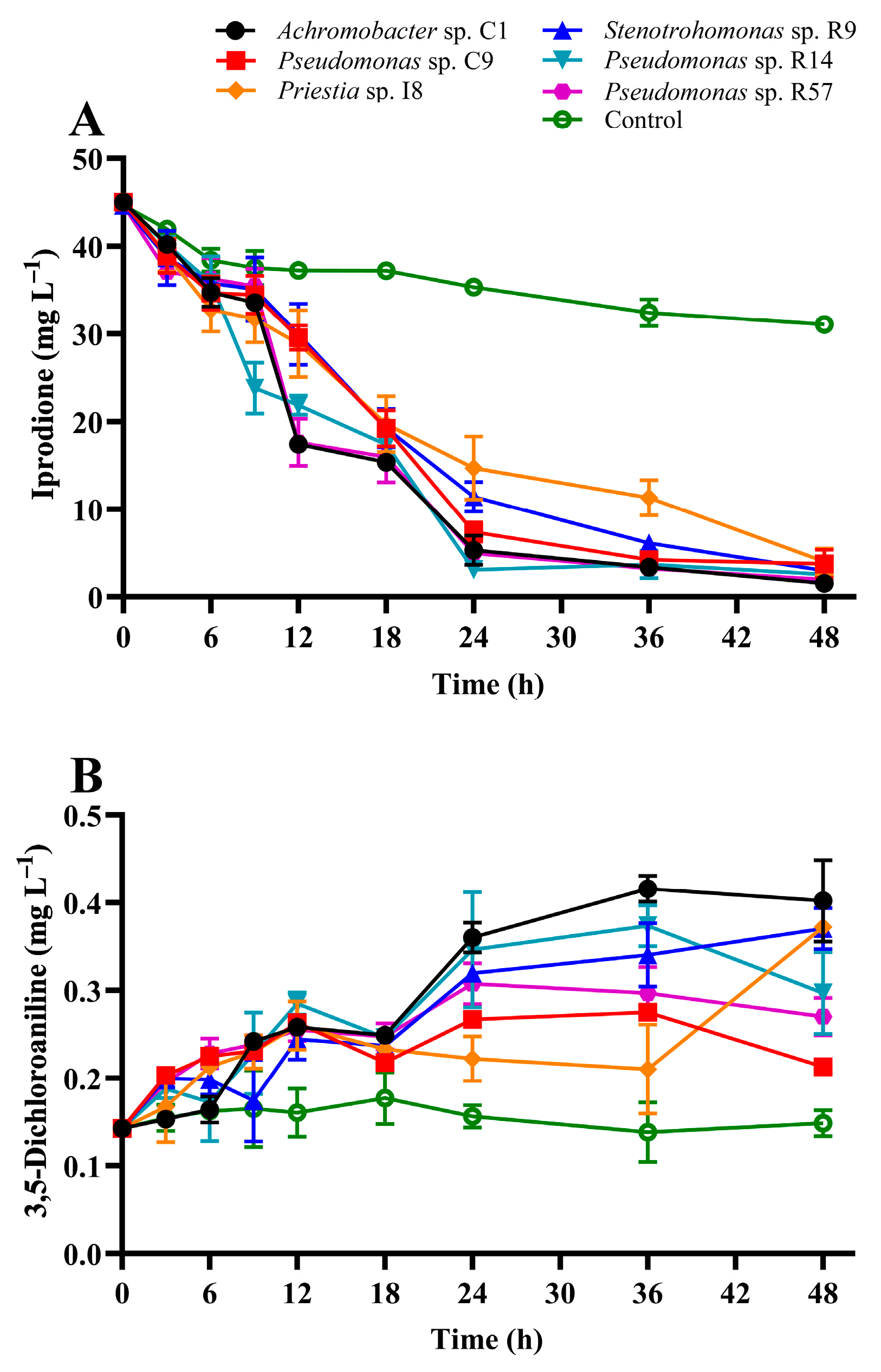
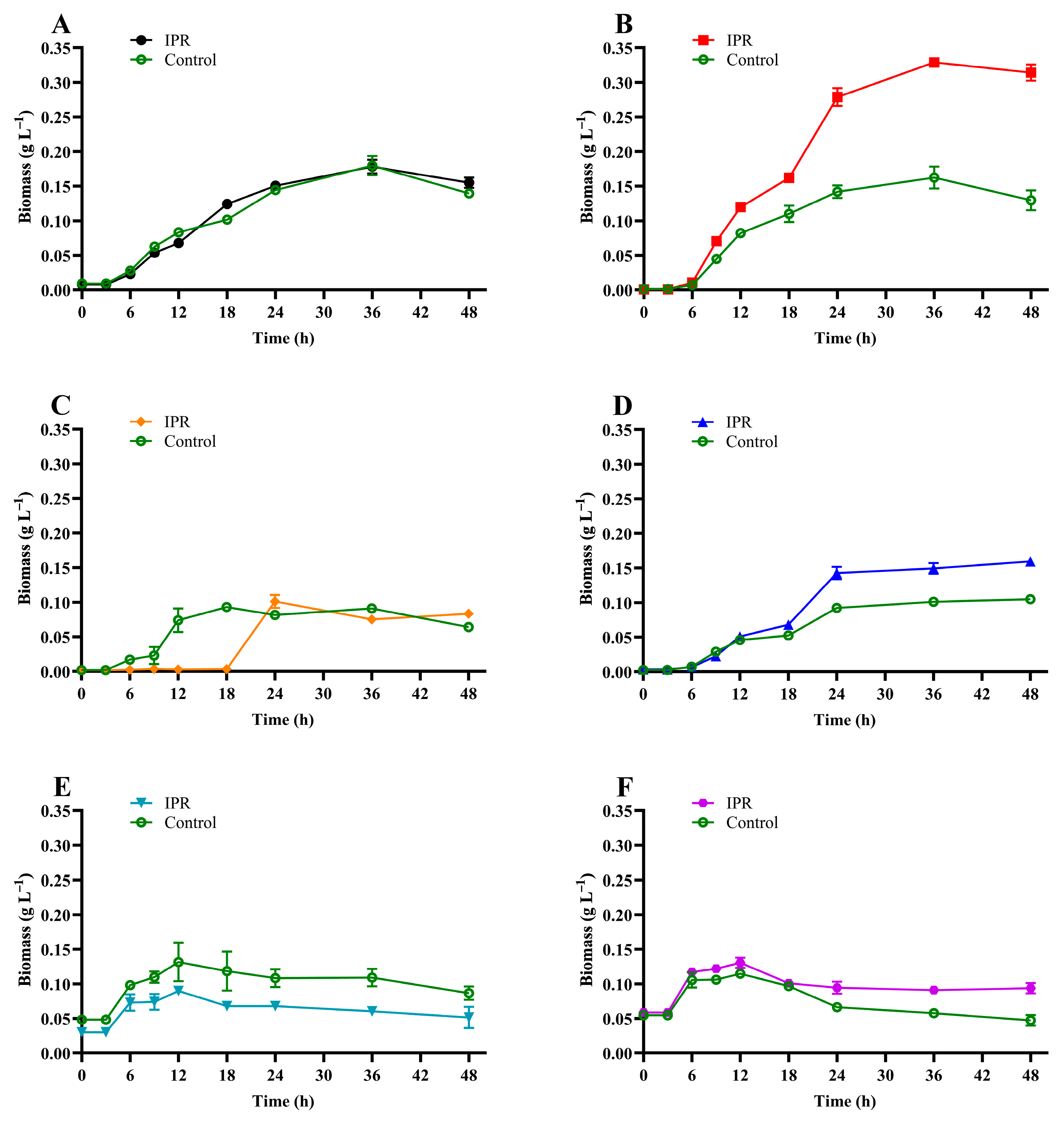
3.3. IPR Removal and Growth of IPR-Tolerant Bacteria
3.4. Comparative Proteomic Study
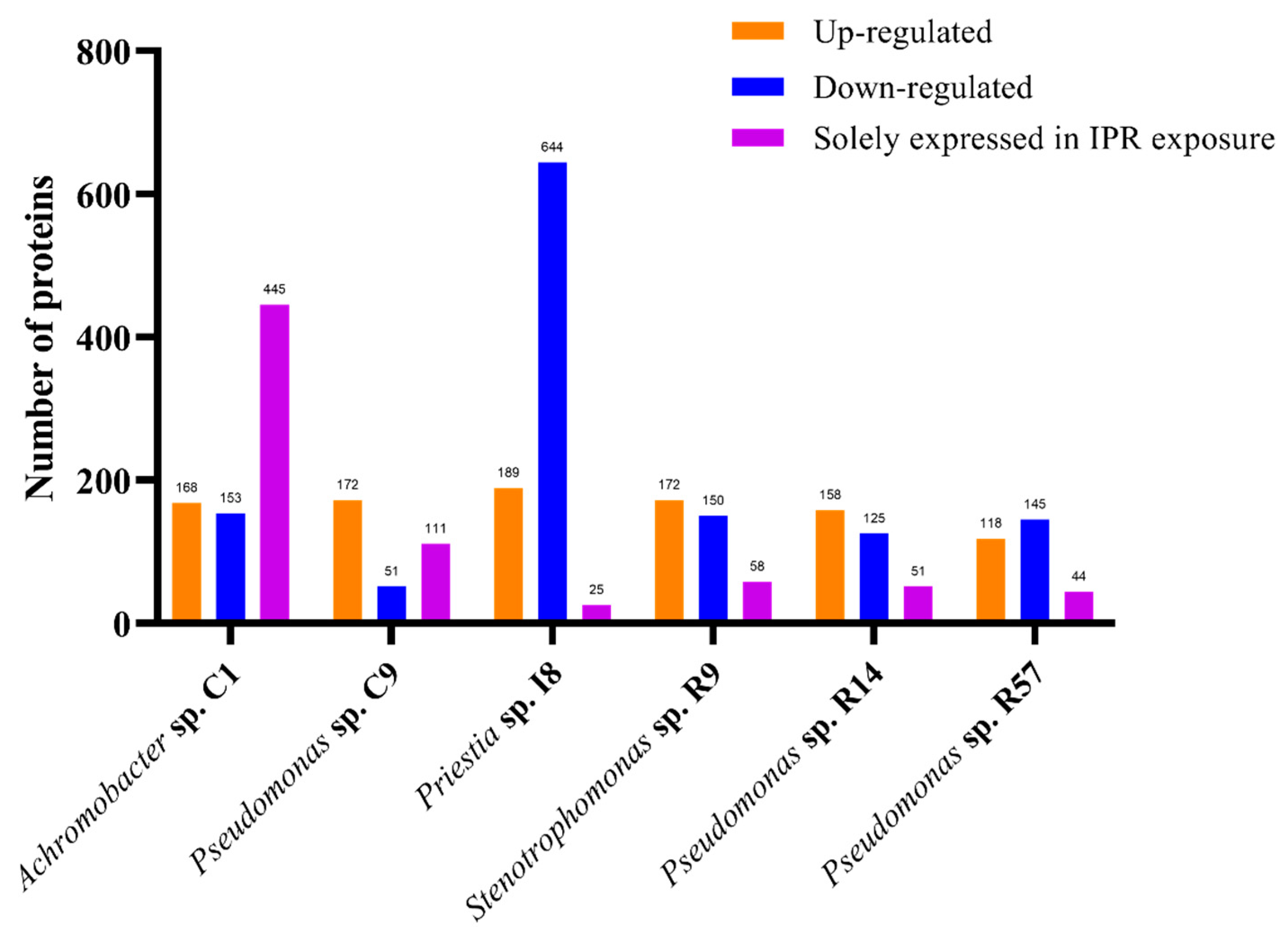
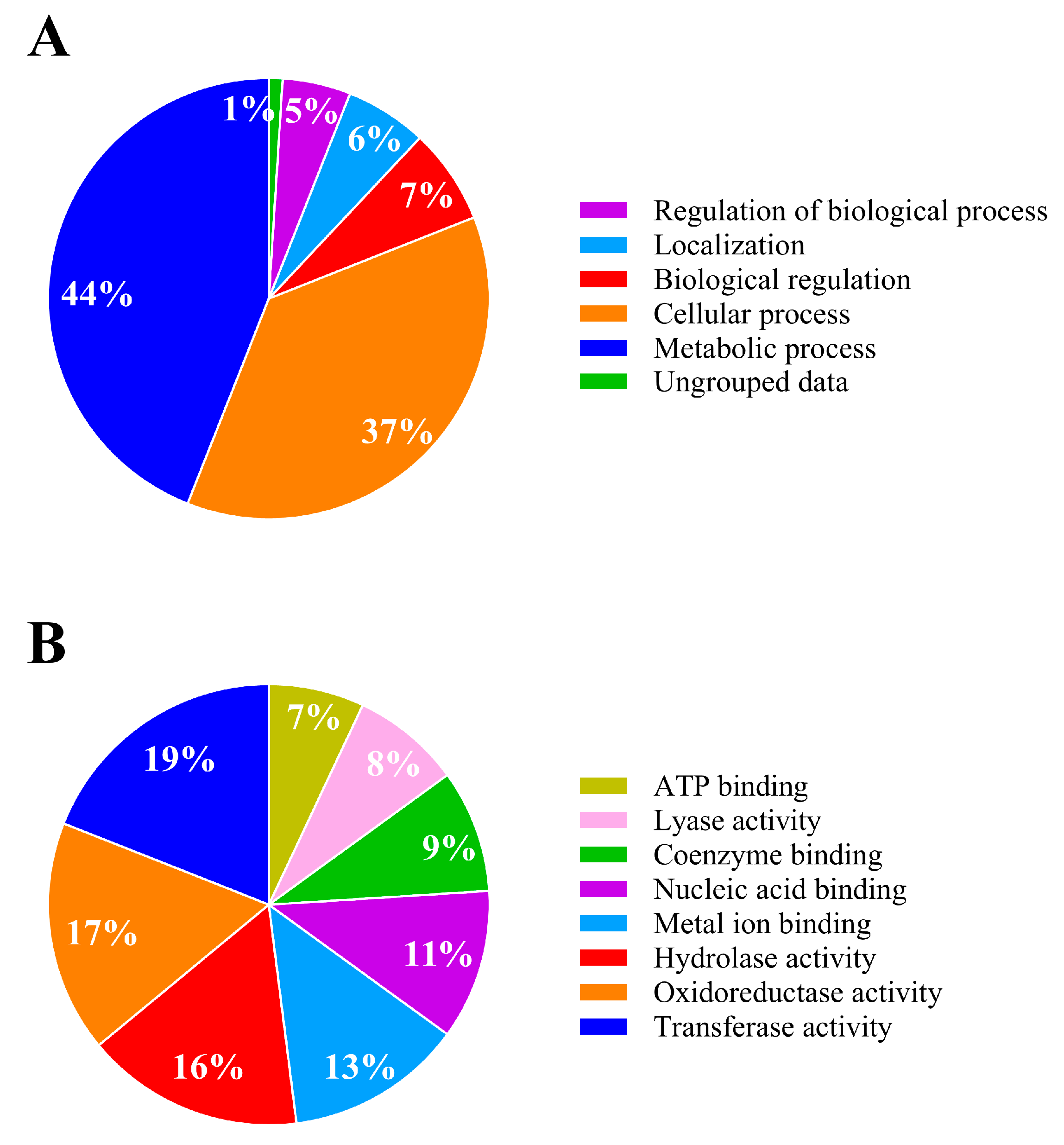
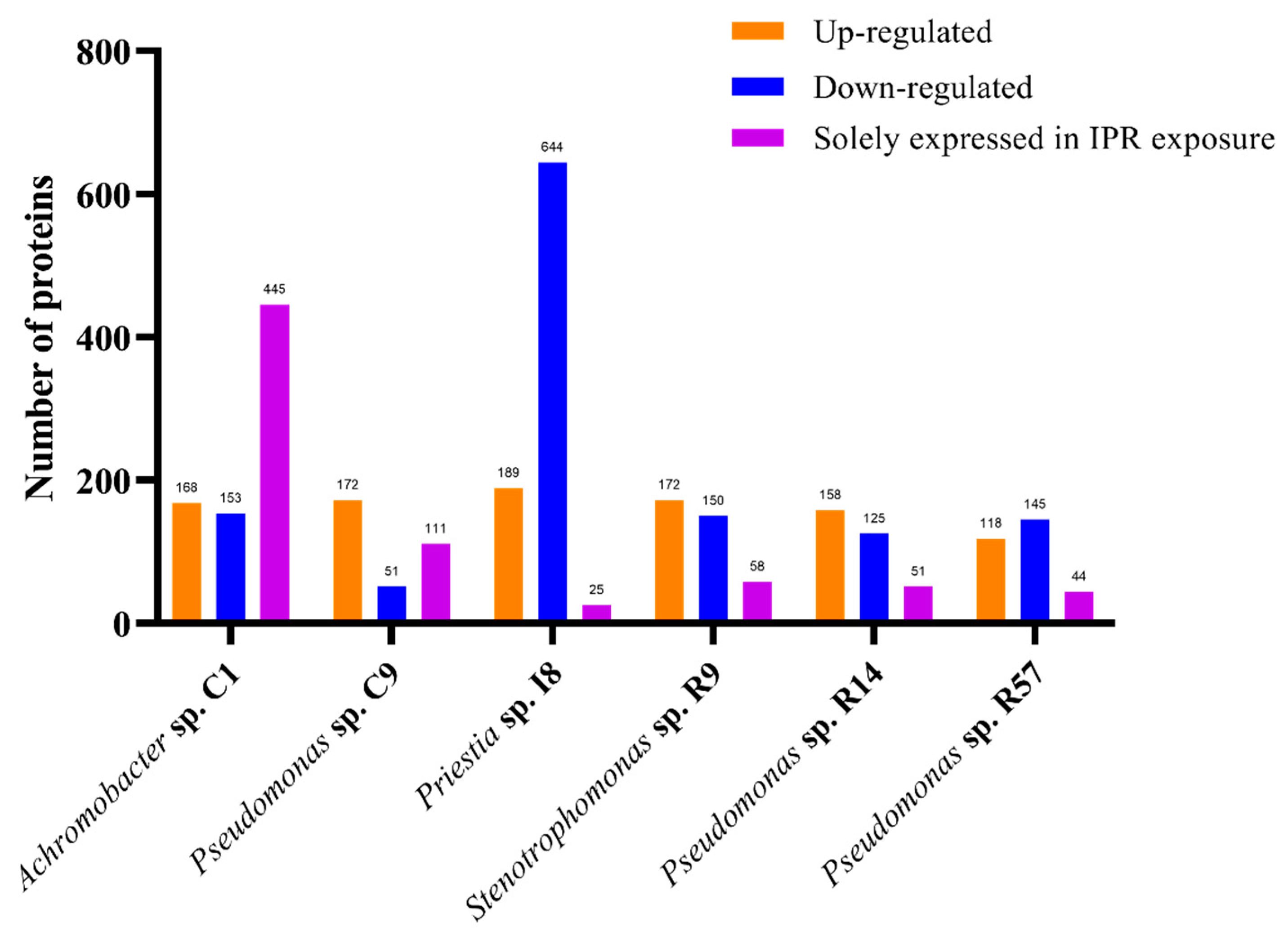
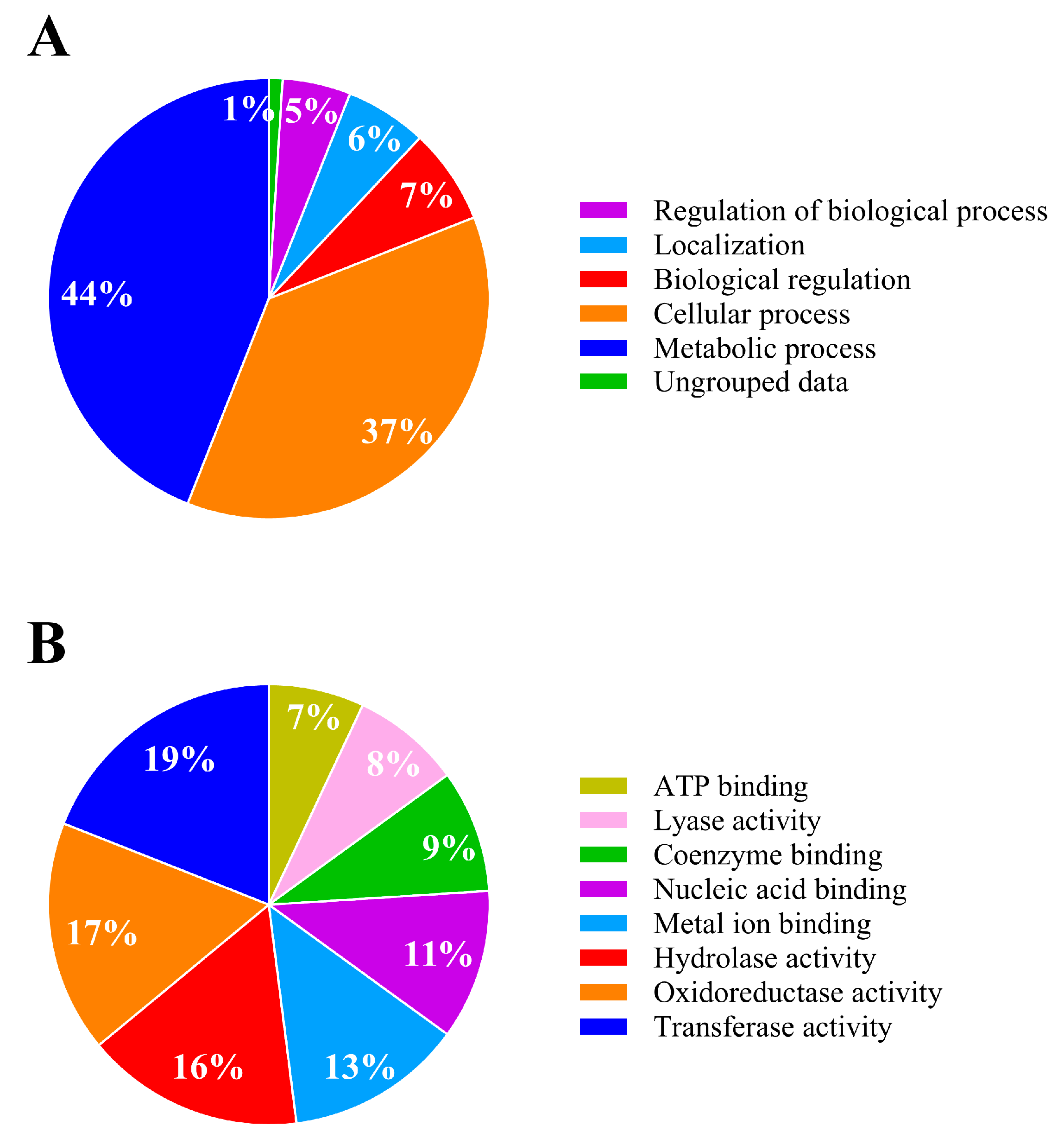
3.4.1. Overall Findings
3.4.2. Analysis of Differentially Expressed Proteins of IPR-Treated and Untreated Achromobacter sp. C1
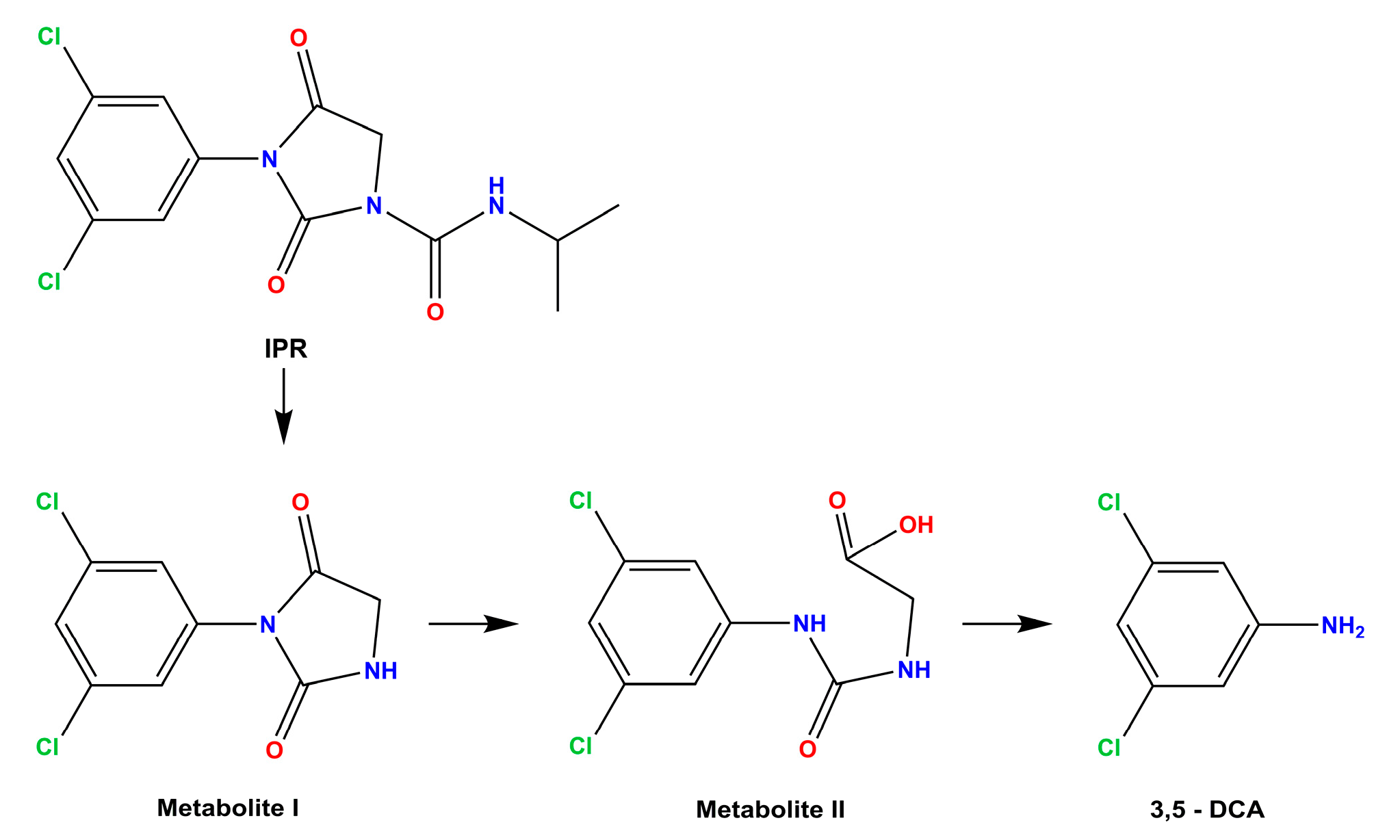
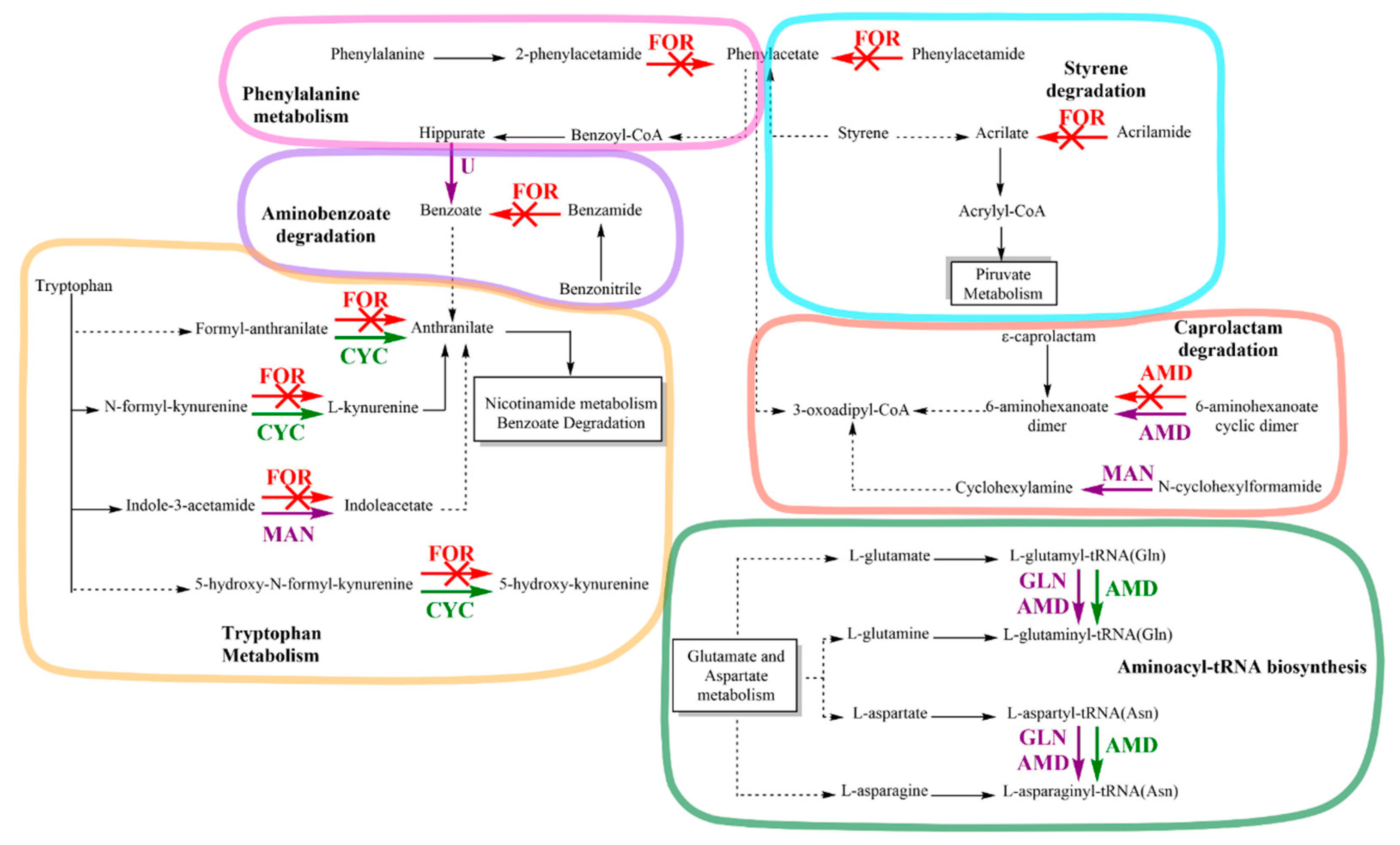
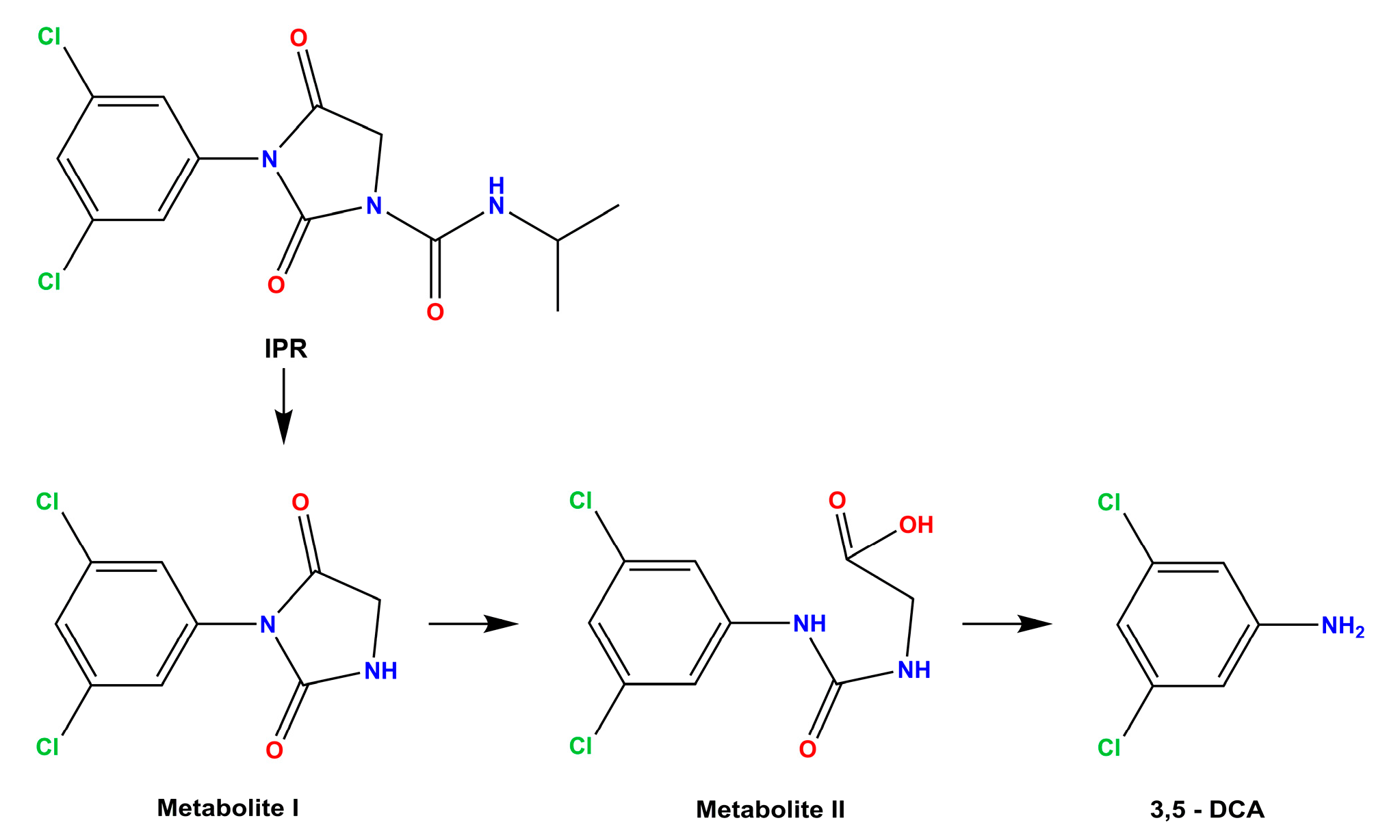
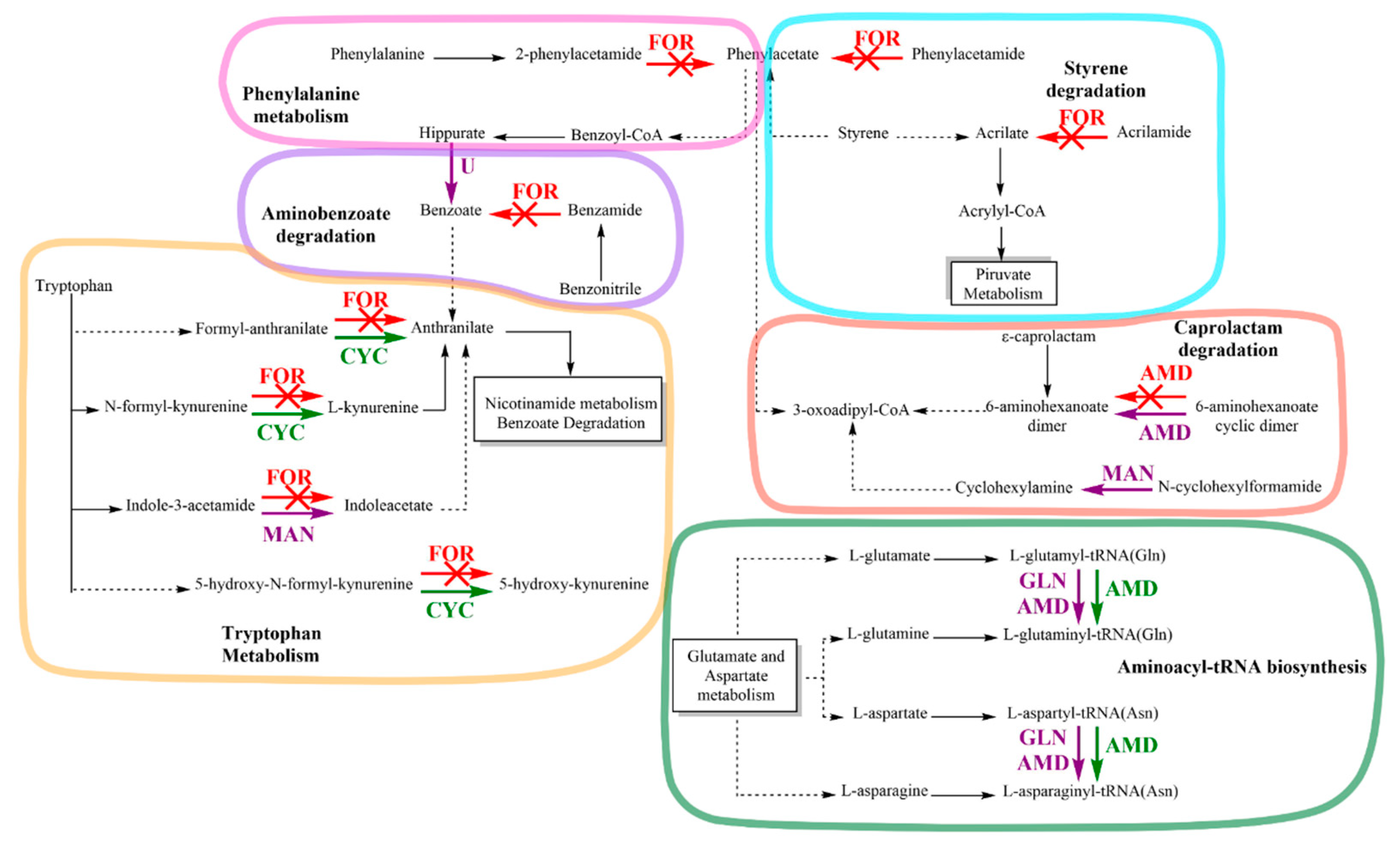
3.5. Amidase Protein Expression of Achromobacter sp. C1 and Effect on Amidase Metabolic Pathways in Response to IPR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tudi, M.; Daniel Ruan, H.; Wang, L.; Lyu, J.; Sadler, R.; Connell, D.; Chu, C.; Phung, D.T. Agriculture development, pesticide application and its impact on the environment. Int. J. Environ. Res. Public Health 2021, 18, 1112. [Google Scholar] [CrossRef] [PubMed]
- Correia, M.; Rodrigues, M.; Paíga, P.; Delerue-Matos, C. Fungicides. In Encyclopedia of Food and Health; Elsevier: Amsterdam, The Netherlands, 2016; pp. 169–176. [Google Scholar]
- Dutra, P.S.S.; Pereira, W.V.; May De Mio, L.L. Brazilian isolates of Monilinia fructicola from peach do not present reduced sensitivity to iprodione. Eur. J. Plant Pathol. 2019, 153, 1341–1346. [Google Scholar] [CrossRef]
- PubChem. Compound Summary Iprodione. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Iprodione (accessed on 14 December 2022).
- Strömqvist, J.; Jarvis, N. Sorption, degradation and leaching of the fungicide iprodione in a golf green under Scandinavian conditions: Measurements, modelling and risk assessment. Pest. Manag. Sci. 2005, 61, 1168–1178. [Google Scholar] [CrossRef] [PubMed]
- Sarker, A.; Lee, S.-H.; Kwak, S.-Y.; Nandi, R.; Kim, J.-E. Comparative catalytic degradation of a metabolite 3,5-dichloroaniline derived from dicarboximide fungicide by laccase and MnO2 mediators. Ecotoxicol. Environ. Saf. 2020, 196, 110561. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Shi, W.; Shu, R.; Pang, J.; Liu, Y.; Zhang, X.; Lei, Y. Isolation and characterization of a bacterium able to degrade high concentrations of iprodione. Can. J. Microbiol. 2018, 64, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, W.; Zhang, Y.; Teng, Y.; Xu, Z. Effects of fungicide iprodione and nitrification inhibitor 3,4-dimethylpyrazole phosphate on soil enzyme and bacterial properties. Sci. Total Environ. 2017, 599–600, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Verdenelli, R.A.; Lamarque, A.L.; Meriles, J.M. Short-term effects of combined iprodione and vermicompost applications on soil microbial community structure. Sci. Total Environ. 2012, 414, 210–219. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Follow Up Assessment of MRLs for the Active Substance Iprodione; EFSA Supporting Publications: Parma, Italy, 2018; p. 15.
- Andrioli, N.B.; Chaufan, G. Dose-independent genotoxic response in A549 cell line exposed to fungicide Iprodione. Arch. Toxicol. 2021, 95, 1071–1079. [Google Scholar] [CrossRef]
- Campos, M.; Perruchon, C.; Vasilieiadis, S.; Menkissoglu-Spiroudi, U.; Karpouzas, D.G.; Diez, M.C. Isolation and characterization of bacteria from acidic pristine soil environment able to transform iprodione and 3,5-dichloraniline. Int. Biodeterior. Biodegrad. 2015, 104, 201–211. [Google Scholar] [CrossRef]
- Campos, M.; Karas, P.S.; Perruchon, C.; Papadopoulou, E.S.; Christou, V.; Menkissoglou-Spiroudi, U.; Diez, M.C.; Karpouzas, D.G. Novel insights into the metabolic pathway of iprodione by soil bacteria. Environ. Sci. Pollut. Res. 2017, 24, 152–163. [Google Scholar] [CrossRef]
- Bergsveinson, J.; Perry, B.J.; Sheedy, C.; Braul, L.; Reedyk, S.; Gossen, B.D.; Yost, C.K. Identifying the core bacterial and fungal communities within four agricultural biobeds used for the treatment of pesticide rinsates. J. Appl. Microbiol. 2018, 125, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Briceño, G.; Lamilla, C.; Leiva, B.; Levio, M.; Donoso-Piñol, P.; Schalchli, H.; Gallardo, F.; Diez, M.C. Pesticide-tolerant bacteria isolated from a biopurification system to remove commonly used pesticides to protect water resources. PLoS ONE 2020, 15, e0234865. [Google Scholar] [CrossRef] [PubMed]
- Góngora-Echeverría, V.R.; García-Escalante, R.; Rojas-Herrera, R.; Giácoman-Vallejos, G.; Ponce-Caballero, C. Pesticide bioremediation in liquid media using a microbial consortium and bacteria-pure strains isolated from a biomixture used in agricultural areas. Ecotoxicol. Environ. Saf. 2020, 200, 110734. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Liu, C.; Zhang, Z.; Zheng, R.; Zheng, Y. Amidase as versatile tool in amide-bond cleavage: From molecular features to biotechnological applications. Biotechnol. Adv. 2020, 43, 107574. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, K.O.; Silva, A.R.M.; da Silva, B.F.; Milagre, H.M.S.; Milagre, C.D.F. Insights into the microbial degradation pathways of the ioxynil octanoate herbicide. Biocatal. Agric. Biotechnol. 2018, 13, 258–264. [Google Scholar] [CrossRef]
- Sun, L.; Gao, X.; Chen, W.; Huang, K.; Bai, N.; Lyu, W.; Liu, H. Characterization of the propham biodegradation pathway in Starkeya sp. strain YW6 and cloning of a novel amidase gene mmH. J. Agric. Food Chem. 2019, 67, 4193–4199. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, Q.; Hang, P.; Zhou, X.; Jiang, J. Characterization of an arylamidase from a newly isolated propanil-transforming strain of Ochrobactrum sp. PP-2. Ecotoxicol. Environ. Saf. 2019, 167, 122–129. [Google Scholar] [CrossRef]
- Chacko, S.; Ramteke, P.W.; Joseph, B. A comparative study on the production of amidase using immobilized and dehydrated immobilized cells of Pseudomonas putida MTCC 6809. J. Genet. Eng. Biotechnol. 2020, 10, 121–127. [Google Scholar] [CrossRef]
- Chen, Q.; Chen, K.; Ni, H.; Zhuang, W.; Wang, H.; Zhu, J.; He, Q.; He, J. A novel amidohydrolase (DmhA) from Sphingomonas sp. that can hydrolyze the organophosphorus pesticide dimethoate to dimethoate carboxylic acid and methylamine. Biotechnol. Lett. 2016, 38, 703–710. [Google Scholar] [CrossRef]
- Yang, Z.; Jiang, W.; Wang, X.; Cheng, T.; Zhang, D.; Wang, H.; Qiu, J.; Cao, L.; Wang, X.; Hong, Q. An amidase gene, ipaH, is responsible for the initial step in the iprodione degradation pathway of Paenarthrobacter sp. strain YJN-5. Appl. Environ. Microbiol. 2018, 84, 1150–1168. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 1991; pp. 115–175. [Google Scholar]
- Turner, S.; Pryer, K.M.; Miao, V.P.W.; Palmer, J.D. Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit rRNA sequence analysis. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Lamilla, C.; Braga, D.; Castro, R.; Guimarães, C.; de Castilho, L.V.A.; Freire, D.M.G.; Barrientos, L. Streptomyces luridus So3.2 from Antarctic soil as a novel producer of compounds with bioemulsification potential. PLoS ONE 2018, 13, e0196054. [Google Scholar] [CrossRef]
- Diez, M.C.; Elgueta, S.; Rubilar, O.; Tortella, G.R.; Schalchli, H.; Bornhardt, C.; Gallardo, F. Pesticide dissipation and microbial community changes in a biopurification system: Influence of the rhizosphere. Biodegradation 2017, 28, 395–412. [Google Scholar] [CrossRef]
- Andrade, M.T.; Neto, D.F.M.; Nascimento, J.R.S.; Soares, E.L.; Coutinho, Í.C.; Velásquez, E.; Domont, G.B.; Nogueira, F.C.S.; Campos, F.A.P. Proteome Dynamics of the Developing Açaí Berry Pericarp (Euterpe oleracea Mart.). J. Proteome Res. 2020, 19, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talon, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef] [PubMed]
- NCBI Resource Coordinators. Database resources of the National Center for Biotechnology Information (NCBI). Nucleic Acids Res. 2016, 44, D7–D19. [Google Scholar] [CrossRef]
- Mercadier, C.; Garcia, D.; Vega, D.; Bastide, J.; Coste, C. Metabolism of iprodione in adapted and non-adapted soils; Effect of soil inoculation with an iprodione-degrading Arthrobacter strain. Soil Biol. Biochem. 1996, 28, 1791–1796. [Google Scholar] [CrossRef]
- Yang, H.; Hu, S.; Wang, X.; Chuang, S.; Jia, W.; Jiang, J. Erratum: Pigmentiphaga sp. Strain D-2 uses a novel amidase to initiate the catabolism of the neonicotinoid insecticide acetamiprid. Appl. Environ. Microbiol. 2020, 86, e02425-19, Erratum in Appl. Environ. Microbiol. 2020, 86, e02425-20. [Google Scholar] [CrossRef]
- Briceño, G.; Fuentes, M.S.; Palma, G.; Jorquera, M.A.; Amoroso, M.J.; Diez, M.C. Chlorpyrifos biodegradation and 3,5,6-trichloro-2-pyridinol production by actinobacteria isolated from soil. Int. Biodeterior. Biodegrad. 2012, 73, 1–7. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, J.Q.; Yuan, Q.Y.; Li, C.; Yan, X.; Hong, Q.; Li, S.P. Characterization of the propanil biodegradation pathway in Sphingomonas sp. Y57 and cloning of the propanil hydrolase gene prpH. J. Hazard. Mater. 2011, 196, 412–419. [Google Scholar] [CrossRef]
- Gupta, R.S.; Patel, S.; Saini, N.; Chen, S. Robust demarcation of 17 distinct Bacillus species clades, proposed as novel Bacillaceae genera, by phylogenomics and comparative genomic analyses: Description of Robertmurraya kyonggiensis sp. nov. and proposal for an emended genus Bacillus limiting it only to the members of the Subtilis and Cereus clades of species. Int. J. Syst. Evol. Microbiol. 2020, 70, 5753–5798. [Google Scholar]
- Deng, S.; Chen, Y.; Wang, D.; Shi, T.; Wu, X.; Ma, X.; Li, X.; Hua, R.; Tang, X.; Li, Q.X. Rapid biodegradation of organophosphorus pesticides by Stenotrophomonas sp: G1. J. Hazard. Mater. 2015, 297, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Le, T.H.; Hoang, Q.C.; Vu, D.D.; Vo, T.H.T. Biodegradation of organophosphorus insecticide methyl parathion by soil microorganisms. E3S Web Conf. 2021, 265, 03002. [Google Scholar] [CrossRef]
- Zhao, J.; Jiang, Y.; Gong, L.; Chen, X.; Xie, Q.; Jin, Y.; Du, J.; Wang, S.; Liu, G. Mechanism of β-cypermethrin metabolism by Bacillus cereus GW-01. Chem. Eng. J. 2022, 430, 132961. [Google Scholar] [CrossRef]
- Mercadier, C.; Vega, D.; Bastide, J. Iprodione degradation by isolated soil microorganisms. FEMS Microbiol. Ecol. 1997, 23, 207–215. [Google Scholar] [CrossRef]
- Zhang, C.; Wu, X.; Wu, Y.; Li, J.; An, H.; Zhang, T. Enhancement of dicarboximide fungicide degradation by two bacterial cocultures of Providencia stuartii JD and Brevundimonas naejangsanensis J3. J. Hazard. Mater. 2021, 403, 123888. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency (USEPA). Iprodione Drinking Water Assessment, Including Proposed New Use on Pistachio (IR4) and Label Revisions Affecting Rates for Strawberries, Stone Fruits, and Grapes; Office of Prevention, Pesticides and Toxic Substances: Washington, DC, USA, 2007.
- Levio-Raiman, M.; Briceño, G.; Leiva, B.; López, S.; Schalchli, H.; Lamilla, C.; Bornhardt, C.; Diez, M.C. Treatment of pesticide-contaminated water using a selected fungal consortium: Study in a batch and packed-bed bioreactor. Agronomy 2021, 11, 743. [Google Scholar] [CrossRef]
- Moreno-Medina, D.A.; Sánchez-Salinas, E.; Ortiz-Hernández, M.L. Removal of methyl parathion and coumaphos pesticides by a bacterial consortium immobilized in luffa cylindrica. Rev. Int. Contam. Ambient. 2014, 30, 51–63. [Google Scholar]
- Silambarasan, S.; Cornejo, P.; Vangnai, A.S. Biodegradation of 4-nitroaniline by novel isolate Bacillus sp. strain AVPP64 in the presence of pesticides. Environ. Pollut. 2022, 306, 119453. [Google Scholar] [CrossRef]
- Tian, J.; Yu, C.; Xue, Y.; Zhao, R.; Wang, J.; Chen, L. Performance of trichlorfon degradation by a novel Bacillus tequilensis strain PA F-3 and its proposed biodegradation pathway. Biodegradation 2016, 27, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Aswathi, A.; Pandey, A.; Madhavan, A.; Sukumaran, R.K. Chlorpyrifos induced proteome remodelling of Pseudomonas nitroreducens AR-3 potentially aid efficient degradation of the pesticide. Environ. Technol. Innov. 2021, 21, 101307. [Google Scholar] [CrossRef]
- Bardot, C.; Besse-Hoggan, P.; Carles, L.; Le Gall, M.; Clary, G.; Chafey, P.; Federici, C.; Broussard, C.; Batisson, I. How the edaphic Bacillus megaterium strain Mes11 adapts its metabolism to the herbicide mesotrione pressure. Environ. Pollut. 2015, 199, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Castrejón-Godínez, M.L.; Tovar-Sanchez, E.; Ortíz-Hernandez, M.L.; Encarnación-Guevara, S.; Martínez-Batallar, A.G.; Hernández-Ortiz, M.; Sánchez-Salina, E.; Rodríguez, A.; Mussali-Galante, P. Proteomic analysis of Burkholderia zhejiangensis CEIB S4–3 during the methyl parathion degradation process. Pestic. Biochem. Phys. 2022, 187, 105197. [Google Scholar] [CrossRef] [PubMed]
- Jurtshuk, P. Bacterial Metabolism. In Medical Microbiology; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. Available online: http://www.ncbi.nlm.nih.gov/pubmed/21413278 (accessed on 14 December 2022).
- Ortiz-Hernández, M.L.; Sánchez-Salinas, E.; Castrejón, M.L.; Dantan, E.; Popoca, E.C. Mechanisms and strategies for pesticide biodegradation: Opportunity for waste, soils and water cleaning. Rev. Int. Contam. Ambient. 2013, 29, 85–104. [Google Scholar]
- Pasquarelli, F.; Spera, A.; Cantarella, L.; Cantarella, M. Biodegradation of bromoxynil using the cascade enzymatic system nitrile hydratase/amidase from Microbacterium imperiale CBS 498-74. Comparison between free enzymes and resting cells. RSC Adv. 2015, 5, 36913–36923. [Google Scholar] [CrossRef]
- Chang, A.; Jeske, L.; Ulbrich, S.; Hofmann, J.; Koblitz, J.; Schomburg, I.; Neumann-Schaal, M.; Jahn, D.; Schomburg, D. BRENDA, the ELIXIR core data resource in 2021: New developments and updates. Nucleic Acids Res. 2021, 49, D498–D508. [Google Scholar] [CrossRef] [PubMed]
- Kurnasov, O.; Jablonski, L.; Polanuyer, B.; Dorrestein, P.; Begley, T.; Osterman, A. Aerobic tryptophan degradation pathway in bacteria: Novel kynurenine formamidase. FEMS Microbiol. Lett. 2023, 227, 219–227. [Google Scholar] [CrossRef]
- Zhang, M.; Li, Y.; Mu, Q.; Feng, F.; Yu, X.; Ge, Y.; Zhang, Y.; Nie, J. Effects of chlorpyrifos on the metabolic profiling of Bacillus megaterium strain RRB. Chemosphere 2022, 297, 134189. [Google Scholar] [CrossRef]
- Pankaj; Negi, G.; Gangola, S.; Khati, P.; Kumar, G.; Srivastava, A.; Sharma, A. Differential expression and characterization of cypermethrin-degrading potential proteins in Bacillus thuringiensis strain, SG4. 3 Biotech. 2016, 6, 225. [Google Scholar] [CrossRef]
- Hodgson, E. Biotransformation (Metabolism) of pesticides. In Pesticide Biotransformation and Disposition; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Zhao, Y.X.; Guo, L.; Wang, L.; Jiang, N.-D.; Chen, K.-C.; Dai, Y.-J. Biodegradation of the pyridinecarboxamide insecticide flonicamid by Microvirga flocculans and characterization of two novel amidases involved. Ecotoxicol. Environ. Saf. 2021, 220, 112384. [Google Scholar] [CrossRef] [PubMed]
- InterPro. Classification of Protein Families, Amidase Signature Domain. Rev. January 2023. Available online: http://www.ebi.ac.uk/interpro/entry/InterPro/IPR023631/ (accessed on 14 January 2023).
- Singh, S.; Kumar, V.; Chauhan, A.; Datta, S.; Wani, A.B.; Singh, N.; Singh, J. Toxicity, degradation and analysis of the herbicide atrazine. Environ. Chem. Lett. 2018, 16, 211–237. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | C1 | C9 | I8 | R9 | R14 | R57 |
|---|---|---|---|---|---|---|
| Regression equation | −6.001x + 51.809 | −5.623x + 52.203 | −4.895x + 49.670 | −5.482x + 52.288 | −5.831x + 50.658 | −5.908x+ 51.450 |
| K (h−1) | 0.076 | 0.073 | 0.047 | 0.060 | 0.067 | 0.073 |
| T1/2 (h−1) | 9.1 | 9.5 | 14.6 | 11.5 | 10.3 | 9.5 |
| R2 | 0.960 | 0.948 | 0.982 | 0.962 | 0.955 | 0.941 |
| Specific growth rate (µ) | 0.140 | 0.092 | 0.299 | 0.123 | 0.034 | 0.018 |
| Biomass duplication time (h) | 4.94 | 7.55 | 2.32 | 5.63 | 20.24 | 39.12 |
| Accession Number | Protein Name | Fold | Function |
|---|---|---|---|
| Upregulated proteins | |||
| A0A3Q9KM03 A0A3Q9KKP8 | Aspartyl/glutamyl-tRNA(Asn/Gln) amidotransferase subunit C Amidase | 3.6 1.9 | asparaginyl-tRNA synthase (glutamine-hydrolyzing) activity, glutaminyl-tRNA synthase (glutamine-hydrolyzing) activity, transferase activity, ligase activity, ATP binding, nucleotide binding Amidase and indoleacetamide hydrolase activity |
| A0A2S0I6B2 | Cyclase | 1.6 | Arylformamidase activity |
| Downregulated proteins | |||
| A0A2S0I9W4 | Formamidase | 2.6 | Hydrolase activity, acting on carbon–nitrogen (but not peptide) bonds, in linear amides |
| A0A2S5GVY3 | Amidase | 2.7 | Hydrolase activity |
| Expressed solely on IPR treatment proteins | |||
| A0A3S4NGC5 | Glutamyl-tRNA(Gln) Amidotransferase subunit A | Unique | ATP binding, glutaminyl-tRNA synthase (glutaminehydrolyzing) activity, hydrolase activity and transferase activity |
| A0A3S4PMM6 | Glutamyl-tRNA(Gln) amidotransferase subunit A | Unique | Hydrolase activity, ligase activity and transferase activity |
| A0A2S5GXA3 | Glutamyl-tRNA(Gln) amidotransferase subunit A | Unique | ATP binding, glutaminyl-tRNA synthase (glutaminehydrolyzing) activity, hydrolase activity and transferase activity |
| A0A3Q9KJB0 | Amidase | Unique | Hydrolase activity |
| A0A2S5GNM1 | Amidase | Unique | Hydrolase activity |
| A0A448CBV6 | Uncharacterized hydrolase YxeP | Unique | Hydrolase, amidohydrolase activity |
| A0A3S4Q6N4 | Uncharacterized hydrolase YxeP | Unique | Hydrolase, amidohydrolase activity |
| A0A2S5GW20 | N-ethylammeline chlorohydrolase | Unique | Hydrolase activity, amidohydrolase-related |
| A0A2S5GLS7 | N-formylglutamate deformylase | Unique | N-formylglutamate deformylase, n-formylglutamate amidohydrolase activity |
| A0A2S0I4Y1 | N-formylglutamate amidohydrolase | Unique | Hydrolase, N-formylglutamate amidohydrolase activity |
| A0A448C6N6 | Mandelamide hydrolase | Unique | Mandelamide amidase activity |
| A0A3S9YT14 | Amidase | Unique | Hydrolase activity |
| A0A3S9YSH8 | Amidohydrolase | Unique | Hydrolase, amidohydrolase activity |
| A0A2S5GNK0 | Amidohydrolase | Unique | Hydrolase, amidohydrolase activity |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donoso-Piñol, P.; Briceño, G.; Evaristo, J.A.M.; Nogueira, F.C.S.; Leiva, B.; Lamilla, C.; Schalchli, H.; Diez, M.C. Metabolic Profiling and Comparative Proteomic Insight in Respect of Amidases during Iprodione Biodegradation. Microorganisms 2023, 11, 2367. https://doi.org/10.3390/microorganisms11102367
Donoso-Piñol P, Briceño G, Evaristo JAM, Nogueira FCS, Leiva B, Lamilla C, Schalchli H, Diez MC. Metabolic Profiling and Comparative Proteomic Insight in Respect of Amidases during Iprodione Biodegradation. Microorganisms. 2023; 11(10):2367. https://doi.org/10.3390/microorganisms11102367
Chicago/Turabian StyleDonoso-Piñol, Pamela, Gabriela Briceño, Joseph A. M. Evaristo, Fábio C. S. Nogueira, Barbara Leiva, Claudio Lamilla, Heidi Schalchli, and María Cristina Diez. 2023. "Metabolic Profiling and Comparative Proteomic Insight in Respect of Amidases during Iprodione Biodegradation" Microorganisms 11, no. 10: 2367. https://doi.org/10.3390/microorganisms11102367
APA StyleDonoso-Piñol, P., Briceño, G., Evaristo, J. A. M., Nogueira, F. C. S., Leiva, B., Lamilla, C., Schalchli, H., & Diez, M. C. (2023). Metabolic Profiling and Comparative Proteomic Insight in Respect of Amidases during Iprodione Biodegradation. Microorganisms, 11(10), 2367. https://doi.org/10.3390/microorganisms11102367