
Ophthalmic Solutions with a Broad Antiviral Action: Evaluation of Their Potential against Ocular Herpetic Infections

 , , ,
, , ,  ,
,  ,
,  , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Compounds
2.2. Cell and Virus Culture
2.3. Antiviral Activity
2.4. Real-Time PCR
2.5. Statistical Analysis
3. Results and Discussion
3.1. Antiviral Activity against HSV-1
3.2. Real-Time PCR
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization (WHO). Available online: https://www.who.int/news-room/fact-sheets/detail/herpes-simplex-virus (accessed on 1 June 2022).
- Duarte, L.F.; Farias, M.A.; Alvarez, D.M.; Bueno, S.M.; Riedel, C.A.; Gonzalez, P.A. Herpes Simplex Virus Type 1 Infection of the Central Nervous System: Insights into Proposed Interrelationships With Neurodegenerative Disorders. Front. Cell. Neurosci. 2019, 13, 46. [Google Scholar] [CrossRef] [PubMed]
- Nicoll, M.P.; Proenca, J.T.; Efstathiou, S. The molecular basis of herpes simplex virus latency. FEMS Microbiol. Rev. 2012, 36, 684–705. [Google Scholar] [CrossRef]
- Cohen, J.I. Herpesvirus latency. J. Clin. Investig. 2020, 130, 3361–3369. [Google Scholar] [CrossRef] [PubMed]
- Koyuncu, O.O.; Hogue, I.B.; Enquist, L.W. Virus infections in the nervous system. Cell Host Microbe 2013, 13, 379–393. [Google Scholar] [CrossRef] [PubMed]
- Kimberlin, D.W. Neonatal herpes simplex infection. Clin. Microbiol. Rev. 2004, 17, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Knezevic, A.; Martic, J.; Stanojevic, M.; Jankovic, S.; Nedeljkovic, J.; Nikolic, L.; Pasic, S.; Jankovic, B.; Jovanovic, T. Disseminated neonatal herpes caused by herpes simplex virus types 1 and 2. Emerg. Infect. Dis. 2007, 13, 302–304. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, M.J.; Venkatesan, A. Herpes Simplex Virus-1 Encephalitis in Adults: Pathophysiology, Diagnosis, and Management. Neurotherapeutics 2016, 13, 493–508. [Google Scholar] [CrossRef]
- Costa, B.K.D.; Sato, D.K. Viral encephalitis: A practical review on diagnostic approach and treatment. J. Pediatr. 2020, 96 (Suppl. S1), 12–19. [Google Scholar] [CrossRef]
- Kaye, S.; Choudhary, A. Herpes simplex keratitis. Prog. Retin. Eye Res. 2006, 25, 355–380. [Google Scholar] [CrossRef]
- Rowe, A.M.; St Leger, A.J.; Jeon, S.; Dhaliwal, D.K.; Knickelbein, J.E.; Hendricks, R.L. Herpes keratitis. Prog. Retin. Eye Res. 2013, 32, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.D. Herpes Simplex Virus Keratitis. Home Healthc. Now 2019, 37, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Sibley, D.; Larkin, D.F.P. Update on Herpes simplex keratitis management. Eye 2020, 34, 2219–2226. [Google Scholar] [CrossRef] [PubMed]
- Glick, J.E.; Bar, J. Herpes Simplex Virus-Associated Keratitis. J. Emerg. Med. 2021, 61, e11–e12. [Google Scholar] [CrossRef]
- Al-Dujaili, L.J.; Clerkin, P.P.; Clement, C.; McFerrin, H.E.; Bhattacharjee, P.S.; Varnell, E.D.; Kaufman, H.E.; Hill, J.M. Ocular herpes simplex virus: How are latency, reactivation, recurrent disease and therapy interrelated? Future Microbiol. 2011, 6, 877–907. [Google Scholar] [CrossRef]
- Reddy, J.C.; Rapuano, C.J. Current Concepts in the Management of Herpes Simplex Anterior Segment Eye Disease. Curr. Ophthalmol. Rep. 2013, 1, 194–203. [Google Scholar] [CrossRef][Green Version]
- Wang, L.; Wang, R.; Xu, C.; Zhou, H. Pathogenesis of Herpes Stromal Keratitis: Immune Inflammatory Response Mediated by Inflammatory Regulators. Front. Immunol. 2020, 11, 766. [Google Scholar] [CrossRef] [PubMed]
- Gimenez, F.; Suryawanshi, A.; Rouse, B.T. Pathogenesis of herpes stromal keratitis--a focus on corneal neovascularization. Prog. Retin. Eye Res. 2013, 33, 1–9. [Google Scholar] [CrossRef]
- Lobo, A.M.; Agelidis, A.M.; Shukla, D. Pathogenesis of herpes simplex keratitis: The host cell response and ocular surface sequelae to infection and inflammation. Ocul. Surf. 2019, 17, 40–49. [Google Scholar] [CrossRef]
- Tsatsos, M.; MacGregor, C.; Athanasiadis, I.; Moschos, M.M.; Hossain, P.; Anderson, D. Herpes simplex virus keratitis: An update of the pathogenesis and current treatment with oral and topical antiviral agents. Clin. Exp. Ophthalmol. 2016, 44, 824–837. [Google Scholar] [CrossRef]
- Chodosh, J.; Ung, L. Adoption of Innovation in Herpes Simplex Virus Keratitis. Cornea 2020, 39 (Suppl. S1), S7–S18. [Google Scholar] [CrossRef]
- Wilhelmus, K.R. Diagnosis and management of herpes simplex stromal keratitis. Cornea 1987, 6, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Morfin, F.; Thouvenot, D. Herpes simplex virus resistance to antiviral drugs. J. Clin. Virol. 2003, 26, 29–37. [Google Scholar] [CrossRef]
- Bacon, T.H.; Levin, M.J.; Leary, J.J.; Sarisky, R.T.; Sutton, D. Herpes simplex virus resistance to acyclovir and penciclovir after two decades of antiviral therapy. Clin. Microbiol. Rev. 2003, 16, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Piret, J.; Boivin, G. Antiviral resistance in herpes simplex virus and varicella-zoster virus infections: Diagnosis and management. Curr. Opin. Infect. Dis. 2016, 29, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, A.; Pharm, S.B.; Gueudry, J.; Deback, C.; Haigh, O.; Schweitzer, C.; Boutolleau, D.; Labetoulle, M. Acyclovir-Resistant Herpes Simplex Virus 1 Keratitis: A Concerning and Emerging Clinical Challenge. Am. J. Ophthalmol. 2022, 238, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Majewska, A.; Mlynarczyk-Bonikowska, B. 40 Years after the Registration of Acyclovir: Do We Need New Anti-Herpetic Drugs? Int. J. Mol. Sci. 2022, 23, 3431. [Google Scholar] [CrossRef]
- Koerner, J.C.; George, M.J.; Kissam, E.A.; Rosco, M.G. Povidone-iodine concentration and in vitro killing time of bacterial corneal ulcer isolates. Digit. J. Ophthalmol. 2018, 24, 24–26. [Google Scholar] [CrossRef]
- Singh, S.; Sawant, O.B.; Mian, S.I.; Kumar, A. Povidone-Iodine Attenuates Viral Replication in Ocular Cells: Implications for Ocular Transmission of RNA Viruses. Biomolecules 2021, 11, 753. [Google Scholar] [CrossRef]
- Musumeci, R.; Bandello, F.; Martinelli, M.; Calaresu, E.; Cocuzza, C.E. In vitro bactericidal activity of 0.6% povidone-iodine eye drops formulation. Eur. J. Ophthalmol. 2019, 29, 673–677. [Google Scholar] [CrossRef]
- Paduch, R.; Urbanik-Sypniewska, T.; Kutkowska, J.; Choragiewicz, T.; Matysik-Wozniak, A.; Zweifel, S.; Czarnek-Chudzik, A.; Zaluska, W.; Rejdak, R.; Toro, M.D. Ozone-Based Eye Drops Activity on Ocular Epithelial Cells and Potential Pathogens Infecting the Front of the Eye. Antioxidants 2021, 10, 968. [Google Scholar] [CrossRef]
- Caruso, C.; Eletto, D.; Tosco, A.; Pannetta, M.; Scarinci, F.; Troisi, M.; Porta, A. Comparative Evaluation of Antimicrobial, Antiamoebic, and Antiviral Efficacy of Ophthalmic Formulations. Microorganisms 2022, 10, 1156. [Google Scholar] [CrossRef]
- Petrillo, F.; Chianese, A.; De Bernardo, M.; Zannella, C.; Galdiero, M.; Reibaldi, M.; Avitabile, T.; Boccia, G.; Galdiero, M.; Rosa, N.; et al. Inhibitory Effect of Ophthalmic Solutions against SARS-CoV-2: A Preventive Action to Block the Viral Transmission? Microorganisms 2021, 9, 1550. [Google Scholar] [CrossRef]
- Zannella, C.; Chianese, A.; Palomba, L.; Marcocci, M.E.; Bellavita, R.; Merlino, F.; Grieco, P.; Folliero, V.; De Filippis, A.; Mangoni, M.; et al. Broad-Spectrum Antiviral Activity of the Amphibian Antimicrobial Peptide Temporin L and Its Analogs. Int. J. Mol. Sci. 2022, 23, 2060. [Google Scholar] [CrossRef] [PubMed]
- Falanga, A.; Del Genio, V.; Kaufman, E.A.; Zannella, C.; Franci, G.; Weck, M.; Galdiero, S. Engineering of Janus-Like Dendrimers with Peptides Derived from Glycoproteins of Herpes Simplex Virus Type 1: Toward a Versatile and Novel Antiviral Platform. Int. J. Mol. Sci. 2021, 22, 6488. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Lyu, Y.N.; Xin, H.Y.; Cheng, J.T.; Liu, X.Q.; Wang, X.W.; Peng, X.C.; Xiang, Y.; Xin, V.W.; Lu, C.B.; et al. Identification of Putative UL54 (ICP27) Transcription Regulatory Sequences Binding to Oct-1, v-Myb, Pax-6 and Hairy in Herpes Simplex Viruses. J. Cancer 2019, 10, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Lalli, J.; Sedlackova-Slavikova, L.; Rice, S.A. Functional comparison of herpes simplex virus 1 (HSV-1) and HSV-2 ICP27 homologs reveals a role for ICP27 in virion release. J. Virol. 2015, 89, 2892–2905. [Google Scholar] [CrossRef] [PubMed]
- Muylaert, I.; Zhao, Z.; Elias, P. UL52 primase interactions in the herpes simplex virus 1 helicase-primase are affected by antiviral compounds and mutations causing drug resistance. J. Biol. Chem. 2014, 289, 32583–32592. [Google Scholar] [CrossRef]
- Bermek, O.; Williams, R.S. The three-component helicase/primase complex of herpes simplex virus-1. Open Biol. 2021, 11, 210011. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Xin, V.W.; Liu, X.Q.; Wang, Y.Y.; Zhang, Y.; Cheng, J.T.; Cai, W.Q.; Xiang, Y.; Peng, X.C.; Wang, X.; et al. Identification of 22 Novel Motifs of the Cell Entry Fusion Glycoprotein B of Oncolytic Herpes Simplex Viruses: Sequence Analysis and Literature Review. Front. Oncol. 2020, 10, 1386. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.; Kopp, S.J.; Connolly, S.A.; Longnecker, R. Structure-Based Mutations in the Herpes Simplex Virus 1 Glycoprotein B Ectodomain Arm Impart a Slow-Entry Phenotype. mBio 2017, 8, e00614-17. [Google Scholar] [CrossRef]
- Kosovsky, J.; Vojvodova, A.; Oravcova, I.; Kudelova, M.; Matis, J.; Rajcani, J. Herpes simplex virus 1 (HSV-1) strain HSZP glycoprotein B gene: Comparison of mutations among strains differing in virulence. Virus Genes 2000, 20, 27–33. [Google Scholar] [CrossRef]
- Bagga, B.; Kate, A.; Joseph, J.; Dave, V.P. Herpes simplex infection of the eye: An introduction. Community Eye Health 2020, 33, 68–70. [Google Scholar]
- Petrillo, F.; Pignataro, D.; Lavano, M.; Santella, B.; Folliero, V.; Zannella, C.; Astarita, C.; Gagliano, C.; Franci, G.; Avitabile, T.; et al. Current Evidence on the Ocular Surface Microbiota and Related Diseases. Microorganisms 2020, 8, 1033. [Google Scholar] [CrossRef]
- Franci, G.; Falanga, A.; Zannella, C.; Folliero, V.; Martora, F.; Galdiero, M.; Galdiero, S.; Morelli, G.; Galdiero, M. Infectivity inhibition by overlapping synthetic peptides derived from the gH/gL heterodimer of herpes simplex virus type 1. J. Pept. Sci. 2017, 23, 311–319. [Google Scholar] [CrossRef]
- Lim, N.A.; Teng, O.; Ng, C.Y.H.; Bao, L.X.Y.; Tambyah, P.A.; Quek, A.M.L.; Seet, R.C.S. Repurposing povidone-iodine to reduce the risk of SARS-CoV-2 infection and transmission: A narrative review. Ann. Med. 2022, 54, 1488–1499. [Google Scholar] [CrossRef] [PubMed]
- Shet, M.; Westover, J.; Hong, R.; Igo, D.; Cataldo, M.; Bhaskar, S. In vitro inactivation of SARS-CoV-2 using a povidone-iodine oral rinse. BMC Oral Health 2022, 22, 47. [Google Scholar] [CrossRef] [PubMed]
- Kawana, R.; Kitamura, T.; Nakagomi, O.; Matsumoto, I.; Arita, M.; Yoshihara, N.; Yanagi, K.; Yamada, A.; Morita, O.; Yoshida, Y.; et al. Inactivation of human viruses by povidone-iodine in comparison with other antiseptics. Dermatology 1997, 195 (Suppl. S2), 29–35. [Google Scholar] [CrossRef] [PubMed]
- Sabracos, L.; Romanou, S.; Dontas, I.; Coulocheri, S.; Ploumidou, K.; Perrea, D. The in vitro effective antiviral action of povidone-iodine (PVP-I) may also have therapeutic potential by its intravenous administration diluted with Ringer’s solution. Med. Hypotheses 2007, 68, 272–274. [Google Scholar] [CrossRef]
- Eggers, M.; Koburger-Janssen, T.; Eickmann, M.; Zorn, J. In Vitro Bactericidal and Virucidal Efficacy of Povidone-Iodine Gargle/Mouthwash Against Respiratory and Oral Tract Pathogens. Infect. Dis. Ther. 2018, 7, 249–259. [Google Scholar] [CrossRef]
- Sriwilaijaroen, N.; Wilairat, P.; Hiramatsu, H.; Takahashi, T.; Suzuki, T.; Ito, M.; Ito, Y.; Tashiro, M.; Suzuki, Y. Mechanisms of the action of povidone-iodine against human and avian influenza A viruses: Its effects on hemagglutination and sialidase activities. Virol. J. 2009, 6, 124. [Google Scholar] [CrossRef]
- Hosseini, H.; Ashraf, M.J.; Saleh, M.; Nowroozzadeh, M.H.; Nowroozizadeh, B.; Abtahi, M.B.; Nowroozizadeh, S. Effect of povidone-iodine concentration and exposure time on bacteria isolated from endophthalmitis cases. J. Cataract. Refract. Surg. 2012, 38, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Sauerbrei, A. Bactericidal and virucidal activity of ethanol and povidone-iodine. Microbiologyopen 2020, 9, e1097. [Google Scholar] [CrossRef] [PubMed]
- Chesnay, A.; Bailly, E.; Desoubeaux, G. Demonstration of the yeasticidal efficacy of povidone-iodine-based commercial antiseptic solutions against Candida auris. J. Mycol. Med. 2021, 31, 101173. [Google Scholar] [CrossRef] [PubMed]
- Kredics, L.; Narendran, V.; Shobana, C.S.; Vagvolgyi, C.; Manikandan, P.; Indo-Hungarian Fungal Keratitis Working, G. Filamentous fungal infections of the cornea: A global overview of epidemiology and drug sensitivity. Mycoses 2015, 58, 243–260. [Google Scholar] [CrossRef] [PubMed]
- Ong, H.S.; Fung, S.S.M.; Macleod, D.; Dart, J.K.G.; Tuft, S.J.; Burton, M.J. Altered Patterns of Fungal Keratitis at a London Ophthalmic Referral Hospital: An Eight-Year Retrospective Observational Study. Am. J. Ophthalmol. 2016, 168, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Stelitano, D.; Franci, G.; Chianese, A.; Galdiero, S.; Morelli, G.; Galdiero, M. HSV membrane glycoproteins, their function in viral entry and their use in vaccine studies. In Amino Acids, Peptides and Proteins: Volume 43; The Royal Society of Chemistry: London, UK, 2019; Volume 43, pp. 14–43. [Google Scholar]
- Franci, G.; Crudele, V.; Della Rocca, M.T.; Melardo, C.; Chianese, A.; Finamore, E.; Bencivenga, F.; Astorri, R.; Vitiello, M.; Galdiero, E.; et al. Epstein-Barr Virus Seroprevalence and Primary Infection at the University Hospital Luigi Vanvitelli of Naples from 2007 to 2017. Intervirology 2019, 62, 15–22. [Google Scholar] [CrossRef]
- Spadea, L.; Tonti, E.; Spaterna, A.; Marchegiani, A. Use of Ozone-Based Eye Drops: A Series of Cases in Veterinary and Human Spontaneous Ocular Pathologies. Case Rep. Ophthalmol. 2018, 9, 287–298. [Google Scholar] [CrossRef]
- Elvis, A.M.; Ekta, J.S. Ozone therapy: A clinical review. J. Nat. Sci. Biol. Med. 2011, 2, 66–70. [Google Scholar] [CrossRef]
- Sechi, L.A.; Lezcano, I.; Nunez, N.; Espim, M.; Dupre, I.; Pinna, A.; Molicotti, P.; Fadda, G.; Zanetti, S. Antibacterial activity of ozonized sunflower oil (Oleozon). J. Appl. Microbiol. 2001, 90, 279–284. [Google Scholar] [CrossRef]
- Santella, B.; Folliero, V.; Pirofalo, G.M.; Serretiello, E.; Zannella, C.; Moccia, G.; Santoro, E.; Sanna, G.; Motta, O.; De Caro, F.; et al. Sepsis-A Retrospective Cohort Study of Bloodstream Infections. Antibiotics 2020, 9, 851. [Google Scholar] [CrossRef]
- Pinna, A.; Donadu, M.G.; Usai, D.; Dore, S.; D’Amico-Ricci, G.; Boscia, F.; Zanetti, S. In vitro antimicrobial activity of a new ophthalmic solution containing povidone-iodine 0.6% (IODIM((R))). Acta Ophthalmol. 2020, 98, e178–e180. [Google Scholar] [CrossRef] [PubMed]
- Ugazio, E.; Tullio, V.; Binello, A.; Tagliapietra, S.; Dosio, F. Ozonated Oils as Antimicrobial Systems in Topical Applications. Their Characterization, Current Applications, and Advances in Improved Delivery Techniques. Molecules 2020, 25, 334. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
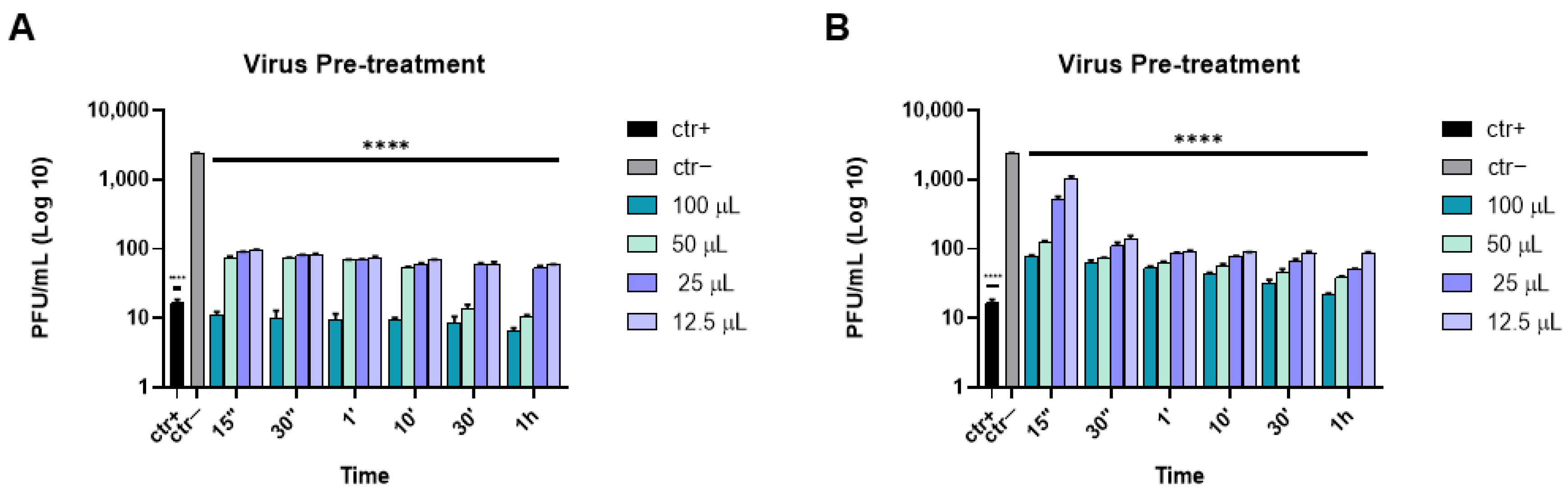{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence |
|---|---|
| UL54 | Forward: TGGCGGACATTAAGGACATTG Reverse: TGGCCGTCAACTCGCAG |
| UL52 | Forward: GACCGACGGGTGCGTTATT Reverse: GAAGGAGTCGCCATTTAGCC |
| UL27 | Forward: GCCTTCTTCGCCTTTCGC Reverse: CGCTCGTGCCCTTCTTCTT |
| GAPDH | Forward: CCTTTCATTGAGCTCCAT Reverse: CGTACATGGGAGCGTC |
| Thermocycler conditions | |
| 95 °C for 10 min | |
| 95 °C for 15 s 60 °C for 1 min 72 °C for 20 s | 40 cycles |
| EYE DROP | INCUBATION TIME | CC50 (μL) | Antiviral Assay | IC50 (μL) | TI |
|---|---|---|---|---|---|
| Iodim | 15 s | >100 | Co-treatment | 30 | 3.3 |
| 30 s | 25 | 4 | |||
| 1 min | <12.5 | <8 | |||
| 10 min | <12.5 | <8 | |||
| 30 min | <12.5 | <8 | |||
| 1 h | <12.5 | <8 | |||
| Iodim | 15 s | Virus pre-treatment | 25 | 4 | |
| 30 s | 12.5 | 8 | |||
| 1 min | <12.5 | <8 | |||
| 10 min | <12.5 | <8 | |||
| 30 min | <12.5 | <8 | |||
| 1 h | <12.5 | <8 | |||
| Ozodrop | 15 s | >100 | Co-treatment | 30 | 3.3 |
| 30 s | 25 | 4 | |||
| 1 min | <12.5 | <8 | |||
| 10 min | <12.5 | <8 | |||
| 30 min | <12.5 | <8 | |||
| 1 h | <12.5 | <8 | |||
| Ozodrop | 15 s | Virus pre-treatment | 75 | 1.3 | |
| 30 s | 25 | 4 | |||
| 1 min | <12.5 | <8 | |||
| 10 min | <12.5 | <8 | |||
| 30 min | <12.5 | <8 | |||
| 1 h | <12.5 | <8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zannella, C.; Chianese, A.; De Bernardo, M.; Folliero, V.; Petrillo, F.; De Filippis, A.; Boccia, G.; Franci, G.; Rosa, N.; Galdiero, M. Ophthalmic Solutions with a Broad Antiviral Action: Evaluation of Their Potential against Ocular Herpetic Infections. Microorganisms 2022, 10, 1728. https://doi.org/10.3390/microorganisms10091728
Zannella C, Chianese A, De Bernardo M, Folliero V, Petrillo F, De Filippis A, Boccia G, Franci G, Rosa N, Galdiero M. Ophthalmic Solutions with a Broad Antiviral Action: Evaluation of Their Potential against Ocular Herpetic Infections. Microorganisms. 2022; 10(9):1728. https://doi.org/10.3390/microorganisms10091728
Chicago/Turabian StyleZannella, Carla, Annalisa Chianese, Maddalena De Bernardo, Veronica Folliero, Francesco Petrillo, Anna De Filippis, Giovanni Boccia, Gianluigi Franci, Nicola Rosa, and Massimiliano Galdiero. 2022. "Ophthalmic Solutions with a Broad Antiviral Action: Evaluation of Their Potential against Ocular Herpetic Infections" Microorganisms 10, no. 9: 1728. https://doi.org/10.3390/microorganisms10091728
APA StyleZannella, C., Chianese, A., De Bernardo, M., Folliero, V., Petrillo, F., De Filippis, A., Boccia, G., Franci, G., Rosa, N., & Galdiero, M. (2022). Ophthalmic Solutions with a Broad Antiviral Action: Evaluation of Their Potential against Ocular Herpetic Infections. Microorganisms, 10(9), 1728. https://doi.org/10.3390/microorganisms10091728